The Changbai Alpine Shrub Tundra Will Be Replaced by Herbaceous Tundra under Global Climate Change

 , ,
, ,
Abstract
:1. Introduction
2. Results
2.1. Statistical Characteristics of the Dominant Species
2.2. Spatial Distribution of the Dominant Species
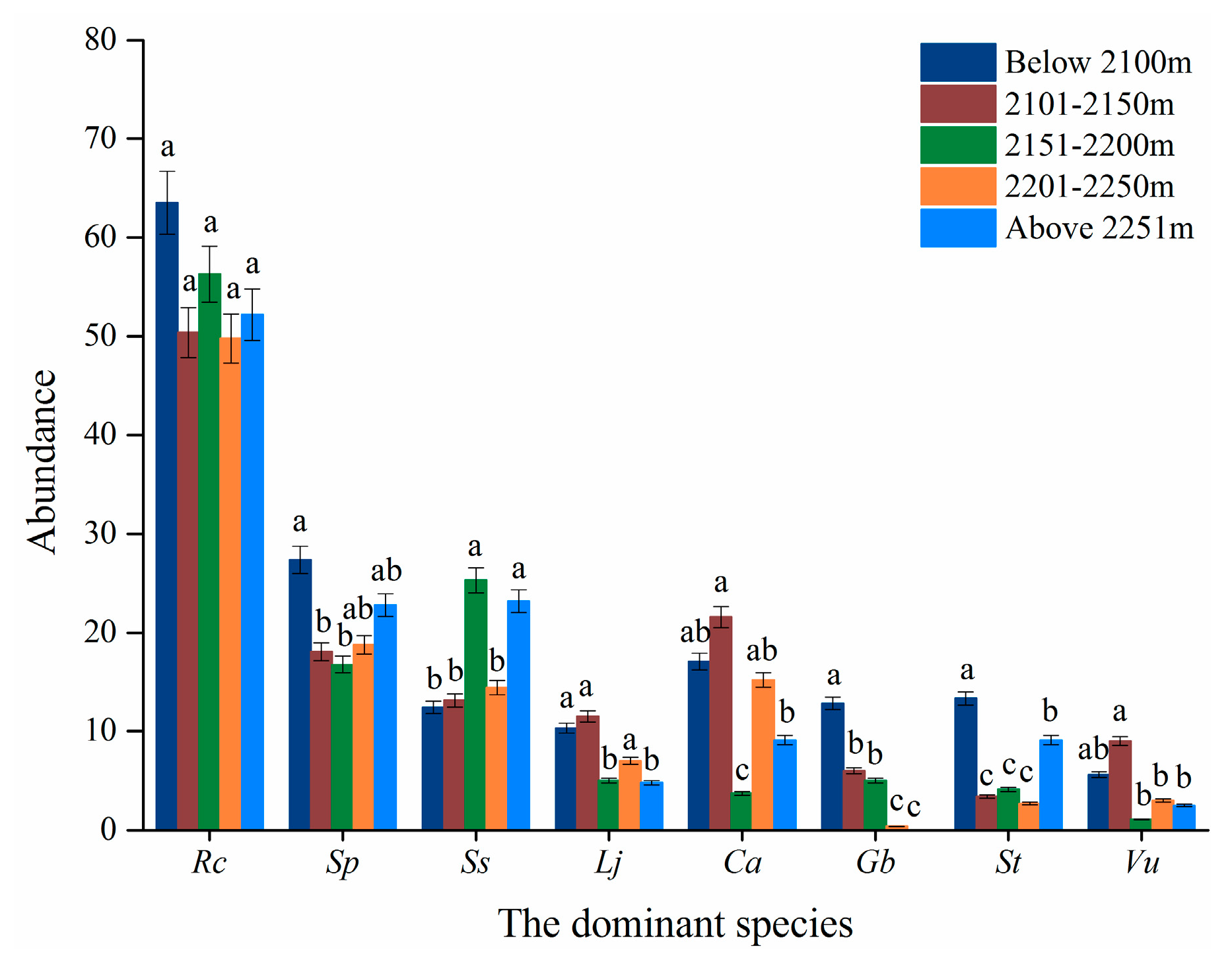
2.2.1. Altitudinal Factor
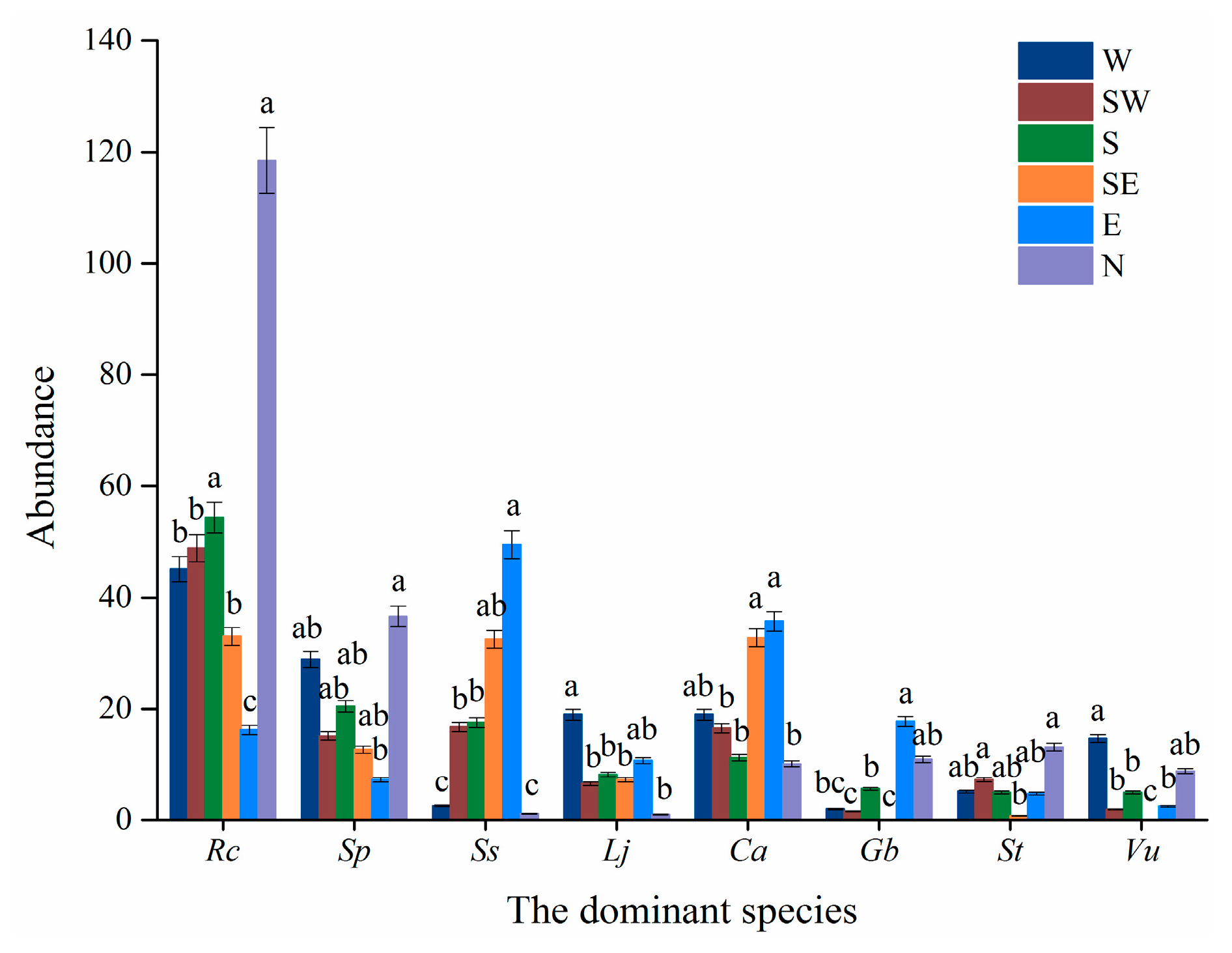
2.2.2. Slope Aspects
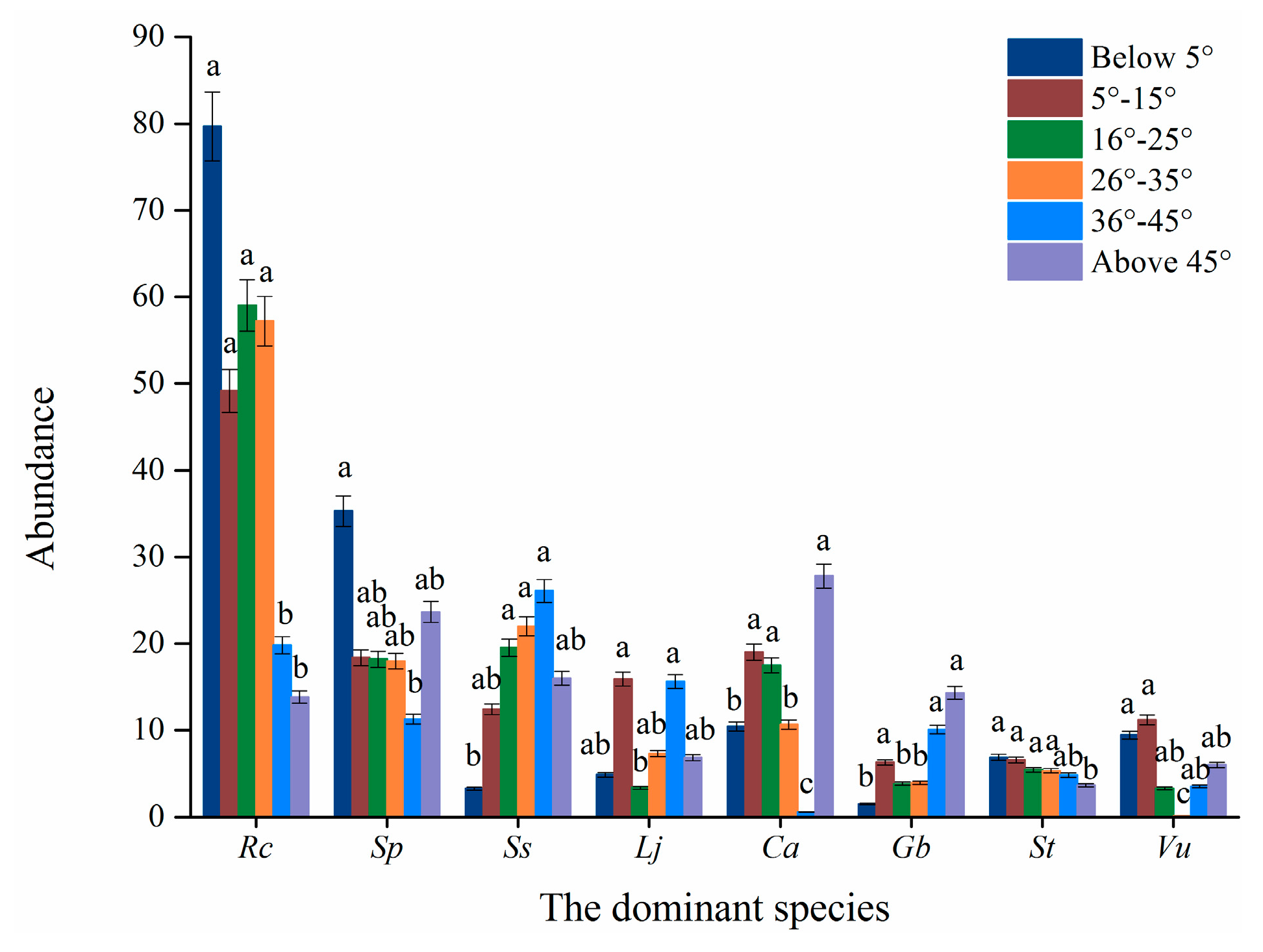
2.2.3. Slope Grade
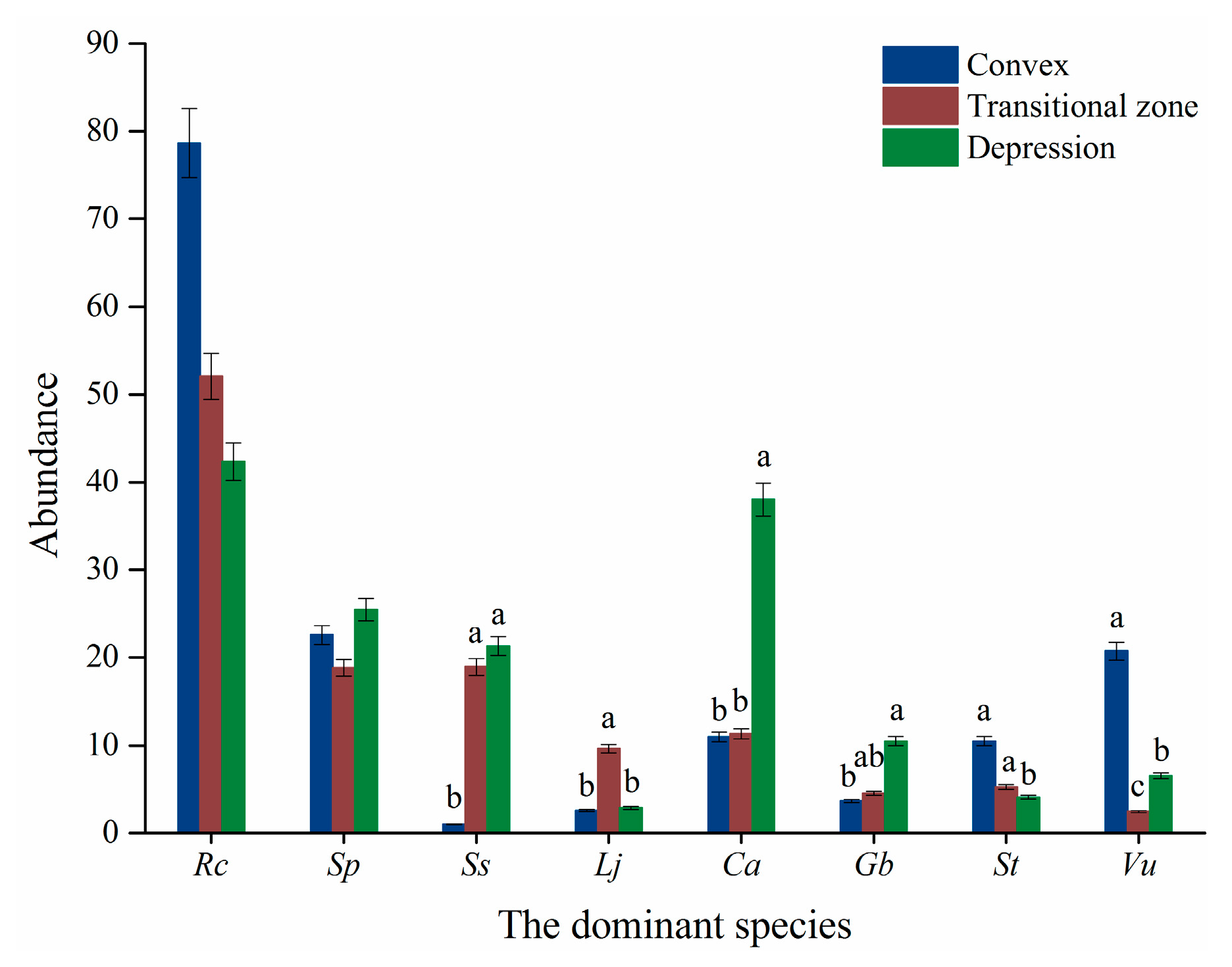
2.2.4. Micro-Topography
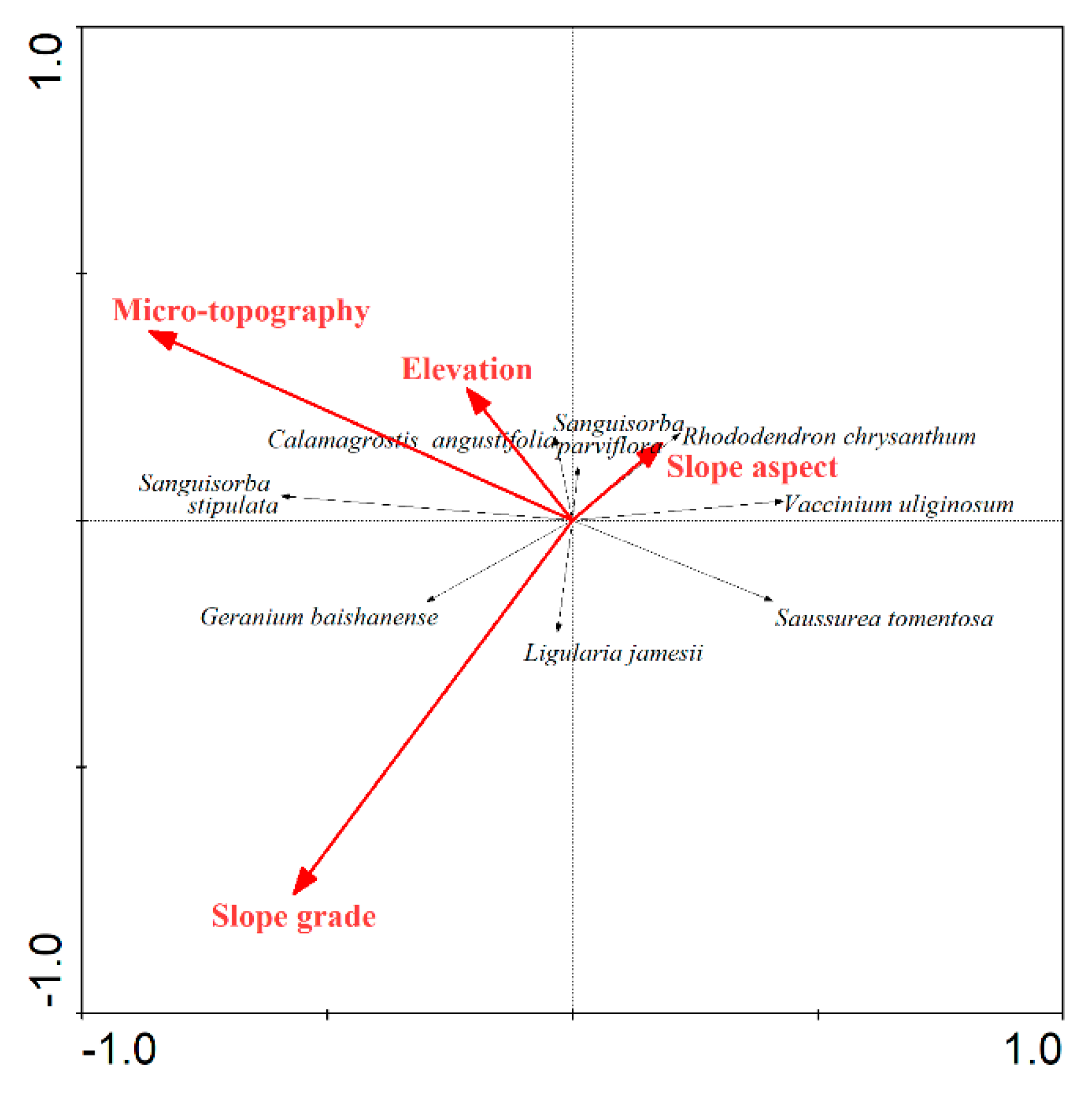
2.3. Correlation Analysis of Dominant Species and Altitudinal and Topographic Factors
3. Discussion
3.1. Expansion of Herbaceous Plants
3.2. Implication for Possible Causes of Changes in the Tundra Vegetation
4. Materials and Methods
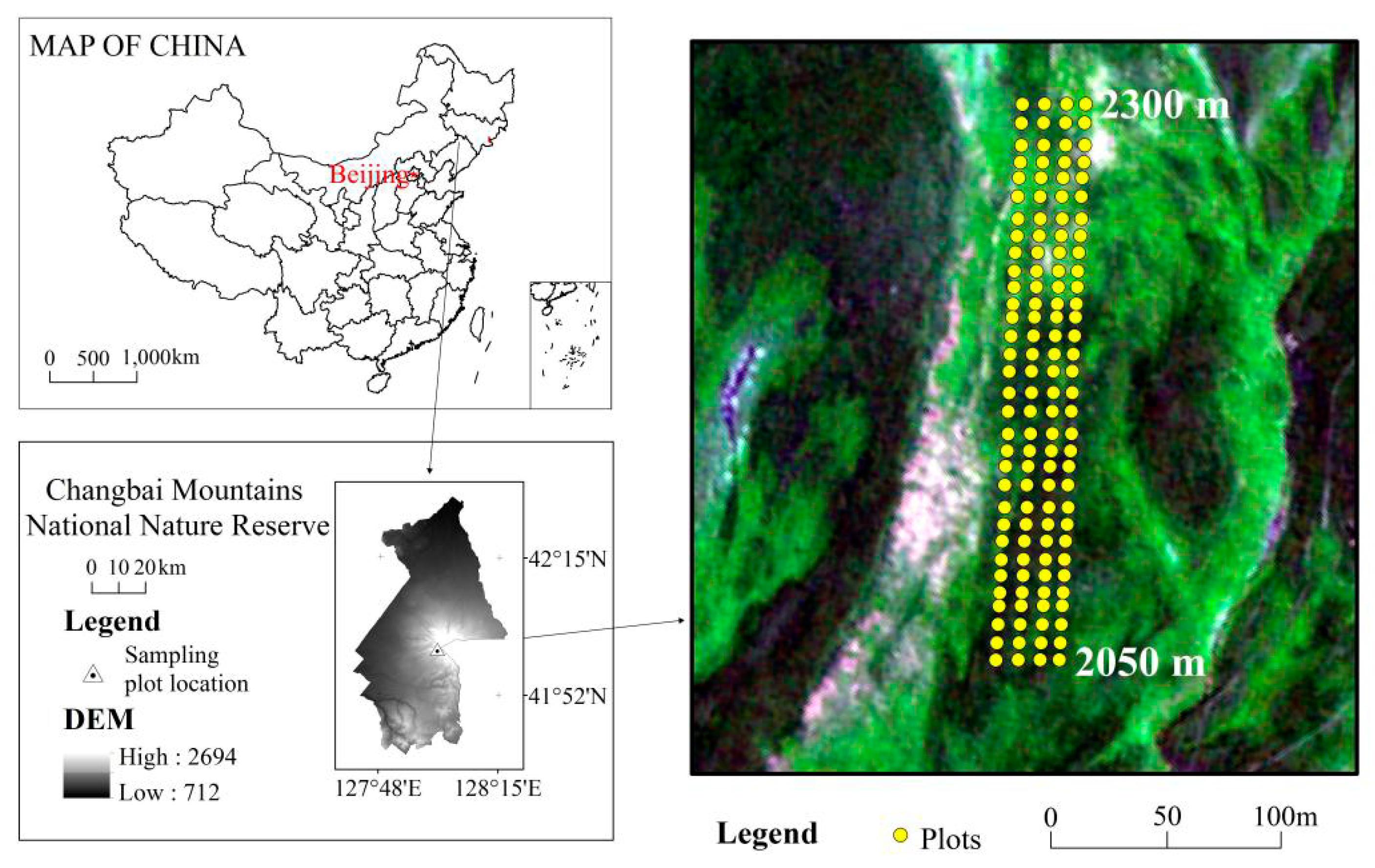
4.1. Study Area
4.2. Field Surveys
4.3. Data Processing
4.3.1. Determination of Dominant Species
4.3.2. Descriptive Statistics of Dominant Species
4.3.3. The Shannon-Wiener Index (SW)
4.3.4. One-way Analysis of Variance (ANOVA) and Generalized Linear Models (GLMs)
4.3.5. Redundancy Analysis (RDA)
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Körner, C. Alpine Plant Life: Functional Plant Ecology of High Mountain Ecosystems; Springer: Berlin/Heidelberg, Germany, 2003; pp. 21–30. [Google Scholar]
- Wei, J.; Jiang, P.; Yu, D.Y.; Wu, G.; Fu, H.W. Distribution patterns of vegetation biomass and nutrients biocycle in alpine tundra ecosystem on Changbai Mountains, Northeast China. J. For. Res. 2007, 18, 271–278. [Google Scholar] [CrossRef]
- Mavris, C.; Furrer, G.; Dahms, D.; Anderson, S.P.; Blum, A.; Goetze, J.; Wells, A.; Egli, M. Decoding potential effects of climate and vegetation change on mineral weathering in alpine soils: An experimental study in the Wind River Range (Wyoming, USA). Geoderma 2015, 255, 12–26. [Google Scholar] [CrossRef]
- Seo, J.; Jang, I.; Jung, J.Y.; Lee, Y.K.; Kang, H. Warming and increased precipitation enhance phenol oxidase activity in soil while warming induces drought stress in vegetation of an Arctic ecosystem. Geoderma 2015, 259, 347–353. [Google Scholar] [CrossRef]
- Lesica, P.; McCune, B. Decline of arctic-alpine plants at the southern margin of their range following a decade of climatic warming. J. Veg. Sci. 2004, 15, 679–690. [Google Scholar] [CrossRef]
