1. Introduction
Durum wheat (
Triticum turgidum L. ssp.
durum Desf.) is an important cereal crop, particularly in regions with limited water availability [
1,
2]. Algeria, being a country with diverse climatic conditions, including harsh environments, faces challenges in durum wheat cultivation due to water scarcity and other biotic and abiotic stresses [
3]. To address these challenges and enhance durum wheat production, researchers and breeders focus on exploiting the genetic variability of diverse quantitative and some qualitative traits within old and modern germplasm [
4,
5,
6,
7].
Durum wheat germplasm represents a diverse collection of genetic resources, which includes different varieties, landraces, and wild relatives [
8,
9]. This genetic diversity possesses a range of traits, including highly heritable characteristics like heading date, plant height, and kernel weight, but also a large number of quantitative traits, such as yield potential, drought tolerance, disease resistance, and nutrient use efficiency, which exhibit continuous variation [
10,
11,
12].
Through the systematic evaluation of diverse durum wheat germplasm collections, breeders have identified cultivars that possess desirable traits. This can be based on phenotypic traits like plant height, flowering time, grain yield, and quality characteristics, as well as genotypic traits revealed through molecular markers [
13,
14,
15]. After crossing cultivars carrying desirable traits, breeders—through traditional breeding methods—select superior progenies over multiple generations [
16]. Molecular breeding approaches, such as marker-assisted selection and genomic selection, facilitate the identification and utilization of specific genes or genomic regions associated with target traits [
17,
18].
Genetic and non-genetic parameters, such as phenotypic (
CVp) and genotypic coefficient of variation (
CVg), along with heritability (
h2bs), serve as valuable biometric indicators for assessing the genetic variation and adaptability within a germplasm collection. They provide insights into the relative contributions of genetic and environmental factors to observed variations in specific traits among individuals [
19,
20,
21].
Because grain yield is a polygenic complex trait that is controlled by many factors, and is extremely influenced by environmental variation, selection for high-yielding genotypes should not be based on yield only, but other yield-contributing traits should be taken into consideration during the selection process [
22,
23]. Hence, knowledge of the degree of relationship between yield and different yield-related components can identify the traits that could be used as indirect selection criteria, increasing the efficiency of the selection process. The genotypic and phenotypic correlations are concepts used by breeders to describe the association between the genetic makeup (genotype) and the observable traits (phenotype) of individuals within a population [
24]. Genotypic correlation focuses on the genetic basis of traits, while phenotypic correlation considers the observable expression of these traits in individuals, influenced by both genetic and environmental factors [
25].
In fact, the correlation coefficient only quantifies the link between each pair of traits and does not truly indicate the relative importance of secondary traits on the yield. Path coefficient is a common approach used in the field of crop breeding, where researchers are interested in understanding the complex relationships between grain yield and its related traits in order to identify those with significant effects on yield materialization to be used as selection criteria [
26]. This method has the advantage of portioning the correlation coefficient into direct and indirect relationships among a set of variables, helping to understand the complex interplay between the different factors in a theoretical model [
26]. The direct effect refers to the influence of one trait on another without the mediation of any other traits, whereas the indirect effect considers the impact of one trait on another that is mediated through one or more intervening traits in the model [
27]. According to Boulelouah et al. (2022) [
28], both correlation and path coefficient techniques could achieve knowledge regarding the proper cause-and-effect relationship between yield and yield components.
By understanding the morpho-genetic dissimilarity among diverse cultivars within the available germplasm, breeders can strategically choose parents for crossing with the goal of maximizing the potential for genetic gain. Cluster analysis is a statistical method used in the context of genetic diversity studies to classify cultivars of germplasm collections in specific heterotic groups [
29]. In this context, the generalized Mahalanobis (1936) distance method provides greater precision when selecting genotypes for future crosses. It was largely used as a consistent measure of dissimilarity in various crop species, including durum wheat [
30,
31]. This information is valuable for both understanding the genetic structure of the population and for guiding breeding strategies to harness the genetic diversity for crop improvement.
In view of this, the present investigation was conducted to explore the genetic divergence and variability present within a durum wheat germplasm collection through the phenotypic evaluation of various desirable characteristics of particular importance for local adaptation to semi-arid regions of Algeria. This evaluation allows breeders to identify individuals as potential parents for future breeding programs targeting the development of wheat varieties with enhanced productivity and resilience.
2. Results
2.1. Sources of Variation Analysis
The results showed highly significant differences (
p ≤ 0.01) among the tested genotypes for the majority of traits subjected to one-way analysis of variance (ANOVA), except for MT and BY, which were non-significant (
Table 1).
