

The Impact of Phenological Gaps on Leaf Characteristics and Foliage Dynamics of an Understory Dwarf Bamboo, Sasa kurilensis
Abstract
:1. Introduction
2. Results
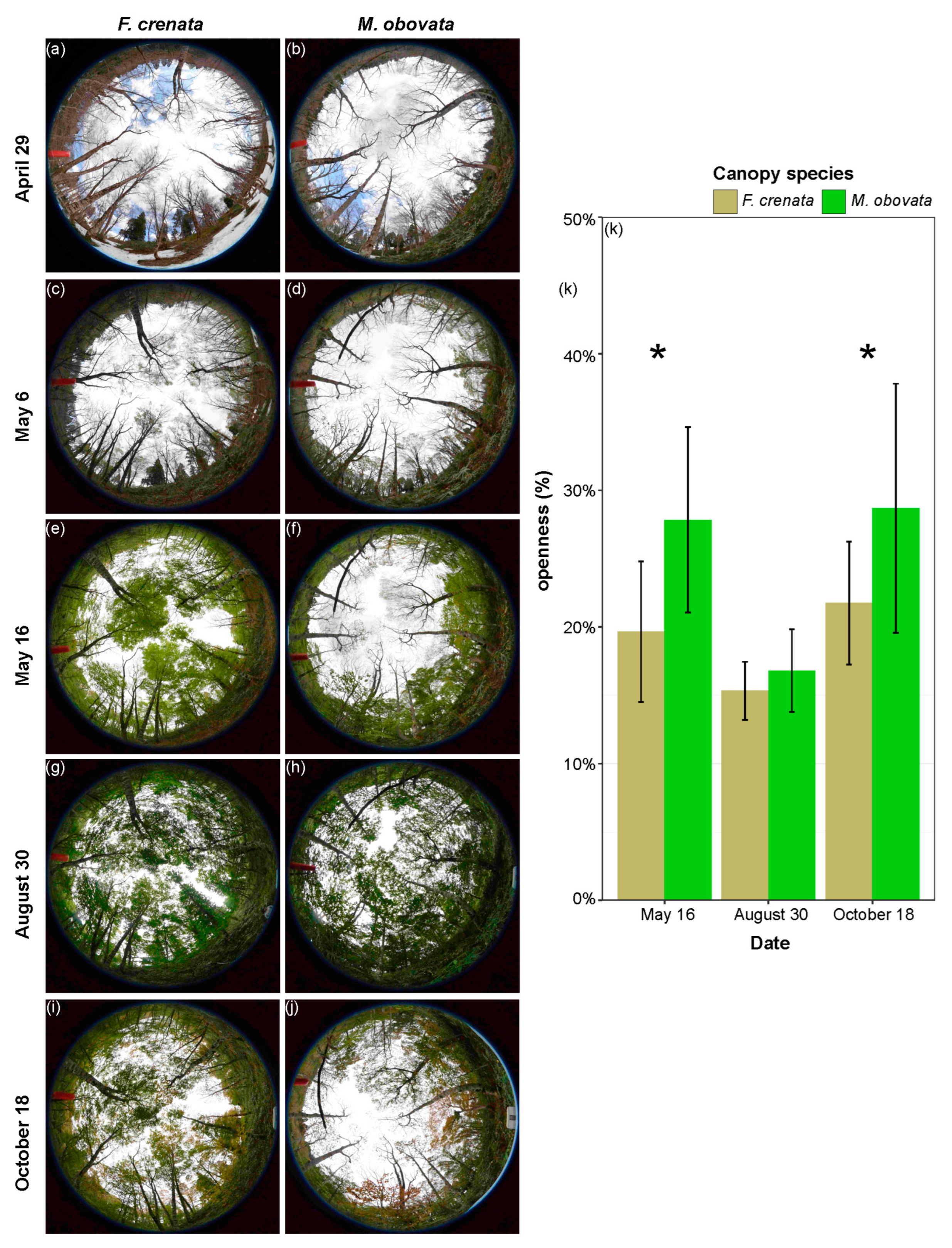
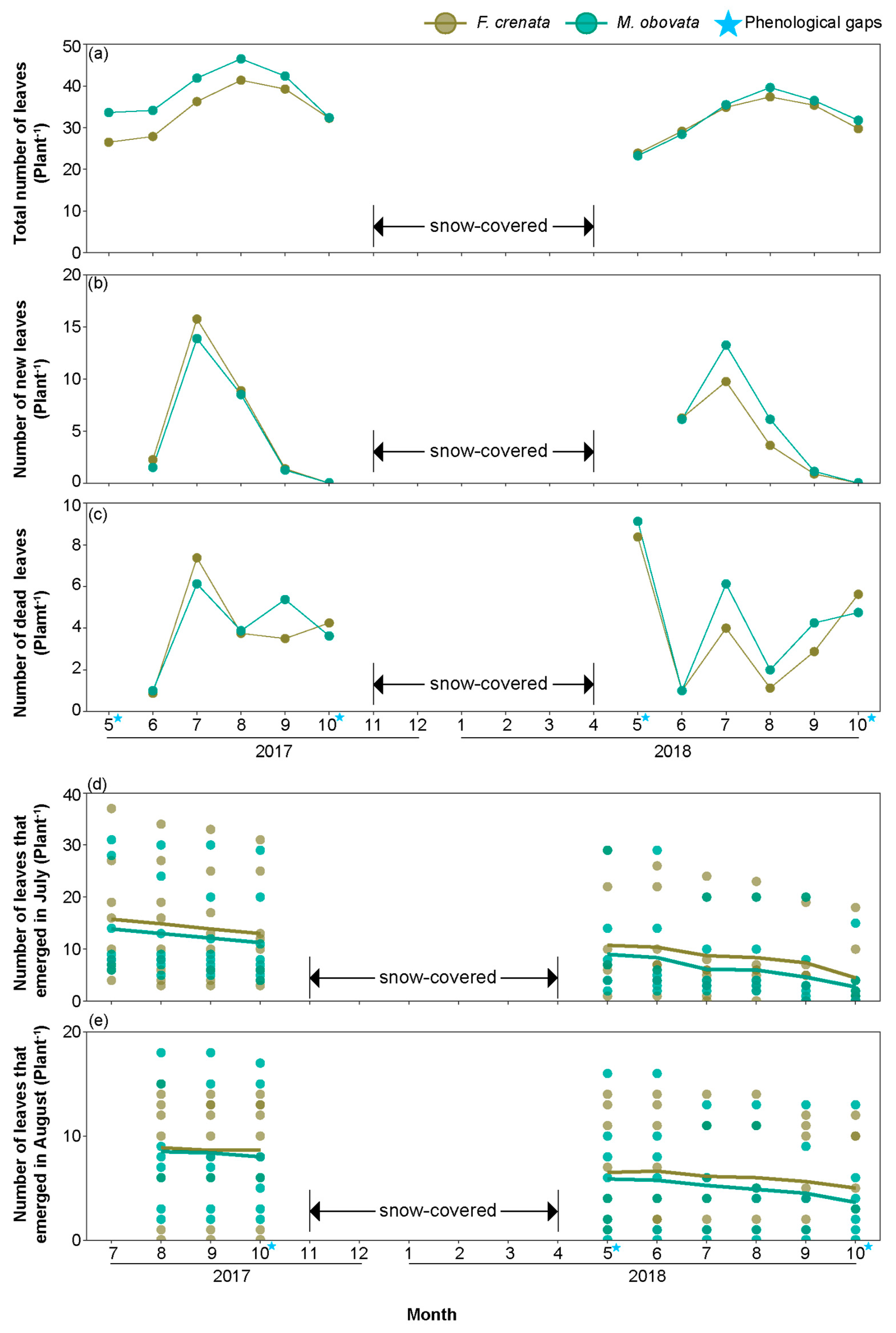
2.1. Phenological Gaps and Microhabitat Conditions under Different Canopy Species
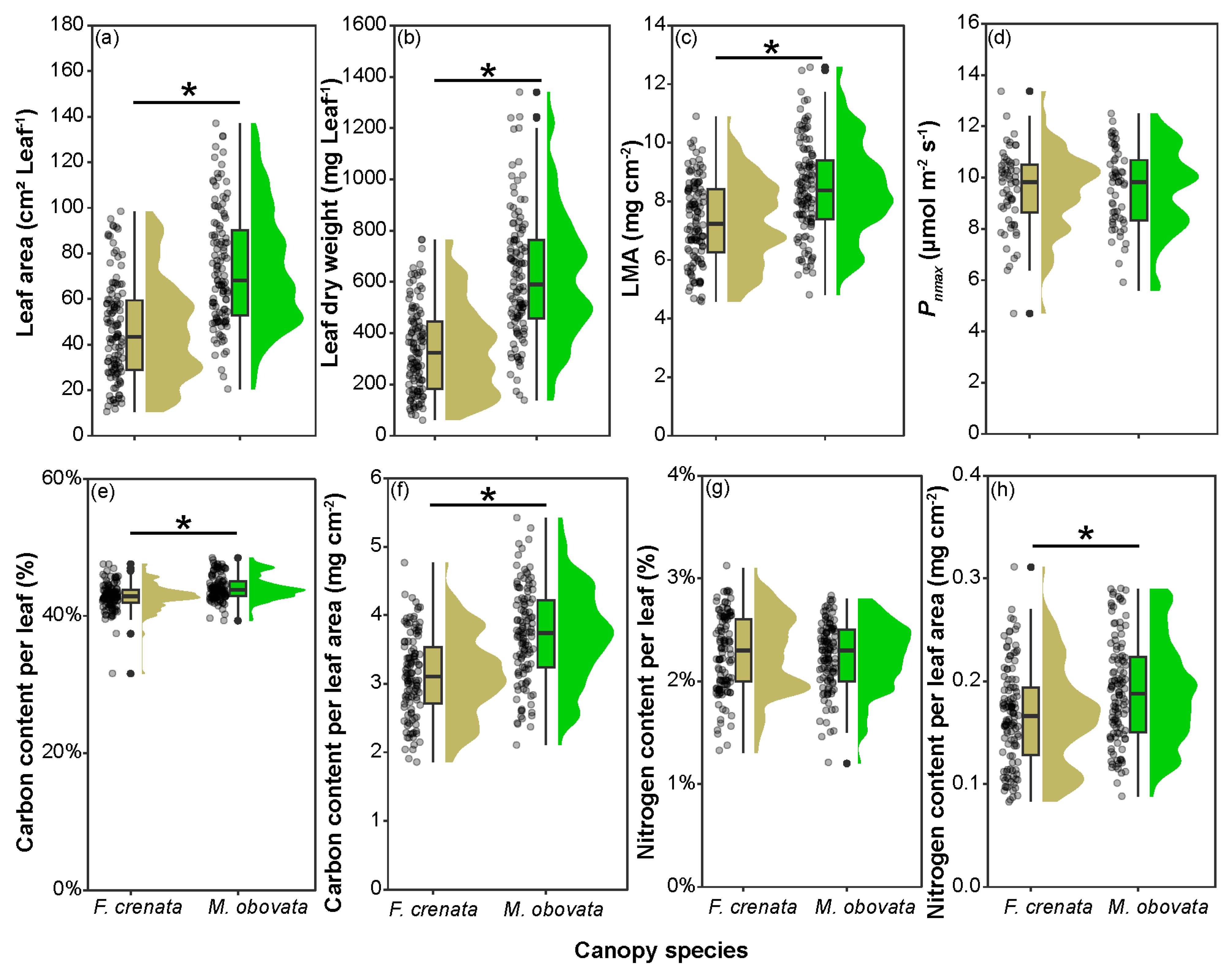
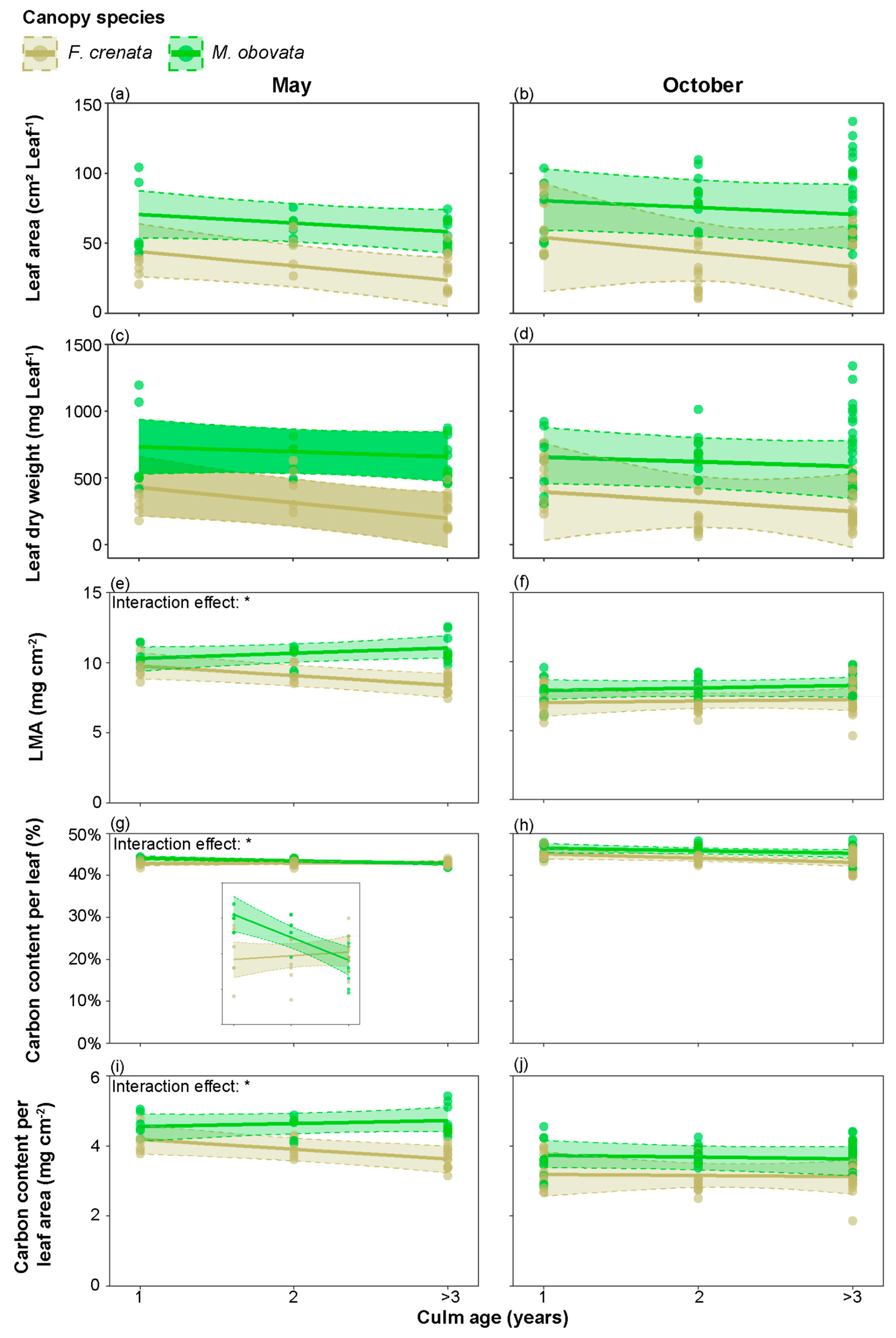
2.2. Leaf Characteristics under Different Canopy Species
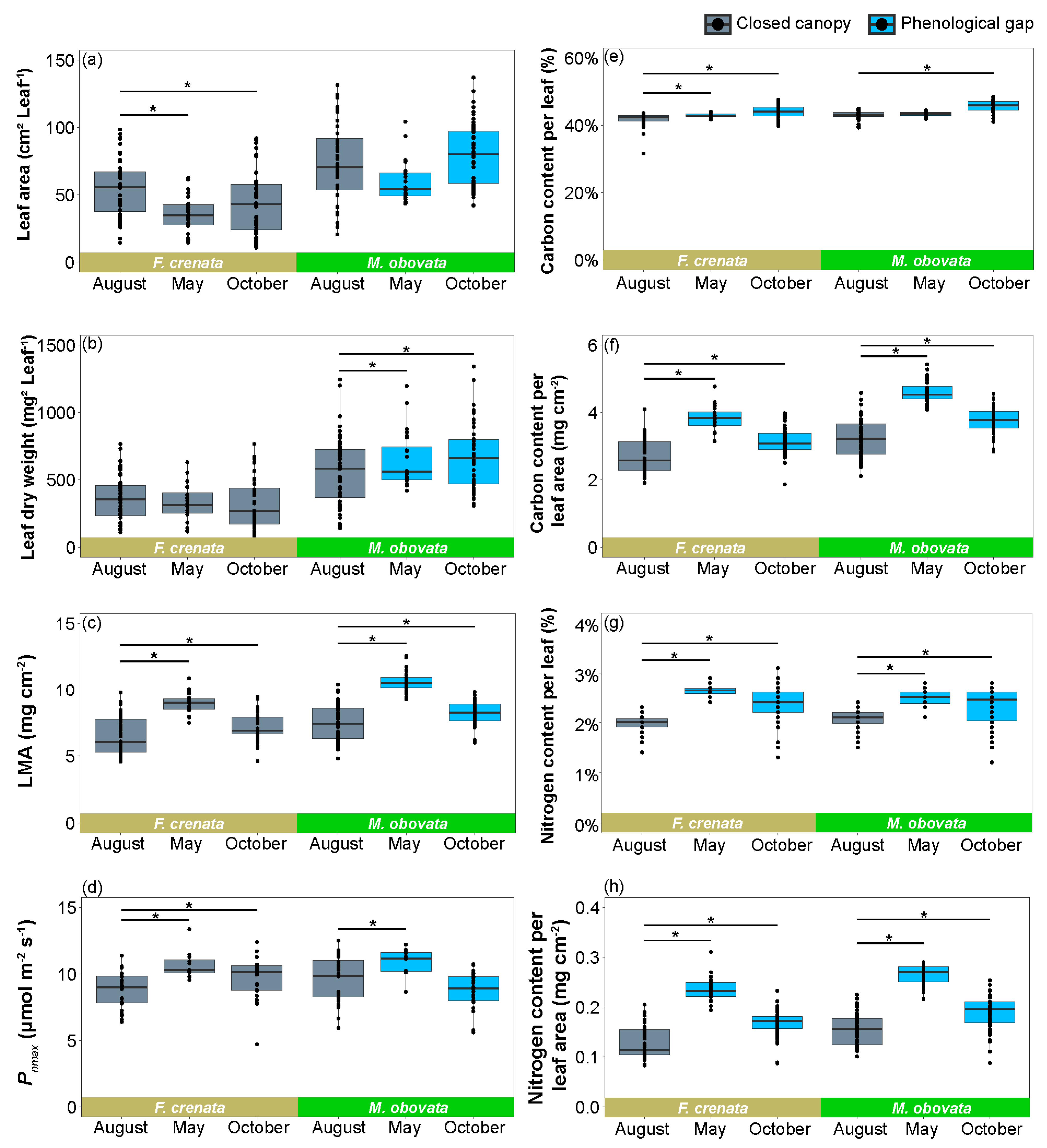
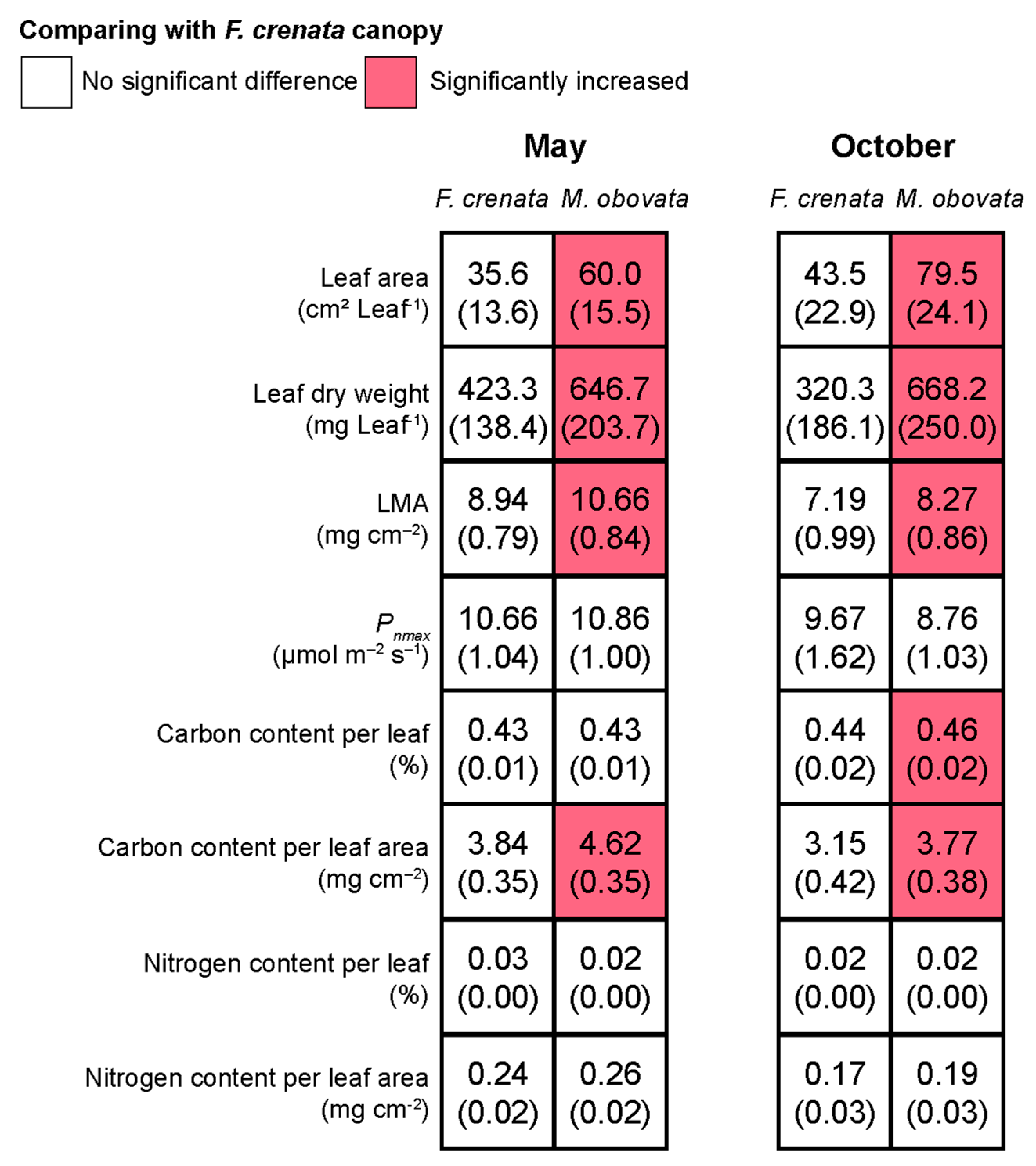
2.3. The Influence of Phenological Gaps on Leaf Characteristics
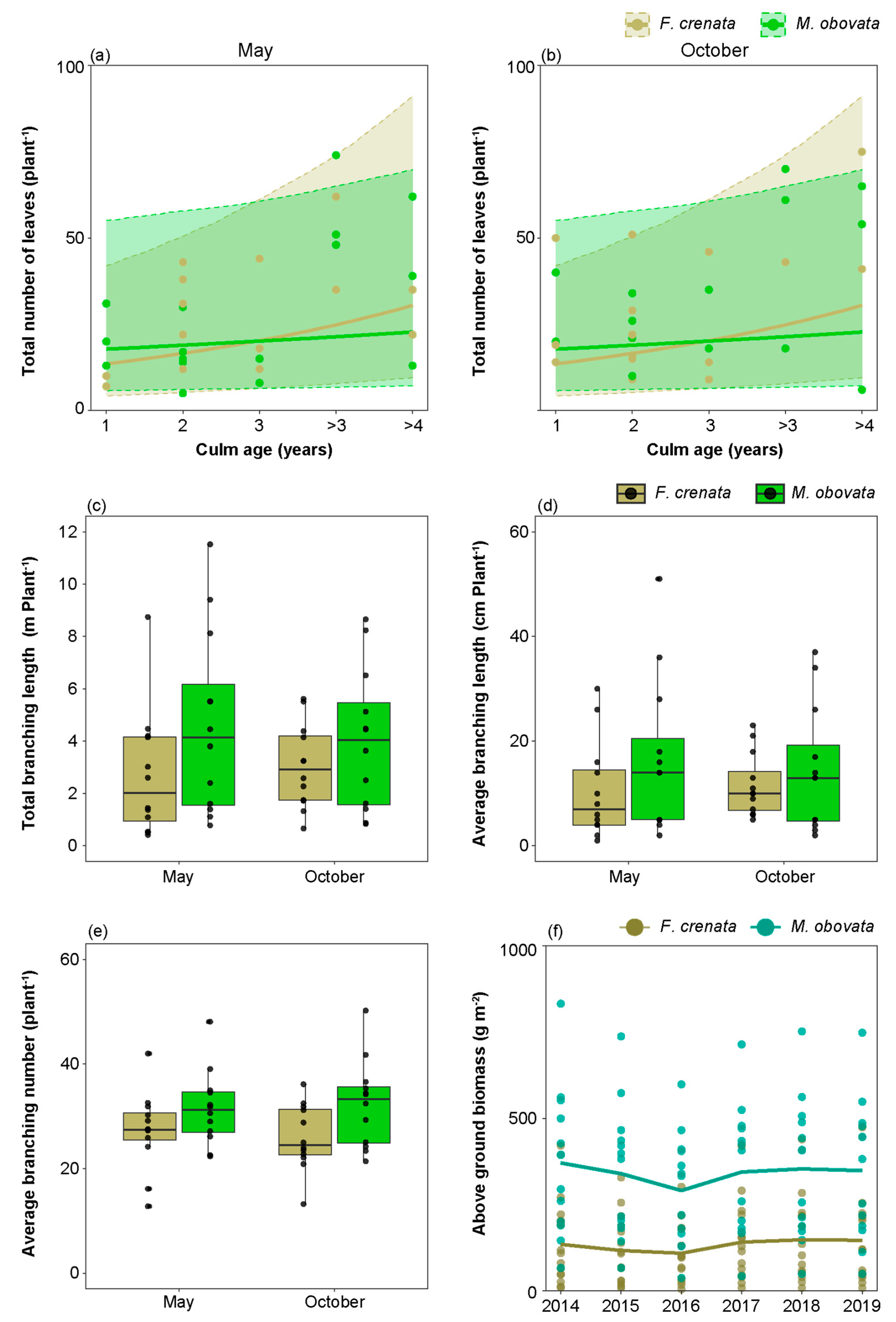
2.4. Foliage Dynamics, Branching Characteristics and Above Ground Biomass under Different Canopy Species
3. Discussion
3.1. Seasonal Variations in the Leaf Characteristics of S. kurilensis
3.2. The Impact of the Phenological Gap on the Leaf Characteristics of S. kurilensise
3.3. The Impact of Phenological Gaps on the Foliage Dynamics and Branching Characteristics of S. kurilensis
4. Materials and Methods
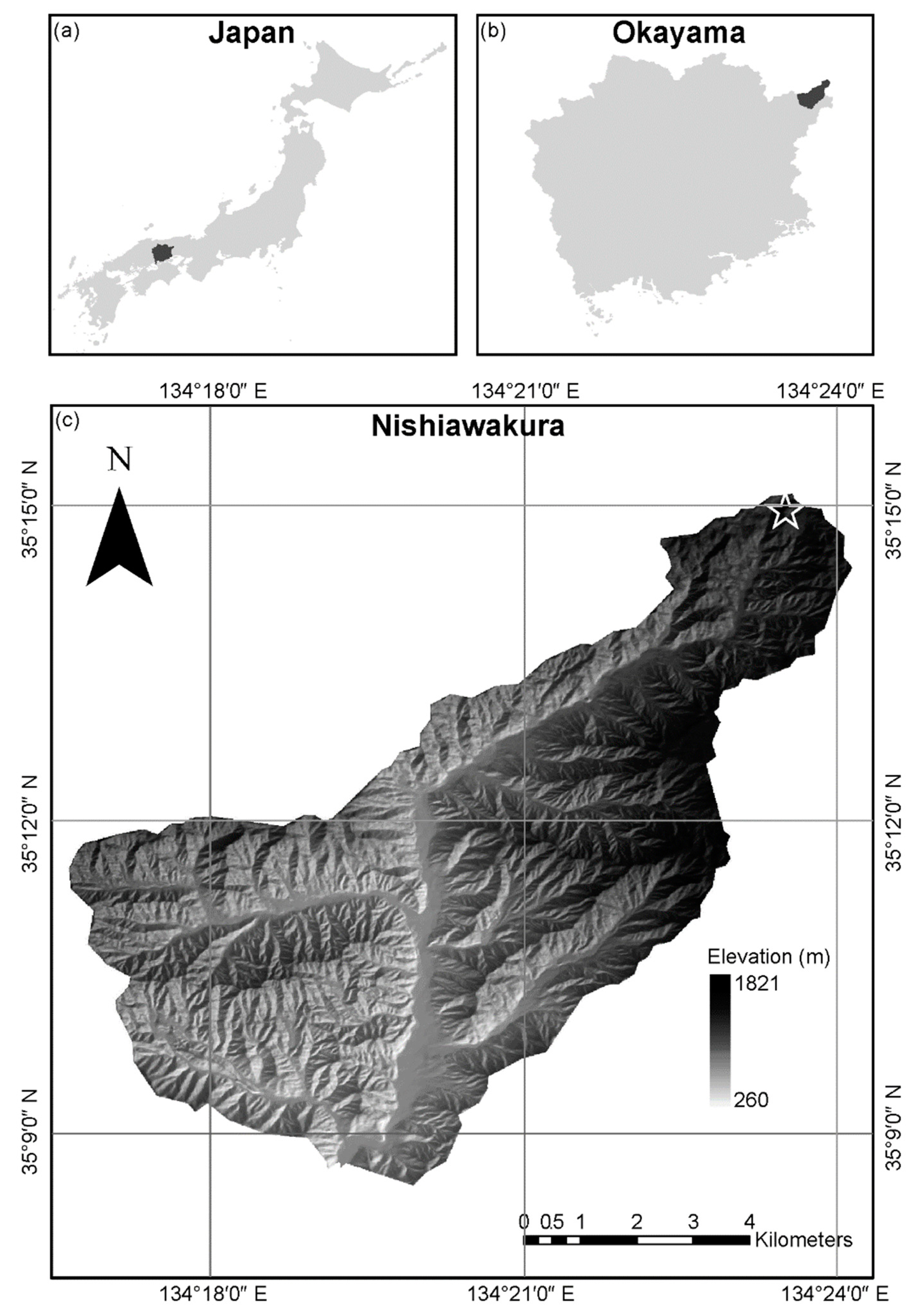
4.1. Study Area
4.2. Establishment of Study Plot
4.3. Leaf Characteristics
4.4. Leaf Maximum Net Photosynthetic Rate
4.5. Foliage Dynamics and Branching Characteristics
4.6. Estimation of Above Ground Biomass
4.7. Light Environment and Microhabitat Conditions
4.8. Model Analysis
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Franklin, J.F.; Maeda, T.; Ohsumi, Y.; Matsui, M.; Yagi, H.; Hawk, G.M. Subalpine coniferous forests of central Honshu, Japan. Ecol. Monogr. 1979, 49, 311–334. [Google Scholar] [CrossRef]