- Huelber, K.; Gottfried, M.; Pauli, H.; Reiter, K.; Winkler, M.; Grabherr, G. Phenological responses of snowbed species to snow removal dates in the Central Alps: Implications for climate warming. Arct. Antarct. Alp. Res. 2006, 38, 99–103. [Google Scholar] [CrossRef]
- Diaz, H.F.; Eischeid, J.K. Disappearing “alpine tundra” Koppen climatic type in the western United States. Geophys. Res. Lett. 2007, 34, 1–4. [Google Scholar] [CrossRef]
- Inouye, D.W. Effects of climate change on phenology, frost damage, and floral abundance of montane wildflowers. Ecology 2008, 89, 353–362. [Google Scholar] [CrossRef] [PubMed]
- Wilson, S.D.; Nilsson, C. Arctic alpine vegetation change over 20 years. Glob. Chang. Biol. 2009, 15, 1676–1684. [Google Scholar] [CrossRef] [Green Version]
- Ackerly, D.D.; Loarie, S.R.; Cornwell, W.K.; Weiss, S.B.; Hamilton, H.; Branciforte, R.; Kraft, N.J.B. The geography of climate change: Implications for conservation biogeography. Divers. Distrib. 2010, 16, 476–487. [Google Scholar] [CrossRef]
- Klanderud, K.; Birks, H.J.B. Recent increases in species richness and shifts in altitudinal distributions of Norwegian mountain plants. Holocene 2016, 13, 1–6. [Google Scholar] [CrossRef]
- McDougall, K.L.; Morgan, J.W.; Walsh, N.G.; Williams, R.J. Plant invasions in treeless vegetation of the Australian Alps. Perspect. Plant Ecol. Evol. Syst. 2005, 7, 159–171. [Google Scholar] [CrossRef]
- Beckage, B.; Osborne, B.; Gavin, D.G.; Pucko, C.; Siccama, T.; Perkins, T. A rapid upward shift of a forest ecotone during 40 years of warming in the green mountains of Vermont. Proc. Natl. Acad. Sci. USA 2008, 105, 4197–4202. [Google Scholar] [CrossRef]
- Kelly, A.E.; Goulden, M.L. Rapid shifts in plant distribution with recent climate change. Proc. Natl. Acad. Sci. USA 2008, 105, 11823–11826. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parolo, G.; Rossi, G. Upward migration of vascular plants following a climate warming trend in the Alps. Basic Appl. Ecol. 2008, 9, 100–107. [Google Scholar] [CrossRef]
- Odland, A.; Hoitomt, T.; Olsen, S.L. Increasing vascular plant richness on 13 high mountain summits in Southern Norway since the early 1970s. Arct. Antarct. Alp. Res. 2010, 42, 458–470. [Google Scholar] [CrossRef]
- Walther, G.R.; Beißner, S.; Burga, C.A. Trends in the upward shift of Alpine plants. J. Veg. Sci. 2010, 16, 541–548. [Google Scholar] [CrossRef]
- Pauli, H.; Gottfried, M.; Reiter, K.; Klettner, C.; Grabherr, G. Signals of range expansions and contractions of vascular plants in the high Alps: Observations (1994–2004) at the Gloria master site Schrankogel, Tyrol, Austria. Glob. Chang. Biol. 2007, 13, 147–156. [Google Scholar] [CrossRef]
