
Hepatoprotective Effects of Four Brazilian Savanna Species on Acetaminophen-Induced Hepatotoxicity in HepG2 Cells

 , ,
, ,  , ,
, ,  and
and
Abstract
:1. Introduction
2. Results
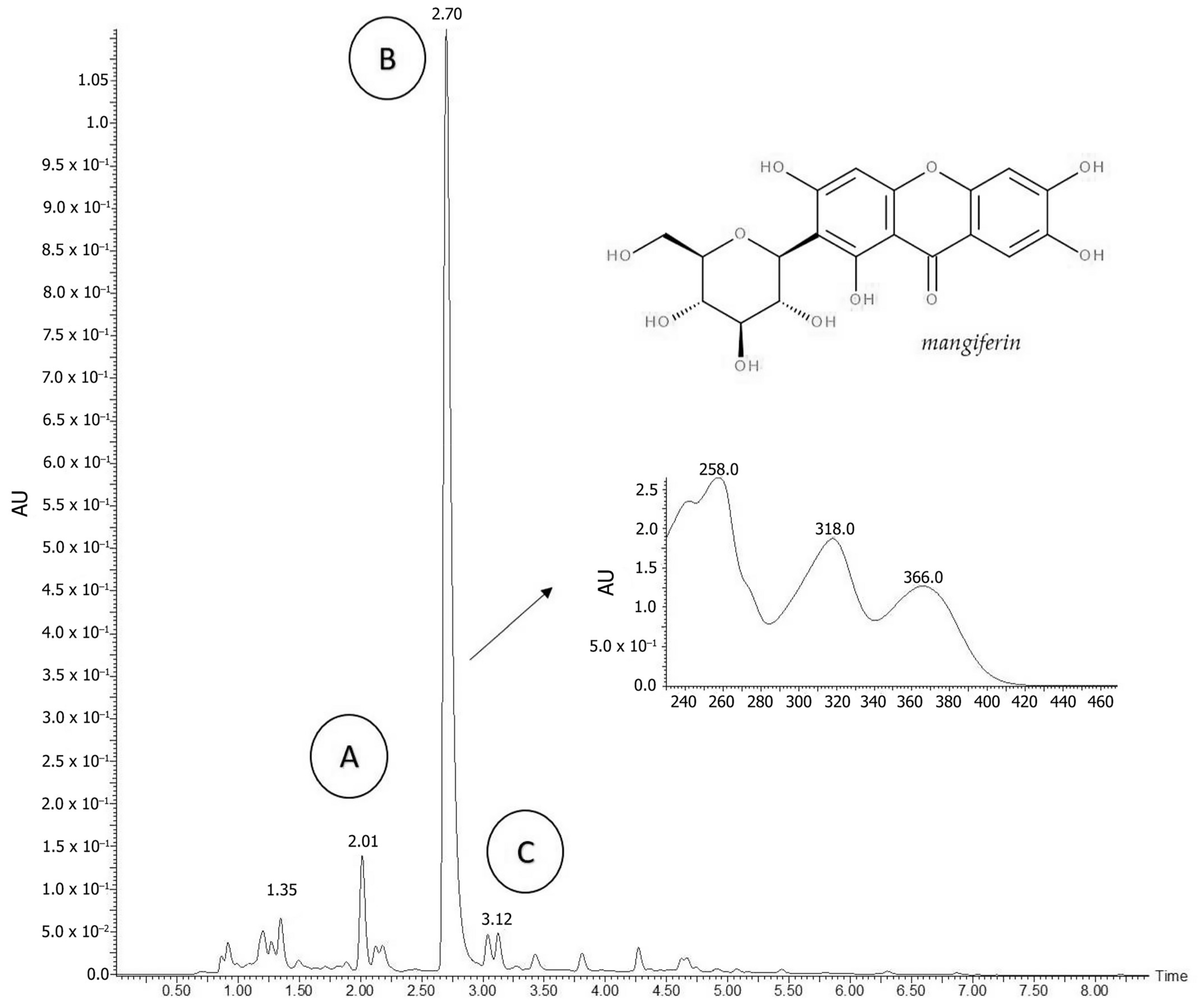
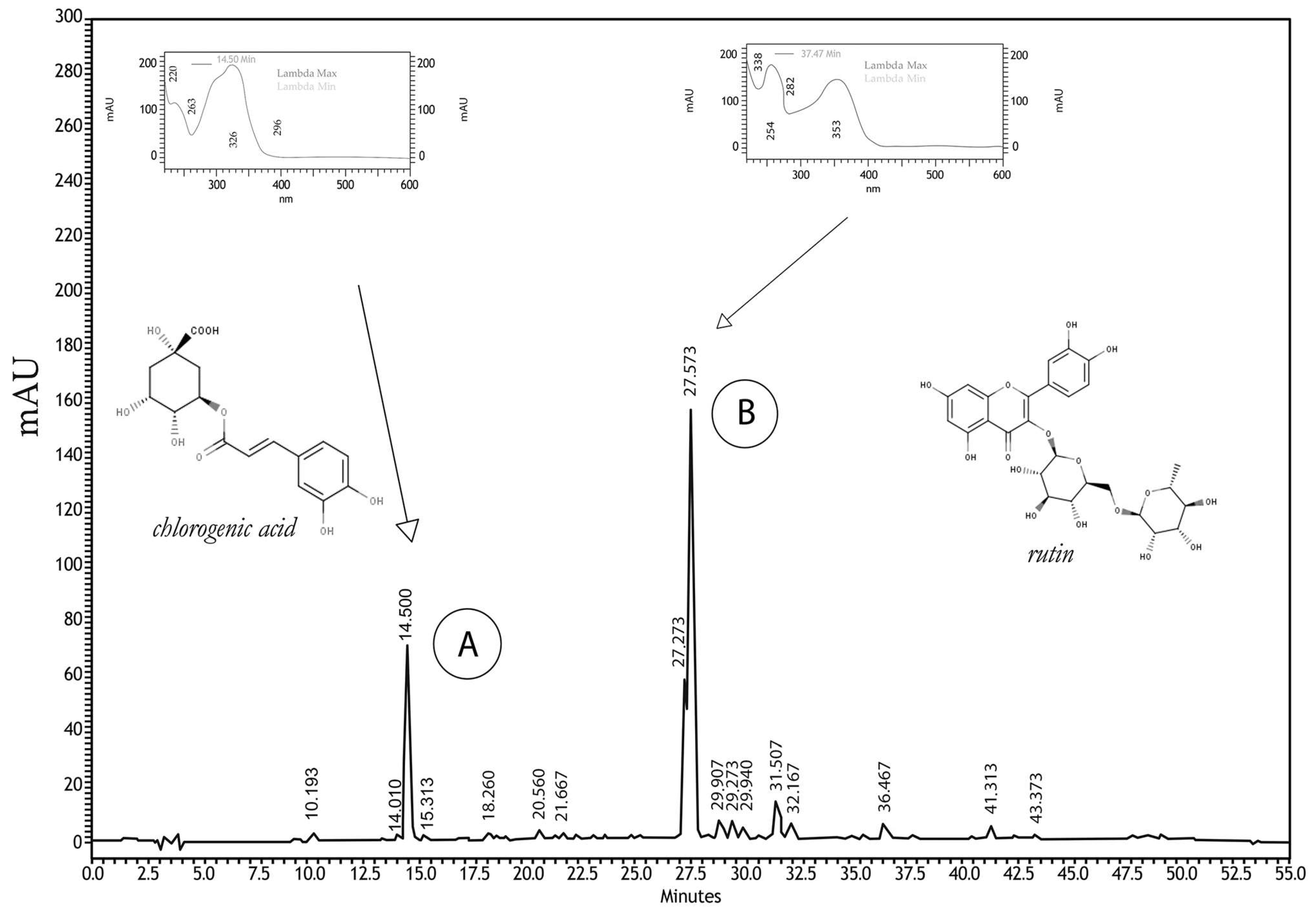
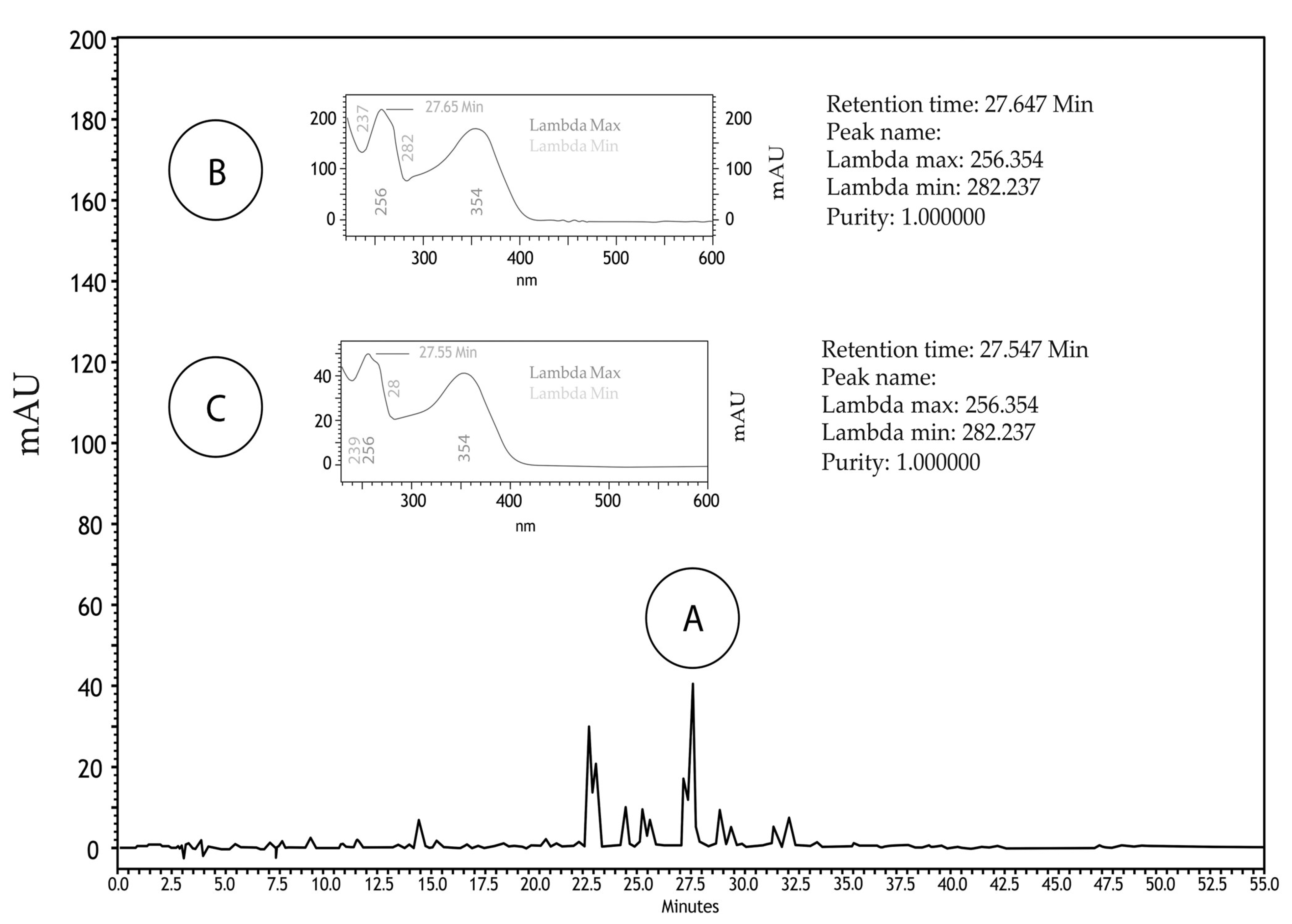
