
Nutritional Characterization of Chilean Landraces of Common Bean
 , , , ,
, , , ,  , ,
, ,
Abstract
:1. Introduction
2. Results
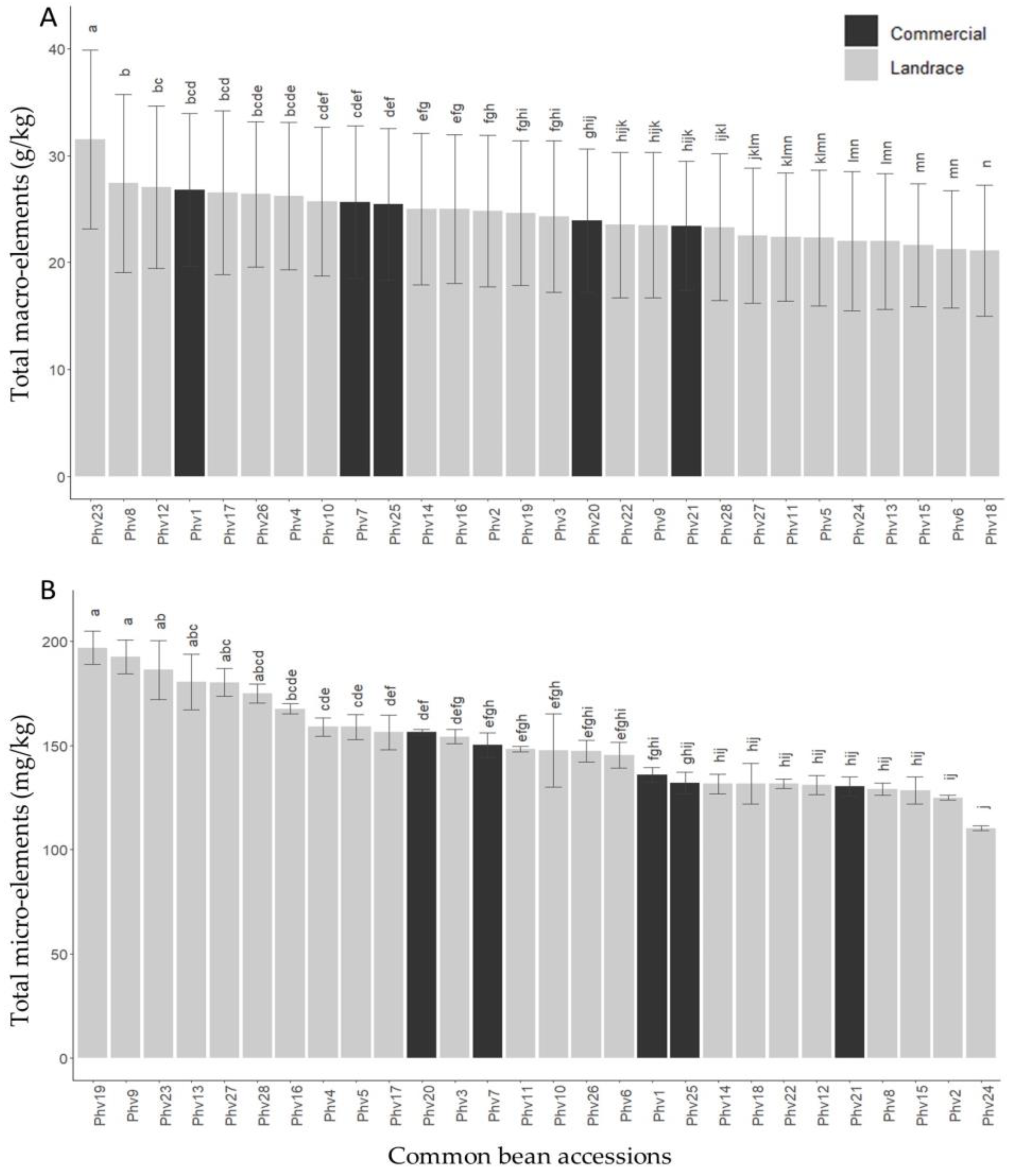
2.1. Variability among Chilean Common Beans
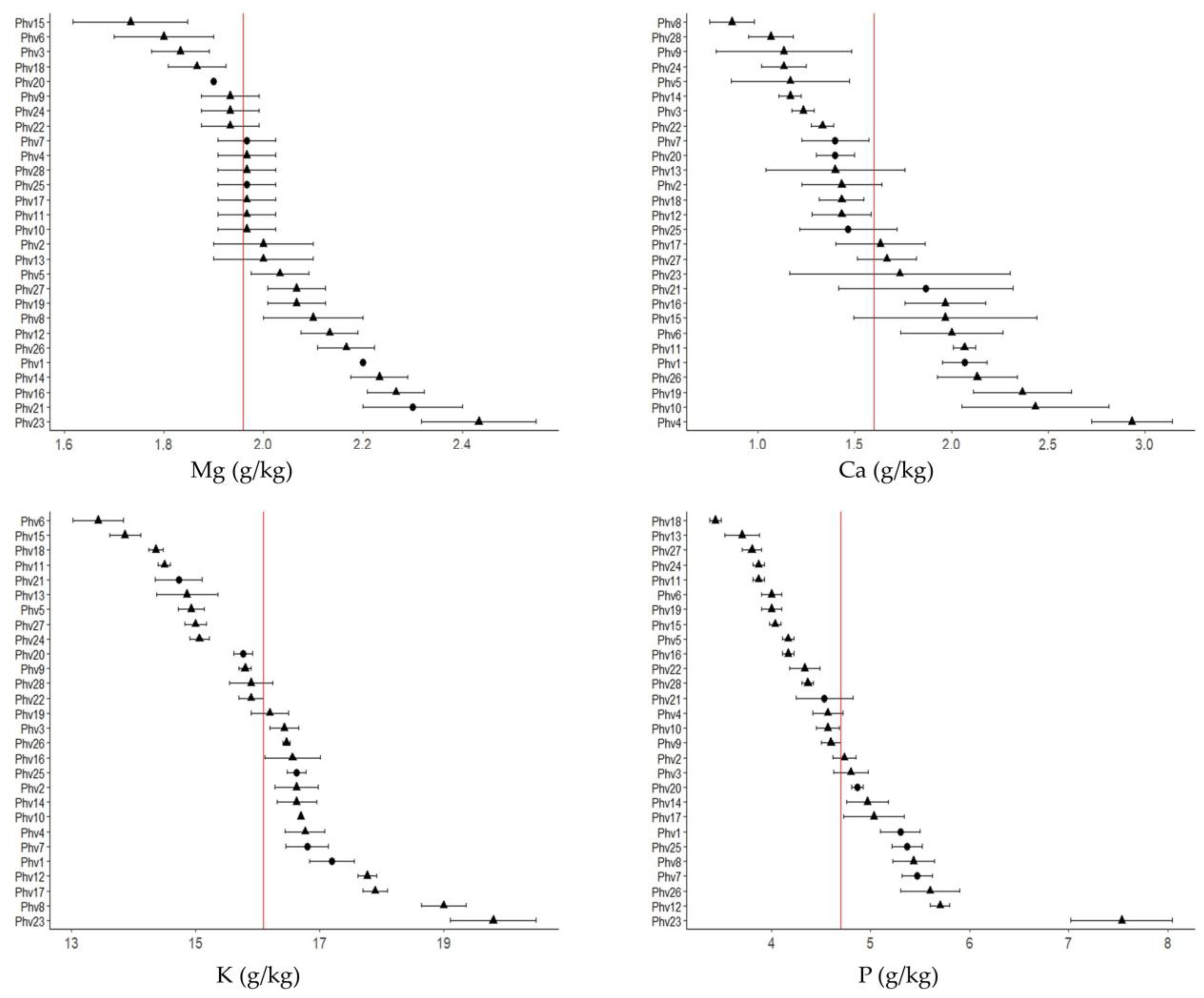
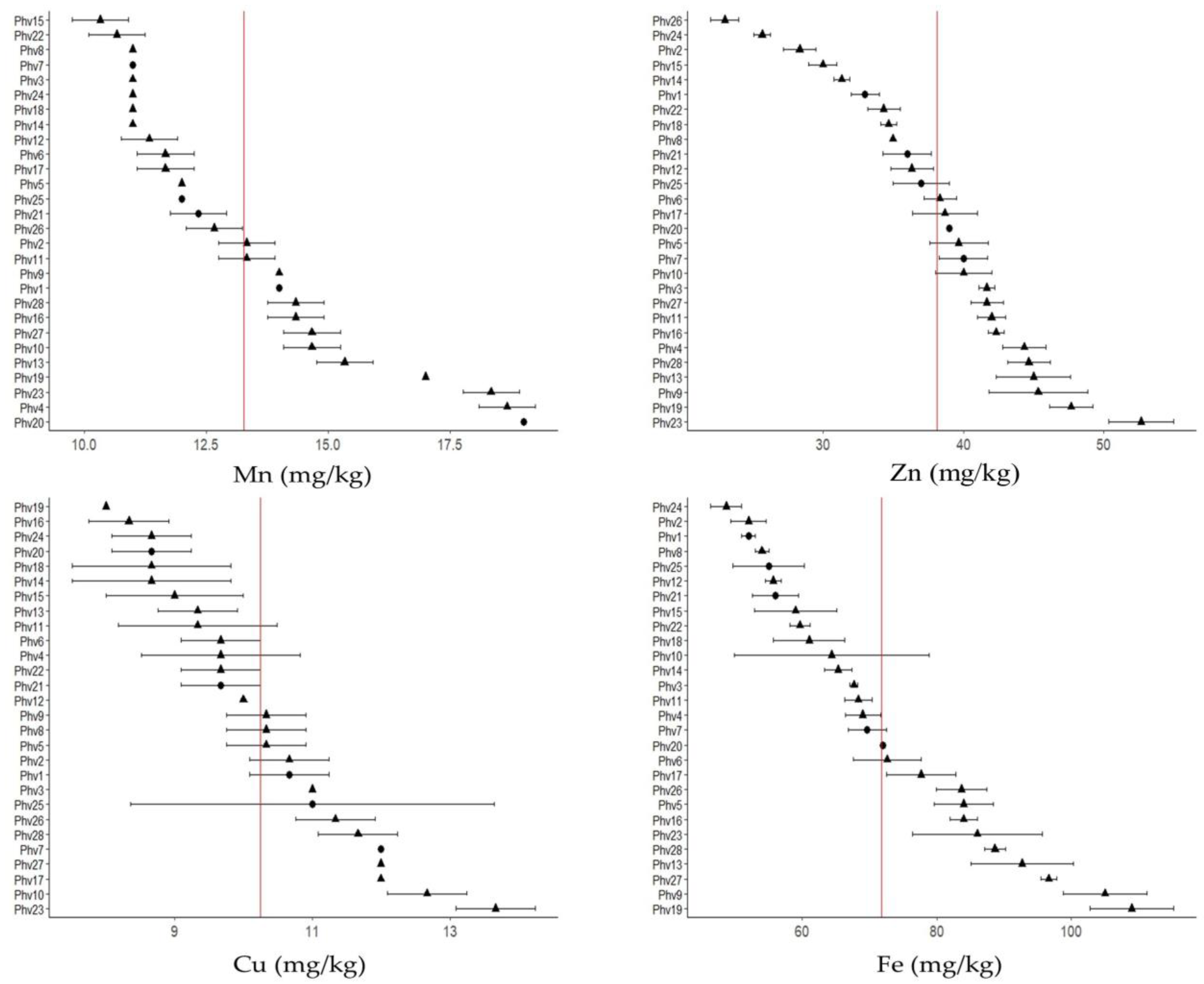
2.2. Mineral Contents
2.3. Amino Acid Profile and Protein Content
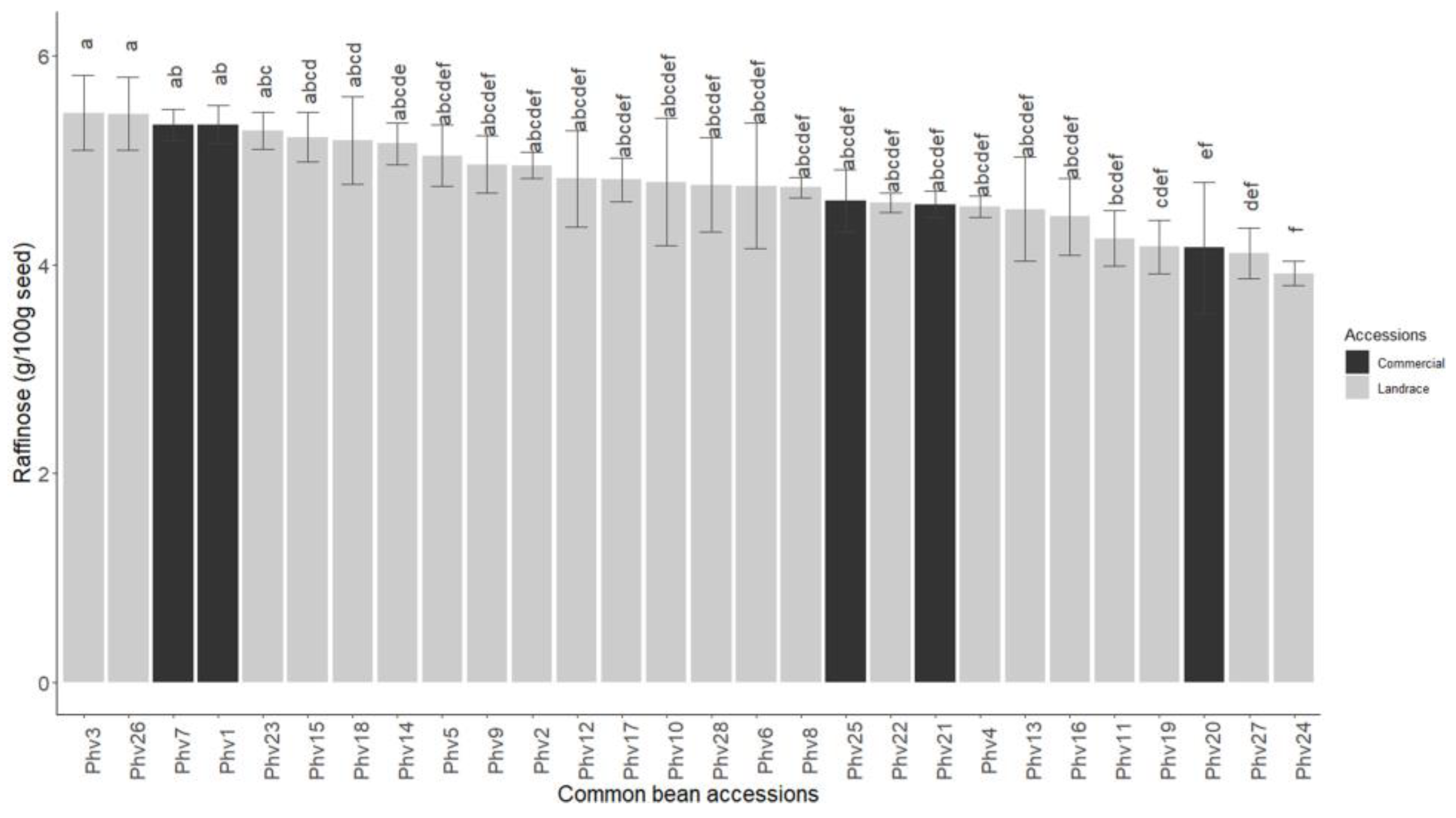
2.4. Sucrose and Raffinose Content
2.5. Antioxidant Activity and Phenolic Concentration
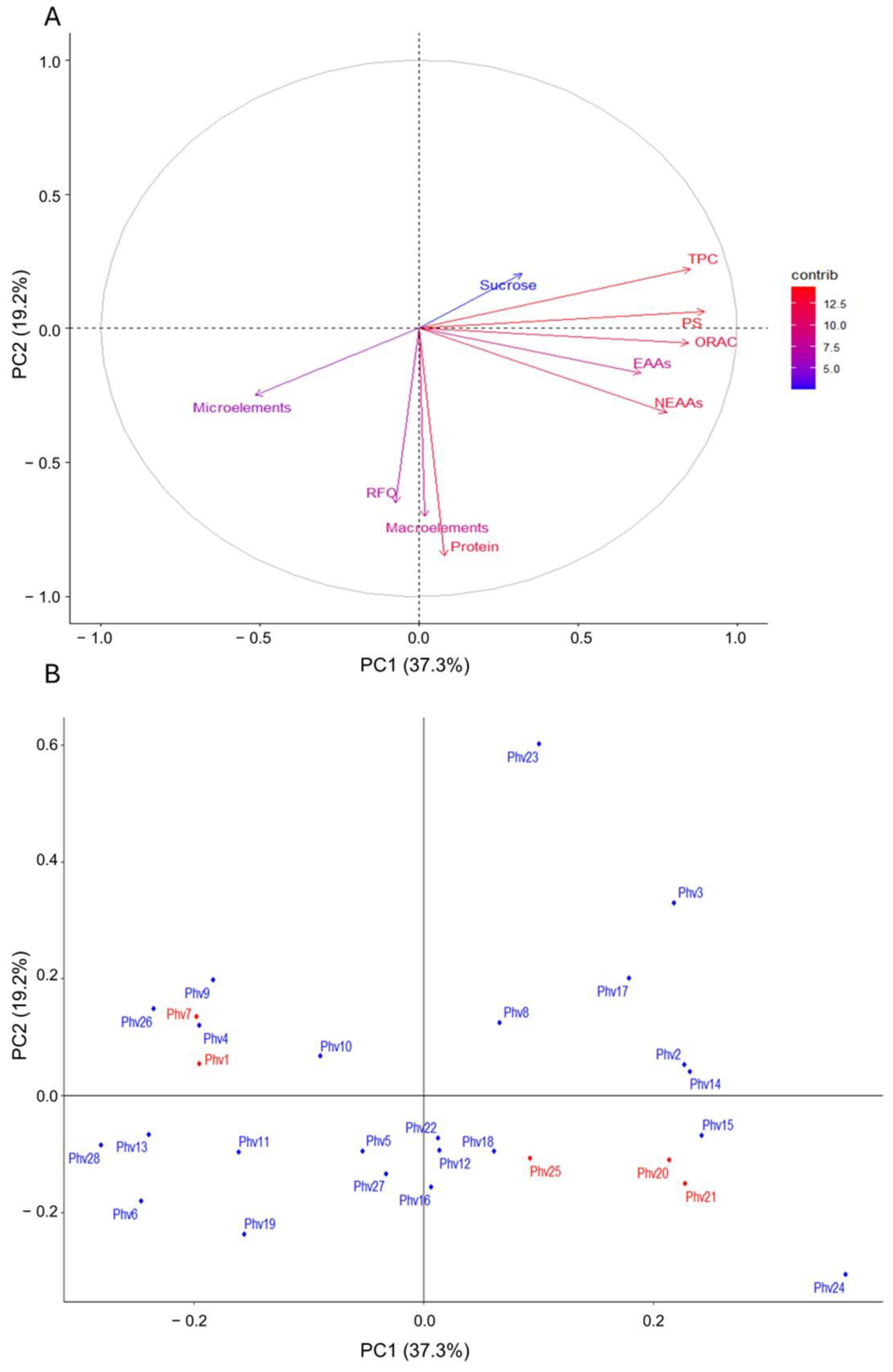
2.6. PCA and Correlations between the Traits
3. Discussion
