Fly-Ash Pollution Modulates Growth, Biochemical Attributes, Antioxidant Activity and Gene Expression in Pithecellobium Dulce (Roxb) Benth

 ,
,  ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area and its Climate
2.2. Study Site Selection
2.3. Sampling of Plant Material
2.4. Measurement of pH and Suspended Particulate Matter (SPM)
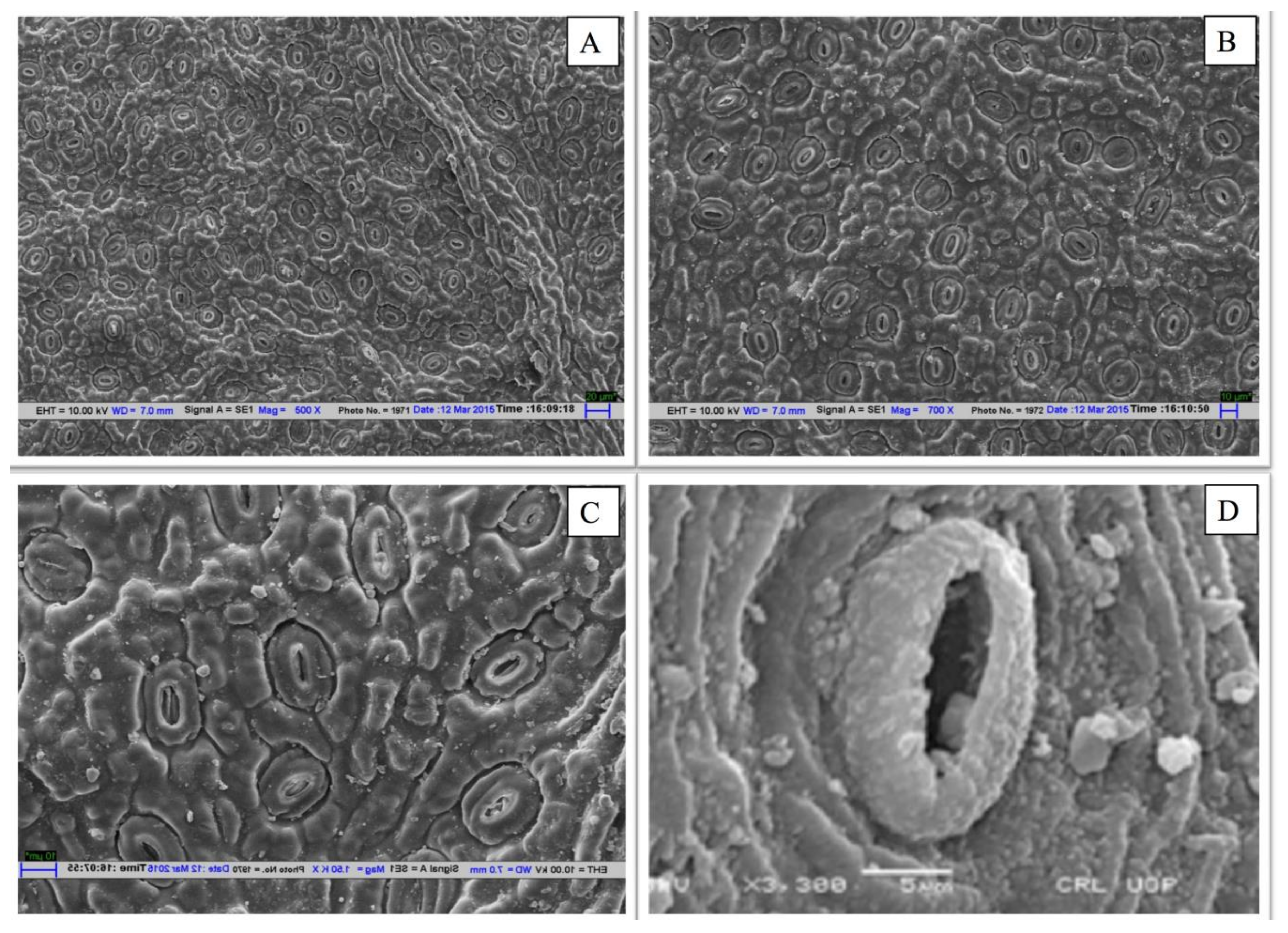
2.5. Epidermal Studies
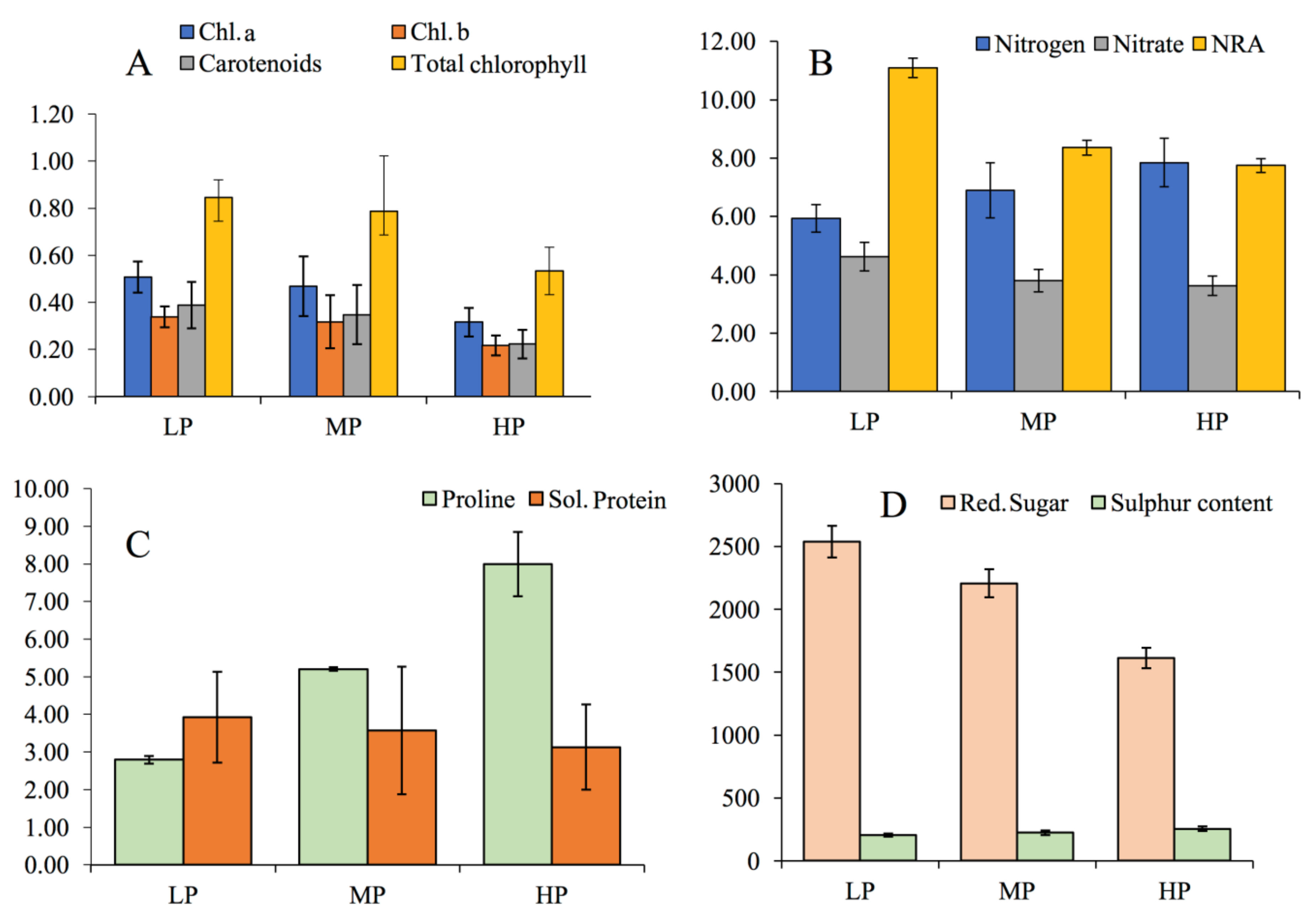
2.6. Photosynthesis and Photosynthetic Pigments
2.7. Nitrogen Assimilation Related Parameters
2.8. Sugar, Sulphur, Proline, and Protein Contents
2.9. Air Pollution Tolerance Index (APTI)
2.10. Estimation of Malondialdehyde (MDA) and Antioxidant Enzyme Activities
2.11. Ascorbate Peroxidase (APX EC1.11.1.11)
2.12. Catalase (CAT, EC1.11.1.6)
2.13. Superoxide Dismutase (SOD, EC1.15.1.1)
2.14. Peroxidase (POD, EC1.11.1.7)
2.15. Expression Analysis of Antioxidant and Stress Responsive Genes
2.15.1. Primer Design
2.15.2. RNA Extraction and Semi-Quantitative PCR Analysis
2.15.3. Quantitative Real-Time PCR
2.16. Statistical Analysis
3. Results and Discussion
4. Conclusions
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Pandey, A.K.; Pandey, M.; Mishra, A.; Tiwary, S.M.; Tripathi, B.D. Air pollution tolerance index and anticipated performance index of some plant species for development of urban forest. Urban For. Urban Green. 2015, 14, 866–871. [Google Scholar] [CrossRef]
- Haynes, R. Reclamation and revegetation of fly ash disposal sites—Challenges and research needs. J. Environ. Manag. 2009, 90, 43–53. [Google Scholar] [CrossRef] [PubMed]
- George, J.; Masto, R.E.; Ram, L.C.; Das, T.B.; Rout, T.K.; Mohan, M. Human exposure risks for metals in soil near a coal-fired power-generating plant. Arch. Environ. Contam. Toxicol. 2015, 68, 451–461. [Google Scholar] [CrossRef] [PubMed]
- Pandey, V.C.; Singh, J.S.; Singh, R.P.; Singh, N.; Yunus, M. Arsenic hazards in coal fly ash and its fate in Indian scenario. Resour. Conserv. Recycl. 2011, 55, 819–835. [Google Scholar] [CrossRef]
- Gupta, M.; Iqbal, M. Ontogenetic histological changes in the wood of mango (Mangifera indica L. cv Deshi) exposed to coal-smoke pollution. Environ. Exp. Bot. 2005, 54, 248–255. [Google Scholar] [CrossRef]
- Pandey, V.C.; Prakash, P.; Bajpai, O.; Kumar, A.; Singh, N. Phytodiversity on fly ash deposits: Evaluation of naturally colonized species for sustainable phytorestoration. Environ. Sci. Pollut. Res. 2015, 22, 2776–2787. [Google Scholar] [CrossRef] [PubMed]
- Pandey, A.K.; Pandey, M.; Tripathi, B.D. Air Pollution Tolerance Index of climber plant species to develop Vertical Greenery Systems in a polluted tropical city. Landsc. Urban Plan. 2015, 144, 119–127. [Google Scholar] [CrossRef]
- Ribeiro, J.; Silva, T.; Mendonça Filho, J.; Flores, D. Fly ash from coal combustion—An environmental source of organic compounds. Appl. Geochem. 2014, 44, 103–110. [Google Scholar] [CrossRef]
- Pandey, V.C. Assisted phytoremediation of fly ash dumps through naturally colonized plants. Ecol. Eng. 2015, 82, 1–5. [Google Scholar] [CrossRef]
- Pandey, V.C.; Singh, B. Rehabilitation of Coal Fly Ash Basins: Current Need to Use Ecological Engineering; Elsevier Academic Press: New York, NY, USA, 2012. [Google Scholar]
- Jambhulkar, H.P.; Juwarkar, A.A. Assessment of bioaccumulation of heavy metals by different plant species grown on fly ash dump. Ecotoxicol. Environ. Saf. 2009, 72, 1122–1128. [Google Scholar] [CrossRef]