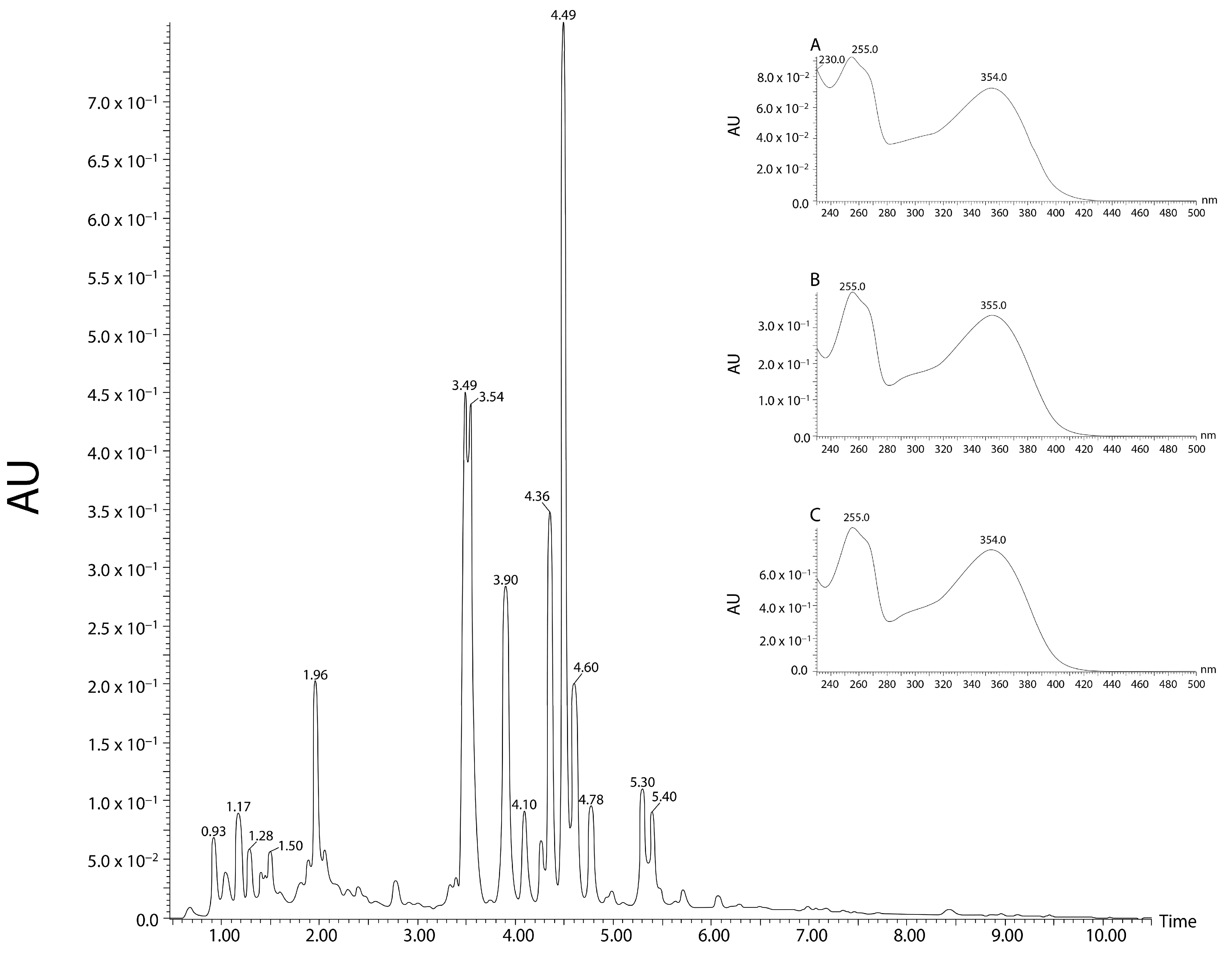
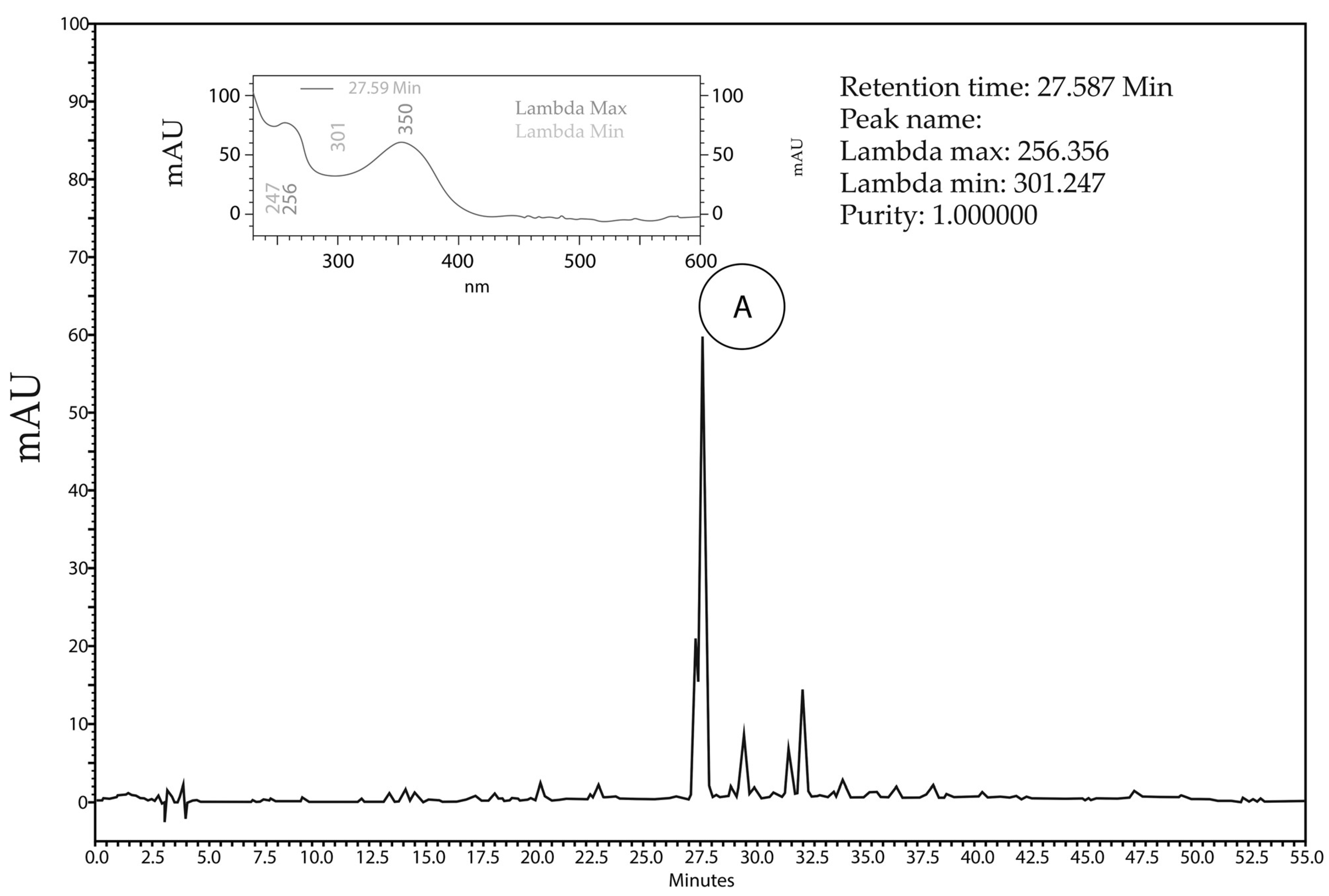
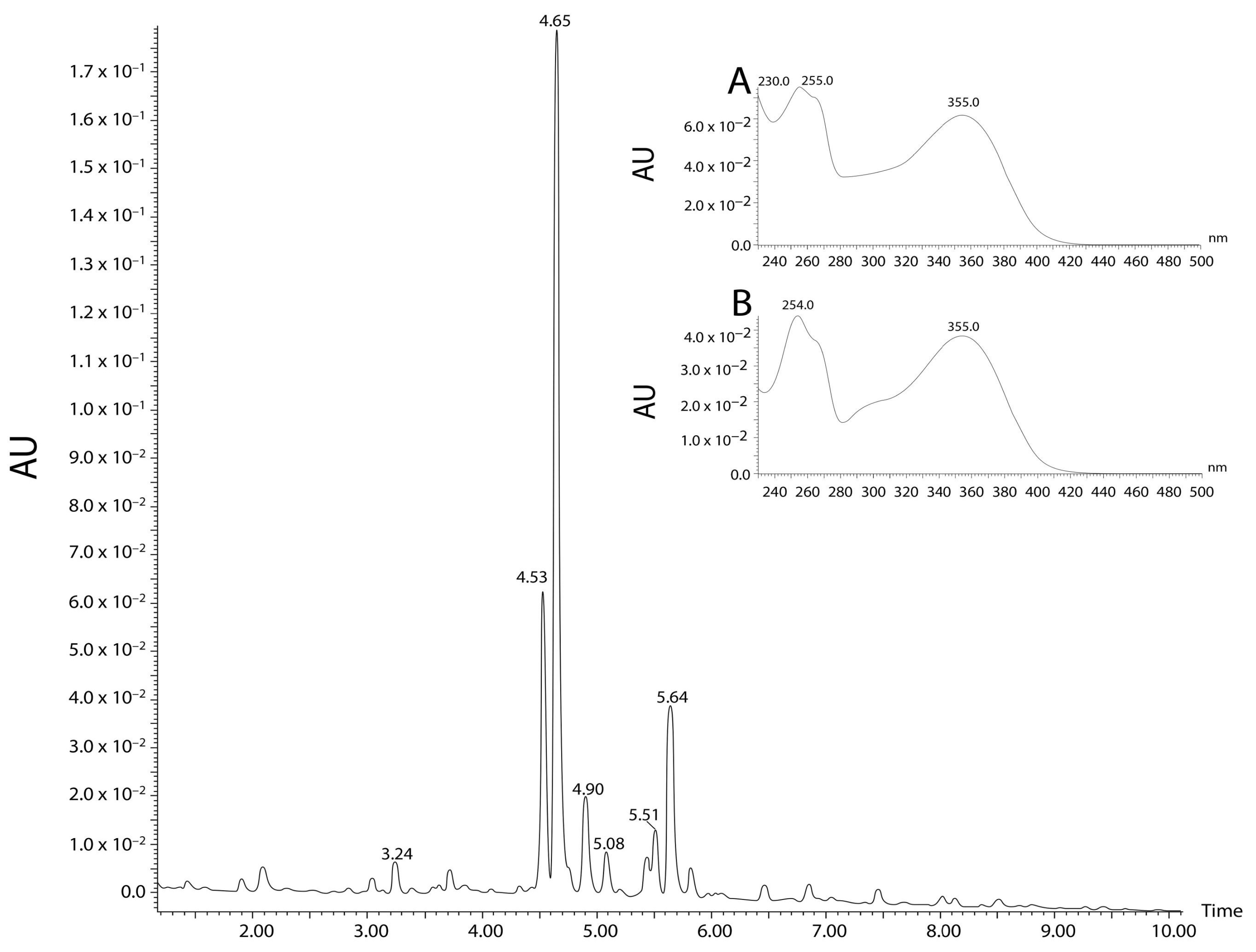
2.1. Phytochemical Analyses
2.2. Cytotoxicity Test
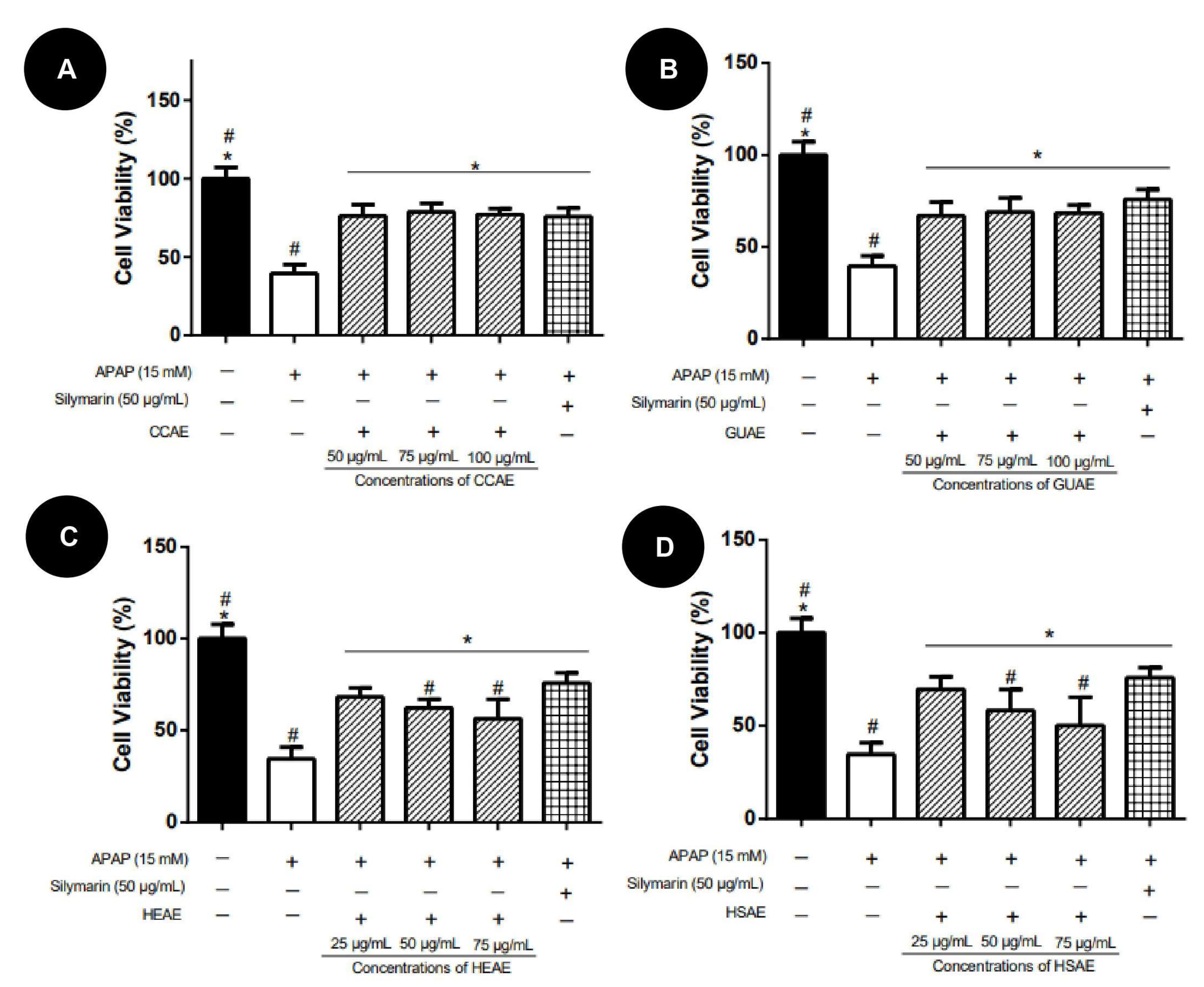
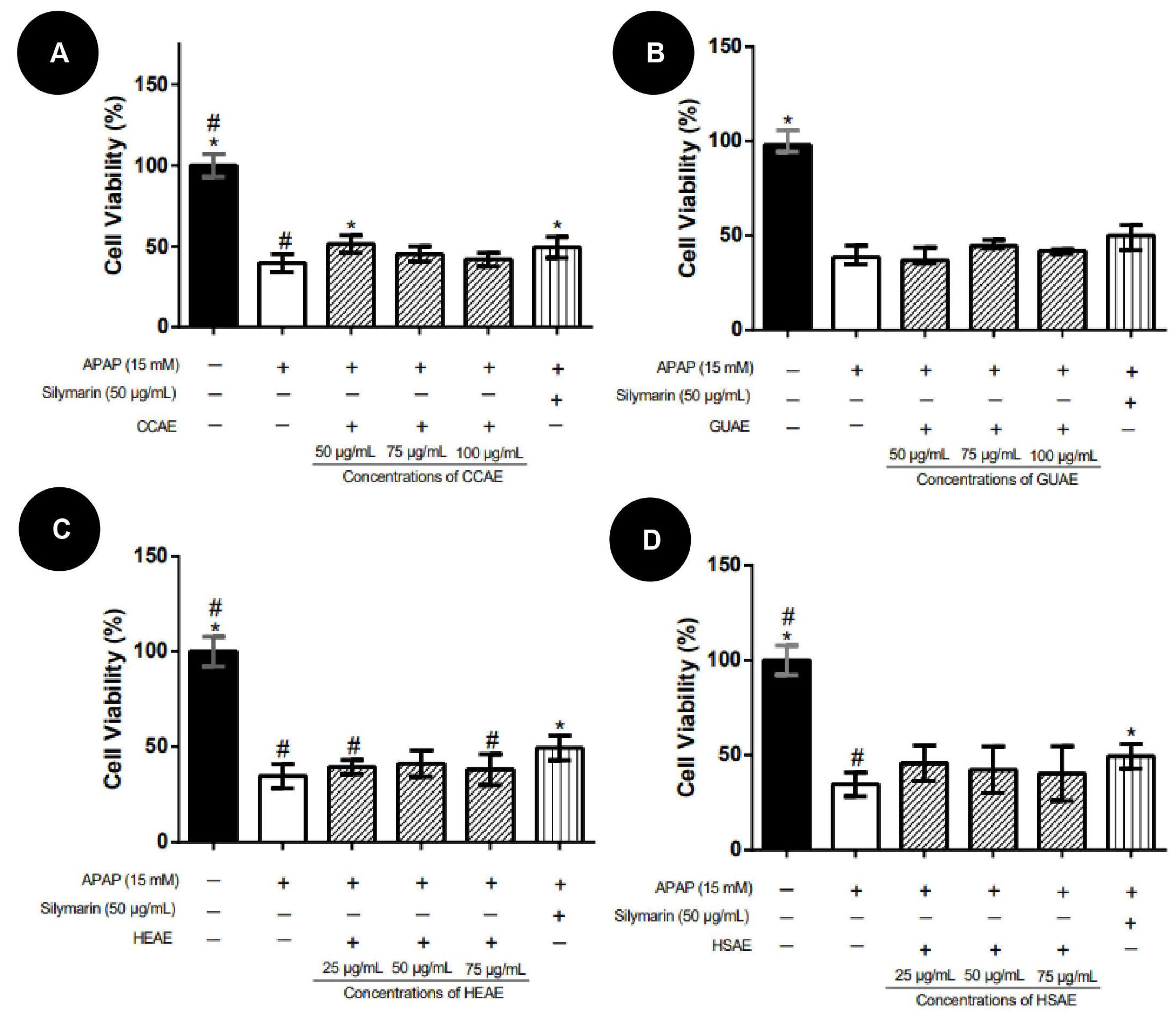