3.1. Variability among Chilean Common Beans
3.2. Mineral Contents
3.3. Amino Acid Profile and Protein Content
3.4. Sucrose and Raffinose Content
3.5. Antioxidant Activity and Phenolic Content
4. Materials and Methods
4.1. Plant Materials
4.2. Mineral Composition
4.3. Amino Acids Composition
4.4. Sugars Content
4.5. Total Phenolic Content (TPC)
4.6. Antioxidant Activity
4.7. Determination of Tryptophan and Its Oxidation Products
4.8. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Broughton, W.J.; Hernández, G.; Blair, M.; Beebe, S.; Gepts, P.; Vanderleyden, J. Beans (Phaseolus spp.)—Model food legumes. Plant Soil 2003, 252, 55–128. [Google Scholar] [CrossRef]
- Araújo, S.S.; Beebe, S.; Crespi, M.; Delbreil, B.; González, E.M.; Gruber, V.; Lejeune-Henaut, I.; Link, W.; Monteros, M.J.; Prats, E.; et al. Abiotic stress responses in legumes: Strategies used to cope with environmental challenges. Crit. Rev. Plant Sci. 2015, 34, 237–280. [Google Scholar] [CrossRef]
- Labastida, D.; Ingvarsson, P.K.; Rendon-Anaya, M. Dissecting the genetic basis of drought responses in common bean using natural variation. Front Plant Sci. 2023, 14, 1143873. [Google Scholar] [CrossRef]
- Kristensen, M.D.; Bendsen, N.T.; Christensen, S.M.; Astrup, A.; Raben, A. Meals based on vegetable protein sources (beans and peas) are more satiating than meals based on animal protein sources (veal and pork)—A randomized cross-over meal test study. Food Nutr. Res. 2016, 60, 32634. [Google Scholar] [CrossRef]
- Tas, A.A.; Shah, A.U. The replacement of cereals by legumes in extruded snack foods: Science, technology and challenges. Trends Food Sci. Technol. 2021, 116, 701–711. [Google Scholar] [CrossRef]
- FAO. FAOSTAT Statistical Database. 2022. Available online: https://www.fao.org/faostat/en/#home (accessed on 1 March 2024).