- Nakashizuka, T.; Numata, M. Regeneration process of climax beech forests: I. Structure of a beech forest with the undergrowth of Sasa. Jpn. J. Ecol. 1982, 32, 57–67. [Google Scholar] [CrossRef]
- Wada, N. Dwarf bamboos affect the regeneration of zoochorous trees by providing habitats to acorn-feeding rodents. Oecologia 1993, 94, 403–407. [Google Scholar] [CrossRef]
- Noguchi, M.; Yoshida, T. Factors influencing the distribution of two co-occurring dwarf bamboo species (Sasa kurilensis and S. senanensis) in a conifer-broadleaved mixed stand in northern Hokkaido. Ecol. Res. 2005, 20, 25–30. [Google Scholar] [CrossRef]
- Kudo, G.; Amagai, Y.; Hoshino, B.; Kaneko, M. Invasion of dwarf bamboo into alpine snow-meadows in northern Japan: Pattern of expansion and impact on species diversity. Ecol. Evol. 2011, 1, 85–96. [Google Scholar] [CrossRef]
- Tsunoda, Y.; Furukawa, S.; Mizunaga, H. How does the longevity of Sasa kurilensis ramets respond to a light gradient? An analysis of ontogenetic changes to hydraulic resistance and carbon budget within a ramet. Ecol. Res. 2017, 32, 117–128. [Google Scholar] [CrossRef]
- Matsuo, A.; Suyama, Y.; Makita, A. The relationship between rhizome habits and spatial distribution pattern of genets in two dwarf bamboos. Jpn. J. Ecol. 2010, 60, 81–88. [Google Scholar] [CrossRef]
- Itô, H.; Hino, T. Dwarf bamboo as an ecological filter for forest regeneration. Ecol. Res. 2007, 22, 706–711. [Google Scholar] [CrossRef]