- Hughes, I.I. Biological consequences of global warming: Is the signal already apparent? Trends Ecol. Evol. 2010, 15, 56–61. [Google Scholar] [CrossRef]
- Walther, G.-R. Plants in a warmer world. Perspect. Plant. Ecol. Evol. Syst. 2003, 6, 169–185. [Google Scholar] [CrossRef]
- Thuiller, W.; Richardson, D.M.; Midgley, G.F. Will climate change promote alien plant invasions? Biol. Invasions 2006, 193, 197–211. [Google Scholar]
- Smith, J.G.; Sconiers, W.; Spasojevic, M.J.; Ashton, I.W.; Suding, K.N. Phenological changes in alpine plants in response to increased snowpack, temperature, and nitrogen. Arct. Antarct. Alp. Res. 2018, 44, 135–142. [Google Scholar] [CrossRef]
- Huang, X.C.; Li, C.G. An analysis on the ecology of alpine tundra landscape of Changbai Mountains. Sci. Geogr. Sin. 1984, 39, 285–297. [Google Scholar]
- Ni, J.; Zhang, X.S. Climate variability, ecological gradient and the Northeast China Transect (NECT). J. Arid Environ. 2000, 46, 313–325. [Google Scholar] [CrossRef] [Green Version]
- Zhou, X.F.; Wang, X.C.; Han, S.J.; Zou, C.J. The effect of global climate change on the dynamics of Betulaermanii—Tundra ecotone in the Changbai mountains. Earth Sci. Front. 2002, 9, 227–231. [Google Scholar]
- Wang, J.; Guan, D.; Jin, C.; Wang, A.; Wu, J.; Yuan, F. Non-uniformity of precipitation distribution in Changbai Mountains in growth season. Chin. J. Ecol. 2011, 30, 131–137. [Google Scholar]
- Zong, S.W.; Wu, Z.F.; Du, H.B. Study on Climate Change in Alpine Tundra of the Changbai Mountain in Growing Season in Recent 52 Years. Arid Zone Res. 2013, 30, 41–49. [Google Scholar]
- Wang, X.D.; Liu, H.Q. The Dynamics Response of Betulaermanii Population and Climate Change on Different Slopes Aspect of North Slope, Changbai Mountains. Sci. Geogr. Sin. 2012, 32, 199–206. [Google Scholar]
- Hu, N.F.; Wang, A.Z.; Guan, D.X.; Yuan, F.H.; Jin, C.J.; Wu, J.B.; Wang, J.J. Multiple time scale analysis of precipitation series in Changbai Mountain Region in 1959–2006. Chin. J. Appl. Ecol. 2010, 21, 549–556. [Google Scholar]
- Shi, Y.; Gao, X.J.; Wu, J.; Giorgi, F.; Dong, W.J. Simulation of the Changes in Snow Cover over China under Global Warming by a High Resolution RCM. J. Glaciol. Geocryol. 2010, 32, 215–222. [Google Scholar]
- Qian, J.J.; Zhang, W.Z. A brief report on the research of the Changbaishan alpine tundra vegetation. J. Northeast. Normal Univ. 1980, 1, 51–67. [Google Scholar]