2.2. Mean Performance of Genotypes
The mean performance of 20 durum wheat genotypes for 15 morpho-physiological and agronomic traits at the Setif location is presented in
Table 2. The HD elapsed between 128.0 days in Ofanto and 138.0 days in Guemgoum Rkhem, with a mean of 132.4 days. The mean CC was 43.6 CCI. The minimum CC was 29.3 CCI, which was exhibited by Bousselam, whereas the maximum value of CC was recorded in Simeto, at 54.3 CCI. The CT varied between 29.7 °C in Megress and 36.9 °C in Bousselam, with an average value of 32.8 °C. The mean RWC was 70.6%. Belikh02 was the cultivar with the lowest RWC, i.e., 62.3%, whereas Mohamed Ben Bachir was the genotype with the highest water content in the leaves, at 81.7%. Despite the wide range observed for MT, the difference among the genotypes for this trait was not significant. The mean value of MT assessed in the wheat genotypes was 80.6%. Ofanto had the lowest cell MT, which was 51.4%, whereas Amar06 had the highest MT, which was 95.2%.
The mean FLA was 30.3 cm2. Belikh02 had the smallest leaves, at 21.9 cm2, whereas Waha had the largest leaves, at 36.3 cm2. The mean PH was 79.6 cm. Mexicali75 was the shortest cultivar, with a plant height of 69.3 cm, whereas Guemgoum Rkhem was the tallest genotype, with a PH of 108.7 cm. The mean value of all of the genotypes for SN was 378.7 spikes m−2. The lowest mean value of SN (225.0 spikes m−2) was obtained in the Guemgoum Rkhem cultivar, while the highest mean value of this trait (496.7 spikes m−2) was exhibited by Waha. The overall average SW was 4.1 t ha−1, with a range from 2.3 (Guemgoum Rkhem) to 5.2 t ha−1 (Wahbi). The mean value of TKW was 30.1 g. The lowest kernel weight (21.1 g) was exhibited by Bousselam, while the highest TKW (41.0 g) was recorded for Boutaleb. The minimum NGS recorded was 4.3 grain spike−1 for Vitron, whereas a maximum of 15.8 grains spike−1 was found for Manssourah, with an average value of 8.8 grains spike−1. The mean GY was 1.9 t ha−1. Zb/Fl had the lowest GY, at 1.1 t ha−1, whereas Manssourah had the highest GY, at 3.2 t ha−1. The minimum BY recorded was 7.2 t ha−1 for Belikh02, whereas a maximum of 12.3 t ha−1 was registered for Saoura, with a mean value of 10.9 t ha−1. Mohamed Ben Bachir had the lowest HI, i.e., 9.6% t ha−1, whereas Manssourah displayed the highest HI, i.e., 32.1%, with an average HI value of 17.9%.
2.3. Estimation of Components of Variation
The estimates of genetic and non-genetic parameters for the measured traits in the wheat cultivars are illustrated in
Table 3. The phenotypic coefficient of variation (
CVp) values varied between 1.99 for HD and 38.95% for GY. The genotypic coefficient of variation (
CVg) ranged from 1.99 for HD to 32.62% for HI. The
CVp, as well as the
CVg, values were categorized as low (<10%), moderate (10 to 20%), and high (>20%) [
32]. A high
CVp indicates a considerable variation for the trait within the population, which may be due to genetic differences, environmental influences, or their interactions. In this study, next to GY (37.63%), HI (37.63%), NGS (32.29%), SN (26.64%), SW (26.34%), CC (23.14%), and TKW (20.22%) also presented high
