Chloroplast Genome Sequence of Artemisia scoparia: Comparative Analyses and Screening of Mutational Hotspots
by
 , , and
, , and
Shabina Iram
1,* ,
,
Muhammad Qasim Hayat
1,*,
Muhammad Tahir
1,
Alvina Gul
1,
Abdullah
2 and
and
Ibrar Ahmed
3 1
Department of Plant Biotechnology, Atta-Ur-Rahman School of Applied Biosciences (ASAB), National University of Sciences and Technology (NUST), H-12 Islamabad 44000, Pakistan
2
Department of Biochemistry, Quaid-i-Azam University, Islamabad 45320, Pakistan
3
Alpha Genomics Private Limited, Islamabad 45710, Pakistan
*
Authors to whom correspondence should be addressed.
Plants 2019, 8(11), 476; https://doi.org/10.3390/plants8110476
Submission received: 20 September 2019
/
Revised: 14 October 2019
/
Accepted: 29 October 2019
/
Published: 6 November 2019
Abstract
:Artemisia L. is among the most diverse and medicinally important genera of the plant family Asteraceae. Discrepancies arise in the taxonomic classification of Artemisia due to the occurrence of multiple polyploidy events in separate lineages and its complex morphology. The discrepancies could be resolved by increasing the genomic resources. A. scoparia is one of the most medicinally important species in Artemisia. In this paper, we report the complete chloroplast genome sequence of Artemisia scoparia. The genome was 151,060 bp (base pairs), comprising a large single copy (82,834 bp) and small single copy (18,282 bp), separated by a pair of long inverted repeats (IRa and IRb: 24,972 bp each). We identified 114 unique genes, including four ribosomal RNAs, 30 transfer RNAs, and 80 protein-coding genes. We analysed the chloroplast genome features, including oligonucleotide repeats, microsatellites, amino acid frequencies, RNA editing sites, and codon usage. Transversion substitutions were twice as frequent as transition substitutions. Mutational hotspot loci included ccsA-ndhD, trnH-psbA, ndhG-ndhI, rps18-rpl20, and rps15-ycf1. These loci can be used to develop cost-effective and robust molecular markers for resolving the taxonomic discrepancies. The reconstructed phylogenetic tree supported previous findings of Artemisia as a monophyletic genus, sister to the genus Chrysanthemum, whereby A. scoparia appeared as sister to A. capillaris.
1. Introduction
The genus Artemisia L. includes over 500 species and occupies the top position in the family Asteraceae in terms of its bio-prospection [1]. Members of this genus are mainly hairy shrubs and herbs with a cosmopolitan distribution in arid and semi-arid habitats [2]. Western and central Asia is considered its centre of origin in the northern hemisphere, although some species are also found in the southern hemisphere [3]. Several species of Artemisia have wide and varied medicinal applications in the pharmaceutical industry, as well as in folk remedies [1].
The vernacular name of Artemisia scoparia Waldst. et. Kit (red stem wormwood) is Jhahoo or Jaukay in Pakistan [2]. It is a branched perennial herb with a bitter aroma [2]. The herb has a wild distribution in Southwest Asia and central Europe [4]. In northern Pakistan, A. scoparia is found growing up to an altitude of 4000 m in the summer season along the field boundaries, on stony grounds, rural tracks, and sandy soils of barren areas after rainfall [5].
Recently, A. scoparia has received immense attention due to its potential therapeutic impacts on indigenous communities. This species has been reported to contain anti-viral [6], anti-cancer [7], anti-inflammatory [8], anti-allergic [9], anti-oxidant, anti-malarial, insecticidal [10], anti-microbial [11], anti-hypertensive [12], and anti-obesity [13] properties. This species also has renal-protective [5], hepato-protective [14], hypo-lipidemic [15], and urease inhibitory [16] properties. Moreover, it has been utilized for curing Alzheimer’s disease [17].
The chloroplast (cp) genome in most of the plants has a characteristic quadripartite and circular structure, in which a small single copy region (SSC) and a large single copy region (LSC) are separated by a pair of large inverted repeats (IRs) [18,19]. Linear chloroplast genomes are also reported in some species [20]. The size of the cp genome ranges from 107 to 218 kb [21]. The cp genome is conservative in nature and is mostly inherited maternally [21]. Chloroplast genome polymorphism has been used in phylogenetic reconstructions from population genetics [22] to investigate deep divergence at a genera and family level [23,24,25,26]. The slow evolving nature and maternal inheritance of the chloroplast genome in comparison to the nuclear genome make it suitable for such studies [21]. Therefore, the chloroplast genome structure can be exploited for the taxonomic classification, inferring of phylogeny, and molecular barcoding of the medicinal plants like A. scoparia. Knowledge about the chloroplast genome structure is also helpful in the transformation of foreign genes to obtain high protein expression [21,27,28].
Based on its extensive medicinal applications and ecological value, it is important to further explore the genetic and phylogenetic characteristics of A. scoparia. Highly efficient molecular markers are required to select pharmaceutically efficient germplasm for large-scale breeding purposes in Artemisia [27]. The acquisition of genomic information can help to develop medicinally potent genetic breeds of the plant. Traditionally, the taxonomic identification of Artemisia species has relied on morphological characteristics. However, controversies with regards to the taxonomic relationships have arisen due to the plastic nature, complex genetics, and polyploidy of Artemisia species, limiting the role of morphological characteristics as the sole factor for classification [29,30]. The development of high-resolution genetic markers is important for fostering attempts regarding species identification, inferring phylogeny, and improving the medicinal value of A. scoparia.
In this study, we de novo assembled the chloroplast genome of A. scoparia and compared the chloroplast genome features of nine Artemisia species. We analysed the codon usage, amino acid frequency, RNA editing sites, and repeats in the A. scoparia genome. We also identified twenty polymorphic loci which might be appropriate for the design of authentic, cost-effective, and high-efficacy molecular markers. The phylogeny was inferred to understand the phylogenetic position of A. scoparia. The findings of our study are expected to be useful in phylogenetics, population genetics, and genetic engineering studies of A. scoparia.
2. Results
2.1. Chloroplast Genome Features of A. scoparia
Hiseq2500 generated 6.6 GB sequencing data that contained 18.54 million reads from the paired-end run. This whole genome sequence data was used to de novo assemble the chloroplast genome of A. scoparia with an average coverage depth of 64 ×. The Artemisia scoparia chloroplast genome had a quadripartite structure with a length of 151,060 bp (base pairs). The chloroplast genome contained an LSC (82,834 bp) region and an SSC (18,282 bp) region, which were separated by two copies of IRs (IRa and IRb: 24,972 bp each).
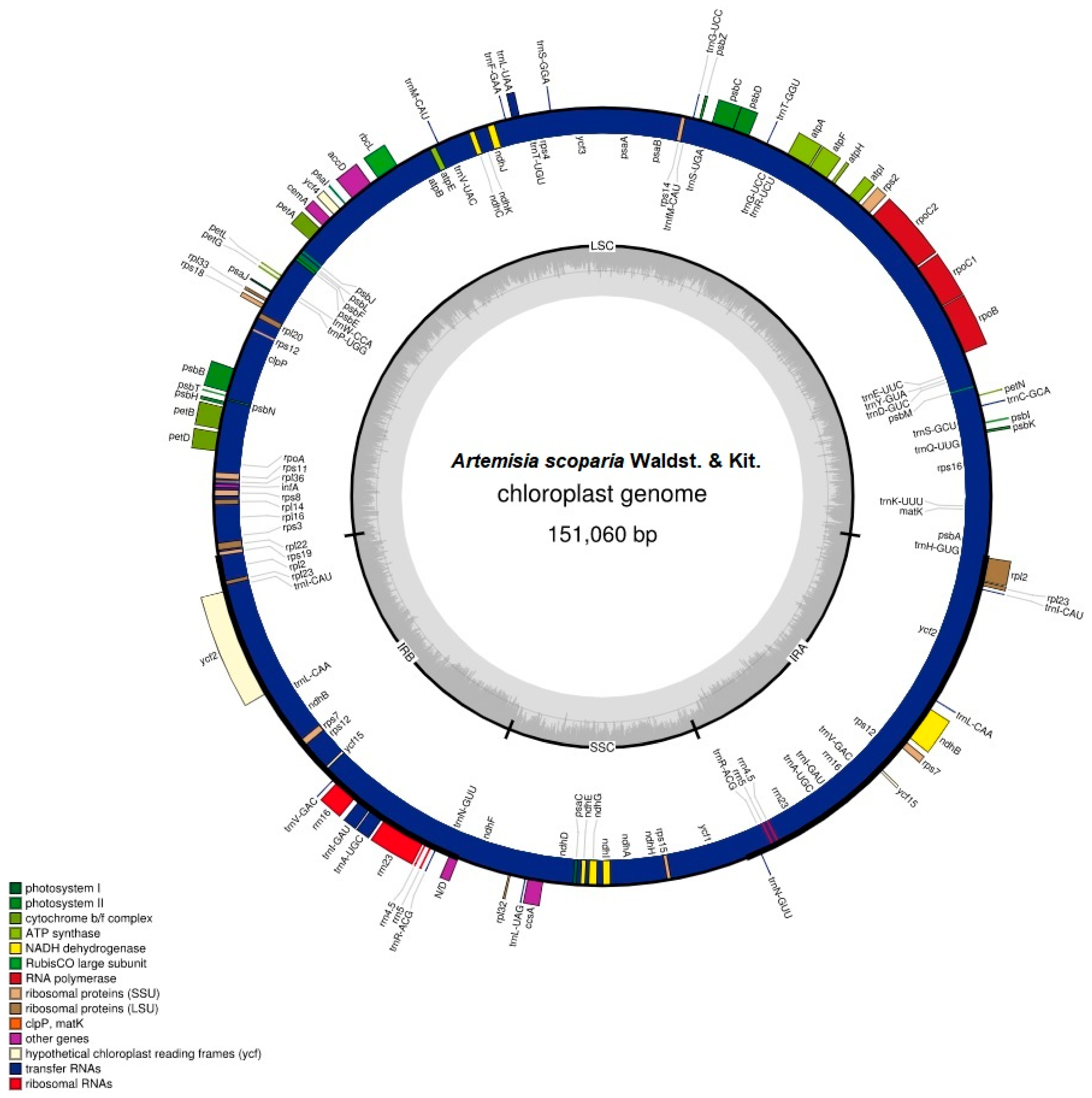
The chloroplast genome contained 114 unique genes in which 18 genes were also duplicated in IRs, except for the pseudogene copy of ycf1Ψ. Among 114 unique genes, 30 were tRNA genes, 4 were rRNA genes, and 80 were protein-coding genes. Eighteen genes that were duplicated in IRs comprised 4 rRNA genes, 7 tRNA genes, and 7 protein-coding genes. We also found 18 intron-containing genes that included 6 tRNA genes and 12 protein-coding genes. Among the 18 genes, 16 genes contained one intron, whereas 2 genes (ycf3 and clpP) contained two introns. The rps12 gene was a trans-splicing gene in which the 5′ end existed in the LSC as a single copy, whereas the 3′ existed in the IRs regions in duplicate. The ycf1 gene started from the IRa region and extended into the SSC region, and thus left a truncated copy of ycf1Ψ at the border of the IRb region. The arrangement of genes in the chloroplast genome has been shown in Figure 1. The gene content of the genome has been shown, along with their function, in Table S1.