- Qian, H. Numerical classification and ordination of plant communities in Mt. Changbai. J. Appl. Ecol. 1990, 1, 254–263. [Google Scholar]
- Zong, S.W.; Xu, J.W.; Wu, Z.F. Investigation and Mechanism Analysis on the Invasion of Deyeuxia angustifolia to Tundra Zone in Western Slope of Changbai Mountain. J. Mt. Sci. 2013, 31, 448–455. [Google Scholar]
- Jiawei, X.; Feihu, Z. Several main questions of physical geography research of Changbai Mountains. In Geosystems and Ecological Security of the Changbai Mountainss: IV. Changchun; Northeast Normal University Press: Beijing, China, 2010; pp. 266–274. [Google Scholar]
- Jin, Y.H.; Xu, J.W.; Liang, Y.; Zong, S.W. Effects of Volcanic Interference on the Vegetation Distribution of Changbai Mountain. Sci. Geogr. Sin. 2013, 33, 203–208. [Google Scholar]
- Jin, Y.H.; Xu, J.W.; Liu, L.N.; Tao, Y.; He, H.S.; Zong, S.W.; Du, H.B.; Wang, L.; Zhu, R.S. Spatial distribution pattern and associations of dominant plant species in the alpine tundra of the Changbai Mountains. Sci. Geogr. Sin. 2016, 36, 1212–1218. [Google Scholar]
- Jin, Y.H.; Xu, J.W.; Wang, Y.Q.; Wang, S.X.; Chen, Z.S.; Huang, X.T.; Niu, L.J. Effects of Nitrogen Deposition on Tundra Vegetation Undergoing Invasion by Deyeuxia angustifolia in Changbai Mountains. Chin. Geogr. Sci. 2016, 26, 99–108. [Google Scholar] [CrossRef]
- Jin, Y.H.; Zhang, Y.J.; Xu, J.W.; Tao, Y.; He, H.S.; Guo, M.; Wang, A.L.; Liu, Y.X.; Niu, L.P. Comparative Assessment of Tundra Vegetation Changes Between North and Southwest Slopes of Changbai Mountains, China, in Response to Global Warming. Chin. Geogr. Sci. 2018, 28, 665–679. [Google Scholar] [CrossRef] [Green Version]
- Jin, Y.H.; Xu, J.W.; Zong, S.W.; Wang, P. Experimental study on the effects of nitrogen deposition on the tundra vegetation of the Changbai Mountains. Sci. Geogr. Sin. 2014, 34, 1526–1532. [Google Scholar]
- Vonlanthen, C.M.; Kammer, P.M.; Eugster, W.; Bühler, A.; Veit, H. Alpine vascular plant species richness: The importance of daily maximum temperature and pH. Plant. Ecol. 2006, 184, 13–25. [Google Scholar] [CrossRef]
- Schöb, C.; Kammer, P.M.; Choler, P.; Veit, H. Small-scale plant species distribution in snowbeds and its sensitivity to climate change. Plant. Ecol. 2009, 200, 91–104. [Google Scholar] [CrossRef]
- Jiménez-Alfaro, B.; Marcenó, C.; Bueno, Á.; Gavilán, R.; Obeso, J.R. Biogeographic deconstruction of alpine plant communities along altitudinal and topographic gradients. J. Veg. Sci. 2014, 25, 160–171. [Google Scholar] [CrossRef]