CVp estimates. By contrast, low values of
CVp were found in HD (1.99%), CT (5.76%), and RWC (6.31%), whereas the remaining recorded traits exhibited moderate values of
CVp.
2.4. Relationships among Measured Traits
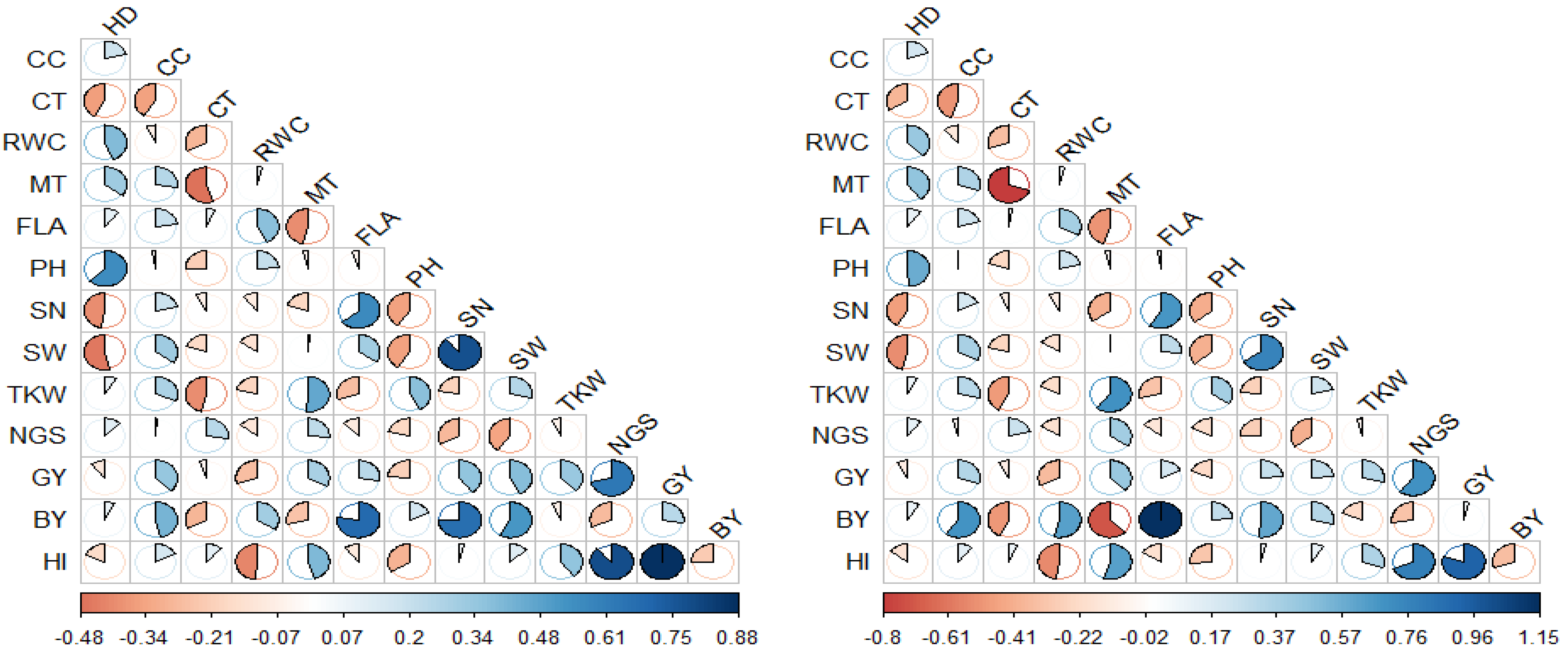
Figure 1 represents the phenotypic (
rp) and genotypic (
rg) correlation coefficients among the 13 morpho-physiological and agronomic traits measured in the durum wheat cultivars. At both genotypic and phenotypic levels, GY showed significant to highly significant and positive relationships with CC (
rp = 0.332 ** and
rg = 0.343 **), SN (
rp = 0.443 ** and
rg = 0.270 *), SW (
rp = 0.497 ** and
rg = 0.270 *), TKW (
rp = 0.337 ** and
rg = 0.332 **), NGS (
rp = 0.558 ** and
rg = 0.693 **), and HI (
rp = 0.833 ** and
rg = 0.926**). BY, in addition to its relation to GY, it had other strong correlations, including positives with CC (
rp = 0.256 * and
rg = 0.628 **), FLA (
rp = 0.429 ** and
rg = 1.152 **), SN (
rp = 0.739 ** and
rg = 0.599 **), and SW (
rp = 0.651 ** and
rg = 0.332 **); and a negative correlation with NGS (
rp = −0.262 * and
rg = −0.326 *). As discussed above, the flag leaf area and the biological yield were largely influenced by similar environmental conditions (
Table 3); therefore, the environmental factors that affected both traits may explain the overestimation of the genetic correlation coefficient value. On the other hand, HI demonstrated significant negative correlations with RWC (
rp = −0.270 * and
rg = −0.552 **) and PH (
rp = −0.287 * and
rg = −0.311 *), and positive associations with TKW (
rp = 0.336 ** and
rg = 0.357 **) and NGS (
rp = 0.762 ** and
rg = 0.792 **).
Worthy of note is the negative correlation of SN with HD (rp = −0.342 ** and rg = −0.474 **), PH (rp = −0.290 * and rg = −0.416 **), and NGS (rp = −0.292 * and rg = −0.288 *); and the positive correlations between SN and FLA (rp = 0.458 ** and rg = 0.671 **) and SN and SW (rp = 0.790 ** and rg = 0.762 **). The SW also showed negative relationships with HD (rp = −0.384 ** and rg = −0.531 **) and PH (rp = −0.293 * and rg = −0.414 **), and a positive correlation with FLA (rp = 0.307 * and rg = 0.299 *). The HD, in addition to its negative relations with SN and SW, showed highly significant correlations, including a negative correlation with CT (rp = −0.336 ** and rg = −0.381 **) and positive correlations with RWC (rp = 0.321 * and rg = 0.438 **) and PH (rp = 0.530 ** and rg = 0.570 **). The TKW exhibited a significant negative relationship with CT (rp = −0.490 ** and rg = −0.286 *) and significant positive correlations with MT (rp = 0.286 * and rg = 0.699 **) and PH (rp = 0.326 * and rg = 0.390 **). Furthermore, MT significantly correlated with CT (rp = −0.259 * and rg = −0.804 **) and FLA (rp = −0.361 ** and rg = −0.512 **). FLA and RWC were positively inter-correlated (rp = 0.358 ** and rg = 0.382 **) at both genotypic and phenotypic levels.