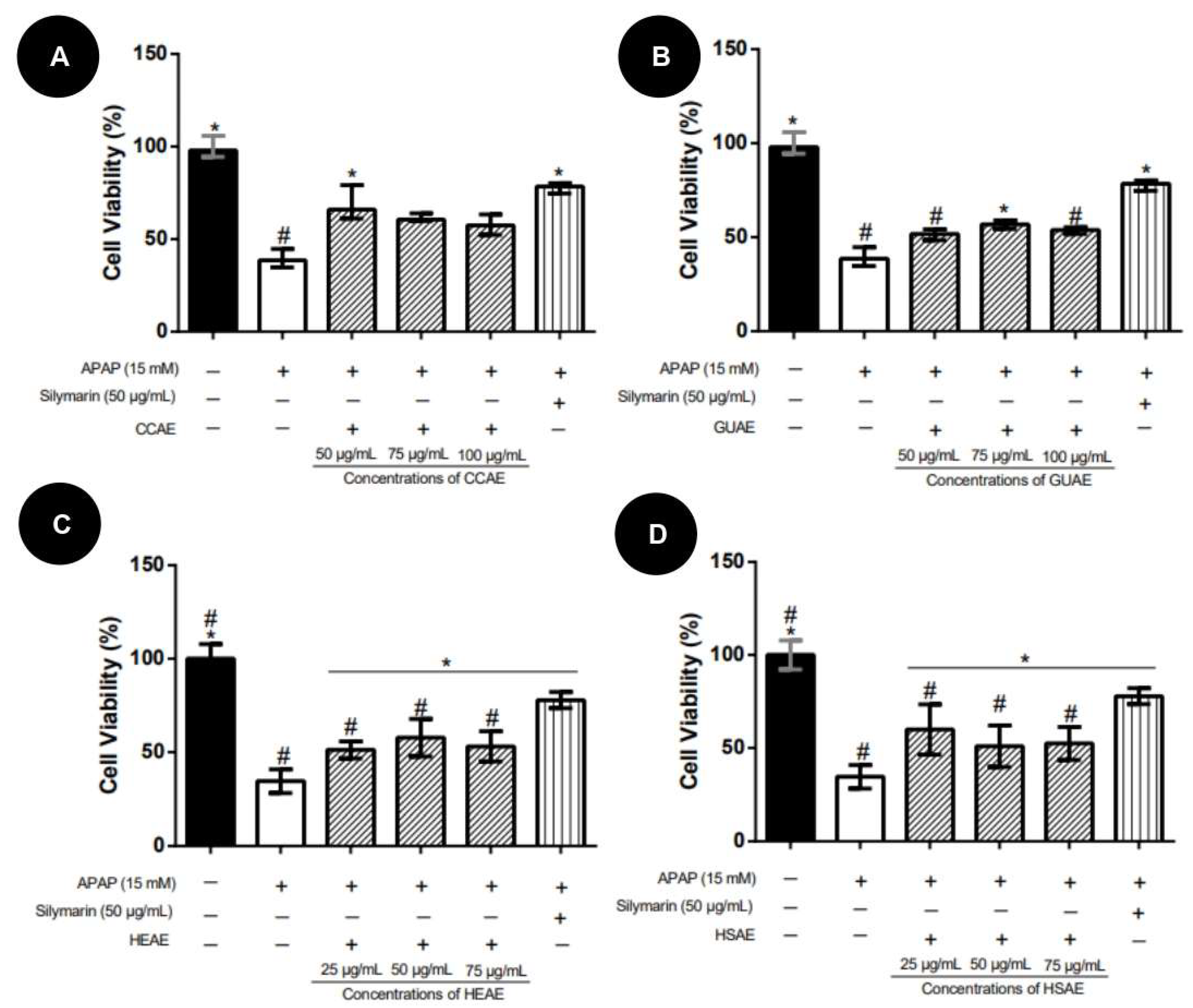
2.3. Activity against APAP Toxicity
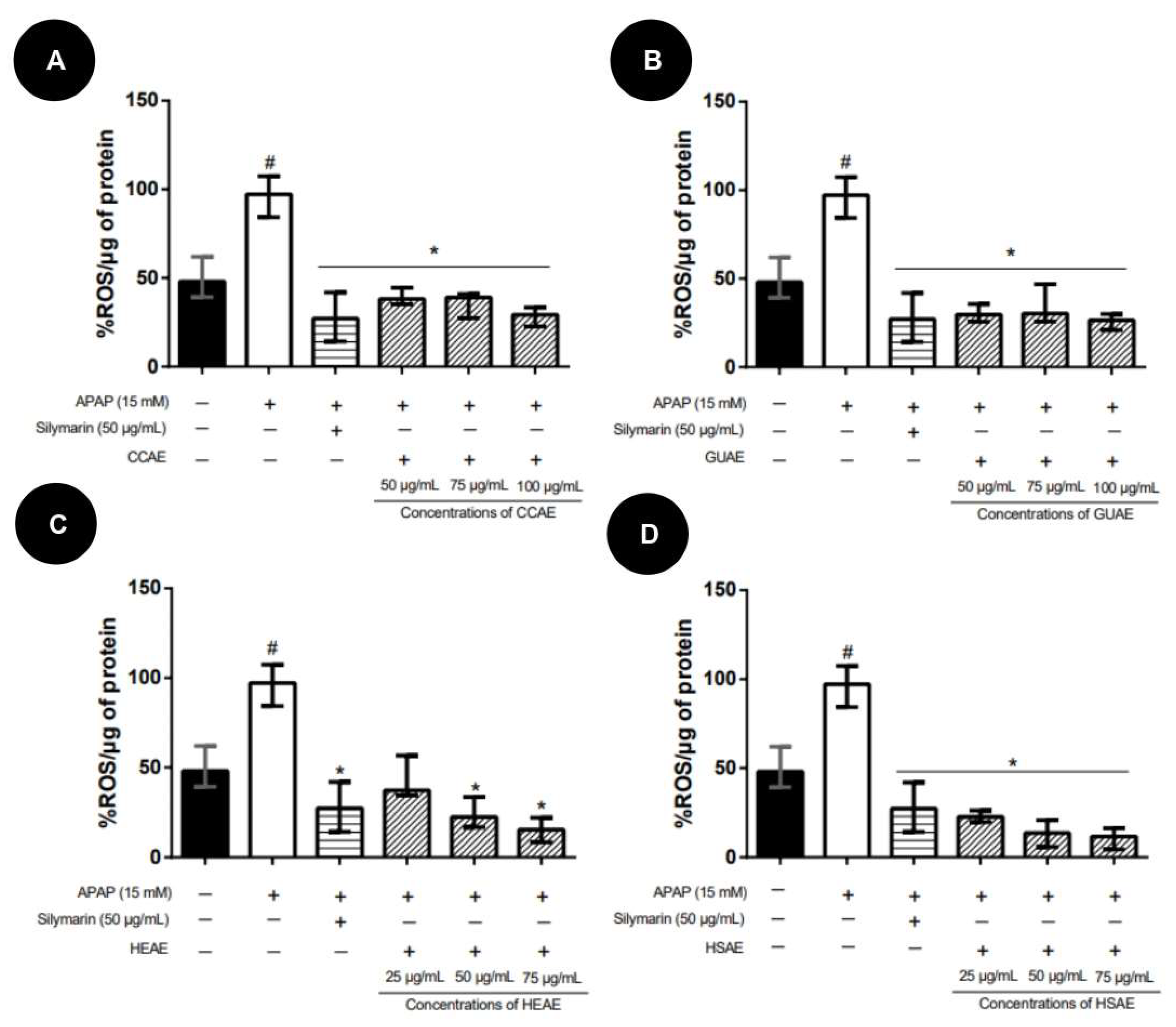
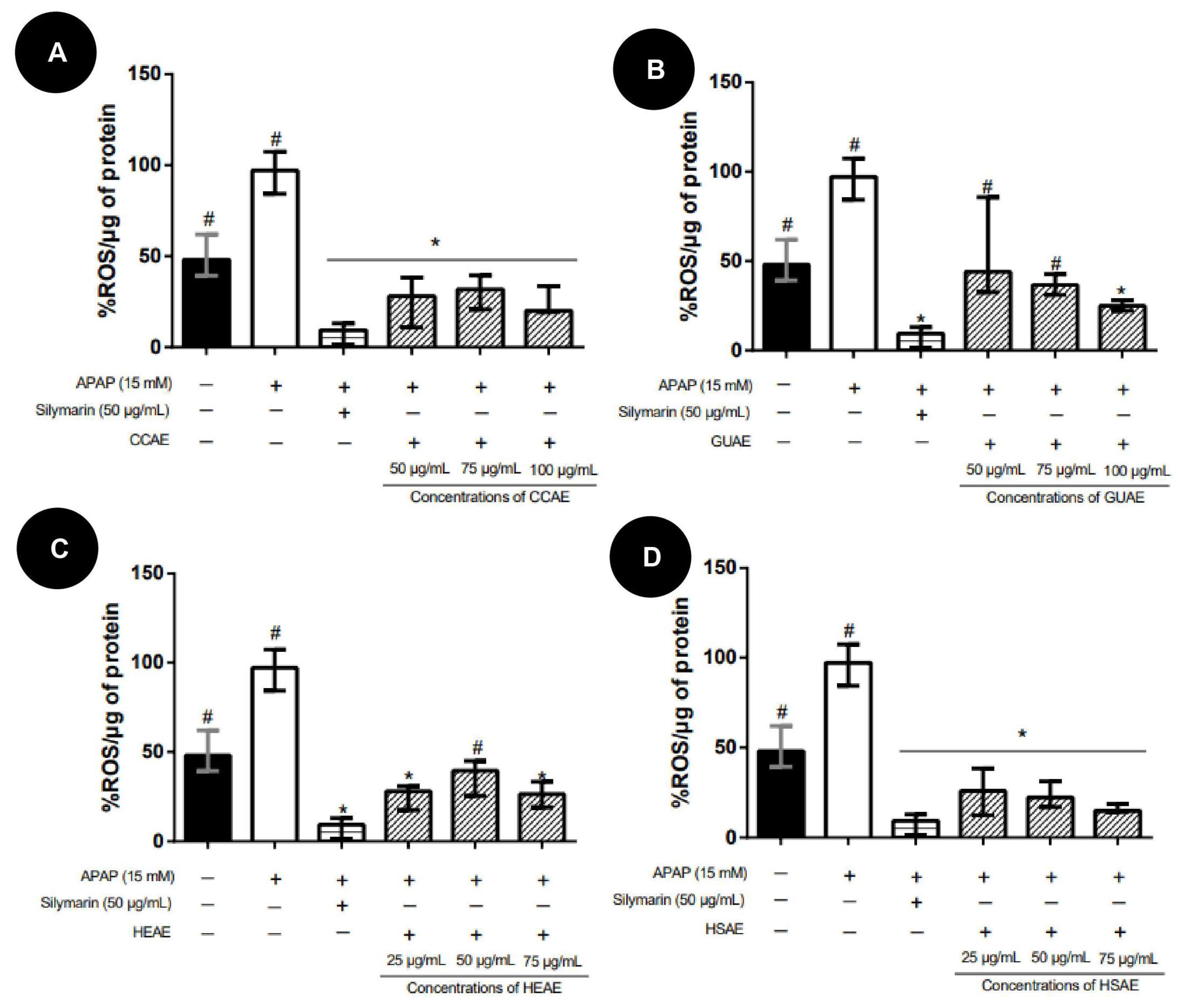
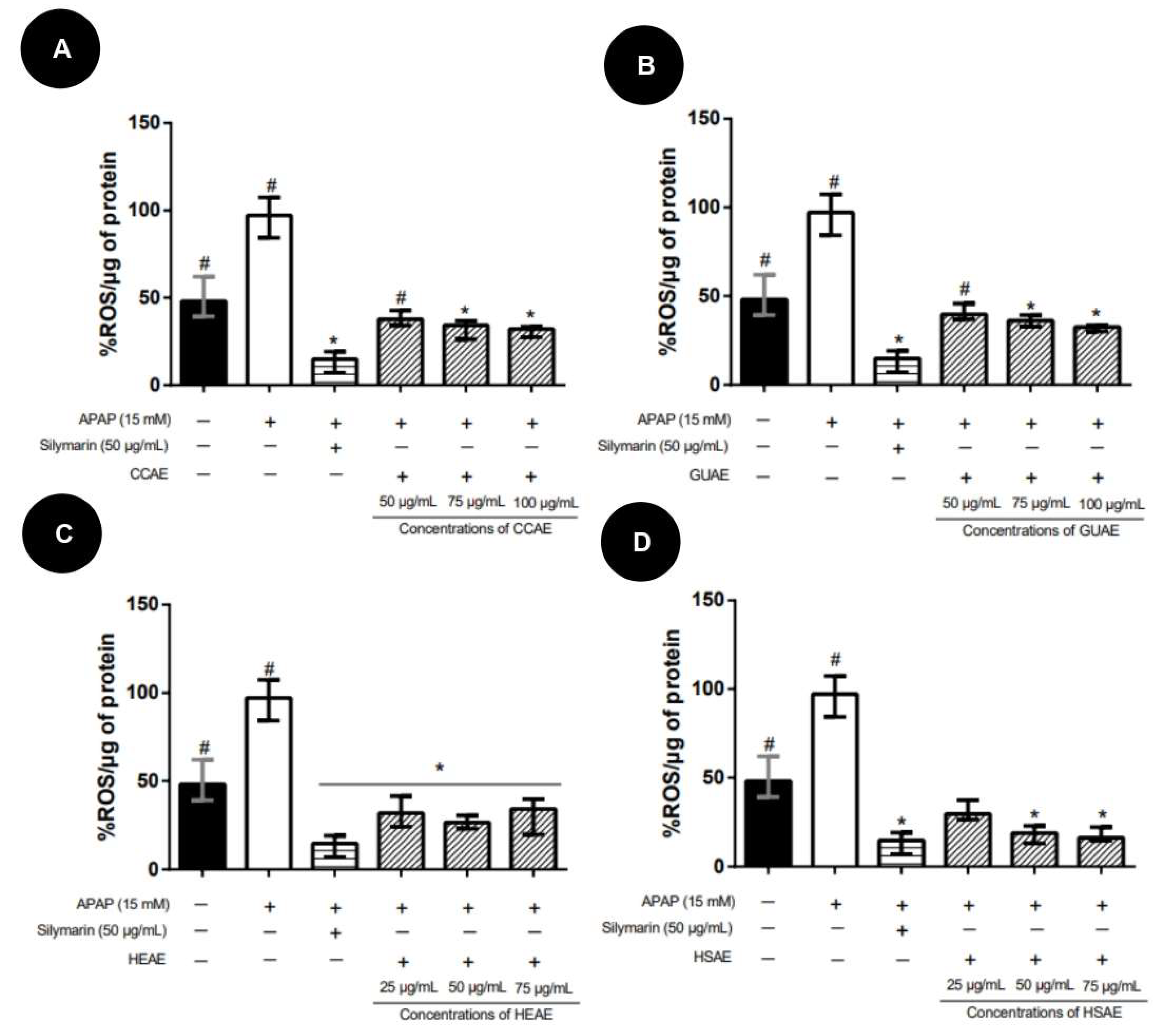
2.4. Quantification In Vitro of ROS
3. Discussion
4. Materials and Methods
4.1. Plant Material and Preparation of Extract
4.2. Phytochemical Analyzes
4.2.1. High-Performance Liquid Chromatography Analysis (HPLC)
4.2.2. Ultra-High Performance Liquid Chromatography with Tandem Mass Spectrometry Analysis (UHPLC-MS/MS)
4.3. In Vitro APAP-Induced Toxicity Assay
4.3.1. Cell Culture
4.3.2. Cytotoxicity Test
4.3.3. Activity against APAP-Induced Toxicity
4.3.4. In Vitro Quantification of Reactive Oxygen Species (ROS)
4.4. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Levy, B. Fisiologia, 6th ed.; Elsevier: Rio de Janeiro, Brazil, 2009. [Google Scholar]
- Carvalho, H.F. Células: Uma Abordagem Multidisciplinar; Manole: Barueri, Brazil, 2005; p. 450. [Google Scholar]
- Brasil. Cadernos de Atenção Básica—Práticas Integrativas e Complementares: Plantas Medicinais e Fitoterapia na Atenção Básica, 1st ed.; Ministério da Saúde: Brasília, Brazil, 2012; Volume 31, p. 156.