The chloroplast genome of A. scoparia is AT-rich. The complete chloroplast genome is comprised of 62.5% AT content and 37.5% GC content. The GC content of the three regions varies in the genome. The IR regions contain a high GC content (43.1%), compared to the LSC (35.6%) and SSC (30.8%) regions. The high GC content of IRs is due to the presence of high GC-comprising genes: rRNAs (55%) and tRNAs (52.7%). The genomic features have been provided in detail in Table 1.
2.2. Codon Usage and Amino Acid Frequency
Codon usage analyses revealed a high encoding of codons that contained A or T at the 3′ end among synonymous codons compared to codons that ended with C or G (Table S2). The amino acid frequency analyses revealed an abundance of leucine and isoleucine, whereas cysteine was the least abundant amino acid (Figure S1).
2.3. RNA Editing Sites
The predictive RNA editor for plant chloroplast genes (PREP-cp) predicted 51 post-transcriptional RNA editing modifications in 21 protein-coding genes. Most of the RNA editing sites were found in ndhB (9 editing sites) and ndhD (6 editing sites), whereas matK, rpoC1, and accD contained four RNA editing sites each. All the changes took place on the first and second nucleotide of codons. However, the conversion rate at the first nucleotide was observed to be about four times higher than that at the second nucleotide. Most of the RNA editing sites resulted in the conversion of serine to leucine and phenylalanine. Moreover, most of the conversion led to hydrophobic amino acids, including methionine, phenylalanine, valine, leucine etc. (Table S3).
2.4. Comparison of Genus Artemisia Genomes and the Contraction and Expansion of Large Inverted Repeats
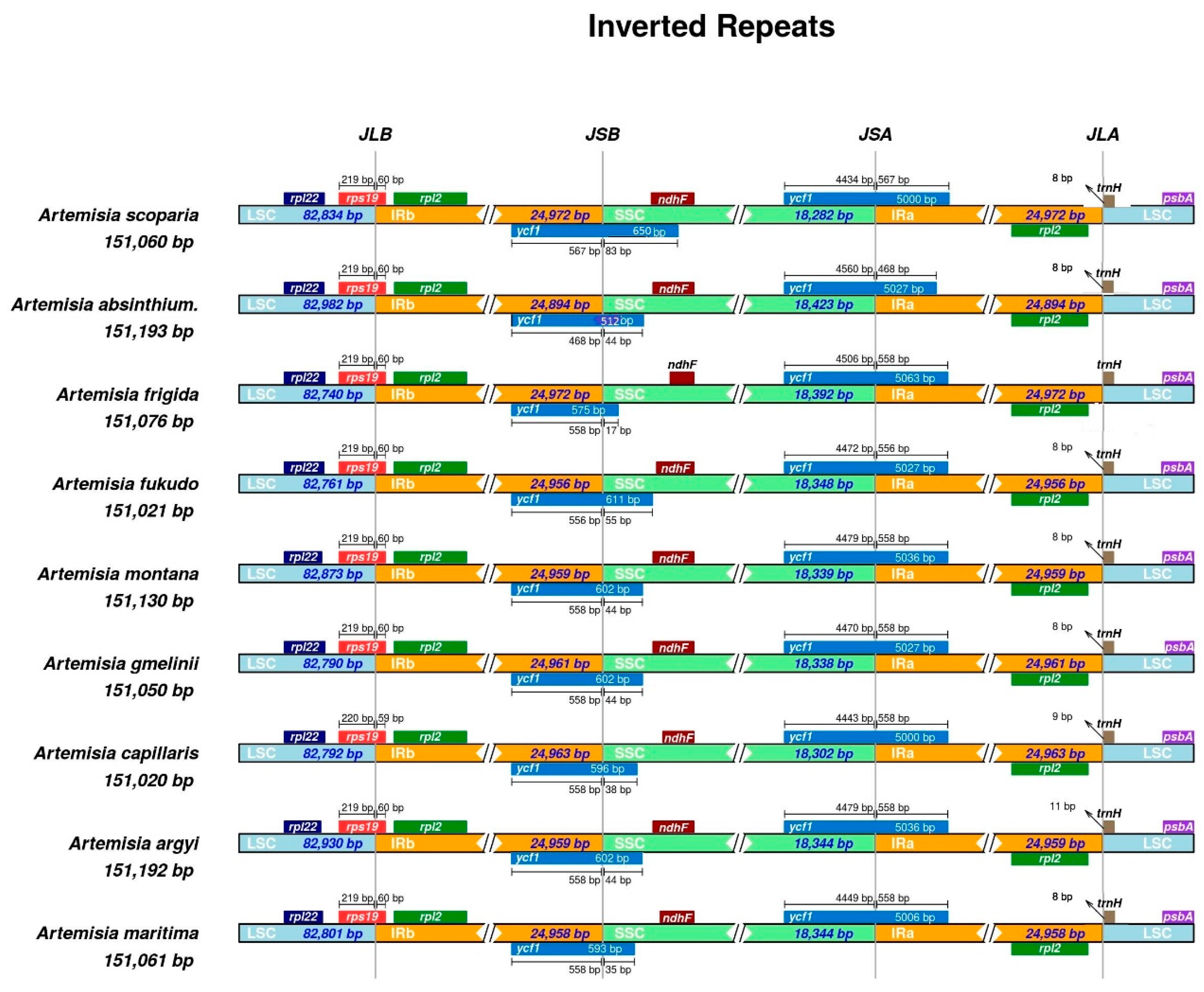
We compared the chloroplast genome features of nine Artemisia species and also evaluated the phenomenon of the IR contraction and expansion. The size of the complete chloroplast genome varied from 151,011 bp (in A. fukudo) to 151,318 bp (in A. gmelinii); the LSC region varied from 82,740 bp (in A. frigida) to 83,061 bp (in A. gmelinii); the SSC region varied from 18,282 bp (in A. scoparia) to 18,423 bp (in A. absinthium); and each IR varied from 24,892 bp (in A. absinthium) to 24,972 bp (in A. frigida and A. scoparia) (Table S4).
The IR contraction and expansion were also evaluated among nine species of the genus Artemisia. The pseudogene of rps19Ψ was observed at the junction of LSC and Ira, whereas the pseudogene of ycf1Ψ was observed at the junction of IRb and SSC. The rps19 gene integrated from the LSC region to the IRb region with 59 to 60 bp extending into the IRb region, leaving rps19Ψ at junctions of IRa and LSC. The size of ycf1Ψ varied from 512 bp (in A. absinthium) to 650 bp (in A. scoparia) and integrated from the IRb region to SSC region with 17 bp (in A. frigida) to 83 bp (in A. scoparia). The ndhF gene was present at the junction of SSC and IRb completely in the SSC region, but also overlapped with ycf1Ψ up to 30 bp in Artemisia scoparia. The gene trnH at the junction of IRa and LSC remained completely in the LSC region, whereas the rpl2 gene remained completely in the repeated region at the junction of LSC and IRb/IRa. The graphical representation of IR contraction and expansion has been shown in Figure 2.