- Hirobe, M.; Miyamoto, S.; Sakamoto, K.; Kondo, J.; Otoda, T.; Akaji, Y.; Yamanaka, N. The spatial distributions of understory trees in relation to dwarf bamboo cover in a cool-temperate deciduous broadleaf forest in Japan. J. For. Res. 2017, 20, 357–362. [Google Scholar] [CrossRef]
- Fukuzawa, K.; Shibata, H.; Takagi, K.; Satoh, F.; Koike, T.; Sasa, K. Vertical distribution and seasonal pattern of fine-root dynamics in a cool–temperate forest in northern Japan: Implication of the understory vegetation, Sasa dwarf bamboo. Ecol. Res. 2007, 22, 485–495. [Google Scholar] [CrossRef]
- Fukuzawa, K.; Shibata, H.; Takagi, K.; Nomura, M.; Kurima, N.; Fukazawa, T.; Satoh, F.; Sasa, K. Effects of clear-cutting on nitrogen leaching and fine root dynamics in a cool-temperate forested watershed in northern Japan. For. Ecol. Manag. 2006, 225, 257–261. [Google Scholar] [CrossRef]
- Toyooka, H.; Sato, A.; Ishizuka, M. The Influence of light Intensity in Sasa senanensis growth. Trans. Meet. Hokkaido Branch Jpn. For. Soc. 1982, 30, 139–141. (In Japanese) [Google Scholar] [CrossRef]
- Tobita, H.; Utsugi, H.; Kitao, M.; Kayama, M.; Maruyama, Y. Photosynthetic active of Sasa senanensis under gap formed from typhoon. Trans. Meet. Hokkaido Branch Jpn. For. Soc. 2006, 54, 70–72. (In Japanese) [Google Scholar] [CrossRef]
- Ala, T.; Sakamoto, K.; Miki, N.; Hirobe, M.; Yoshikawa, K. Leaf photosynthetic characteristics of Pleioblastus pubescens Nakai growing under different light conditions. J. Jpn. Soc. Reveg. Technol. 2008, 34, 636–640. (In Japanese) [Google Scholar] [CrossRef]