- Juwarkar, A.A.; Jambhulkar, H.P. Restoration of fly ash dump through biological interventions. Environ. Monit. Assess. 2008, 139, 355–365. [Google Scholar] [CrossRef] [PubMed]
- Ram, L.C.; Jha, S.K.; Tripathi, R.C.; Masto, R.E.; Selvi, V.A. Remediation of fly ash landfills through plantation. Remediation 2008, 18, 71–90. [Google Scholar] [CrossRef]
- Qadir, S.U.; Raja, V.; Siddiqui, W.A. Morphological and biochemical changes in Azadirachta indica from coal combustion fly ash dumping site from a thermal power plant in Delhi, India. Ecotoxicol. Environ. Saf. 2016, 129, 320–328. [Google Scholar] [CrossRef] [PubMed]
- Iqbal, M.; Jacob, M.; Mehta, S.; Mahmooduzzafar; Srivastava, P. Plant responses to particulate matters suspended in the air. In Environmental Hazards: Plants and People; CBS Publishers: New Delhi, India, 2000; pp. 80–112. [Google Scholar]
- Goswami, S.; Das, S. Copper phytoremediation potential of Calandula officinalis L. and the role of antioxidant enzymes in metal tolerance. Ecotoxicol. Environ. Saf. 2016, 126, 211–218. [Google Scholar] [CrossRef] [PubMed]
- Pandey, V.C.; Singh, J.S.; Kumar, A.; Tewari, D. Accumulation of heavy metals by chickpea grown in fly ash treated soil: Effect on antioxidants. CLEAN Soil Air Water 2010, 38, 1116–1123. [Google Scholar] [CrossRef]
- Tauqeer, H.M.; Ali, S.; Rizwan, M.; Ali, Q.; Saeed, R.; Iftikhar, U.; Ahmad, R.; Farid, M.; Abbasi, G.H. Phytoremediation of heavy metals by Alternanthera bettzickiana: Growth and physiological response. Ecotoxicol. Environ. Saf. 2016, 126, 138–146. [Google Scholar] [CrossRef]
- Sharma, P.; Jha, A.B.; Dubey, R.S.; Pessarakli, M. Reactive oxygen species, oxidative damage, and antioxidative defense mechanism in plants under stressful conditions. J. Bot. 2012, 2012, 1–26. [Google Scholar] [CrossRef]
- Iqbal, M.; Abdin, M.Z.; Mahmooduzzafar; Yunus, M.; Agrawal, M. Resistance mechanisms in plants against air pollution. In Plant Response to Air Pollution; Yunus, M., Iqbal, M., Eds.; John Willey & Sons: Chichester, UK, 1996; pp. 195–240. [Google Scholar]
- Ali, I.; Liu, B.; Farooq, M.A.; Islam, F.; Azizullah, A.; Yu, C.; Su, W.; Gan, Y. Toxicological effects of bisphenol A on growth and antioxidant defense system in Oryza sativa as revealed by ultrastructure analysis. Ecotoxicol. Environ. Saf. 2016, 124, 277–284. [Google Scholar] [CrossRef]
- Pandey, V.C.; Singh, K.; Singh, R.P.; Singh, B. Naturally growing Saccharum munja L. on the fly ash lagoons: A potential ecological engineer for the revegetation and stabilization. Ecol. Eng. 2012, 40, 95–99. [Google Scholar] [CrossRef]
- Kumari, A.; Pandey, V.C.; Rai, U.N. Feasibility of fern Thelypteris dentata for revegetation of coal fly ash landfills. J. Geochem. Explor. 2013, 128, 147–152. [Google Scholar] [CrossRef]
- Pandey, V.C.; Bajpai, O.; Singh, N. Plant regeneration potential in fly ash ecosystem. Urban For. Urban Green. 2016, 15, 40–44. [Google Scholar] [CrossRef]
- Chaturvedi, R.; Prasad, S.; Rana, S.; Obaidullah, S.; Pandey, V.; Singh, H. Effect of dust load on the leaf attributes of the tree species growing along the roadside. Environ. Monit. Assess. 2013, 185, 383–391. [Google Scholar] [CrossRef] [PubMed]
- Lukaszkiwicz, J. Determining the age of alley trees with the dbh-height-based multi-factoral model. In Annals of Warsaw University of Life Sciences-SGGW; Horticulture and Landscape Architecture: West Lafayette, IN, USA, 2010; pp. 53–60. [Google Scholar]