At the phenotypic level, GY presented significant to highly significant positive relationships with FLA (rp = 0.272 * and rg = 0.210 ns) and BY (rp = 0.390 ** and rg = 0.045 ns). Similarly, the data regarding the TKW measurements showed a significant positive correlation with SW (rp = 0.278 * and rg = 0.238 ns). At the genotypic level, the GY showed strong correlations, including a negative correlation with RWC (rp = −0.152 ns and rg = −0.366 **) and a positive correlation with MT (rp = 0.651 ** and rg = 0.332 ns). The BY significantly correlated with most of the physiological traits, including CT (rp = −0.176 ns and rg = −0.497 **), RWC (rp = 0.111 ns and rg = 0.625 **), and MT (rp = −0.059 ns and rg = −0.721 **). The BY also displayed a positive genotypic correlation with PH (rp = 0.085 ns and rg = 0.281 *) and a strong negative correlation with HI (rp = −0.152 ns and rg = −0.354 **). The latter also demonstrated a highly significant positive association with MT (rp = 0.241 ns and rg = 0.629 **). Other significant correlations were also observed among the measured traits. For example, negative correlations of SN with MT (rp = −0.033 ns and rg = −0.409 **) and TKW (rp = −0.113 ns and rg = −0.269 *) were found. The latter trait also demonstrated other important correlations, including a positive correlation with CC (rp = 0.222 ns and rg = 0.322 *) and a negative with correlation FLA (rp = −0.200 ns and rg = −0.326 *). The CC exhibited strong positive relationships with MT (rp = 0.186 ns and rg = 0.353**) and SW (rp = 0.241 ns and rg = 0.378 **) and a negative correlation with CT (rp = −0.186 ns and rg = −0.508 **). The CT and RWC were negatively inter-related (rp = −0.221 ns and rg = −0.349 **). The data regarding the MT measurements showed highly significant positive correlations with HD (rp = 0.218 ns and rg = 0.460 **) and NGS (rp = 0.105 ns and rg = 0.398 **).
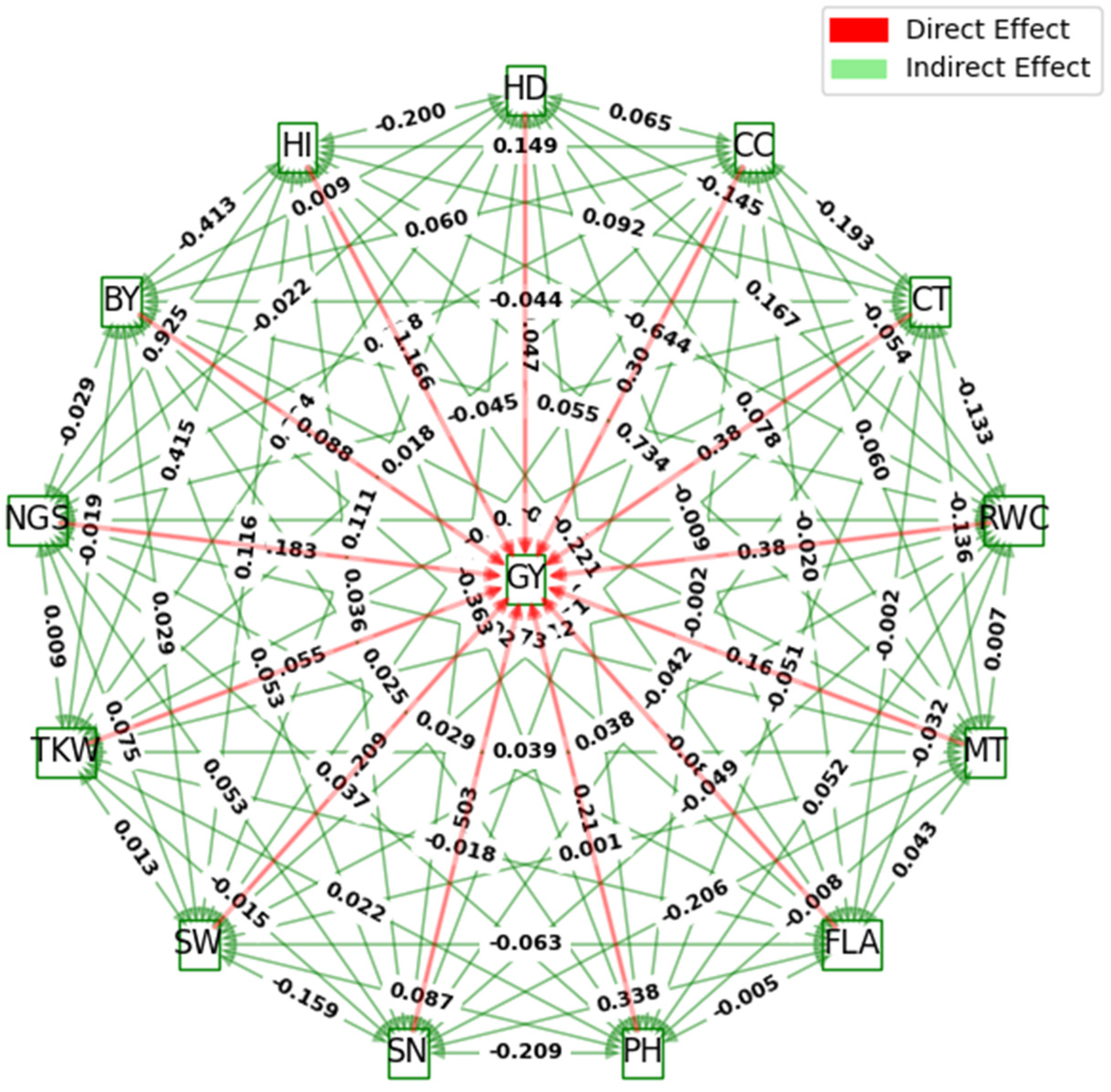
2.5. Estimate of Direct and Indirect Effects at a Genotypic Level
The results of the path coefficient analysis of 12 measured traits to GY are given in
Table 4 and
Figure 2. Dewey and Lu (1959) [