- Hayat, I.; Ahmad, A.; Masud, T.; Ahmed, A.; Bashir, S. Nutritional and health perspectives of beans (Phaseolus vulgaris L.): An overview. Cr. Rev. Food Sci. Nutr. 2014, 54, 580–592. [Google Scholar] [CrossRef]
- Maphosa, Y.; Jideani, V.A.; Maphosa, Y.; Jideani, V.A. The Role of Legumes in Human Nutrition. In Functional Food—Improve Health through Adequate Food; IntechOpen: Rijeka, Croatia, 2017; pp. 103–122. [Google Scholar]
- Amarowicz, R.; Pegg, R.B. Legumes as a source of natural antioxidants. Eur. J. Lipid Sci. Tech. 2008, 110, 865–878. [Google Scholar] [CrossRef]
- Messina, V. Nutritional and health benefits of dried beans. Am. J. Clin. Nutr. 2014, 100, 437–442. [Google Scholar] [CrossRef] [PubMed]
- Carbas, B.; Machado, N.; Oppolzer, D.; Ferreira, L.; Queiroz, M.; Brites, C.; Rosa, E.A.; Barros, A.I. Nutrients, antinutrients, phenolic composition, and antioxidant activity of common bean cultivars and their potential for food applications. Antioxidants 2020, 9, 186. [Google Scholar] [CrossRef] [PubMed]
- Saleh, S.; Liu, G.; Liu, M.; Ji, Y.; He, H.; Gruda, N. Effect of irrigation on growth, yield, and chemical composition of two green bean cultivars. Horticulturae 2018, 4, 3. [Google Scholar] [CrossRef]
- Nicoletto, C.; Zanin, G.; Sambo, P.; Dalla Costa, L. Quality assessment of typical common bean genotypes cultivated in temperate climate conditions and different growth locations. Sci. Hortic. 2019, 256, 108599. [Google Scholar] [CrossRef]
- Shang, R.; Wu, H.; Guo, R.; Liu, Q.; Pan, L.; Li, J.; Hu, Z.; Chen, C. The diversity of four anti-nutritional factors in common bean. Hortic. Plant J. 2016, 2, 97–104. [Google Scholar] [CrossRef]
- Ribeiro, N.D.; dos Santos, G.G.; Maziero, S.M.; dos Santos, G.G. Genetic diversity and selection of bean landraces and cultivars based on technological and nutritional traits. J. Food Compos. Anal. 2021, 96, 103721. [Google Scholar] [CrossRef]
- Smýkal, P.; Nelson, M.N.; Berger, J.D.; Von Wettberg, E.J. The impact of genetic changes during crop domestication on healthy food development. Agronomy 2018, 8, 26. [Google Scholar] [CrossRef]
- Assefa, T.; Assibi Mahama, A.; Brown, A.V.; Cannon, E.K.; Rubyogo, J.C.; Rao, I.M.; Blair, M.W.; Cannon, S.B. A review of breeding objectives, genomic resources, and marker-assisted methods in common bean (Phaseolus vulgaris L.). Mol. Breed. 2019, 39, 1–23. [Google Scholar] [CrossRef]
- Schmutz, J.; McClean, P.E.; Mamidi, S.; Wu, G.A.; Cannon, S.B.; Grimwood, J.; Jenkins, J.; Shu, S.; Song, Q.; Chavarro, C.; et al. A reference genome for common bean and genome-wide analysis of dual domestications. Nat. Genet. 2014, 46, 707–713. [Google Scholar] [CrossRef] [PubMed]
- Trucchi, E.; Benazzo, A.; Lari, M.; Iob, A.; Vai, S. Ancient genomes reveal early Andean farmers selected common beans while preserving diversity. Nat. Plants 2021, 7, 123–128. [Google Scholar] [CrossRef] [PubMed]
- Paredes, M.; Becerra, V.; Tay, J. Inorganic nutritional composition of common bean (Phaseolus vulgaris L.) genotypes race Chile. Chil. J. Agr. Res. 2009, 69, 486–495. [Google Scholar] [CrossRef]
- Celmeli, T.; Sari, H.; Canci, H.; Sari, D.; Adak, A.; Eker, T.; Toker, C. The nutritional content of common bean (Phaseolus vulgaris L.) landraces in comparison to modern varieties. Agronomy 2018, 8, 166. [Google Scholar] [CrossRef]
- Blair, M.W.; Diaz, J.M.; Hidalgo, R.; Diaz, L.M.; Duque, M.C. Microsatellite characterization of Andean races of common bean (Phaseolus vulgaris L.). Theor. Appl. Genet. 2007, 116, 29–43. [Google Scholar] [CrossRef] [PubMed]
- Becerra, V.; Paredes, M.; Rojo, R.; Díaz, M.; Blair, M.W. Microsatellite marker characterization of Chilean common bean (Phaseolus vulgaris L.) germplasm. Crop Sci. 2010, 50, 1932–1941. [Google Scholar] [CrossRef]
- Murube, E.; Beleggia, R.; Pacetti, D.; Nartea, A.; Frascarelli, G.; Lanzavecchia, G.; Bellucci, E.; Nanni, L.; Gioia, T.; Marciello, U.; et al. Characterization of nutritional quality traits of a common bean germplasm collection. Foods 2021, 10, 1572. [Google Scholar] [CrossRef]