- Madan, N.J. Snow Ecology: An Interdisciplinary Examination of Snow-covered Ecosystems. J. Ecol. 2001, 89, 1096–1099. [Google Scholar] [CrossRef]
- Choler, P. Consistent shifts in alpine plant traits along a mesotopographical gradient. Arct. Antarct. Alp. Res. 2005, 37, 444–453. [Google Scholar] [CrossRef]
- Bruun, H.H.; Moen, J.; Virtanen, R.; Grytnes, J.A.; Oksanen, L.; Angerbjörn, A. Effects of altitude andtopography on species richness of vascular plants, bryophytes and lichens in alpine communities. J. Veg. Sci. 2006, 17, 37–46. [Google Scholar] [CrossRef]
- Pickering, C.M.; Green, K. Vascular plant distribution in relation to topography, soils and micro-climate at five GLORIA sites in the Snowy Mountains, Australia. Aust. J. Bot. 2009, 57, 189–199. [Google Scholar] [CrossRef] [Green Version]
- Gutiérrez-Girón, A.; Gavilán, R.G. Spatial patterns and interspecific relations analysis help to betterunderstand species distribution patterns in a Mediterranean high mountain grassland. Plant. Ecol. 2010, 210, 137–151. [Google Scholar] [CrossRef]
- Cannone, N.; Sgorbati, S.; Guglielmin, M. Unexpected impacts of climate change on alpine vegetation. Front. Ecol. Environ. 2007, 5, 360–364. [Google Scholar] [CrossRef] [Green Version]
- Grabherr, G.; Gottfried, M.; Pauli, H. Climate change impacts in alpine environments. Geogr. Compass 2010, 4, 1133–1153. [Google Scholar] [CrossRef]
- Ferrer-Castán, D.; Vetaas, O.R. Floristic variation, chorological types and diversity: Do they correspond at broad and local scales? Divers. Distrib. 2003, 9, 221–235. [Google Scholar] [CrossRef]
- Blamires, D.; de Oliveira, G.; de Souza Barreto, B.; Diniz-Filho, J.A.F. Habitat use and deconstruction of richness patterns in Cerrado birds. Acta Oecol. 2008, 33, 97–104. [Google Scholar] [CrossRef]
- Azeria, E.T.; Bouchard, M.; Pothier, D.; Fortin, D.; Hébert, C. Using biodiversity deconstruction to disentangle assembly and diversity dynamics of understory plants along post-fire succession in boreal forest. Glob. Ecol. Biogeogr. 2011, 20, 119–133. [Google Scholar] [CrossRef]
- Huang, D.; Haack, R.A.; Zhang, R. Does global warming increase establishment rates of invasive alien species? A centurial time series analysis. PLoS ONE 2011, 6, e24733. [Google Scholar] [CrossRef] [PubMed]