26] classified path coefficient values as very high (>1.0), high (0.30–0.99), moderate (0.2–0.29), low (0.1–0.19), and negligible (0.00–0.09). Based on this, the harvest index (1.166) and RWC (0.381) exhibited very high and high positive direct effects on GY, respectively. Traits such as SN (0.503), CT (0.380), and CC (0.302) exerted moderate positive direct effects on GY, similar to their genotypic correlations with this primary trait (
Table 4;
Figure 1). The PH (0.215) and MT (0.169) expressed moderate-to-low direct effects on GY. Conversely, SW (−0.209), next to NGS (−0.183), yielded the first and second highest negative direct effects on GY, respectivley. While HD, FLA, and BY had non-significant associations with GY at a genotypic level, it has been stated in
Table 4 and
Figure 2 that the SW and NGS demonstrated positive and statistically significant correlations with the grain yield per hectare.
The NGS (0.925), MT (0.734), and TKW (0.415) depicted the highest positive indirect effects on GY via HI. Furthermore, SW (0.383), FLA (0.338), and BY (0.301) exhibited high positive indirect effects on GY through SN. The BY expressed moderate positive indirect effects on GY through RWC (0.238) and CC (0.206). On the other hand, the RWC (−0.644), followed by the BY (−0.413), PH (−0.363), FLA (−0.221), and HD (−0.200), demonstrated high-to-moderate negative indirect effects on GY via HI. Similarly, the HD (−0.239), next to the PH (−0.209) and MT (−0.206), had moderate negative indirect effects on GY through SN. MT, via CT (−0.306), and HI, via RWC (−0.210), also exerted high and moderate negative indirect effects on GY, respectively. The rest of the indirect effects on GY were low-to-negligible, as shown in
Table 4 and
Figure 2.
2.6. Divergence among Wheat Cultivars
The 20 durum wheat cultivars were classified into seven distinct clusters or groups, namely, G1, G2, G3, G4, G5, G6, and G7 (
Table 5). Eight cultivars were found in the first cluster (G1), accounting for 40% of total genotypes, followed by four cultivars classified in the second cluster (G2), with a relative contribution of 20% to the genetic diversity of the durum wheat germplasm. The third cluster (G3) contained three genotypes, while the fourth cluster (G4) was formed by two cultivars only. Besides these four clusters, the remaining three clusters only had one cultivar (
Table 5).