- Akond, A.G.M.; Crawford, H.; Berthold, J.; Talukder, Z.I.; Hossain, K. Minerals (Zn, Fe, Ca and Mg) and antinutrient (phytic acid) constituents in common bean. Am. J. Food Technol. 2011, 6, 235. [Google Scholar] [CrossRef]
- Mambrin, R.B.; Sausen, D.; Moura, D.; Carvalho, I.R.; Szareski, V.J. Phosphorus nutrition in beans. Commun. Plant Sci. 2021, 11, 46–56. [Google Scholar]
- Ramírez-Cárdenas, L.; Leonel, A.J.; Costa, N.M.B.; Reis, F.P. Zinc bioavailability in different beans as affected by cultivar type and cooking conditions. Food Res. Int. 2010, 43, 573–581. [Google Scholar] [CrossRef]
- Destro de Lima, J.; Ribeiro Rivadavea, W.; Frehner Kavalco, S.A.; Gonçalves Junior, A.C.; Daniela Lopes, A.; Jaqueline da Silva, G. Chemical and nutritional characterization of bean genotypes (Phaseolus vulgaris L.). AIMS Agric. Food 2021, 6, 932–944. [Google Scholar] [CrossRef]
- Islam, F.M.A.; Basford, K.E.; Jara, C.; Redden, R.J.; Beebe, S. Seed compositional and disease resistance differences among gene pools in cultivated common bean. Genet. Resour. Crop Evol. 2002, 49, 285–293. [Google Scholar] [CrossRef]
- Mojica, L.; de Mejía, E.G. Characterization and Comparison of Protein and Peptide Profiles and their Biological Activities of Improved Common Bean Cultivars (Phaseolus vulgaris L.) from Mexico and Brazil. Plant Foods Hum. Nutr. 2015, 70, 105–112. [Google Scholar] [CrossRef] [PubMed]
- Saboori-Robat, E.; Joshi, J.; Pajak, A.; Solouki, M.; Mohsenpour, M.; Renaud, J.; Marsolais, F. Common Bean (Phaseolus vulgaris L.) Accumulates Most S-Methylcysteine as Its γ-Glutamyl Dipeptide. Plants 2019, 8, 126. [Google Scholar] [CrossRef] [PubMed]
- Nosworthy, M.G.; Neufeld, J.; Frohlich, P.; Young, G.; Malcolmson, L.; House, J.D. Determination of the protein quality of cooked Canadian pulses. Food Sci. Nutr. 2017, 5, 896–903. [Google Scholar] [CrossRef] [PubMed]
- Rombouts, C.; Hemeryck, L.Y.; Van Hecke, T.; De Smet, S.; De Vos, W.H.; Vanhaecke, L. Untargeted metabolomics of colonic digests reveals kynurenine pathway metabolites, dityrosine and 3-dehydroxycarnitine as red versus white meat discriminating metabolites. Sci. Rep. 2017, 7, 42514. [Google Scholar] [CrossRef] [PubMed]
- Reuter, M.; Zamoscik, V.; Plieger, T.; Bravo, R.; Ugartemendia, L.; Rodriguez, A.B.; Kirsch, P. Tryptophan-rich diet is negatively associated with depression and positively linked to social cognition. Nutr. Res. 2021, 85, 14–20. [Google Scholar] [CrossRef] [PubMed]
- Doria, E.; Campion, B.; Sparvoli, F.; Tava, A.; Nielsen, E. Anti-nutrient components and metabolites with health implications in seeds of 10 common bean (Phaseolus vulgaris L. and Phaseolus lunatus L.) landraces cultivated in southern Italy. J. Food Compos. Anal. 2012, 26, 72–80. [Google Scholar] [CrossRef]
- Elango, D.; Rajendran, K.; Van der Laan, L.; Sebastiar, S.; Raigne, J.; Thaiparambil, N.A.; El Haddad, N.; Raja, B.; Wang, W.; Ferela, A.; et al. Raffinose Family Oligosaccharides: Friend or Foe for Human and Plant Health? Front. Plant Sci. 2022, 13, 829118. [Google Scholar] [CrossRef] [PubMed]
- Yan, S.; Liu, Q.; Li, W.; Yan, J.; Fernie, A.R. Raffinose Family Oligosaccharides: Crucial Regulators of Plant Development and Stress Responses. Cr. Rev. Plant Sci. 2022, 41, 286–303. [Google Scholar] [CrossRef]
- Karner, U.; Peterbauer, T.; Raboy, V.; Jones David, A.; Hedley, C.L.; Richter, A. Myo-Inositol and sucrose concentrations affect the accumulation of raffinose family oligosaccharides in seeds. J. Exp. Bot. 2004, 55, 1981–1987. [Google Scholar] [CrossRef]
- Gangola, M.P.; Jaiswal, S.; Kannan, U.; Gaur, P.M.; Båga, M.; Chibbar, R.N. Galactinol synthase enzyme activity influences raffinose family oligosaccharides (RFO) accumulation in developing chickpea (Cicer arietinum L.) seeds. Phytochemistry 2016, 125, 88–98. [Google Scholar] [CrossRef]