- McDougall, K.L.; Khuroo, A.A.; Loope, L.L.; Parks, C.G.; Pauchard, A.; Reshi, Z.A.; Rushworth, I.; Kueffer, C. Plant invasions in mountains: Global lessons for better management. Mt. Res. Dev. 2011, 31, 380–387. [Google Scholar] [CrossRef]
- Malanson, G.P.; Fagre, D.B. Spatial contexts for temporal variability in alpine vegetation underongoing climate change. Plant. Ecol. 2013, 214, 1309–1319. [Google Scholar] [CrossRef]
- Inouye, D.W.; Mcguire, A.D. Effects of Snowpack on Timing and Abundance of Flowering in Delphinium-Nelsonii (Ranunculaceae)—Implications for Climate Change. Am. J. Bot. 1991, 78, 997–1001. [Google Scholar] [CrossRef]
- Roux, P.C.L.; Luoto, M. Earth surface processes drive the richness, composition and occurrence of plant species in an arctic-alpine environment. J. Veg. Sci. 2014, 25, 45–54. [Google Scholar] [CrossRef]
- Understory Response to Overstory and Soil Gradients in Mixed Versus Monospecifc Mediterranean Pine Forests. Available online: https://link.springer.com/article/10.1007/s10342-019-01215-0 (accessed on 26 July 2019).
- Wipf, S.; Stoeckli, V.; Bebi, P. Winter climate change in alpine tundra: Responses to changes in snow depth and snowmelt timing. Clim. Chang. 2009, 94, 105–121. [Google Scholar] [CrossRef]
- Wipf, S.; Rixen, C. A review of snow manipulation experiments in Arctic and alpine tundra ecosystems. Polar Res. 2010, 29, 95–109. [Google Scholar] [CrossRef]
- Bjorkman, A.D.; Elmendorf, S.C.; Beamish, A.L.; Vellend, M.; Henry, G.H.R. Contrasting effects of warming and increased snowfall on arctic tundra plant phenology over the past two decades. Glob. Chang. Biol. 2015, 21, 4651–4661. [Google Scholar] [CrossRef]
- Zong, S.; Jin, Y.; Xu, J.; Wu, Z.; He, H.; Du, H.; Wang, L. Nitrogen deposition but not climate warming promotes Deyeuxia angustifolia encroachment in alpine tundra of the Changbai Mountains, Northeast China. Sci. Total Environ. 2016, 544, 85–93. [Google Scholar] [CrossRef]
- Zhao, D.C. Preliminary study on the relationship between volcanic eruption and vegetation succession in Changbai Mountains. Resour. Sci. 1984, 1, 72–78. [Google Scholar]
- Mueller-Dombois, D.; Ellenberg, H. Aims and Methods of Vegetation Ecology; John Wiley and Sons: New York, NY, USA, 1974. [Google Scholar]
- Webster, M.R.; Oliver, M.A. Geostatistics for Environmental Scientists, 2nd ed.; Wiley: Chichester, UK, 2007; pp. 11–35. [Google Scholar]
- Lei, Z.D.; Yang, S.X.; Xie, S.C. Soil Dynamics; Tsinghua University Press: Beijing, China, 1988. [Google Scholar]