- Ghouse, A.; Yunus, M. Preparation of epidermal peels from leaves of gymnosperms by treatment with hot, 60 per cent HNO3. Stain Technol. 1972, 47, 322. [Google Scholar] [CrossRef] [PubMed]
- Salisbury, E.J.I. On the causes and ecological significance of stomatal frequency, with special reference to the woodland flora. Philos. Trans. R. Soc. Lond. Ser. B Contain. Pap. Biol. Character 1928, 216, 1–65. [Google Scholar] [CrossRef]
- Karcz, J.; Bernas, T.; Nowak, A.; Talik, E.; Woznica, A. Application of lyophilization to prepare the nitrifying bacterial biofilm for imaging with scanning electron microscopy. Scanning 2012, 34, 26–36. [Google Scholar] [CrossRef] [PubMed]
- Hiscox, J.D.; Israelstam, G.F. A method for the extraction of chlorophyll from leaf tissue without maceration. Can. J. Bot. 1979, 57, 1332–1334. [Google Scholar] [CrossRef]
- Duxbury, A.C.; Yentsch, C.S. Plankton Pigment Nomographs. J. Mar. Biol. 1956, 16, 145–150. [Google Scholar]
- Maclachlan, S.; Zalik, S. Plastid structure, chlorophyll concentration and free amino acid composition of a chlorophyll mutant of barley. Can. J. Bot. 1963, 41, 1053–1062. [Google Scholar] [CrossRef]
- Klepper, L.; Flesher, D.; Hageman, R.H. Generation of Reduced Nicotinamide Adenine Dinucleotide for Nitrate Reduction in Green Leaves. Plant Physiol. 1971, 48, 580–590. [Google Scholar] [CrossRef]
- Grover, H.; Nair, T.; Abrol, Y. Nitrogen metabolism of the upper three leaf blades of wheat at different soil nitrogen levels: I. Nitrate reductase activity and content of various nitrogenous constituents. Physiol. Plant. 1978, 42, 287–292. [Google Scholar] [CrossRef]
- Evans, H.J.; Nason, A. Pyridine Nucleotide-Nitrate Reductase from Extracts of Higher Plants. Plant Physiol. 1953, 28, 233–254. [Google Scholar] [CrossRef] [PubMed]
- Lindner, R. Rapid analytical methods for some of the more common inorganic constituents of plant tissues. Plant Physiol. 1944, 19, 76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dey, P.M. Carbohydrates. In Methods in Plant Biochemistry; Accademic Press: London, UK, 1990; Volume 2, pp. 189–218. [Google Scholar] [CrossRef]
- Chesnin, L.; Yien, C. Turbidimetric determination of available sulfates 1. Soil Sci. Soc. Am. J. 1951, 15, 149–151. [Google Scholar] [CrossRef]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid determination of free proline for water-stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Bradford, M. A Rapid and Sensitive Method for the Quantitation of Microgram Quantities of Protein Utilizing the Principle of Protein-Dye Binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Keller, T.; Schwager, H. Air pollution and ascorbic acid. Eur. J. For. Pathol. 1977, 7, 338–350. [Google Scholar] [CrossRef]
- Qadir, S.U.; Siddiqui, W.A. Effect of fly ash on some biochemical parameters of selected plants growing at dumping site of badarpur thermal power plant in delhi. Int. J. Res. Appl. Nat. Soc. Sci. 2014, 2, 7–14. [Google Scholar]
- Singh, S.K.; Rao, D.N.; Agrawal, M.; Pandey, J.; Naryan, D. Air pollution tolerance index of plants. J. Environ. Manag. 1991, 32, 45–55. [Google Scholar] [CrossRef]
- Heath, R.L.; Packer, L. Photoperoxidation in isolated chloroplasts. Arch. Biochem. Biophys. 1968, 125, 189–198. [Google Scholar] [CrossRef]