- Ala, T.; Sakamoto, K.; Miki, N.; Hirobe, M.; Yoshikawa, K. Seasonal changes of ecological characteristics on culms and leaves of Pleioblastus pubescens Nakai growing under different light conditions. J. Jpn. Soc. Reveg. Technol. 2008, 34, 524–533. (In Japanese) [Google Scholar] [CrossRef]
- Wu, C.; Ryota, T.; Kyohei, F.; Itsuka, H.; Yasuaki, A.; Muneto, H.; Keiji, S. Environmental factors affecting community structure of Sasa kurilensis in the understory of a beech forest. J. Jpn. Soc. Reveg. Technol. 2019, 45, 103–108. (In Japanese) [Google Scholar] [CrossRef]
- Kobayashi, T.; Muraoka, H.; Shimano, K. Photosynthesis and biomass allocation of beech (Fagus crenata) and dwarf-bamboo (Sasa kurilensis) in response to contrasting light regimes in a Japan sea-type beech forest. J. For. Res. 2017, 5, 103–107. [Google Scholar] [CrossRef]
- Tomita, M.; Seiwa, K. Influence of canopy tree phenology on understorey populations ofFagus crenata. J. Veg. Sci. 2004, 15, 379–388. [Google Scholar] [CrossRef]
- Komiyama, A.; Kato, S.; Teranishi, M. Differential overstory leaf flushing contributes to the formation of a patchy understory. J. For. Res. 2001, 6, 163–171. [Google Scholar] [CrossRef]
- Niinemets, U.; Portsmuth, A.; Tobias, M. Leaf shape and venation pattern alter the support investments within leaf lamina in temperate species: A neglected source of leaf physiological differentiation? Funct. Ecol. 2007, 21, 28–40. [Google Scholar] [CrossRef]
- Niinemets, U.; Kull, O.; Tenhunen, J.D. Variability in leaf morphology and chemical composition as a function of canopy light environment in coexisting deciduous trees. Int. J. Plant Sci. 1999, 160, 837–848. [Google Scholar] [CrossRef]