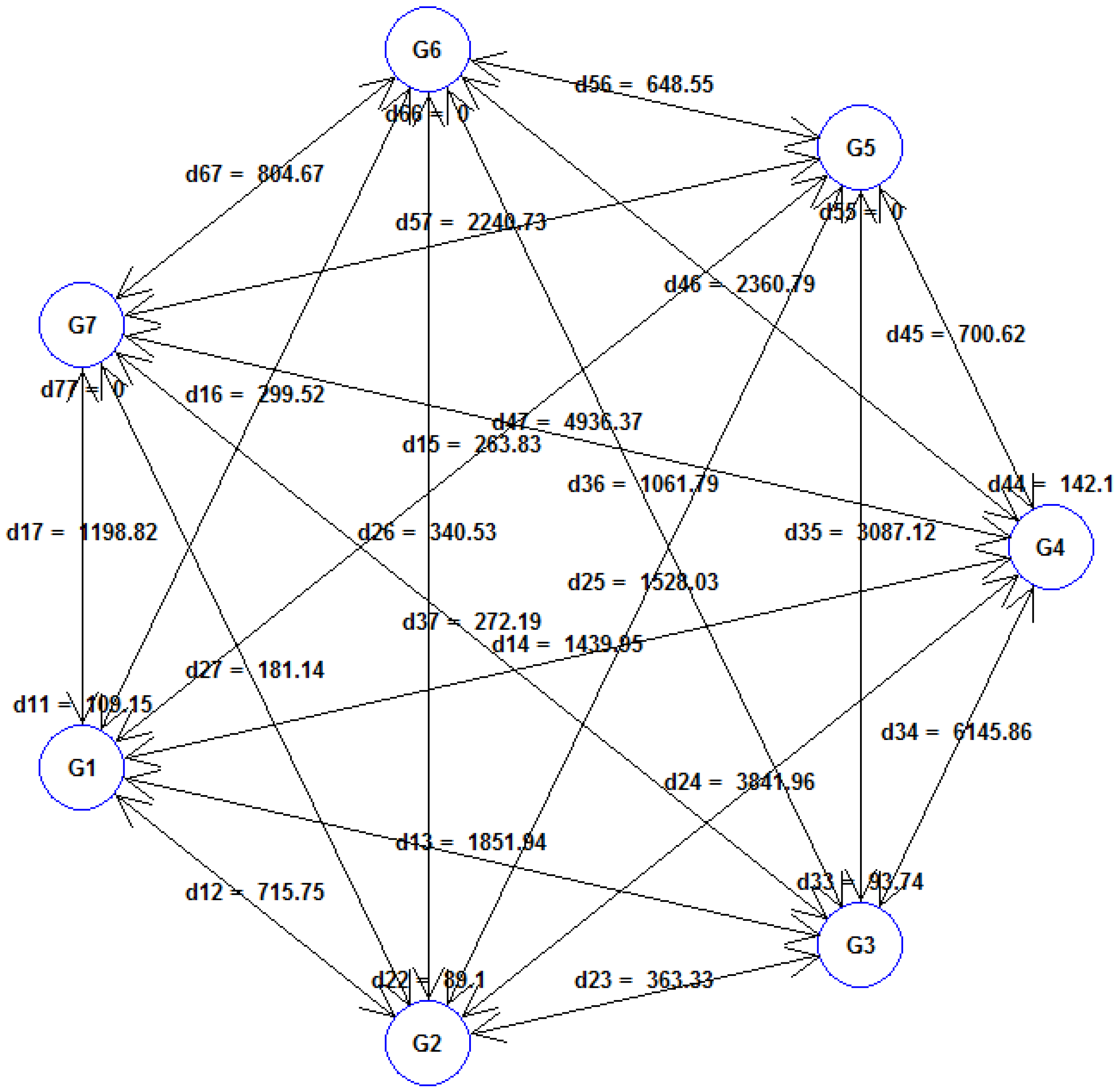
Figure 3 illustrates the average distances within and between the seven clusters formed by Tocher’s method based on Mahalanobis Euclidean
2 distance. The average inter-cluster distance was observed at maximum (
D2 = 6145.86) between cluster G3 and cluster G4. The lowest inter-cluster distance (
D2 = 181.14) was recorded between clusters G2 and G7, which exhibited more genetic similarity. The maximum intra-cluster distance was obtained in cluster G4 (
D2 = 142.10), followed by cluster G1 (
D2 = 109.16), indicating that the cultivars belonging to these clusters were far diverged from those within clusters G2 (
D2 = 89.10) and G3 (
D2 = 93.75). Clusters G5, G6, and G7 did not show any intra-cluster distance. Among the fourteen traits studied, the greatest contribution to cluster divergence was made by HD (92.33%), followed by plant height (2.08%), thousand kernel weight (1.61%), and spike weight (1.52%).
The average of the studied traits for each cluster has been represented in
Table 6. Accordingly, cluster G4 was characterized by having the highest cluster mean for days to 50% heading (137.5 days), while cluster G3 had the lowest HD mean (128.7 days). G3 and G4 exhibited, respectively, the lowest (67.0) and the highest (75.4 CCI) chlorophyll content in the leaves. The cultivars grouped in cluster G1 showed the highest CT (47.2%), GY (2.4 t ha
−1), BY (11.6 t ha
−1), and HI (21.2%) values.
Cluster G2 showed the highest SN (411.3 spikes m−2), whereas the lowest SN (240.8 spikes m−2) was observed in cluster G4, which had also the longest plant stature (104.7 cm) and the lowest SW (2.4 t ha−1), GY (1.1 t ha−1), and HI (9.8%) values. Cluster G5 is characterized by the shortest plants (73.3 cm) and the highest RWC (91.2%) and NGS (10.1 grains spike−1). Cluster G6 had the coolest canopy cover (29.3 °C), the largest leaves (33.8 cm2), the highest MT (36.9%), and the smallest grain size (21.1 g). Cluster G7 had the lowest cluster mean for MT (31.5%), the smallest leaves (24.7 cm2), the lowest spike fertility (4.3 grains spike−1), and the lowest BY (9.2 t ha−1). This cluster is characterized also by the highest SW (5.0 t ha−1) and the largest grain size (40.1 g).