- Nakano, Y.; Asada, K. Hydrogen peroxide is scavenged by ascorbate-specific peroxidase in spinach chloroplasts. Plant Cell Physiol. 1981, 22, 867–880. [Google Scholar]
- Aebi, H. Catalase In Vitro. Methods Enzymol. 1984, 105, 121–126. [Google Scholar] [PubMed]
- Zhang, W.F.; Zhang, F.; Raziuddin, R.; Gong, H.J.; Yang, Z.M.; Lu, L.; Ye, Q.F.; Zhou, W.J. Effects of 5-Aminolevulinic Acid on Oilseed Rape Seedling Growth under Herbicide Toxicity Stress. J. Plant Growth Regul. 2008, 27, 159–169. [Google Scholar] [CrossRef]
- Zhou, W.; Leul, M. Uniconazole-induced alleviation of freezing injury in relation to changes in hormonal balance, enzyme activities and lipid peroxidation in winter rape. Plant Growth Regul. 1998, 26, 41–47. [Google Scholar] [CrossRef]
- Liu, S.; Chen, C.; Chen, G.; Cao, B.; Chen, Q.; Lei, J. RNA-sequencing tag profiling of the placenta and pericarp of pungent pepper provides robust candidates contributing to capsaicinoid biosynthesis. Plant Cell Tissue Organ Culture 2012, 110, 111–121. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Iqbal, M.; Bano, R.; Wali, B. Plant Growth Responses to Air Pollution. Plant Biodiversity, Microbial Interaction and Environmental Biology; Avishkar Publishers: Jaipur, India, 2005; pp. 166–188. [Google Scholar]
- Wali, B.; Iqbal, M.; Mahmooduzzafar. Anatomical and functional responses of Calendula officinalis L. to SO2 stress as observed at different stages of plant development. Flora Morphol. Distrib. Funct. Ecol. Plants 2007, 202, 268–280. [Google Scholar] [CrossRef]
- Iqbal, M.; Mahmooduzzafar; Nighat, F.; Khan, P.R. Photosynthetic, metabolic and growth responses of Triumfetta rhomboideato coal-smoke pollution at different stages of plant ontogeny. J. Plant Interact. 2010, 5, 11–19. [Google Scholar] [CrossRef] [Green Version]
- Ots, K.; Indriksons, A.; Varnagiryte-Kabasinskiene, I.; Mandre, M.; Kuznetsova, T.; Klõšeiko, J.; Tilk, M.; Kõresaar, K.; Lukjanova, A.; Kikamägi, K. Changes in the canopies of Pinus sylvestris and Picea abies under alkaline dust impact in the industrial region of Northeast Estonia. For. Ecol. Manag. 2011, 262, 82–87. [Google Scholar] [CrossRef]
- Saha, D.C.; Padhy, P.K. Effects of stone crushing industry on Shorea robusta and Madhuca indica foliage in Lalpahari forest. Atmos. Pollut. Res. 2011, 2, 463–476. [Google Scholar] [CrossRef] [Green Version]
- Tiwari, S.; Agrawal, M.; Marshall, F. Evaluation of ambient air pollution impact on carrot plants at a sub urban site using open top chambers. Environ. Monit. Assess. 2006, 119, 15–30. [Google Scholar] [CrossRef]
- Wagh, N.; Shukla, P.V.; Tambe, S.B.; Ingle, S. Biological monitoring of roadside plants exposed to vehicular pollution in Jalgaon city. J. Environ. Biol. 2006, 37, 419–421. [Google Scholar]
- Verma, A.; Singh, S.N. Biochemical and Ultrastructural Changes in Plant Foliage Exposed to Auto-Pollution. Environ. Monit. Assess. 2006, 120, 585–602. [Google Scholar] [CrossRef] [PubMed]
- Joshi, P.; Swami, A. Air pollution induced changes in the photosynthetic pigments of selected plant species. J. Environ. Biol. 2009, 30, 295–298. [Google Scholar]
- Joshi, N.; Chauhan, A.; Joshi, P.C. Impact of industrial air pollutants on some biochemical parameters and yield in wheat and mustard plants. Environmentalist 2009, 29, 398–404. [Google Scholar] [CrossRef]