- Xu, F.; Guo, W.; Xu, W.; Wei, Y.; Wang, R. Leaf morphology correlates with water and light availability: What consequences for simple and compound leaves? Prog. Nat. Sci. 2009, 19, 1789–1798. [Google Scholar] [CrossRef]
- Wang, J.; Lu, W.; Tong, Y.; Yang, Q. Leaf morphology, photosynthetic performance, chlorophyll fluorescence, stomatal development of lettuce (lactuca sativa L.) exposed to different ratios of red light to blue light. Front. Plant Sci. 2016, 7, 250. [Google Scholar] [CrossRef] [PubMed]
- Hogewoning, S.W.; Trouwborst, G.; Maljaars, H.; Poorter, H.; van Ieperen, W.; Harbinson, J. Blue light dose-responses of leaf photosynthesis, morphology, and chemical composition of Cucumis sativus grown under different combinations of red and blue light. J. Exp. Bot. 2010, 61, 3107–3117. [Google Scholar] [CrossRef] [PubMed]
- Morin, X.; Lechowicz, M.J.; Augspurger, C.; O’keefe, J.; Viner, D.; Chuine, I. Leaf phenology in 22 North American tree species during the 21st century. Glob. Chang. Biol. 2009, 15, 961–975. [Google Scholar] [CrossRef]
- Long, S.; Humphries, S.; Falkowski, P. Photoinhibition of Photosynthesis in Nature. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1994, 45, 633–662. [Google Scholar] [CrossRef]
- Kitao, M.; Lei, T.T.; Koike, T.; Tobita, H.; Maruyama, Y. Tradeoff between shade adaptation and mitigation of photoinhibition in leaves of Quercus mongolica and Acer mono acclimated to deep shade. Tree Physiol. 2006, 26, 441–448. [Google Scholar] [CrossRef] [PubMed]