2.5. Microsatellite and Oligonucleotide Repeats Analyses
We used the Perl script of MIcroSAtellite identification tools (MISA) for the identification of microsatellite repeats and detected 58 microsatellite repeats and eight compound repeats. All compound repeats were found in LSC regions. Among the 58 microsatellite repeats, 45 repeats existed in the LSC region, 4 in IR regions, and 9 in the SSC region. The five types of repeats were detected and varied in number, such as Mononucleotide (40) > tetranucleotide (11) > dinucleotide (9) > trinucleotide (5) > pentanucleotide (1). The mononucleotide repeats were made of 10–20 repeat units, dinucleotide repeats were made of 5–7 repeat units, trinucleotide repeats were made of 4–5 repeat units, tetranucleotide repeats were made of 3–4 repeat units, and the pentanucleotide repeat was made of 3 repeat units. All types of repeats were AT-rich (Table 2). The position, type, and region of each microsatellite has been given Table S5.
We used the REPuter program to detect oligonucleotide repeats. The REPuter program detected 14 oligonucleotide repeats, including 10 forward repeats, three palindromic repeats, and one reverse repeat. The size of repeats varied from 30 to 60 bp. The LSC region contained five repeats, IR region contained three repeats, and SSC region contained one repeat. Moreover, three repeats were shared between LSC and IR, one repeat in LSC and SSC, and one repeat in SSC and IR. As per further categorization based on the functional regions of the chloroplast genome, six repeats were observed in intergenic spacer regions, four in coding regions, and two in intronic regions, and two repeat pairs were shared in intronic and intergenic spacer regions (Table 3).
2.6. Types of Substitutions and InDels Events
We analysed the types of substitutions and InDel events by using A. scoparia as a reference in pairwise alignment with the A. capillaris cp genome. We found 88 substitutions in the complete chloroplast genome: 59 substitutions were found in LSC, 23 in SSC, and 6 in IR regions. Among a total of 54 InDels, 42 were in LSC, 9 in SSC, and 3 in IR regions (Table 4).
There were 29 transition substitutions and 59 transversion substitutions. Therefore, we observed a transitions to transversions (Ts/Tv) ratio of 0.49. The Ts/Tv values were 0.49 in the LSC region, 0.55 in the SSC region, and 5 × (five among six were transitions) in IR regions.
2.7. Screening of Divergence Regions
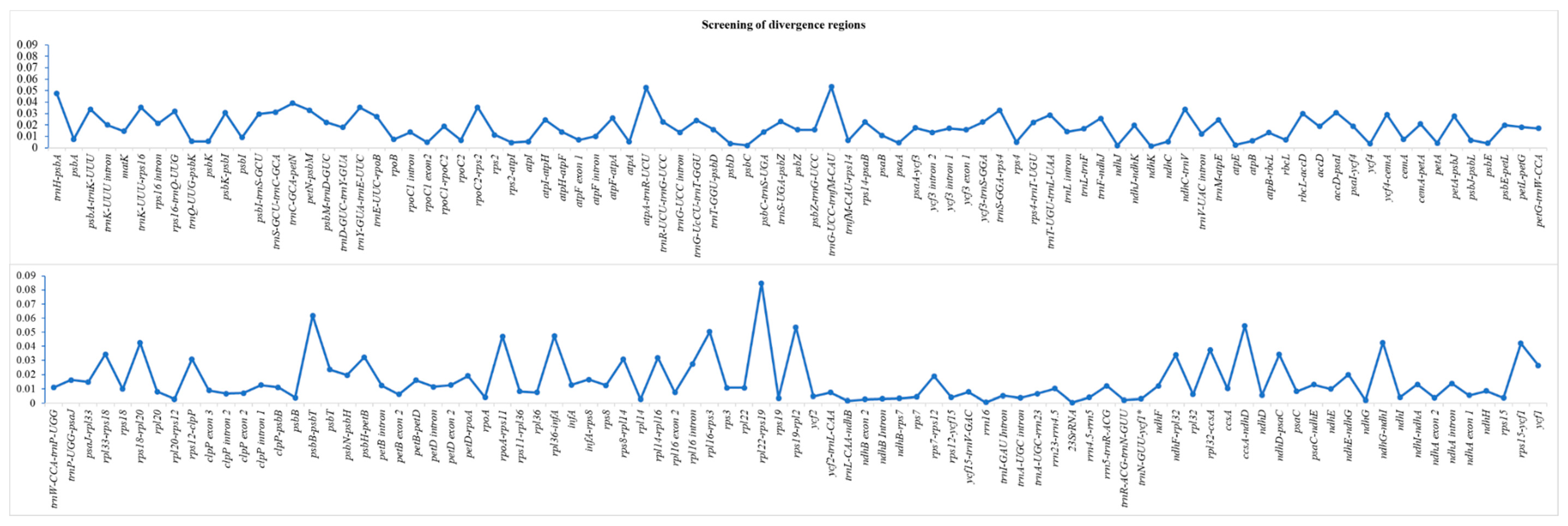
We conducted a multiple sequence alignment of complete chloroplast genomes of nine Artemisia species and screened intergenic spacer regions, intronic regions, and protein-coding sequences for the identification of highly diverse regions. The average divergence (π) in complete chloroplast genomes was 0.014. The highest average divergence was recorded for intergenic spacer regions (0.022), followed by introns (0.012) and then protein-coding sequences (0.005). The nucleotide diversity of each region has been shown in Figure 3. We also identified twenty mutational hotspots from intergenic spacer regions that might be adopted as appropriate loci for population genetic and phylogeographic studies. Notable among these loci are ccsA-ndhD, trnH-psbA, ndhG-ndhI, rps18-rpl20, and rps15-ycf1 (Table 5). Among protein-coding genes, ycf1, psbD, and accD exhibited remarkable polymorphism.
2.8. Inferring of Phylogeny in the Genus Artemisia and Family Asteraceae
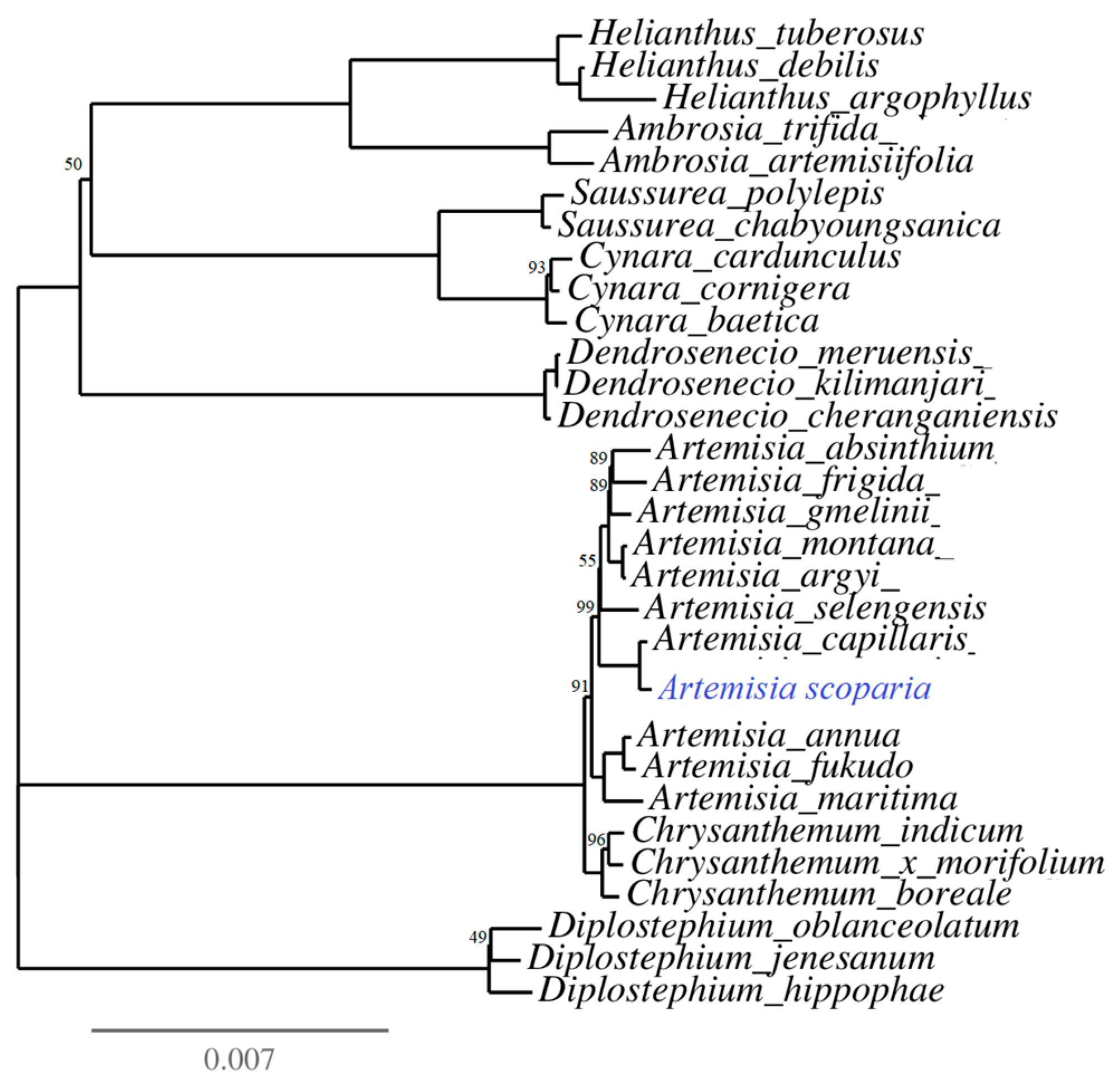
The phylogenetic tree was reconstructed for 30 species of the family Asteraceae (Table S6). The phylogeny was inferred based on coding sequences of 77 protein-coding genes using the IQ-tree online program with the best fit model TVM + F + I + G4. The multiple sequence alignment consisted of 57,226 nucleotide sites, in which 53,270 (93.09%) nucleotide sites were constant (invariant sites), 3360 were parsimony informative sites, and 970 showed a distinct pattern. The phylogeny analyses showed Artemisia as a monophyletic genus which was sister to the genus Chrysanthemum. Moreover, A. scoparia was found as sister taxa to A. capillaris (Figure 4).