3. Discussion
The results of the present study have revealed considerable variability in plant material, suggesting ample opportunities for wheat breeders to exploit this genetic diversity in breeding programs. Previous research in the Eastern Algerian High Plateaus supports these findings [
33,
34,
35]. Researchers have noticed a wide variation in physiological, yield, and yield-related traits. The lack of significant differences in aboveground biomass may be due to water stress and increased temperatures at the post-anthesis growth stage, reducing cell division and elongation, leaf area, and grain filling period. The absence of significant differences in cell membrane thermostability could prompt further genetic studies by local breeders to enhance heat stress tolerance through hybridization and selection programs.
This study demonstrates that the mean trait values vary among the different genotypes of durum wheat, indicating distinct trait profiles and underlying genetic differences that influence morpho-physiological and agronomic features. This variability is crucial for selecting and breeding durum wheat varieties with desired characteristics, such as a higher yield, disease resistance, environmental stress tolerance, and enhanced nutritional content. By identifying the superior genotypes for specific traits, breeders can strategically design breeding programs to develop improved durum wheat varieties suited to various growing conditions and end-use requirements. [
36].
This study has revealed varying levels of
CVg in durum wheat germplasm, with some traits exhibiting high-to-moderate
CVg values, offering opportunities for improvement through selective breeding. Conversely, the
CVg estimates were low for other traits, implying limited genetic variability that may hinder traditional selective breeding methods [
37]. Additionally, the difference between
CVp and
CVg values has highlighted the influence of environmental factors on trait variability, particularly for traits like CC, MT, FLA, and yield components, excluding TKW. In such a situation, breeders should consider this information when developing strategies to enhance genotypic stability and environmental adaptability in wheat varieties [
38,
39]. Apart from MT and FLA, most of the measured traits in the durum wheat germplasm displayed high genetic control (
CVg/
CVe > 1), making them favorable targets for breeding programs, due to their responsiveness to selection efforts. On the other hand, those traits with a variation index below one (
CVg/
CVe < 1) are predominantly influenced by environmental factors, rendering them less predictable across various growing conditions [
40]. Consequently, indirect selection, or the consideration of environmental factors, may be necessary to effectively improve these traits [
22]. The broad-sense heritability (
h2bs) has further supported these findings, indicating traits with high genetic determinism, like HD, CT, PH, TKW, NGS, and HI, which are desirable for breeding programs, due to their amenability to selection efforts.
h2bs was just intermediate for CC, RWC, FLA, SN, SW, and GY, suggesting that both genetic and environmental factors contribute to the observed variation in these traits. Conversely, less heritable traits pose challenges in breeding, due to their susceptibility to environmental influences.
Bendjama and Ramdani (2022) [
20] also observed high heritability for most agro-morphological traits in local durum wheat varieties grown in Algeria. However, broad-sense heritability varies depending on the population and environmental conditions [
41], and it does not reveal the specific genetic mechanisms underlying the traits [
19]. It only quantifies the overall genetic contribution to trait variation. Understanding the balance between genetic and environmental factors is essential for assessing the risks in genotype selection and minimizing environmental impacts on breeding outcomes.
This study has uncovered significant relationships between various traits in durum wheat, at both genotypic and phenotypic levels. The genotypic coefficients of correlation were generally larger than their corresponding phenotypic coefficients. Some significant genotypic correlations were not statistically significant at the phenotypic level, possibly due to environmental factors or sample size limitations [
42]. Conversely, instances where phenotypic correlation coefficients exceeded genotypic ones could indicate gene–environment interactions, where the environmental conditions modulate the impact of the genetic factors on traits, altering the observed correlations between genotypic and phenotypic levels [
43].
The results indicate that all of the cultivars had a long growth cycle with fewer fertile tillers and light spikes, but they showed tolerance to water deficit with high above-ground biomass. Despite this, they had a low harvest index and productivity. The tall cultivars had a larger grain size, due to a longer grain-filling period, allowing for more nutrient and energy accumulation, leading to higher yields under favorable conditions [
44]. Conversely, the short cultivars produced heavier spikes with better fertility, more spikes per unit surface, and efficient assimilate translocation, resulting in increased biological and grain yields. This suggests a link between dwarfing genes and genes controlling spike-related traits and the plant’s capacity to allocate assimilates to grains (source–sink relationship).
Fellahi et al. (2023) [
45] found that semi-dwarf wheat breeding lines carrying the Rht-D1b and Rht-B1b mutant alleles outperformed the tall lines in yield under supplemental irrigation and in spike number under rainfed conditions, along with other yield-related traits, such as harvest index and number of grains per spike, regardless of the environment. This observation is supported by the present investigation, which has revealed strong correlations between the yield and various yield attributes. Rabti et al. (2020) [
5] similarly emphasized the significant influence of the spike number, spike fertility, and harvest index on the grain yield in both old and modern durum wheat varieties. These findings suggest that improving these agronomic components could substantially increase yield in the screened plant material.