- Mirzazadeh, A.; Abdollahpor, S.; Vahed, M.M. Incorporating Skewness and Kurtosis in Improvement of Combine Harvester Cleaning System Performance. Int. J. Agric. Innov. Res. 2015, 3, 1412–1416. [Google Scholar]
- Zhang, C.Y.; Huang, Q.L.; Ma, Z.B.; Zheng, Q.R.; Wang, H. Diameter distribution in each storey and law of typical natural broad-leaved forest in mid-subtropical zone. Scientia Silvae Sinicae 2017, 53, 18–27. [Google Scholar]
- Shannon, S.E.; Weaver, W.; Wiener, N. The Mathematical Theory of Communication; Illinois University Press: Chicago, IL, USA, 1949. [Google Scholar]
- Nobes, K.; Uthicke, S.; Hendeson, R. Is light the limiting factor for the distribution of benthic bearing foraminifera on the Great Barrier Reef? J. Exp. Mar. Biol. Ecol. 2008, 363, 48–57. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plant Species | Dominant Shrubs | Dominant Herbs | ||||||
|---|---|---|---|---|---|---|---|---|
| R. chrysanthum | V. uliginosum | S. stipulata | S. parviflora | C. angustifolia | L. jamesii | S. tomentosa | G. baishanense | |
| IV | 20.56 | 3.62 | 8.88 | 8.59 | 6.76 | 4.70 | 3.81 | 3.07 |
| Plant Species | Abundance (mean ± 1SE) | CV | SK | KU | |
|---|---|---|---|---|---|
| Dominant Shrubs | R. chrysanthum | 53.83 ± 4.67 | 0.95 | 0.53 | −1.07 |
| V. uliginosum | 4.81 ± 1.36 | 3.23 | 6.27 | 47.77 | |
| Dominant Herbs | S. parviflora | 19.93 ± 1.81 | 1.04 | 1.04 | 0.63 |
| S. stipulata | 17.28 ± 2.39 | 1.59 | 1.74 | 2.11 | |
| C. angustifolia | 14.13 ± 2.49 | 2.03 | 3.1 | 13.77 | |
| L. jamesii | 8.18 ± 1.52 | 2.13 | 3.68 | 17.32 | |
| S. tomentosa | 5.67 ± 0.83 | 1.69 | 2.22 | 5.55 | |
| G. baishanense | 5.04 ± 0.87 | 1.97 | 2.39 | 5.09 |
| df | R. chrysanthum | V. uliginosum | S. parviflora | S. stipulata | L. jamesii | C. angustifolia | S. tomentosa | G. baishanense | Shrubs/Herbs | SW | |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Elevation | 4 | 0.583 | 0.003 | 0.062 | 0.006 | 0.035 | 0.00 | 0.00 | 0.00 | 0.286 | 0.000 |
| Micro-toPography | 2 | 0.505 | 0.000 | 0.119 | 0.000 | 0.012 | 0.000 | 0.006 | 0.002 | 0.001 | 0.277 |
| Slope Aspects | 5 | 0.012 | 0.012 | 0.165 | 0.000 | 0.010 | 0.072 | 0.054 | 0.000 | 0.053 | 0.081 |
| Slopes | 5 | 0.000 | 0.000 | 0.001 | 0.001 | 0.000 | 0.006 | 0.554 | 0.000 | 0.015 | 0.534 |
| Ordination Axes | Correlation Coefficient | Eigenvalues | Species-Environment Correlations | Cumulative Percentage Variance of Species-Environment Relation (%) | Monte Carlo Test | ||||