- Bashir, H.; Ibrahim, M.M.; Bagheri, R.; Ahmad, J.; Arif, I.A.; Baig, M.A.; Qureshi, M.I. Influence of sulfur and cadmium on antioxidants, phytochelatins and growth in Indian mustard. AoB Plants 2015, 7. [Google Scholar] [CrossRef] [Green Version]
- Iqbal, M.; Jura-Morawiec, J.; WŁoch, W.; Mahmooduzzafar. Foliar characteristics, cambial activity and wood formation in Azadirachta indica A. Juss. as affected by coal–smoke pollution. Flora-Morphol. Distrib. Funct. Ecol. Plants 2010, 205, 61–71. [Google Scholar] [CrossRef]
- Grantz, D.; Garner, J.; Johnson, D. Ecological effects of particulate matter. Environ. Int. 2003, 29, 213–239. [Google Scholar] [CrossRef]
- Sharma, A.P.; Tripathi, B.D. Biochemical responses in tree foliage exposed to coal-fired power plant emission in seasonally dry tropical environment. Environ. Monit. Assess. 2008, 158, 197–212. [Google Scholar] [CrossRef]
- Kumar, A.; Vajpayee, P.; Ali, M.B.; Tripathi, R.D.; Singh, N.; Rai, U.N.; Singh, S.N. Biochemical Responses of Cassia siamea Lamk. Grown on Coal Combustion Residue (Fly-ash). Bull. Environ. Contam. Toxicol. 2002, 68, 675–683. [Google Scholar] [CrossRef]
- Hayat, S.; Hayat, Q.; Alyemeni, M.N.; Wani, A.S.; Pichtel, J.; Ahmad, A. Role of proline under changing environments. Plant Signal. Behav. 2012, 7, 1456–1466. [Google Scholar] [CrossRef] [Green Version]
- Tripathi, A.; Gautam, M. Biochemical parameters of plants as indicators of air pollution. J. Environ. Biol. 2007, 28, 127. [Google Scholar] [PubMed]
- Krishnaveni, M. Air pollution tolerance index and antioxidant activity of Parthenium hysterophorus. J. Pharm. Res. 2013, 7, 296–298. [Google Scholar] [CrossRef]
- Kaur, R.; Nayyar, H. Ascorbic acid: A potent defender against environmental stresses. In Oxidative Damage to Plants; Elsevier Academic Press: New York, NY, USA, 2014; pp. 235–287. [Google Scholar]
- Aref, I.M.; Khan, P.R.; Khan, S.; El-Atta, H.; Ahmed, A.I.; Iqbal, M. Modulation of antioxidant enzymes in Juniperus procera needles in relation to habitat environment and dieback incidence. Trees 2016, 30, 1669–1681. [Google Scholar] [CrossRef]
- Zouari, M.; Ahmed, C.B.; Zorrig, W.; Elloumi, N.; Rabhi, M.; Delmail, D.; Rouina, B.B.; Labrousse, P.; Abdallah, F.B. Exogenous proline mediates alleviation of cadmium stress by promoting photosynthetic activity, water status and antioxidative enzymes activities of young date palm (Phoenix dactylifera L.). Ecotoxicol. Environ. Saf. 2016, 128, 100–108. [Google Scholar] [CrossRef]
- Raja, V.; Majeed, U.; Kang, H.; Andrabi, K.I.; John, R. Abiotic stress: Interplay between ROS, hormones and MAPKs. Environ. Exp. Bot. 2017, 137, 142–157. [Google Scholar] [CrossRef]
- Foyer, C.H.; Ruban, A.V.; Noctor, G. Viewing oxidative stress through the lens of oxidative signalling rather than damage. Biochem. J. 2017, 474, 877–883. [Google Scholar] [CrossRef] [Green Version]
- Petrov, V.; Hille, J.; Mueller-Roeber, B.; Gechev, T.S. ROS-mediated abiotic stress-induced programmed cell death in plants. Front. Plant Sci. 2015, 6, 69. [Google Scholar] [CrossRef] [Green Version]
- Hu, Z.; Fan, J.; Xie, Y.; Amombo, E.; Liu, A.; Gitau, M.M.; Khaldun, A.B.M.; Chen, L.; Fu, J. Comparative photosynthetic and metabolic analyses reveal mechanism of improved cold stress tolerance in bermudagrass by exogenous melatonin. Plant Physiol. Biochem. 2016, 100, 94–104. [Google Scholar] [CrossRef] [Green Version]
- Jan, N.; Majeed, U.; Andrabi, K.I.; John, R. Cold stress modulates osmolytes and antioxidant system in Calendula officinalis. Acta Physiol. Plant. 2018, 40. [Google Scholar] [CrossRef]