- Evans, J.R.; Poorter, H. Photosynthetic acclimation of plants to growth irradiance: The relative importance of specific leaf area and nitrogen partitioning in maximizing carbon gain. Plant Cell Environ. 2001, 24, 755–767. [Google Scholar] [CrossRef]
- Crawley, M.J. Plant Ecology; Blackwell Scientific Publications: Hoboken, NJ, USA, 1986. [Google Scholar]
- Winkler, D.E.; Amagai, Y.; Huxman, T.E.; Kaneko, M.; Kudo, G. Seasonal dry-down rates and high stress tolerance promote bamboo invasion above and below treeline. Plant Ecol. 2016, 217, 1219–1234. [Google Scholar] [CrossRef]
- James, S.A.; Bell, D.T. Influence of light availability on leaf structure and growth of two Eucalyptus globulus ssp. globulus provenances. Tree Physiol. 2000, 20, 1007–1018. [Google Scholar] [CrossRef]
- Chapin, F.S., III; Kedrowski, R.A. Seasonal changes in nitrogen and phosphorus fractions and autumn retranslocation in evergreen and deciduous taiga trees. Ecology 1983, 64, 376–391. [Google Scholar] [CrossRef]
- Kleunen, M.V.; Fischer, M.; Schmid, B. Effects of intraspecific competition on size variation and reproductive allocation in a clonal plant. Oikos 2001, 94, 515–524. [Google Scholar] [CrossRef]
- Poorter, L. Light-dependent changes in biomass allocation and their importance for growth of rain forest tree species. Funct. Ecol. 2001, 15, 113–123. [Google Scholar] [CrossRef]
- Pearcy, R.W.; Muraoka, H.; Valladares, F. Crown architecture in sun and shade environments: Assessing function and trade-offs with a three-dimensional simulation model. New Phytol. 2005, 166, 791–800. [Google Scholar] [CrossRef]
- Stoll, P.; Prati, D. Intraspecific aggregation alters competitive interactions in experimental plant communities. Ecology 2001, 82, 319–327. [Google Scholar] [CrossRef]