3. Discussion
In the current study, we de novo assembled the chloroplast genome of A. scoparia and compared its genome with the chloroplast genome of eight publicly available Artemisia species. We evaluated the IR contractions and expansion phenomenon in these species, which revealed the origination of the pseudogene ycf1Ψ. We also analysed types of substitutions, InDels events, and repeats within the genomes.
All nine species of the genus Artemisia showed similarity in terms of the gene content, intron content, and composition of GC content. The chloroplast genome of angiosperms is highly conserved and several previous studies have also reported high similarity in chloroplast genome sequences of the same lineage [26,31,32,33,34,35]. However, some studies have reported variation in gene numbers and gain or loss of introns at a genus level, as well as at a family level [36,37,38]. We found a functional infA gene, encoding the translation initiation factor, in all species of Artemisia, whereas in some other plant lineages, the complete loss or non-function copy of the infA gene has been reported [37,38,39].
RNA editing sites are commonly reported in protein-coding genes of the chloroplast. We identified 51 RNA editing sites in 21 protein-coding genes of A. scoparia. We observed most of the conversion for serine to leucine and conversion of hydrophilic amino acids into hydrophobic amino acids. Our study is in line with previous studies of other angiosperms in which a similar effect of RNA editing events has been reported [26,36,37].
The contraction and expansion of IR regions, along with the variation in the length of the intergenic spacer region, caused variation in the length of the chloroplast genome, even within same lineages at a family level, as well as at a genus level [26,36,40,41]. The contraction and expansion of the IR regions lead to the duplication of genes, to the loss of one copy of genes, or to the origination of pseudogenes in the chloroplast genome of angiosperms [37,38,42,43]. Our results are also in agreement with these studies and we observed the origination of ycf1Ψ and rps19Ψ at the junction of IRb/SSC and LSC/IRa, respectively. The IR contraction and expansion might be helpful in the study of evolutionary patterns [38] and the closely related species show a high resemblance at the junction of the chloroplast genome compared to more diverse species [44]. Our study also agreed with this study and the species of Artemisia revealed a closed resemblance at junctions of the chloroplast genome, but this view needs further confirmation in more diverse species (a family level comparison).
We analysed microsatellites and oligonucleotide repeats in the genome of A. scoparia. Microsatellites are good markers in studies of population genetics of plant species. Therefore, the identified loci might be suitable for the population genetic studies of A. scoparia. The identified microsatellite loci showed an abundance of A/T content, which might be due to the AT-rich composition of the chloroplast genome [36]. In the current study, the mononucleotide repeats were followed by tetranucleotide repeats. However, in other Artemisia species, the abundance of dinucleotide repeats was also reported after mononucleotide repeats [45]. In other plant lineages of angiosperm, an abundance of trinucleotide repeats was also found after mononucleotide repeats [36,37]. We found most of the repeats in the LSC region compared to SSC and IR regions. These results are also in agreement with several other studies of angiosperms [36,46,47,48,49].
We also analysed oligonucleotide repeats in the chloroplast genome of A. scoparia. Oligonucleotide repeats are considered to produce substitutions, deletion, inversion, addition, and rearrangements within the genome [22,37,39,50,51]. Moreover, these repeats could also be used for the identification of polymorphic loci, as suggested previously [39,51]. Most of the repeats were found in intergenic spacer regions compared to intronic regions and protein-coding sequences. Our results agreed with previous studies of the genus Artemisia and other species of angiosperm [26,27,36,40,45,52]. However, protein-coding sequences are also reported to contain the highest number of oligonucleotide repeats [38].
Previous studies reported a higher existence of InDels and substitutions in LSC and SSC regions compared to IR regions [36,39,45,53]. In the current study, an abundance of substitutions and InDels was also observed in LSC and SSC regions compared to IR regions. The most common types of substitutions that occurred in the chloroplast genome were transversion substitutions. In the current study, the Ts/Tv ratio was found to be 0.49, which indicates higher transversion substitutions compared to transition substitutions and is also in agreement with previous studies of angiosperm chloroplast genomes in which the authors reported Ts/Tv < 1 [36,45,54].
Commonly used molecular markers, including ITS1, ITS2, matK, trnH-psbA, rpoC1, rbcL, and rpoB, did not well-resolve the phylogenetic relationships among closely related species of Artemisia [29,55,56]. Therefore, these loci could not be employed as barcodes. Moreover, certain other limitations make them unsuitable for barcoding, including their problematic, time consuming, and less robust amplification process and inefficient sequencing [57]. The two approaches that have been suggested for the barcoding of plant species are the use of the whole chloroplast genome as a long barcode [57] or use of molecular markers from mutational hotspots [23,57]. The whole chloroplast genome can be the best choice for the barcoding of plant species as it shows significance polymorphism. However, sequencing of the chloroplast genome is not cost-effective and cannot be preferred. Therefore, we followed the second approach, which was also preferred by other researchers [26,36,37,38,45]. Recently, Meng et al. [58] reported mutational hotspots based on a comparative analyses of the chloroplast genome using mVista, whereas Shahzadi et al. [45] reported twenty mutational hotspots based on comparative analyses of four highly diverse species using multiple MAFFT alignment. Shahzadi et al. [45] used a more technical approach. However, they did not utilize certain genomic resources. In the current study, we utilized nine genomes of Artemisia by also including those species that were embedded among the diverse species of Artemisia and identified twenty mutational hotspots. Among the twenty mutational hotspots, sixteen mutational hotspots were those identified by Shahzadi et al. [45]. However, the mutational hotspots varied in the extent of divergence. The four mutational hotspots that differed from the study of Shahzadi et al. [45] were psbA-trnK, trnS-rps4, psbK-psbI, and ycf4-cemA. Therefore, our study confirmed the suitability of a few highly diverse species for the identification of suitable polymorphic loci to design cost-effective, authentic, and robust markers.
The inferring of phylogeny revealed that the genus Artemisia is a monophyletic genus that is sister to the genus Chrysanthemum. A similar result has also been provided by Shahzadi et al. [45] based on the 77 protein-coding sequences of the chloroplast genome.
In summary, our study provides insight into the chloroplast genome structure of the highly medicinal species A. scoparia. The genomic features revealed a close resemblance among Artemisia species. The identified microsatellite and twenty mutational hotspots might be helpful for studies of population genetics and phylogenetics of the genus Artemisia. Moreover, our study confirms the suitability of a few highly diverse species for the identification of suitable polymorphic loci for the development of effective markers.
4. Materials and Methods
4.1. DNA Extraction and Sequencing
The plant of Artemisia scoparia was collected from District Attock in the Potohar region of Pakistan. Leaf tissues that did not show any apparent disease symptoms were collected from the plant of A. scoparia. The leaf tissues were washed with 70% ethanol and silica dried for future DNA extraction. Whole genomic DNA was extracted by the DNeasy Plant Mini Kit (Qiagen, Inc., Germany). The quality and quantity of DNA was determined by 1% agarose gel electrophoresis and Multiskan GO (Thermo Scientific Inc., USA). The high-quality DNA was sent to Novogene, Hong Kong and sequenced from paired ends with 150 bp short reads and a 350 bp insert size using HiSeq2500 (Illumina, USA).
4.2. Genome Assembly and Annotation
The chloroplast genome of A. scoparia was de novo assembled following Abdullah et al. [37], using Kmer values of 121, 111, 91, 71, and 61 in Velvet 1.2.10 [59]. The coverage depth analyses were performed with Burrow wheal aligner (BWA) [60] and boundaries of LSC, SSC, and IR (IRa and IRb) regions were defined by the manual visualization of scaffolds in Geneious R8.1 [61], following Abdullah et al. [37]. The de novo assembled genome was annotated using the GeSeq annotation tool [62], whereas the transfer RNA genes were further verified by tRNAscan-SE version 2.0.3 [63] and ARAGORN v.1.2.38 [64]. The circular diagram of the chloroplast genome was drawn by Organellar Genome DRAW [65]. The required five-column tab-delimited table of genome annotations used to submit our sequence to the National Centre for Biotechnology Information (NCBI) was generated by GB2sequin [66]. GenBank assigned an accession number of MN385624.
4.3. Comparative Analyses and Genomic Features
The genomic features of A. scoparia were determined in Geneious R8.1 [61], which compared its genomic features with eight other publicly available species of the genus Artemisia. The contraction and expansion of inverted repeat regions were also examined among the nine species of the genus Artemisia using IRscope [67].