- Harb, A.; Awad, D.; Samarah, N. Gene expression and activity of antioxidant enzymes in barley (Hordeum vulgare L.) under controlled severe drought. J. Plant Interact. 2015, 10, 109–116. [Google Scholar] [CrossRef]
- Teixeira, F.K.; Menezes-Benavente, L.; Galvão, V.C.; Margis, R.; Margis-Pinheiro, M. Rice ascorbate peroxidase gene family encodes functionally diverse isoforms localized in different subcellular compartments. Planta 2006, 224, 300. [Google Scholar] [CrossRef] [PubMed]
- Hundertmark, M.; Hincha, D.K. LEA (Late Embryogenesis Abundant) proteins and their encoding genes in Arabidopsis thaliana. BMC Genom. 2008, 9, 118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sakuma, Y.; Liu, Q.; Dubouzet, J.G.; Abe, H.; Shinozaki, K.; Yamaguchi-Shinozaki, K. DNA-binding specificity of the ERF/AP2 domain of Arabidopsis DREBs, transcription factors involved in dehydration-and cold-inducible gene expression. Biochem. Biophys. Res. Commun. 2002, 290, 998–1009. [Google Scholar] [CrossRef] [PubMed]
- Nakashima, K.; Shinwari, Z.K.; Sakuma, Y.; Seki, M.; Miura, S.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Organization and expression of two Arabidopsis DREB2 genes encoding DRE-binding proteins involved in dehydration-and high-salinity-responsive gene expression. Plant Mol. Biol. 2000, 42, 657–665. [Google Scholar] [CrossRef] [PubMed]
- Gao, S.-Q.; Chen, M.; Xia, L.-Q.; Xiu, H.-J.; Xu, Z.-S.; Li, L.-C.; Zhao, C.-P.; Cheng, X.-G.; Ma, Y.-Z. A cotton (Gossypium hirsutum) DRE-binding transcription factor gene, GhDREB, confers enhanced tolerance to drought, high salt, and freezing stresses in transgenic wheat. Plant Cell Rep. 2009, 28, 301–311. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sites | Intercellular Carbon Dioxide (mmol, CO2 m−2 S−1) | Stomatal Conductance (mmol, CO2 m−2 S−1) | PN(mmol m−2 S−1) | Stomatal Index (%) | SLA (cm2) | Leaf Length (cm) | Leaf Width (cm) |
|---|---|---|---|---|---|---|---|
| LPS | 313.22 ± 24.70 | 1.15 ± 0.78 | 5.36 ± 1.62 | 14.12 ± 0.48 | 6.31 ± 0.85 | 9.16 ± 0.61 | 3.56 ± 0.26 |
| MPS | 331.38 ± 25.93 | 0.87 ± 0.52 | 4.47 ± 1.60 | 14.15 ± 0.43 | 4.23 ± 0.21 | 6.93 ± 1.20 | 2.59 ± 0.25 |
| HPS | 353.80 ± 26.11 | 0.61 ± 0.24 | 3.36 ± 1.76 | 15.63 ± 0.63 | 3.06 ± 0.23 | 4.94 ± 0.90 | 1.21 ± 0.27 |
| Parameters | Sum of Squares | Df | Mean Square | F | Sig. |
|---|---|---|---|---|---|
| Ascorbic acid | 23.32 | 74 | 11.66 | 54.76 | *** |
| pH | 32.35 | 74 | 16.18 | 441.83 | *** |
| FW | 3268.12 | 74 | 1634.06 | 8006.07 | *** |
| DW | 2768.57 | 74 | 1384.28 | 2976.34 | *** |
| TW | 4135.29 | 74 | 2067.65 | 2685.09 | *** |
| RWC | 71.17 | 74 | 35.58 | 0.27 | NS |
| APTI | 63.11 | 74 | 31.55 | 21.97 | *** |
| SOD | 694.25 | 74 | 347.13 | 961.79 | *** |
| APX | 93.39 | 74 | 46.70 | 6467.29 | *** |
| CAT | 377.16 | 74 | 188.58 | 3644.43 | *** |
| POD | 238.34 | 74 | 119.17 | 488.64 | *** |
| MDA | 25.20 | 74 | 12.60 | 2282.15 | *** |
| Sites | Chl. a | Chl. b | CTDS | T. Chl. | NPR | NRA | Nitrate | Nitrogen | Proline | SPT | RS | SCc | SI | SC | ICD | SPM | SLA | LL | LW | AA | pH | FW | DW | TW | RWC | APTI | SOD | APX | CAT | POD | MDA | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Sites | 1 | |||||||||||||||||||||||||||||||
| Chl. a | −0.649 a | 1 | ||||||||||||||||||||||||||||||
| Chl. b | −0.551 a | 0.854 a | 1 | |||||||||||||||||||||||||||||