- Amadi, C.N.; Orisakwe, O.E. Herb-induced liver injuries in developing nations: An update. Toxics 2018, 6, 24. [Google Scholar] [CrossRef]
- Liao, J.; Lu, Q.; Li, Z.; Li, J.; Zhao, Q.; Li, J. Acetaminophen-induced liver injury: Molecular mechanism and treatments from natural products. Front. Pharm. 2023, 14, 1122632. [Google Scholar] [CrossRef]
- Farias, P.D.O. Aspectos epidemiológicos das intoxicações por analgésicos não opioides e anti-inflamatórios não esteroides em um hospital de urgência e emergência da rede pública do Brasil. Rev. Médica Minas Gerais 2016, 26 (Suppl. S5), S11–S15. [Google Scholar]
- Wolf, M.S.; King, J.; Jacobson, K.; Di Francesco, L.; Bailey, S.C.; Mullen, R.; McCarthy, D.; Serper, M.; Davis, T.C.; Parker, R.M. Risk of unintentional overdose with non-prescription acetaminophen products. J. Gen. Intern. Med. 2012, 27, 1587–1593. [Google Scholar] [CrossRef]
- Sebben, V.C.; Lugoch, R.D.W.; Schlinker, C.S.; Arbo, M.D.; Vianna, R.L. Validação de metodologia analítica e estudo de estabilidade para quantificação sérica de paracetamol. J. Bras. Patol. Med. Lab. 2010, 46, 143–148. [Google Scholar] [CrossRef]
- Saccomano, S.J. Acute acetaminophen toxicity in adults. Nurs. Crit. Care 2019, 14, 10–17. [Google Scholar] [CrossRef]
- Agrawal, S.; Khazaeni, B. Acetaminophen Toxicity; StatPearls Publishing: St. Petersburg, FL, USA, 2023. [Google Scholar]
- Subramanya, S.B.; Venkataraman, B.; Meeran, M.F.N.; Goyal, S.N.; Patil, C.R.; Ojha, S. Therapeutic Potential of Plants and Plant Derived Phytochemicals against Acetaminophen-Induced Liver Injury. Int. J. Mol. Sci. 2018, 19, 3776. [Google Scholar] [CrossRef] [PubMed]
- Costa, E.A.; Santos, L.R.; Pontes, I.S.; Matos, L.G.; Silva, G.A.; Lião, L.M. Analgesic and anti-inflammatory effects of Cheiloclinium cognatum root barks. Braz. J. Pharmacogn. 2007, 17, 508–513. [Google Scholar] [CrossRef]
- Pereira, R.C.G.; Soares, D.C.F.; Oliveira, D.C.P.; de Sousa, G.F.; Vieira-Filho, S.A.; Mercadante-Simoes, M.O.; Lula, I.; Silva-Cunha, A.; Duarte, L.P. Triterpenes from leaves of Cheiloclinium cognatum and their in vivo antiangiogenic activity. Magn. Reason. Chem. 2018, 56, 360–366. [Google Scholar] [CrossRef]
- Jeller, A.H.; Silva, D.H.; Liao, L.M.; Bolzani Vda, S.; Furlan, M. Antioxidant phenolic and quinonemethide triterpenes from Cheiloclinium cognatum. Phytochemistry 2004, 65, 1977–1982. [Google Scholar] [CrossRef]
- Feltrin, A.C.; Boligon, A.A.; Janovik, V.; Athayde, M.L. Antioxidant potential, total phenolic and flavonoid contents from the stem bark of Guazuma ulmifolia Lam. Asian J. Biol. Sci. 2012, 5, 268–272. [Google Scholar] [CrossRef]
- Pereira, G.A.; Peixoto Araujo, N.M.; Arruda, H.S.; Farias, D.P.; Molina, G.; Pastore, G.M. Phytochemicals and biological activities of mutamba (Guazuma ulmifolia Lam.): A review. Food Res. Int. 2019, 126, 108713. [Google Scholar] [CrossRef] [PubMed]
- Geller, F.C.; Teixeira, M.R.; Pereira, A.B.; Dourado, L.P.; Souza, D.G.; Braga, F.C.; Simoes, C.M. Evaluation of the Wound Healing Properties of Hancornia speciosa Leaves. Phytother. Res. 2015, 29, 1887–1893. [Google Scholar] [CrossRef] [PubMed]
- Dos Santos, U.P.; Tolentino, G.S.; Morais, J.S.; de Picoli Souza, K.; Estevinho, L.M.; Dos Santos, E.L. Physicochemical Characterization, Microbiological Quality and Safety, and Pharmacological Potential of Hancornia speciosa Gomes. Oxid. Med. Cell. Longev. 2018, 2018, 2976985. [Google Scholar] [CrossRef]
- Dimech, G.S.; Soares, L.A.; Ferreira, M.A.; de Oliveira, A.G.; Carvalho Mda, C.; Ximenes, E.A. Phytochemical and antibacterial investigations of the extracts and fractions from the stem bark of Hymenaea stigonocarpa Mart. ex Hayne and effect on ultrastructure of Staphylococcus aureus induced by hydroalcoholic extract. Sci. World J. 2013, 2013, 862763. [Google Scholar] [CrossRef]
- Monteiro, A.F.; Batista, J.M., Jr.; Machado, M.A.; Severino, R.P.; Blanch, E.W.; Bolzani, V.S.; Vieira, P.C.; Severino, V.G. Structure and Absolute Configuration of Diterpenoids from Hymenaea stigonocarpa. J. Nat. Prod. 2015, 78, 1451–1455. [Google Scholar] [CrossRef]
- Orsi, P.R.; Seito, L.N.; Di Stasi, L.C. Hymenaea stigonocarpa Mart. ex Hayne: A tropical medicinal plant with intestinal anti-inflammatory activity in TNBS model of intestinal inflammation in rats. J. Ethnopharmacol. 2014, 151, 380–385. [Google Scholar] [CrossRef]
- Rodrigues Orsi, P.; Bonamin, F.; Aparecida Severi, J.; Cassia Santos, R.; Vilegas, W.; Hiruma-Lima, C.A.; Stasi, L.C. Hymenaea stigonocarpa Mart. ex Hayne: A Brazilian medicinal plant with gastric and duodenal anti-ulcer and antidiarrheal effects in experimental rodent models. J. Ethnopharmacol. 2012, 143, 81–90. [Google Scholar] [CrossRef]
- Gillessen, A.; Schmidt, H.H.-J. Silymarin as supportive treatment in liver diseases: A narrative review. Adv. Ther. 2020, 37, 1279–1301. [Google Scholar] [CrossRef]
- ISO I. 10993–5: 2009; Biological Evaluation of Medical Devices—Part 5: Tests for in Vitro Cytotoxicity. International Organization for Standardization: Geneva, Switzerland, 2009.