4.4. Amino Acid Frequency, Codon Usage, and RNA Editing Sites
4.5. Microsatellites and Oligonucleotide Repeats Analyses
We used Perl script MIcroSAtellite identification tools (MISA) [69] to determine microsatellites in the cp genome of A. scoparia. The parameters adjusted for microsatellite detection included 10 for mono-; 5 for di-; 4 for tri-; and 3 each for tetra-, penta-, and hexanucleotide repeats.
To detect oligonucleotide repeats, we used the REPuter [70] program to detect four different types of repeats: forward (F), reverse (R), palindromic (P), and complementary (C) repeats. The parameters of the oligonucleotide program included a minimum repeat size of 30 bp with three mismatches to detect repeats with a minimum of 90% similarities.
4.6. Substitutions and InDel Analyses
We determined the numbers and types of substitutions along with the InDels event in each region of the chloroplast genome. We used A. capillaris as a reference for A. scoparia following Shahzadi et al. [45] and pairwise aligned IR, LSC, and SSC regions of both genomes using MAFFT v.5 (Multiple Alignment with Fast Fourier Transform) [71]. The numbers and types of substitutions were described in Geneious R8.1 [61]. The numbers of InDels events were determined by analysing the pairwise alignment of each region in DnaSP v.5.10 [72].
4.7. Screening of Divergence Regions
We determined the extent of divergence of the protein-coding sequences, intronic regions, and intergenic spacer regions. For this purpose, we multiple aligned nine species of the genus Artemisia by MAFFT [71] and counted the number of substitutions and InDels events within each region by visualizing them in Geneious R8.1 [61], following Shahzadi et al. [45]. The nine species that were included in the comparison for the determination of divergence regions are provided in Figure 2.
4.8. Phylogeny in the Genus Artemisia and Family Asteraceae
We inferred the phylogeny of the genus Artemisia and family Asteraceae among thirty species following the methodological approach of Shahzadi et al. [45] and reconstructed the maximum likelihood tree using the IQ-TREE [73,74,75] program, whereas TreeDyn was used to enhance the phylogenetic tree visualization and analyses [76,77].
Supplementary Materials
The following are available online at https://www.mdpi.com/2223-7747/8/11/476/s1. Table S1: Gene content of the chloroplast genome of Artemisia scoparia, Table S2: Analyses of codon usage in A. scoparia, Table S3: RNA editing sites in A. scoparia, Table S4: Comparison of chloroplast genome features of eight other Artemisia species Table S5: Microsatellite loci in Artemisia scoparia, Table S6: Species used when inferring phylogeny, Figure S1: Amino acid frequency in Artemisia scoparia.
Author Contributions
Conceptualization, S.I. and M.Q.H.; visualization, S.I.; methodology, S.I., I.A., and A.; resources, I.A. and M.Q.H.; formal analysis, S.I., I.A., and A.; data curation, S.I. and A.; investigation, S.I.; writing—original draft preparation, S.I.; writing—review and editing, S.I., A., and I.A.; supervision, M.Q.H., A.G., and M.T.; funding acquisition, M.T.; project administration, M.T., A.G., and M.Q.H.
Funding
The Research Directorate of the National University of Sciences and Technology (NUST), H-12 Islamabad, Pakistan provided funds for this study.
Conflicts of Interest
There are no conflicts of interest between the authors.
References
- Pandey, A.K.; Singh, P. The genus Artemisia: A 2012–2017 literature review on chemical composition, antimicrobial, insecticidal and antioxidant activities of essential oils. Medicines 2017, 4, 68. [Google Scholar] [CrossRef]
- Hayat, M.Q.; Khan, M.A.; Ashraf, M.; Jabeen, S. Ethnobotany of the genus Artemisia L. (Asteraceae) in Pakistan. Ethnobot. Res. Appl. 2009, 7, 147–162. [Google Scholar] [CrossRef]
- Valles, J.; Durant-McArthur, E. Artemisia systematics and phylogeny: Cytogenetic and molecular insights. In Proceedings of the Shrubland Ecosystem Genetics and Biodiversity, Provo, UT, USA, 13–15 June 2000; pp. 67–74. [Google Scholar]
- Abad, M.J.; Bedoya, L.M.; Apaza, L.; Bermejo, P. The Artemisia L. genus: A review of bioactive essential oils. Molecules 2012, 17, 2542–2566. [Google Scholar] [CrossRef]
- Sajid, M.; Khan, M.R.; Shah, N.A.; Ullah, S.; Younis, T.; Majid, M.; Ahmad, B.; Nigussie, D. Proficiencies of Artemisia scoparia against CCl 4 induced DNA damages and renal toxicity in rat. BMC Complement. Altern. Med. 2016, 16, 1–10. [Google Scholar] [CrossRef]
- Wang, L.; Si, L.; Li, Y.; Wang, H.; Xu, F.; Bian, H.; Shi, Y.; Huang, H. Study on mechanism and active ingredient of Artemisia scoparia extracts against influenza virus. Lat. Am. J. Pharm. 2017, 36, 1355–1360. [Google Scholar]
- Choi, E.; Park, H.; Lee, J.; Kim, G. Anticancer, antiobesity, and anti-inflammatory activity of Artemisia species in vitro. J. Tradit. Chin. Med. 2013, 33, 92–97. [Google Scholar] [CrossRef]
- Nam, S.Y.; Han, N.R.; Rah, S.Y.; Seo, Y.; Kim, H.M.; Jeong, H.J. Anti-inflammatory effects of Artemisia scoparia and its active constituent, 3,5-dicaffeoyl-epi-quinic acid against activated mast cells. Immunopharmacol. Immunotoxicol. 2018, 40, 52–58. [Google Scholar] [CrossRef]
- Ryu, K.J.; Yoou, M.S.; Seo, Y.; Yoon, K.W.; Kim, H.M.; Jeong, H.J. Therapeutic effects of Artemisia scoparia Waldst. et Kitaib in a murine model of atopic dermatitis. Clin. Exp. Dermatol. 2018, 43, 798–805. [Google Scholar] [CrossRef]
- Afshar, F.H.; Delazar, A.; Janneh, O.; Nazemiyeh, H.; Pasdaran, A.; Nahar, L.; Sarker, S.D. Evaluation of antimalarial, free-radicalscavenging and insecticidal activities of Artemisia scoparia and A. spicigera, Asteraceae. Braz. J. Pharmacogn. 2011, 21, 986–990. [Google Scholar] [CrossRef]
- Baykan Erel, Ş.; Reznicek, G.; Şenol, S.G.; Karabay Yavaşoǧulu, N.Ü.; Konyalioǧlu, S.; Zeybek, A.U. Batı Anadolu Artemisia L. türlerinin antimikrobiyal ve antioksidan özellikleri. Turk. J. Biol. 2012, 36, 75–84. [Google Scholar] [CrossRef]
- Cho, J.Y.; Park, K.H.; Hwang, D.Y.; Chanmuang, S.; Jaiswal, L.; Park, Y.K.; Park, S.Y.; Kim, S.Y.; Kim, H.R.; Moon, J.H.; et al. Antihypertensive effects of Artemisia scoparia Waldst in spontaneously hypertensive rats and identification of angiotensin I converting enzyme inhibitors. Molecules 2015, 20, 19789–19804. [Google Scholar] [CrossRef]
- Richard, A.J.; Fuller, S.; Fedorcenco, V.; Beyl, R.; Burris, T.P.; Mynatt, R.; Ribnicky, D.M.; Stephens, J.M. Artemisia scoparia enhances adipocyte development and endocrine function in vitro and enhances insulin action in vivo. PLoS ONE 2014, 9, e98897. [Google Scholar] [CrossRef]
- Waris, T.S.; Khan, M.R.; Shah, N.A.; Shuaib, M.; Hussain, F.; Ishaq, M.; Kamal, Z.; Zeb, U.; Ali, S.; Ali, K.; et al. Heptoprotective role of Artemisia scoparia Waldst. and kit against CCl 4 -induced toxicity in rats. Pol. J. Environ. Stud. 2018, 27, 1307–1314. [Google Scholar] [CrossRef]
- Boudreau, A.; Richard, A.J.; Burrell, J.A.; King, W.T.; Dunn, R.; Schwarz, J.M.; Ribnicky, D.M.; Rood, J.; Salbaum, J.M.; Stephens, J.M. An ethanolic extract of Artemisia scoparia inhibits lipolysis in vivo and has antilipolytic effects on murine adipocytes in vitro. Am. J. Physiol. Endocrinol. Metab. 2018, 315, E1053–E1061. [Google Scholar] [CrossRef]
- Khan, M.A.; Khan, H.; Tariq, S.A.; Pervez, S. Urease inhibitory activity of aerial parts of Artemisia scoparia: Exploration in an in vitro study. Ulcers 2014, 2014, 1–5. [Google Scholar] [CrossRef]
- Promyo, K.; Cho, J.Y.; Park, K.H.; Jaiswal, L.; Park, S.Y.; Ham, K.S. Artemisia scoparia attenuates amyloid β accumulation and tau hyperphosphorylation in spontaneously hypertensive rats. Food Sci. Biotechnol. 2017, 26, 775–782. [Google Scholar] [CrossRef] [PubMed]
- Khan, A.; Asaf, S.; Khan, A.L.; Al-Harrasi, A.; Al-Sudairy, O.; AbdulKareem, N.M.; Khan, A.; Shehzad, T.; Alsaady, N.; Al-Lawati, A.; et al. First complete chloroplast genomics and comparative phylogenetic analysis of Commiphora gileadensis and C. foliacea: Myrrh producing trees. PLoS ONE 2019, 14, e0208511. [Google Scholar] [CrossRef] [PubMed]
- Palmer, J.D. Comparative organization of chloroplast genomes. Annu. Rev. Genet. 1985, 19, 325–354. [Google Scholar] [CrossRef]
- Oldenburg, D.J.; Bendich, A.J. The linear plastid chromosomes of maize: Terminal sequences, structures, and implications for DNA replication. Curr. Genet. 2016. [Google Scholar] [CrossRef]
- Daniell, H.; Lin, C.-S.; Yu, M.; Chang, W.-J. Chloroplast genomes: Diversity, evolution, and applications in genetic engineering. Genome Biol. 2016, 17, 134. [Google Scholar] [CrossRef]
- Ahmed, I. Evolutionary Dynamics in Taro. Ph.D. Thesis, Massey University, Palmerston North, New Zealand, August 2014. Available online: https://mro.massey.ac.nz/handle/10179/5610 (accessed on 24 June 2019).