- Adler, P.B.; Smull, D.; Beard, K.H.; Choi, R.T.; Furniss, T.; Kulmatiski, A.; Meiners, J.M.; Tredennick, A.T.; Veblen, K.E. Competition and coexistence in plant communities: Intraspecific competition is stronger than interspecific competition. Ecol. Lett. 2018, 21, 1319–1329. [Google Scholar] [CrossRef]
- Diemer, M. Life span and dynamics of leaves of herbaceous perennials in high-elevation environments: ‘news from the elephant’s leg’. Funct. Ecol. 1998, 12, 413–425. [Google Scholar] [CrossRef]
- Ariya, U.; Hamano, K.-Y.; Makimoto, T.; Kinoshita, S.; Akaji, Y.; Miyazaki, Y.; Hirobe, M.; Sakamoto, K. Temporal and spatial dynamics of an old-growth beech forest in western Japan. J. For. Res. 2015, 21, 73–83. [Google Scholar] [CrossRef]
- Akaji, Y.; Fujiyoshi, K.; Wu, C.; Hattori, I.; Hirobe, M.; Sakamoto, K. Survival and recruitment of Sasa kurilensis culms in response to local light conditions in a cool temperate forest. J. For. Res. 2019, 24, 365–370. [Google Scholar] [CrossRef]
- Wu, C.; Bai, Y.; Cao, Z.; Xu, J.; Xie, Y.; Zheng, H.; Jiang, J.; Mu, C.; Cheng, W.; Fang, H.; et al. Plasticity in the morphology of growing bamboo: A bayesian analysis of exogenous treatment effects on plant height, internode length, and internode numbers. Plants 2023, 12, 1713. [Google Scholar] [CrossRef]
- Feehan, D.M.; Mahmud, A.S. Quantifying population contact patterns in the United States during the COVID-19 pandemic. Nat. Commun. 2021, 12, 893. [Google Scholar] [CrossRef] [PubMed]