| CTDS | −0.565 a | 0.828 a | 0.893 a | 1 | ||||||||||||||||||||||||||||
| T. Chl. | −0.630 a | 0.973 a | 0.951 a | 0.889 a | ||||||||||||||||||||||||||||
| NPR | −0.448 a | 0.644 a | 0.634 a | 0.512 a | 0.664 a | 1 | ||||||||||||||||||||||||||
| NRA | −0.314 a | 0.624 a | 0.295 b | 0.357 a | 0.502 a | 0.608 a | 1 | |||||||||||||||||||||||||
| Nitrate | −0.686 a | 0.508 a | 0.247 b | 0.176 | 0.411 a | 0.585 a | 0.699 a | 1 | ||||||||||||||||||||||||
| Nitrogen | 0.718 a | −0.619 a | −0.397 a | −0.482 a | −0.544 a | −0.357 a | −0.619 a | −0.672 a | 1 | |||||||||||||||||||||||
| Proline | 0.974 a | −0.623 a | −0.516 a | −0.535 a | −0.599 a | −0.381 a | −0.291 b | −0.634 a | 0.686 a | 1 | ||||||||||||||||||||||
| SPT | −0.236 b | 0.672 a | 0.535 a | 0.503 a | 0.636 a | 0.740 a | 0.768 a | 0.496 a | −0.344 a | −0.192 | 1 | |||||||||||||||||||||
| RS | −0.313 a | 0.318 a | 0.520 a | 0.379 a | 0.420 a | 0.660 a | 0.058 | 0.161 | 0.165 | −0.268 b | 0.348 a | 1 | ||||||||||||||||||||
| SCc. | 0.792 a | −0.496 a | −0.578 a | −0.552 a | −0.551 a | −0.334 a | 0.093 | −0.248 b | 0.331 a | 0.799 a | −0.077 | −0.439 a | 1 | |||||||||||||||||||
| SI | 0.711 a | −0.720 a | −0.477 a | −0.543 a | −0.639 a | −0.410 a | −0.510 a | −0.527 a | 0.725 a | 0.725 a | −0.323 a | −0.067 | 0.493 a | 1 | ||||||||||||||||||
| SC | −0.372 a | 0.737 a | 0.681 a | 0.865 a | 0.740 a | 0.500 a | 0.557 a | 0.188 | −0.406 a | −0.351 a | 0.595 a | 0.278 b | −0.292 b | −0.427 a | 1 | |||||||||||||||||
| ICD | 0.551 a | −0.236 b | −0.290 b | 0.004 | −0.269 b | −0.472 a | −0.046 | −0.609 a | 0.273 b | 0.494 a | −0.114 | −0.380 a | 0.404 a | 0.250 b | 0.227 b | 1 | ||||||||||||||||
| SPM | 0.408 a | −0.001 | 0.107 | 0.037 | 0.047 | −0.144 | 0.022 | −0.227 | 0.414 a | 0.397 a | 0.190 | 0.111 | 0.418 a | 0.391 a | 0.146 | 0.182 | 1 | |||||||||||||||
| SLA | −0.922 a | 0.566 a | 0.414 a | 0.432 a | 0.520 a | 0.432 a | 0.384 a | 0.769 a | −0.701 a | −0.889 a | 0.215 | 0.171 | −0.618 a | −0.621 a | 0.309 a | −0.519 a | −0.406 a | 1 | ||||||||||||||
| LL | −0.884 a | 0.555 a | 0.452 a | 0.463 a | 0.530 a | 0.342 a | 0.230 b | 0.576 a | −0.617 a | −0.845 a | 0.115 | 0.339 a | −0.688 a | −0.603 a | 0.316 a | −0.445 a | −0.368 a | 0.791 a | 1 | |||||||||||||
| LW | −0.961 a | 0.677 a | 0.593 a | 0.585 a | 0.666 a | 0.467 a | 0.300 a | 0.645 a | −0.654 a | −0.938 a | 0.294 b | 0.358 a | −0.792 a | −0.693 a | 0.378 a | −0.566 a | −0.332 a | 0.854 a | 0.822 a | 1 | ||||||||||||
| AA | 0.768 a | −0.490 a | −0.334 a | −0.363 a | −0.439 a | −0.446 a | −0.452 a | −0.690 a | 0.619 a | 0.735 a | −0.316 a | −0.217 | 0.453 a | 0.621 a | −0.275 b | 0.494 a | 0.244 b | −0.782 a | −0.568 a | −0.739 a | 1 | |||||||||||
| pH | 0.834 a | −0.679 a | −0.586 a | −0.546 a | −0.664 a | −0.481 a | −0.248 b | −0.506 a | 0.667 a | 0.817 a | −0.213 | −0.231 b | 0.684 a | 0.802 a | −0.304 a | 0.550 a | 0.506 a | −0.746 a | −0.714 a | −0.835 a | 0.692 a | 1 | ||||||||||
| FW | −0.871 a | 0.687 a | 0.600 a | 0.581 a | 0.674 a | 0.411 a | 0.215 | 0.470 a | −0.618 a | −0.868 a | 0.215 | 0.310 a | −0.752 a | −0.808 a | 0.329 a | −0.506 a | −0.391 a | 0.727 a | 0.764 a | 0.885 a | −0.711 a | −0.961 a | 1 | |||||||||
| DW | −0.861 a | 0.673 a | 0.576 a | 0.566 a | 0.656 a | 0.400 a | 0.220 | 0.465 a | −0.619 a | −0.866 a | 0.217 | 0.283 b | −0.748 a | −0.821 a | 0.324 a | −0.491 a | −0.401 a | 0.722 a | 0.743 a | 0.876 a | −0.714 a | −0.954 a | 0.993 a | 1 | ||||||||