- Ahmed, I.; Matthews, P.J.; Biggs, P.J.; Naeem, M.; Mclenachan, P.A.; Lockhart, P.J. Identification of chloroplast genome loci suitable for high-resolution phylogeographic studies of Colocasia esculenta (L.) Schott (Araceae) and closely related taxa. Mol. Ecol. Resour. 2013, 13, 929–937. [Google Scholar] [CrossRef] [PubMed]
- Henriquez, C.L.; Arias, T.; Pires, J.C.; Croat, T.B.; Schaal, B.A. Phylogenomics of the plant family Araceae. Mol. Phylogenet. Evol. 2014, 75, 91–102. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zhang, Z.; Yang, J.; Lv, G. Complete chloroplast genome of seven Fritillaria species, variable DNA markers identification and phylogenetic relationships within the genus. PLoS ONE 2018, 13. [Google Scholar] [CrossRef] [PubMed]
- Mehmood, F.; Abdullah; Shahzadi, I.; Ahmed, I.; Waheed, M.T.; Mirza, B. Characterization of Withania somnifera chloroplast genome and its comparison with other selected species of Solanaceae. Genomics 2019. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Huo, N.; Dong, L.; Wang, Y.; Zhang, S.; Young, H.A.; Feng, X.; Gu, Y.Q. Complete chloroplast genome sequences of Mongolia medicine Artemisia frigida and phylogenetic relationships with other plants. PLoS One 2013, 8. [Google Scholar] [CrossRef]
- Lössl, A.G.; Waheed, M.T. Chloroplast-derived vaccines against human diseases: Achievements, challenges and scopes. Plant Biotechnol. J. 2011, 9, 527–539. [Google Scholar] [CrossRef]
- Haghighi, A.R.; Belduz, A.O.; Vahed, M.M.; Coskuncelebi, K.; Terzioglu, S. Phylogenetic relationships among Artemisia species based on nuclear ITS and chloroplast psbA-trnH DNA markers. Biologia 2014, 69, 834–839. [Google Scholar] [CrossRef]
- Turuspekov, Y.; Genievskaya, Y.; Baibulatova, A.; Zatybekov, A.; Kotuhov, Y.; Ishmuratova, M.; Imanbayeva, A.; Abugalieva, S. Phylogenetic taxonomy of Artemisia L. species from Kazakhstan based on matK analyses. Proc. Latv. Acad. Sci. Sect. B Nat. Exact Appl. Sci. 2018, 72, 29–37. [Google Scholar] [CrossRef]
- Jiang, D.; Zhao, Z.; Zhang, T.; Zhong, W.; Liu, C.; Yuan, Q.J.; Huang, L. The chloroplast genome sequence of Scutellaria baicalensis provides insight into intraspecific and interspecific chloroplast genome diversity in Scutellaria. Genes 2017, 8, 227. [Google Scholar] [CrossRef]
- Saina, J.K.; Li, Z.Z.; Gichira, A.W.; Liao, Y.Y. The complete chloroplast genome sequence of tree of heaven (Ailanthus altissima (Mill.) (Sapindales: Simaroubaceae), an important pantropical tree. Int. J. Mol. Sci. 2018, 19, 929. [Google Scholar] [CrossRef]
- Park, I.; Yang, S.; Kim, W.; Song, J.-H.; Lee, H.-S.; Lee, H.; Lee, J.-H.; Ahn, S.-N.; Moon, B. Sequencing and comparative analysis of the chloroplast genome of Angelica polymorpha and the development of a novel indel marker for species identification. Molecules 2019, 24, 1038. [Google Scholar] [CrossRef] [PubMed]
- Li, D.-M.; Zhao, C.-Y.; Liu, X.-F. Complete chloroplast genome sequences of Kaempferia Galanga and Kaempferia Elegans: Molecular structures and comparative analysis. Molecules 2019, 24, 474. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Rong, C.; Qin, L.; Mo, C.; Fan, L.; Yan, J.; Zhang, M. Complete chloroplast genome sequence of Malus hupehensis: Genome structure, comparative analysis, and phylogenetic relationships. Molecules 2018, 23, 2917. [Google Scholar] [CrossRef] [PubMed]
- Abdullah; Shahzadi, I.; Mehmood, F.; Ali, Z.; Malik, M.S.; Waseem, S.; Mirza, B.; Ahmed, I.; Waheed, M.T. Comparative analyses of chloroplast genomes among three Firmiana species: Identification of mutational hotspots and phylogenetic relationship with other species of Malvaceae. Plant Gene 2019, 19, 100199. [Google Scholar] [CrossRef]
- Abdullah; Mehmood, F.; Shahzadi, I.; Waseem, S.; Mirza, B.; Ahmed, I.; Waheed, M.T. Chloroplast genome of Hibiscus rosa-sinensis (Malvaceae): Comparative analyses and identification of mutational hotspots. Genomics 2019, 1. [Google Scholar] [CrossRef]
- Menezes, A.P.A.; Resende-Moreira, L.C.; Buzatti, R.S.O.; Nazareno, A.G.; Carlsen, M.; Lobo, F.P.; Kalapothakis, E.; Lovato, M.B. Chloroplast genomes of Byrsonima species (Malpighiaceae): Comparative analysis and screening of high divergence sequences. Sci. Rep. 2018, 8, 1–12. [Google Scholar] [CrossRef]
- Ahmed, I.; Biggs, P.J.; Matthews, P.J.; Collins, L.J.; Hendy, M.D.; Lockhart, P.J. Mutational dynamics of aroid chloroplast genomes. Genome Biol. Evol. 2012, 4, 1316–1323. [Google Scholar] [CrossRef]
- Amiryousefi, A.; Hyvönen, J.; Poczai, P. The chloroplast genome sequence of bittersweet (Solanum dulcamara): Plastid genome structure evolution in Solanaceae. PLoS ONE 2018, 13, e196069. [Google Scholar] [CrossRef]
- Yu, X.; Zuo, L.; Lu, D.; Lu, B.; Yang, M.; Wang, J. Comparative analysis of chloroplast genomes of five Robinia species: Genome comparative and evolution analysis. Gene 2019, 689, 141–151. [Google Scholar] [CrossRef]
- Yu, X.Q.; Drew, B.T.; Yang, J.B.; Gao, L.M.; Li, D.Z. Comparative chloroplast genomes of eleven Schima (Theaceae) species: Insights into DNA barcoding and phylogeny. PLoS ONE 2017, 12, e178026. [Google Scholar] [CrossRef]
- He, L.; Qian, J.; Li, X.; Sun, Z.; Xu, X.; Chen, S.; McPhee, D.J. Complete chloroplast genome of medicinal plant Lonicera japonica: Genome rearrangement, intron gain and loss, and implications for phylogenetic studies. Molecules 2017, 22, 249. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Wang, Y.; He, P.; Li, P.; Lee, J.; Soltis, D.E.; Fu, C. Chloroplast genome analyses and genomic resource development for epilithic sister genera Oresitrophe and Mukdenia (Saxifragaceae), using genome skimming data. BMC Genom. 2018, 19, 235. [Google Scholar] [CrossRef] [PubMed]
- Shahzadi, I.; Abdullah; Mehmood, F.; Ali, Z.; Ahmed, I.; Mirza, B. Chloroplast genome sequences of Artemisia maritima and Artemisia absinthium: Comparative analyses, mutational hotspots in genus Artemisia and phylogeny in family Asteraceae. Genomics 2019. [Google Scholar] [CrossRef]
- Yang, Z.; Huang, Y.; An, W.; Zheng, X.; Huang, S.; Liang, L. Sequencing and structural analysis of the complete chloroplast genome of the medicinal plant Lycium Chinense Mill. Plants 2019, 8, 87. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Yang, Z.; Huang, S.; An, W.; Li, J.; Zheng, X. Comprehensive analysis of Rhodomyrtus tomentosa chloroplast genome. Plants 2019, 8, 89. [Google Scholar] [CrossRef] [PubMed]
- Cui, Y.; Nie, L.; Sun, W.; Xu, Z.; Wang, Y.; Yu, J.; Song, J.; Yao, H. Comparative and phylogenetic analyses of ginger (Zingiber officinale) in the family Zingiberaceae based on the complete chloroplast genome. Plants 2019, 8, 283. [Google Scholar] [CrossRef]
- Poczai, P.; Hyvönen, J. The complete chloroplast genome sequence of the CAM epiphyte Spanish moss (Tillandsia usneoides, Bromeliaceae) and its comparative analysis. PLoS ONE 2017, 12, e187199. [Google Scholar] [CrossRef]
- Keller, J.; Rousseau-Gueutin, M.; Martin, G.E.; Morice, J.; Boutte, J.; Coissac, E.; Ourari, M.; Aïnouche, M.; Salmon, A.; Cabello-Hurtado, F.; et al. The evolutionary fate of the chloroplast and nuclear rps16 genes as revealed through the sequencing and comparative analyses of four novel legume chloroplast genomes from Lupinus. DNA Res. 2017, 24, 343–358. [Google Scholar] [CrossRef]
- McDonald, M.J.; Wang, W.C.; Huang, H.D.; Leu, J.Y. Clusters of nucleotide substitutions and insertion/deletion mutations are associated with repeat sequences. PLoS Biol. 2011, 9. [Google Scholar] [CrossRef]
- Cheon, K.-S.; Kim, K.-A.; Kwak, M.; Lee, B.; Yoo, K.-O. The complete chloroplast genome sequences of four Viola species (Violaceae) and comparative analyses with its congeneric species. PLoS ONE 2019, 14, e0214162. [Google Scholar] [CrossRef]