- Del Rio, D.; Rodriguez-Mateos, A.; Spencer, J.P.E.; Tognolini, M.; Borges, G.; Crozier, A. Dietary (Poly)phenolics in Human Health: Structures, Bioavailability, and Evidence of Protective Effects Against Chronic Diseases. Antioxid. Redox Sign. 2013, 18, 1818–1892. [Google Scholar] [CrossRef]
- Grosso, G.; Godos, J.; Lamuela-Raventos, R.; Ray, S.; Micek, A.; Pajak, A.; Sciacca, S.; D’Orazio, N.; Del Rio, D.; Galvano, F. A comprehensive meta-analysis on dietary flavonoid and lignan intake and cancer risk: Level of evidence and limitations. Mol. Nutr. Food Res. 2017, 61, 1600930. [Google Scholar] [CrossRef]
- Nardini, M. Phenolic Compounds in Food: Characterization and Health Benefits. Molecules 2022, 27, 783. [Google Scholar] [CrossRef]
- Rodríguez Madrera, R.; Campa Negrillo, A.; Suárez Valles, B.; Ferreira Fernández, J.J. Phenolic Content and Antioxidant Activity in Seeds of Common Bean (Phaseolus vulgaris L.). Foods 2021, 10, 864. [Google Scholar] [CrossRef] [PubMed]
- Espinosa-Alonso, L.G.; Lygin, A.; Widholm, J.M.; Valverde, M.E.; Paredes-Lopez, O. Polyphenols in Wild and Weedy Mexican Common Beans (Phaseolus vulgaris L.). J. Agric. Food Chem. 2006, 54, 4436–4444. [Google Scholar] [CrossRef]
- Valdés, S.T.; Coelho, C.M.M.; Michelluti, D.J.; Tramonte, V.L.C.G. Association of genotype and preparation methods on the antioxidant activity, and antinutrients in common beans (Phaseolus vulgaris L.). LWT Food Sci. Technol. 2011, 44, 2104–2111. [Google Scholar] [CrossRef]
- Singh, S.P. A key for identification of different growth habits of Phaseolus vulgaris L. Annu. Rep. Bean. Improv. Coop. 1982, 25, 92–95. [Google Scholar]
- Sadzawka, A.; Carrasco, M.; Demanet, R.; Flores, H.; Grez, R.; Mora, M.; Neaman, A. Métodos de análisis de tejidos vegetales. Ser. Actas INIA 2007, 40, 140. [Google Scholar]
- AOAC. Official Methods of Analysis, 15th ed.; Association of Analytical Chemists International: Washington, DC, USA, 1995. [Google Scholar]
- Bidlingmeyer, B.A.; Cohen, S.A.; Tarvin, T.L.; Frost, B. A New, Rapid, High-Sensitivity Analysis of Amino Acids in Food Type Samples. J. AOAC Int. 1987, 70, 241–247. [Google Scholar] [CrossRef]
- Bouhlali, E.T.; Ramchoun, M.; Alem, C.; Ghafoor, K.; Ennassir, J.; Zegzouti, Y.F. Functional composition and antioxidant activities of eight Moroccan date fruit varieties (Phoenix dactylifera L.). J. Saudi Soc. Agric. Sci. 2017, 16, 257–264. [Google Scholar] [CrossRef]
- Ahmad-Qasem, M.H.; Cánovas, J.; Barrajón-Catalán, E.; Micol, V.; Cárcel, J.A.; García-Pérez, J.V. Kinetic and compositional study of phenolic extraction from olive leaves (var. Serrana) by using power ultrasound. Innov. Food Sci. Emerg. Technol. 2013, 17, 120–129. [Google Scholar] [CrossRef]
- Rodríguez, L.; Plaza, A.; Méndez, D.; Carrasco, B.; Tellería, F.; Palomo, I.; Fuentes, E. Antioxidant Capacity and Antiplatelet Activity of Aqueous Extracts of Common Bean (Phaseolus vulgaris L.) Obtained with Microwave and Ultrasound Assisted Extraction. Plants 2022, 11, 1179. [Google Scholar] [CrossRef]
- Ou, B.; Chang, T.; Huang, D.; Prior, R.L. Determination of Total Antioxidant Capacity by Oxygen Radical Absorbance Capacity (ORAC) Using Fluorescein as the Fluorescence Probe: First Action 2012.23. J. AOAC Int. 2013, 96, 1372–1376. [Google Scholar] [CrossRef]
- Wickham, H.; François, R.; Henry, L.; Müller, K.; Wickham, M.H. Package ‘dplyr’. A Grammar of Data Manipulation, R package version 8; 2019. Available online: https://cran.r-hub.io/web/packages/dplyr/dplyr.pdf (accessed on 15 December 2023).
- Wei, T.; Simko, V.; Levy, M.; Xie, Y.; Jin, Y.; Zemla, J. Package ‘corrplot’. Statistician 2017, 56, e24. [Google Scholar]
- De Mendiburu, F.; de Mendiburu, M.F. Package ‘agricolae’, R Package Version 1; 2019. Available online: https://cran.r-project.org/web/packages/agricolae/index.html (accessed on 15 December 2023).