- Harrington, S.M.; Wishingrad, V.; Thomson, R.C. Properties of markov chain monte carlo performance across many empirical alignments. Mol. Biol. Evol. 2021, 38, 1627–1640. [Google Scholar] [CrossRef] [PubMed]
- Bürkner, P.C. Advanced Bayesian multilevel modeling with the R package brms. arXiv 2017, arXiv:1705.11123. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Canopy Species | Topography | Soil Nutrient Content | |||
|---|---|---|---|---|---|
| Slope (°) | Convexity (m) | Water (%) | Carbon (%) | Nitrogen (%) | |
| F. crenata | 26.74 ± 7.29 | −0.13 ± 0.34 | 65.28 ± 1.69 | 16.46 ± 2.27 | 1.10 ± 0.15 |
| M. obovata | 30.49 ± 4.69 | 0.06 ± 0.33 | 68.29 ± 3.13 | 18.62 ± 3.54 | 1.24 ± 0.21 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, C.; Tanaka, R.; Fujiyoshi, K.; Akaji, Y.; Hirobe, M.; Miki, N.; Li, J.; Sakamoto, K.; Gao, J. The Impact of Phenological Gaps on Leaf Characteristics and Foliage Dynamics of an Understory Dwarf Bamboo, Sasa kurilensis. Plants 2024, 13, 719. https://doi.org/10.3390/plants13050719
Wu C, Tanaka R, Fujiyoshi K, Akaji Y, Hirobe M, Miki N, Li J, Sakamoto K, Gao J. The Impact of Phenological Gaps on Leaf Characteristics and Foliage Dynamics of an Understory Dwarf Bamboo, Sasa kurilensis. Plants. 2024; 13(5):719. https://doi.org/10.3390/plants13050719
Chicago/Turabian StyleWu, Chongyang, Ryota Tanaka, Kyohei Fujiyoshi, Yasuaki Akaji, Muneto Hirobe, Naoko Miki, Juan Li, Keiji Sakamoto, and Jian Gao. 2024. "The Impact of Phenological Gaps on Leaf Characteristics and Foliage Dynamics of an Understory Dwarf Bamboo, Sasa kurilensis" Plants 13, no. 5: 719. https://doi.org/10.3390/plants13050719