| TW | −0.874 a | 0.700 a | 0.600 a | 0.586 a | 0.682 a | 0.432 a | 0.251 b | 0.492 a | −0.626 a | −0.871 a | 0.236 b | 0.310 a | −0.740 a | −0.821 a | 0.347 a | −0.495 a | −0.394 a | 0.744 a | 0.761 a | 0.885 a | −0.728 a | −0.961 a | 0.994 a | 0.990 a | 1 | |||||||
| RWC | 0.084 | −0.135 | −0.044 | −0.062 | −0.099 | −0.095 | −0.177 | −0.071 | 0.158 | 0.151 | −0.064 | 0.008 | 0.099 | 0.309 a | −0.065 | −0.074 | 0.122 | −0.051 | −0.104 | −0.073 | 0.053 | 0.071 | −0.070 | −0.118 | −0.103 | 1 | ||||||
| APTI | 0.574 a | −0.423 a | −0.270 b | −0.284 b | −0.371 a | −0.337 a | −0.324 a | −0.432 a | 0.529 a | 0.611 a | −0.163 | −0.111 | 0.433 a | 0.688 a | −0.177 | 0.296 b | 0.382 a | −0.528 a | −0.488 a | −0.553 a | 0.593 a | 0.604 a | −0.594 a | −0.631 a | −0.622 a | 0.796 a | 1 | |||||
| SOD | 0.952 a | −0.620 a | −0.517 a | −0.533 a | −0.598 a | −0.329 a | −0.209 | −0.556 a | 0.660 a | 0.956 a | −0.166 | −0.250 b | 0.800 a | 0.745 a | −0.306 a | 0.494 a | 0.413 a | −0.840 a | −0.827 a | −0.944 a | 0.749 a | 0.887 a | −0.943 a | −0.938 a | −0.941 a | 0.053 | 0.569 a | 1 | ||||
| APX | 0.997 a | −0.647 a | −0.549 a | −0.561 a | −0.628 a | −0.417 a | −0.284 b | −0.663 a | 0.694 a | 0.978 a | −0.208 | −0.306 a | 0.802 a | 0.712 a | −0.365 a | 0.544 a | 0.390 a | −0.917 a | −0.883 a | −0.961 a | 0.762 a | 0.842 a | −0.884 a | −0.875 a | −0.887 a | 0.079 | 0.571 a | 0.962 a | 1 | |||
| CAT | 0.913 a | −0.669 a | −0.568 a | −0.566 a | −0.650 a | −0.425 a | −0.251 b | −0.534 a | 0.656 a | 0.908 a | −0.202 | −0.320 a | 0.746 a | 0.806 a | −0.333 a | 0.519 a | 0.420 a | −0.788 a | −0.812 a | −0.908 a | 0.750 a | 0.955 a | −0.989 a | −0.981 a | −0.988 a | 0.089 | 0.620 a | 0.961 a | 0.923 a | 1 | ||
| POD | 0.965 a | −0.614 a | −0.500 a | −0.490 a | −0.587 a | −0.409 a | −0.262 b | −0.687 a | 0.627 a | 0.949 a | −0.200 | −0.291 b | 0.801 a | 0.699 a | −0.285 b | 0.597 a | 0.397 a | −0.881 a | −0.816 a | −0.951 a | 0.732 a | 0.812 a | −0.853 a | −0.848 a | −0.854 a | 0.082 | 0.557 a | 0.931 a | 0.967 a | 0.884 a | 1 | |
| MDA | 0.932 a | −0.663 a | −0.578 a | −0.595 a | −0.650 a | −0.385 a | −0.215 | −0.508 a | 0.648 a | 0.937 a | −0.190 | −0.285 b | 0.811 a | 0.790 a | −0.352 a | 0.487 a | 0.416 a | −0.808 a | −0.793 a | −0.931 a | 0.752 a | 0.931 a | −0.976 a | −0.973 a | −0.972 a | 0.091 | 0.611 a | 0.979 a | 0.943 a | 0.983 a | 0.914 a | 1 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Qadir, S.U.; Raja, V.; Siddiqui, W.A.; Mahmooduzzafar; Abd_Allah, E.F.; Hashem, A.; Alam, P.; Ahmad, P. Fly-Ash Pollution Modulates Growth, Biochemical Attributes, Antioxidant Activity and Gene Expression in Pithecellobium Dulce (Roxb) Benth. Plants 2019, 8, 528. https://doi.org/10.3390/plants8120528
Qadir SU, Raja V, Siddiqui WA, Mahmooduzzafar, Abd_Allah EF, Hashem A, Alam P, Ahmad P. Fly-Ash Pollution Modulates Growth, Biochemical Attributes, Antioxidant Activity and Gene Expression in Pithecellobium Dulce (Roxb) Benth. Plants. 2019; 8(12):528. https://doi.org/10.3390/plants8120528
Chicago/Turabian StyleQadir, Sami Ullah, Vaseem Raja, Weqar Ahmad Siddiqui, Mahmooduzzafar, Elsayed F. Abd_Allah, Abeer Hashem, Pravej Alam, and Parvaiz Ahmad. 2019. "Fly-Ash Pollution Modulates Growth, Biochemical Attributes, Antioxidant Activity and Gene Expression in Pithecellobium Dulce (Roxb) Benth" Plants 8, no. 12: 528. https://doi.org/10.3390/plants8120528