According to Maeoka et al. (2020) [
46], the improvement in yield of modern wheat varieties is strongly linked to reduced plant height, increased spike fertility, and higher harvest index, with no significant changes in biomass. Correlation analysis highlights the benefit of high above-ground biomass, particularly in adverse environmental conditions like drought. The cultivars with higher biological yield exhibited cooler canopy cover, better water status, and increased chlorophyll content, resulting in enhanced responses to post-anthesis water deficit and heat stress. Similar observations were made by Talebi (2011) [
47], suggesting that wheat genotypes with a lower canopy temperature may have higher rates of transpiration and photosynthesis, leading to increased yield in water-stressed environments.
The genotypic path analysis results showed that the harvest index (HI) had a highly positive direct effect on grain yield (GY), consistent with findings in the existing literature [
48,
49]. Traits like spike number (SN), relative water content (RWC), canopy temperature (CT), and canopy cover (CC) had moderate positive direct effects on GY, reflecting their genotypic correlations with this primary trait. Despite RWC displaying a high positive direct effect on GY, it paradoxically showed a negative correlation with GY, suggesting a positive contribution to the overall crop yield. However, the negative genetic inter-correlation between RWC and GY indicates a tendency for genetic variations in RWC to be inversely related to variations in yield. This underscores the complexity of genetic interactions [
50]. Plant height (PH) and maturity (MT) exhibited moderate-to-low direct effects on GY, with the effect of PH being rendered non-significant, due to the negative indirect effects of other yield components via this secondary morphological trait.
Our results suggest that more fertile tillers, along with a good plant capacity to allocate photosynthesis products (assimilates) from vegetative organs into the formed reproductive parts (grains), lead to incremented grain yield. Thus, the significant positive genotypic correlations of GY with SN and HI are predominately attributed to the direct effect of these two traits on yield per hectare. The results presented here align with the previous research findings documented in the literature [
51,
52,
53]. TKW exerted a negligible direct effect on GY, implying the minimal impact of the grain size on the overall yield in wheat cultivars. However, the positive and significant genotypic inter-correlation between TKW and GY suggests that, while TKW may not directly influence yield, it could be associated with other factors or processes that indirectly contribute to the yield. This hidden contribution warrants further exploration for wheat improvement.
In the scope of this study, SW, followed by NGS, had the highest negative direct effects on GY. Despite this negative impact, they both demonstrated positive and statistically significant correlations with grain yield per hectare, which implies the true relationship between these two secondary traits with the main dependent trait, i.e., GY. This observation could be attributed to the influence of common genes with pleiotropic effects [
54]. While the direct effect of the trait was negative, the other effects of these shared genes could be positive for grain yield. Moreover, traits like spike weight and spike fertility might negatively affect one aspect of the plant’s physiology or morphology that directly contributes to the yield. However, compensatory mechanisms, possibly involving other traits or physiological processes, might be at play, mitigating their negative impacts. In addition, the relationship could be sensitive to environmental conditions. Under certain conditions, the negative direct effect might dominate, while under different conditions, the positive genetic correlation with yield becomes more evident. Thus, selection based on spike weight would be effective but should be practiced cautiously [
38].
Path analysis has illustrated that NGS, MT, and TKW expressed the highest positive indirect effects on GY through HI. Additionally, SW, FLA, and BY depicted high positive indirect effects on GY via SN. BY also exhibited moderate positive indirect effects on GY via RWC and CC. These results confirm the importance of SN and HI and elucidate that the improvement of yield under drought stress during the grain filling stage could be more effective through selection.
Tocher’s clustering partitioned the twenty durum wheat cultivars into seven distinct clusters. Notably, the maximum inter-cluster distance was found to be at maximum between clusters G3 and G4, indicating a wide dissimilarity between the cultivars in these clusters. The greatest contribution to cluster divergence was made by HD, followed by PH, TKW, and SW. This divergence may be due to historical breeding practices, geographic isolation, or other factors influencing the genetic makeup of these wheat cultivars [
55]. Rabti et al. (2020) [
5] have also stated that earliness had a strong influence on grain yield variability in both old and recent durum wheat germplasm.
Understanding the genetic relationships and distances between different accessions enables breeders to strategically select parents for hybridization, enhancing specific traits or overall performance [
56]. Crossing genotypes from genetically divergent clusters can generate transgressive segregants, increasing the genetic variation in the offspring and facilitating the development of new potential varieties [
57]. The genotypes in distant groups may possess unique or rare traits important for crop improvement, particularly in addressing challenges such as diseases, pests, or environmental stress [
58]. However, the agronomic potential of those genotypes must be considered in order to develop potential varieties through cross combinations in wheat breeding programs [
59,
60]. The results have shown that cluster G1 could serve as a parent for developing high-yielding wheat varieties, while the presence of early heading cultivars in cluster G3 suggests the potential for developing early maturing varieties to mitigate heat waves during grain filling. The genotypes in other clusters may be selected based on their trait mean values, depending on the desired direction of selection (i.e., increase or decrease).

 ,
, 




{kind=link}
{kind=link}
{kind=link}
{kind=link}