- Liu, E.; Yang, C.; Liu, J.; Jin, S.; Harijati, N.; Hu, Z.; Diao, Y.; Zhao, L. Comparative analysis of complete chloroplast genome sequences of four major Amorphophallus species. Sci. Rep. 2019, 9, 809. [Google Scholar] [CrossRef]
- Cai, J.; Ma, P.F.; Li, H.T.; Li, D.Z. Complete plastid genome sequencing of four Tilia species (Malvaceae): A comparative analysis and phylogenetic implications. PLoS ONE 2015, 10, e142705. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Zheng, S.; Liu, Y.; Han, J. ITS2, a Better DNA Barcode than ITS in Identification of Species in Artemisia L. Chin. Herb. Med. 2016, 8, 352–358. [Google Scholar] [CrossRef]
- Liu, G.; Ning, H.; Ayidaerhan, N.; Aisa, H.A. Evaluation of DNA barcode candidates for the discrimination of Artemisia L. Mitochondrial DNA Part A 2016, 1394. [Google Scholar] [CrossRef]
- Li, X.; Yang, Y.; Henry, R.J.; Rossetto, M.; Wang, Y.; Chen, S. Plant DNA barcoding: From gene to genome. Biol. Rev. 2014. [Google Scholar] [CrossRef]
- Meng, D.; Xiaomei, Z.; Wenzhen, K.; Id, Z.X. Detecting useful genetic markers and reconstructing the phylogeny of an important medicinal resource plant, Artemisia selengensis, based on chloroplast genomics. PLoS ONE 2019, 14, 1–19. [Google Scholar] [CrossRef]
- Zerbino, D.R.; Birney, E. Velvet: Algorithms for de novo short read assembly using de Bruijn graphs. Genome Res. 2008, 18, 821–829. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Durbin, R. Fast and accurate short read alignment with Burrows-Wheeler transform. Bioinformatics 2009, 25, 1754–1760. [Google Scholar] [CrossRef] [Green Version]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious Basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef]
- Tillich, M.; Lehwark, P.; Pellizzer, T.; Ulbricht-Jones, E.S.; Fischer, A.; Bock, R.; Greiner, S. GeSeq—Versatile and accurate annotation of organelle genomes. Nucleic Acids Res. 2017, 45, W6–W11. [Google Scholar] [CrossRef]
- Lowe, T.M.; Chan, P.P. tRNAscan-SE On-line: Integrating search and context for analysis of transfer RNA genes. Nucleic Acids Res. 2016, 44, W54–W57. [Google Scholar] [CrossRef]
- Laslett, D.; Canback, B. ARAGORN, a program to detect tRNA genes and tmRNA genes in nucleotide sequences. Nucleic Acids Res. 2004, 32, 11–16. [Google Scholar] [CrossRef] [PubMed]
- Lohse, M.; Drechsel, O.; Bock, R. OrganellarGenomeDRAW (OGDRAW): A tool for the easy generation of high-quality custom graphical maps of plastid and mitochondrial genomes. Curr. Genet. 2007, 52, 267–274. [Google Scholar] [CrossRef] [PubMed]
- Lehwark, P.; Greiner, S. GB2sequin—A file converter preparing custom GenBank files for database submission. Genomics 2018. [Google Scholar] [CrossRef] [PubMed]
- Amiryousefi, A.; Hyvönen, J.; Poczai, P. IRscope: An online program to visualize the junction sites of chloroplast genomes. Bioinformatics 2018, 34, 3030–3031. [Google Scholar] [CrossRef]
- Mower, J.P. The PREP suite: Predictive RNA editors for plant mitochondrial genes, chloroplast genes and user-defined alignments. Nucleic Acids Res. 2009, 37, W253–W259. [Google Scholar] [CrossRef]
- Beier, S.; Thiel, T.; Münch, T.; Scholz, U.; Mascher, M. MISA-web: A web server for microsatellite prediction. Bioinformatics 2017, 33, 2583–2585. [Google Scholar] [CrossRef]
- Kurtz, S.; Choudhuri, J.V.; Ohlebusch, E.; Schleiermacher, C.; Stoye, J.; Giegerich, R. REPuter: The manifold applications of repeat analysis on a genomic scale. Nucleic Acids Res. 2001, 29, 4633–4642. [Google Scholar] [CrossRef]
- Katoh, K.; Kuma, K.I.; Toh, H.; Miyata, T. MAFFT version 5: Improvement in accuracy of multiple sequence alignment. Nucleic Acids Res. 2005, 33, 511–518. [Google Scholar] [CrossRef]
- Rozas, J.; Ferrer-Mata, A.; Sanchez-DelBarrio, J.C.; Guirao-Rico, S.; Librado, P.; Ramos-Onsins, S.E.; Sanchez-Gracia, A. DnaSP 6: DNA sequence polymorphism analysis of large data sets. Mol. Biol. Evol. 2017, 34, 3299–3302. [Google Scholar] [CrossRef]
- Nguyen, L.-T.; Schmidt, H.A.; von Haeseler, A.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating Maximum-likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef]
- Hoang, D.T.; Chernomor, O.; von Haeseler, A.; Minh, B.Q.; Vinh, L.S. UFBoot2: Improving the ultrafast bootstrap approximation. Mol. Biol. Evol. 2018, 35, 518–522. [Google Scholar] [CrossRef] [PubMed]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.F.; von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast model selection for accurate phylogenetic estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef] [PubMed]
- Dereeper, A.; Guignon, V.; Blanc, G.; Audic, S.; Buffet, S.; Chevenet, F.; Dufayard, J.-F.; Guindon, S.; Lefort, V.; Lescot, M.; et al. Phylogeny.fr: Robust phylogenetic analysis for the non-specialist. Nucleic Acids Res. 2008, 36, W465–W469. [Google Scholar] [CrossRef] [PubMed]
- Chevenet, F.; Brun, C.; Bañuls, A.-L.; Jacq, B.; Christen, R. TreeDyn: Towards dynamic graphics and annotations for analyses of trees. BMC Bioinform. 2006, 7, 439. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Circular map of the Artemisia scoparia chloroplast genome. The genes present outside of the circle are transcribed anti-clockwise, while those inside the circle are transcribed clockwise. Large single copy (LSC), inverted repeat (IRa, IRb), and small single copy (SSC) regions are indicated. The dashed grey color of the inner circle shows the GC content, whereas the lighter gray color shows the AT content. Color of genes was assigned based on their functions.
Figure 1.
Circular map of the Artemisia scoparia chloroplast genome. The genes present outside of the circle are transcribed anti-clockwise, while those inside the circle are transcribed clockwise. Large single copy (LSC), inverted repeat (IRa, IRb), and small single copy (SSC) regions are indicated. The dashed grey color of the inner circle shows the GC content, whereas the lighter gray color shows the AT content. Color of genes was assigned based on their functions.

Figure 2.
Details of the contraction and expansion of inverted repeats at junction sites. For all plant species, positive strand genes are represented at the top, from right to left, on the corresponding track, whereas negative strand genes are illustrated on the lower side of the track, from left to right. Arrows depict the distance between the start and end of a gene from the junction site. Scale bar present above or below the genes extending from one region to another illustrates the number of base pairs to which genes join in that region. JSA (SSC/IRa), JSB (IRb/SSC), JLA (IRa/LSC), and JLB (IRb/LSC) indicate the junction sites between two corresponding regions of a genome. The plotted genes and distances surrounding the junction site are a scaled projection of the genome.
Figure 2.
Details of the contraction and expansion of inverted repeats at junction sites. For all plant species, positive strand genes are represented at the top, from right to left, on the corresponding track, whereas negative strand genes are illustrated on the lower side of the track, from left to right. Arrows depict the distance between the start and end of a gene from the junction site. Scale bar present above or below the genes extending from one region to another illustrates the number of base pairs to which genes join in that region. JSA (SSC/IRa), JSB (IRb/SSC), JLA (IRa/LSC), and JLB (IRb/LSC) indicate the junction sites between two corresponding regions of a genome. The plotted genes and distances surrounding the junction site are a scaled projection of the genome.