- Kassambara, A.; Mundt, F. Package ‘factoextra’. Extract and Visualize the Results of Multivariate Data Analyses, R Package Version 1.0.7; 2017. Available online: https://CRAN.R-project.org/package=factoextra (accessed on 15 December 2023).
- Wickham, H. ggplot2. Wiley Interdiscip. Rev. Comput. Stat. 2011, 3, 180–185. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Category | Trait | Unit | Mean | Sd | Min | Max | Significance |
|---|---|---|---|---|---|---|---|
| Sugars | Sucrose | g/100 g seed | 2.61 | 0.85 | 1.11 | 4.75 | * |
| RFO | mmol/100 g seed | 4.79 | 0.43 | 3.92 | 5.45 | * | |
| Antioxidant activity | TPC | mg GA/100 g seed | 111.38 | 44.55 | 49.18 | 218.11 | * |
| ORAC | umol eq trolox/100 g extract | 12,821.04 | 5936.16 | 4544.91 | 23,045.01 | * | |
| Protein content | SP | mg/mL | 5.28 | 2.21 | 1.77 | 10.01 | * |
| TP | g/100 g seed | 24.19 | 2.60 | 20.31 | 30.35 | * | |
| Macro-elements | P | g/kg | 4.67 | 0.84 | 3.43 | 7.53 | * |
| K | g/kg | 16.13 | 1.45 | 13.43 | 19.80 | * | |
| Ca | g/kg | 1.64 | 0.48 | 0.87 | 2.93 | * | |
| Mg | g/kg | 2.03 | 0.15 | 1.73 | 2.43 | * | |
| Total | g/kg | 24.47 | 2.31 | 21.10 | 31.50 | * | |
| Micro-elements | Mn | mg/kg | 13.27 | 2.53 | 10.33 | 19.00 | * |
| Zn | mg/kg | 38.13 | 6.71 | 23.00 | 52.67 | * | |
| Cu | mg/kg | 10.25 | 1.42 | 8.00 | 13.67 | * | |
| Fe | mg/kg | 71.76 | 16.66 | 48.67 | 109.00 | * | |
| B | mg/kg | 17.32 | 2.74 | 13.00 | 26.33 | * | |
| Total | mg/kg | 150.74 | 22.62 | 110.33 | 197.00 | * | |
| EAAs | Gly.His | mg/g protein | 61.19 | 4.20 | 49.92 | 68.77 | * |
| Thr | mg/g protein | 35.45 | 2.61 | 30.91 | 40.71 | * | |
| Val | mg/g protein | 39.59 | 2.79 | 34.48 | 45.72 | * | |
| Met | mg/g protein | 7.34 | 1.94 | 3.32 | 11.84 | * | |
| Ile | mg/g protein | 35.75 | 3.13 | 30.63 | 41.82 | * | |
| Leu | mg/g protein | 61.08 | 4.30 | 53.64 | 68.79 | * | |
| Phe | mg/g protein | 42.86 | 5.29 | 35.91 | 59.40 | * | |
| Lys | mg/g protein | 50.99 | 4.93 | 43.78 | 61.62 | * | |
| Total | mg/g protein | 334.24 | 23.23 | 293.27 | 367.94 | * | |
| NEAAs | Asp | mg/g protein | 90.22 | 7.02 | 75.35 | 101.42 | * |
| Glu | mg/g protein | 120.06 | 8.34 | 98.63 | 134.13 | * | |
| 4Hyp | mg/g protein | 1.79 | 0.62 | 0.75 | 3.30 | * | |
| Ser | mg/g protein | 47.40 | 3.99 | 40.20 | 59.44 | * | |
| Arg | mg/g protein | 56.09 | 6.41 | 46.67 | 70.01 | * | |
| Ala | mg/g protein | 37.25 | 3.19 | 31.35 | 43.09 | * | |
| Pro | mg/g protein | 31.56 | 2.70 | 27.65 | 37.50 | * | |
| Tyr | mg/g protein | 26.33 | 4.11 | 18.76 | 34.36 | * | |
| Total | mg/g protein | 410.70 | 24.38 | 369.57 | 452.15 | * |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Márquez, K.; Arriagada, O.; Pérez-Díaz, R.; Cabeza, R.A.; Plaza, A.; Arévalo, B.; Meisel, L.A.; Ojeda, D.; Silva, H.; Schwember, A.R.; et al. Nutritional Characterization of Chilean Landraces of Common Bean. Plants 2024, 13, 817. https://doi.org/10.3390/plants13060817
Márquez K, Arriagada O, Pérez-Díaz R, Cabeza RA, Plaza A, Arévalo B, Meisel LA, Ojeda D, Silva H, Schwember AR, et al. Nutritional Characterization of Chilean Landraces of Common Bean. Plants. 2024; 13(6):817. https://doi.org/10.3390/plants13060817
Chicago/Turabian StyleMárquez, Katherine, Osvin Arriagada, Ricardo Pérez-Díaz, Ricardo A. Cabeza, Andrea Plaza, Bárbara Arévalo, Lee A. Meisel, Daniela Ojeda, Herman Silva, Andrés R. Schwember, and et al. 2024. "Nutritional Characterization of Chilean Landraces of Common Bean" Plants 13, no. 6: 817. https://doi.org/10.3390/plants13060817