|---|---|---|---|---|---|---|---|---|---|
| Elevation | Slope Aspect | Slope Grade | Micro-toPography | Test of Significance of First Canonical Axis | Test of Significance of All Canonical Axis | ||||
| 1 | −0.143 | 0.06 | −0.29 | −0.574 | 0.129 | 0.666 | 75.6 | p = 0.006 | p = 0.002 |
| 2 | 0.102 | 0.186 | −0.0383 | 0.146 | 0.021 | 0.382 | 88.1 | ||
| Linear Regression Equation | Adjusted R Square | Sig. | |
|---|---|---|---|
| R. chrysanthum | Y = 67.902 + 3.403X1 − 5.474X2 + 7.357X3 − 5.611X4 | 0.441 | 0.034 |
| V. uliginosum | Y = 35.880 − 1.693X1 − 6.937X2 − 1.082X3 − 2.067X4 | 0.374 | 0.008 |
| S. parviflora | Y = 13.095 + 0.699X1 + 5.691X2 + 2.323X3 − 2.269X4 | 0.515 | 0.014 |
| S. stipulata | Y = −31.082 + 2.928X1 + 15.025X2 + 3.217X3 + 2.901X4 | 0.617 | 0.001 |
| C. angustifolia | Y = −1.275 − 1.488X1 + 15.901X2 + 0.882X3 − 2.085X4 | 0.568 | 0.011 |
| L. jamesii | Y = 28.393 − 1.955X1 − 0.819X2 − 3.146X3 + 0.189X4 | 0.011 | 0.249 |
| S. tomentosa | Y = 19.978 − 0.549X1 − 3.932X2 + 0.098X3 − 0.696X4 | 0.45 | 0.033 |
| G. baishanense | Y = −4.630 − 1.882X1 + 5.070X2 + 1.486X3 + 1.114X4 | 0.592 | 0.000 |
| Shrub/Herb ratio | Y = 2.065 + 0.022X1 − 0.511X2 + 0.018X3 − 0.075X4 | 0.514 | 0.004 |
| SW | Y = 1.192 − 0.106X1 | 0.572 | 0.000 |
| Elevation (m) X1 | Micro-Topography X2 | Aspects X3 | Slopes (°) X4 | Groups Assignment |
|---|---|---|---|---|
| 2050–2100 | convex | west | <5 | 1 |
| 2101–2150 | transitional zone | southwest | 6–15 | 2 |
| 2151–2200 | depression | south | 16–25 | 3 |
| 2201–2050 | southeast | 26–35 | 4 | |
| 2051–2200 | east | 36–45 | 5 | |
| north | >45 | 6 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jin, Y.; Xu, J.; He, H.; Li, M.-H.; Tao, Y.; Zhang, Y.; Hu, R.; Gao, X.; Bai, Y.; Wang, H.; et al. The Changbai Alpine Shrub Tundra Will Be Replaced by Herbaceous Tundra under Global Climate Change. Plants 2019, 8, 370. https://doi.org/10.3390/plants8100370
Jin Y, Xu J, He H, Li M-H, Tao Y, Zhang Y, Hu R, Gao X, Bai Y, Wang H, et al. The Changbai Alpine Shrub Tundra Will Be Replaced by Herbaceous Tundra under Global Climate Change. Plants. 2019; 8(10):370. https://doi.org/10.3390/plants8100370
Chicago/Turabian StyleJin, Yinghua, Jiawei Xu, Hongshi He, Mai-He Li, Yan Tao, Yingjie Zhang, Rui Hu, Xiang Gao, Yunyu Bai, Huiyun Wang, and et al. 2019. "The Changbai Alpine Shrub Tundra Will Be Replaced by Herbaceous Tundra under Global Climate Change" Plants 8, no. 10: 370. https://doi.org/10.3390/plants8100370