Figure 3.
The x-axis shows the regions of the chloroplast genome and the y-axis shows the nucleotide diversity of each region. The regions with 0 nucleotide diversity are not included in the list.
Figure 3.
The x-axis shows the regions of the chloroplast genome and the y-axis shows the nucleotide diversity of each region. The regions with 0 nucleotide diversity are not included in the list.

Figure 4.
Phylogenetic relationships among 30 species of the family Asteraceae. The number on each node represents the bootstrapping value. The bootstrapping values equal to 100 were not represented on the nodes and omitted from the tree for quality visualization.
Figure 4.
Phylogenetic relationships among 30 species of the family Asteraceae. The number on each node represents the bootstrapping value. The bootstrapping values equal to 100 were not represented on the nodes and omitted from the tree for quality visualization.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Genomic features of Artemisia scoparia.
| Characteristics | Artemisia scoparia | |
|---|---|---|
| Size (Base Pair; bp) | 151,060 | |
| IR length (bp) | 24,972 | |
| LSC length (bp) | 82,834 | |
| SSC length (bp) | 18,282 | |
| Number of genes | 114 | |
| Duplicate genes | 18 | |
| tRNA genes | 30 | |
| rRNA genes | 4 | |
| Protein-coding genes | 80 | |
| GC content | Total (%) | 37.5 |
| CDS (%) | 37.8 | |
| IR (%) | 43.1 | |
| LSC (%) | 35.6 | |
| SSC (%) | 30.8 | |
| rRNA (%) | 55 | |
| tRNA (%) | 52.7 | |
| All gene % | 39.3 | |
Table 2.
Microsatellites in Artemisia scoparia.
| Repeats | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 15 | 16 | 17 | 18 | 19 | 20 | Total |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| A | - | - | - | - | - | - | - | 10 | 2 | 5 | 17 | ||||||||
| T | - | - | - | - | - | - | - | 11 | 3 | 1 | 3 | 2 | 1 | 1 | 1 | 23 | |||
| AT | - | - | 3 | 2 | 1 | 6 | |||||||||||||
| TA | - | - | 2 | 1 | 3 | ||||||||||||||
| ATA | - | 1 | 1 | ||||||||||||||||
| ATT | - | 1 | 1 | ||||||||||||||||
| TAA | - | 1 | 1 | ||||||||||||||||
| TTA | - | 1 | 1 | ||||||||||||||||
| TTC | - | 1 | 1 | ||||||||||||||||
| AATA | 2 | 2 | |||||||||||||||||
| AATC | 2 | 2 | |||||||||||||||||
| AATT | 1 | 1 | |||||||||||||||||
| ATTT | 1 | 1 | |||||||||||||||||
| CAAT | 1 | 1 | |||||||||||||||||
| TAAT | 1 | 1 | |||||||||||||||||
| TATT | 1 | 1 | |||||||||||||||||
| TTTA | 1 | 1 | |||||||||||||||||
| TTTC | 1 | 1 | |||||||||||||||||
| AAATT | 1 | 1 |
Table 3.
Oligonucleotide repeats in Artemisia scoparia.
| S.No | Type | Size | Regions | IGS/CDS/Intron | 1st Position | 2nd Position | Sequence |
|---|---|---|---|---|---|---|---|
| 1 | F | 32 | LSC | psaB/psaA | 37,977 | 40,201 | AGAAAAATAAATGCAATAGCTAAATGATGATG |
| 2 | F | 30 | LSC | psaB/psaA | 37,988 | 40,212 | TGCAATAGCTAAATGATGATGAGCCATATC |
| 3 | R | 30 | LSC | psaA-ycf3/rps4-trnT | 41,744 | 45,790 | ATAAAAAAAAAAAAGATATATATCTAATAT |
| 4 | F | 41 | LSC/IR | ycf3 intron/rps12-ycf15 | 42,974 | 96,836 | TACAGAACCGTACATGAGATTTTCATCTCATACGGCTCCTC |
| 5 | F | 39 | LSC/SSC | ycf3 intron/ndhA intron | 42,976 | 118,122 | CAGAACCGTACATGAGATTTTCATCTCATACGGCTCCTC |
| 6 | F | 35 | LSC/IR | ycf3 intron/ndhB intron | 42,979 | 93,787 | AACCGTACATGAGATTTTCATCTCATACGGCTCCT |
| 7 | F | 30 | LSC | rbcL*-rbcL-accD | 56,187 | 56,211 | AAAAGAGATAAGGTTCGTTCTCTTAAAAGA |
| 8 | F | 30 | LSC/IR | psaJ-rpl33/ycf15-trnV | 66,253 | 97,999 | TAAGAGGATAGCAAGTTACAAATTCTATTT |
| 9 | P | 48 | LSC | psbT-psbN | 72,973 | 72,973 | AATTGAAGTAATGAGCCTCCCAATATTGGGAGGCTCATTACTTCAATT |
| 10 | P | 31 | IR | ycf2 | 86,489 | 86,489 | CGAGAAGCAGATGATTAATCATCTGCTTCCG |
| 11 | F | 60 | IR | ycf2 | 90,062 | 90,080 | CGATATTGATGAGATTGACGATATTGATGCTAGTGACGATATTGATGCTAGTGACGATAT |
| 12 | F | 39 | IR/SSC | rps12-trnV/ndhA intron | 96,838 | 118,122 | CAGAACCGTACATGAGATTTTCACCTCATACGGCTCCTC |
| 13 | F | 34 | IR | rrn4.5-rrn5 | 105,729 | 105,761 | CATTGTTCAACTCTTTAACAACATGAAAAAACCATTGTT |
| 14 | P | 43 | SSC | ndhD-psaC | 115,012 | 115,012 | AAAACATGTGCCCAAAAATAAGATATTTTTGGGCACATGTTTT |
* represents copy of repeat shared between genic and intergenic spacer regions.
Table 4.
Substitutions and InDels events in the chloroplast genome of Artemisia scoparia.
| Substitution Type | LSC | SSC | IR |
|---|---|---|---|
| A/C | 15 | 5 | 1 |
| A/G | 12 | 2 | 2 |
| G/T | 16 | 11 | 0 |
| C/T | 9 | 1 | 3 |
| C/G | 5 | 2 | 0 |
| A/T | 2 | 2 | 0 |
| Total | 59 | 23 | 6 |
| No’s of InDels | 42 | 9 | 3 |
| Average size | 3.86 | 3.11 | 3.67 |
Table 5.
Mutational hotspots in the genus Artemisia.
| Serial No | Region | Nucleotide Diversity | Total No’s of Mutations | Region Length |
|---|---|---|---|---|
| 1 | ccsA-ndhD | 0.054545 | 12 | 220 |
| 2 | trnH-psbA | 0.047382 | 19 | 401 |
| 3 | ndhG-ndhI | 0.042553 | 16 | 376 |
| 4 | rps18-rpl20 | 0.042403 | 12 | 283 |
| 5 | rps15-ycf1 | 0.042056 | 18 | 428 |
| 6 | trnC-GCA-petN | 0.038917 | 23 | 591 |
| 7 | rpl32-trnL | 0.037443 | 39 | 908 |
| 8 | rpoC2-rps2 | 0.035433 | 9 | 254 |
| 9 | trnK-UUU-rps16 | 0.035267 | 31 | 879 |
| 10 | ndhF-rpl32 | 0.033915 | 35 | 1032 |
| 11 | psbA-trnK-UUU | 0.033654 | 7 | 208 |
| 12 | ndhC-trnV | 0.033646 | 43 | 1278 |
| 13 | trnS-GGA-rps4 | 0.032934 | 11 | 334 |
| 14 | petN-psbM | 0.032847 | 18 | 548 |
| 15 | rps16-trnQ-UUG | 0.031729 | 29 | 914 |
| 16 | trnS-GCU-trnC-GCA | 0.031291 | 24 | 767 |
| 17 | accD-psaI | 0.03071 | 16 | 521 |
| 18 | psbK-psbI | 0.030534 | 12 | 393 |
| 19 | rbcL-accD | 0.030019 | 16 | 533 |
| 20 | ycf4-cemA | 0.029091 | 8 | 275 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Iram, S.; Hayat, M.Q.; Tahir, M.; Gul, A.; Abdullah; Ahmed, I. Chloroplast Genome Sequence of Artemisia scoparia: Comparative Analyses and Screening of Mutational Hotspots. Plants 2019, 8, 476. https://doi.org/10.3390/plants8110476
AMA Style
Iram S, Hayat MQ, Tahir M, Gul A, Abdullah, Ahmed I. Chloroplast Genome Sequence of Artemisia scoparia: Comparative Analyses and Screening of Mutational Hotspots. Plants. 2019; 8(11):476. https://doi.org/10.3390/plants8110476
Chicago/Turabian StyleIram, Shabina, Muhammad Qasim Hayat, Muhammad Tahir, Alvina Gul, Abdullah, and Ibrar Ahmed. 2019. "Chloroplast Genome Sequence of Artemisia scoparia: Comparative Analyses and Screening of Mutational Hotspots" Plants 8, no. 11: 476. https://doi.org/10.3390/plants8110476
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.