- Sohn, J.H.; Han, K.-L.; Lee, S.-H.; Hwang, J.-K. Protective effects of panduratin A against oxidative damage of tert-Butylhydroperoxide in human HepG2 cells. Biol. Pharm. Bull. 2005, 28, 1083–1086. [Google Scholar] [CrossRef]
- Behrends, V.; Giskeodegard, G.F.; Bravo-Santano, N.; Letek, M.; Keun, H.C. Acetaminophen cytotoxicity in HepG2 cells is associated with a decoupling of glycolysis from the TCA cycle, loss of NADPH production, and suppression of anabolism. Arch. Toxicol. 2019, 93, 341–353. [Google Scholar] [CrossRef]
- Choi, Y.H.; Lee, H.S.; Chung, C.K.; Kim, E.J.; Kang, I.J. Protective effects of an ethanol extract of Angelica keiskei against acetaminophen-induced hepatotoxicity in HepG2 and HepaRG cells. Nutr. Res. Pract. 2017, 11, 97–104. [Google Scholar] [CrossRef] [PubMed]
- Chowdhury, A.; Lu, J.; Zhang, R.; Nabila, J.; Gao, H.; Wan, Z.; Temitope, I.A.; Yin, X.; Sun, Y. Mangiferin ameliorates acetaminophen-induced hepatotoxicity through APAP-Cys and JNK modulation. Biomed. Pharma. 2019, 117, 109097. [Google Scholar] [CrossRef] [PubMed]
- Truong, V.-L.; Ko, S.-Y.; Jun, M.; Jeong, W.-S. Quercitrin from Toona sinensis (Juss.) M.Roem. Attenuates Acetaminophen-Induced Acute Liver Toxicity in HepG2 Cells and Mice through Induction of Antioxidant Machinery and Inhibition of Inflammation. Nutrients 2016, 8, 431. [Google Scholar] [CrossRef] [PubMed]
- Miret, S.; De Groene, E.M.; Klaffke, W. Comparison of in vitro assays of cellular toxicity in the human hepatic cell line HepG2. J. Biomol. Screen. 2006, 11, 184–193. [Google Scholar] [CrossRef]
- Malta, L.G.; Tessaro, E.P.; Eberlin, M.; Pastore, G.M.; Liu, R.H. Assessment of antioxidant and antiproliferative activities and the identification of phenolic compounds of exotic Brazilian fruits. Food Res. Int. 2013, 53, 417–425. [Google Scholar] [CrossRef]
- Suya, D.H.L.; Lei, T.; Xie, X.; Wang, H.; He, X.; Tong, R.; Wang, Y. Mangiferin: An effective therapeutic agent against several disorders (Review). Mol. Med. Rep. 2018, 18, 4775–4786. [Google Scholar]
- Muhammad Imran, M.S.A.; Butt, M.S.; Kwon, J.-H.; Arshad, M.U.; Sultan, M.T. Mangiferin: A natural miracle bioactive compound against lifestyle related disorders. Lipids Health Dis. 2017, 16, 84. [Google Scholar] [CrossRef]
- Das, J.; Ghosh, J.; Roy, A.; Sil, P.C. Mangiferin exerts hepatoprotective activity against D-galactosamine induced acute toxicity and oxidative/nitrosative stress via Nrf2-NFkappaB pathways. Toxicol. Appl. Pharmacol. 2012, 260, 35–47. [Google Scholar] [CrossRef]
- Domitrovic, R.; Jakovac, H.; Vasiljev Marchesi, V.; Vladimir-Knezevic, S.; Cvijanovic, O.; Tadic, Z.; Romic, Z.; Rahelic, D. Differential hepatoprotective mechanisms of rutin and quercetin in CCl(4)-intoxicated BALB/cN mice. Acta Pharma. Sin. 2012, 33, 1260–1270. [Google Scholar] [CrossRef]
- Reddy, M.K.; Reddy, A.G.; Kumar, B.K.; Madhuri, D.; Boobalan, G.; Reddy, M.A. Protective effect of rutin in comparison to silymarin against induced hepatotoxicity in rats. Vet. World 2017, 10, 74. [Google Scholar] [CrossRef]
- Xu, Y.; Chen, J.; Yu, X.; Tao, W.; Jiang, F.; Yin, Z.; Liu, C. Protective effects of chlorogenic acid on acute hepatotoxicity induced by lipopolysaccharide in mice. Inflamm. Res. 2010, 59, 871–877. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Yang, Y.; Mi, S.; Fan, Q.; Sun, X.; Deng, B.; Wu, G.; Li, Y.; Zhou, Q.; Ruan, Z. Hepatoprotective effect of chlorogenic acid against chronic liver injury in inflammatory rats. J. Funct. Foods 2019, 62, 103540. [Google Scholar] [CrossRef]
- Manov, I.; Hirsh, M.; Iancu, T.C. N-Acetylcysteine does not Protect HepG2Cells against Acetaminophen-Induced Apoptosis. Basic Clin. Pharmacol. Toxicol. 2004, 94, 213–225. [Google Scholar] [CrossRef] [PubMed]
- Hao, Z.Y.; Liang, D.; Luo, H.; Liu, Y.F.; Ni, G.; Zhang, Q.J.; Li, L.; Si, Y.K.; Sun, H.; Chen, R.Y.; et al. Bioactive sesquiterpenoids from the rhizomes of Acorus calamus. J. Nat. Prod. 2012, 75, 1083–1089. [Google Scholar] [CrossRef] [PubMed]
- Abebe, W. Herbal medication: Potential for adverse interactions with analgesic drugs. J. Clin. Pharm. Ther. 2002, 27, 391–401. [Google Scholar] [CrossRef]
- Britza, S.M.; Farrington, R.; Byard, R.W.; Musgrave, I.F. In vitro demonstration of herbal exacerbation of paracetamol-induced hepatotoxicity. Phytomed. Plus 2022, 2, 100267. [Google Scholar] [CrossRef]
- Abdel-Ghaf, O.; Mahmoud, S.T.; Ali Said, A.; Abdel-Azee, F. Hepatoprotective Effect of Rutin Against Oxidative Stress of Isoniazid in Albino Rats. Int. J. Pharmacol. 2017, 13, 516–528. [Google Scholar] [CrossRef]
- Parikh, H.; Pandita, N.; Khanna, A. Phytoextract of Indian mustard seeds acts by suppressing the generation of ROS against acetaminophen-induced hepatotoxicity in HepG2 cells. Pharm. Biol. 2015, 53, 975–984. [Google Scholar] [CrossRef]
- Fan, X.; Lv, H.; Wang, L.; Deng, X.; Ci, X. Isoorientin Ameliorates APAP-Induced Hepatotoxicity via Activation Nrf2 Antioxidative Pathway: The Involvement of AMPK/Akt/GSK3beta. Front. Pharmacol. 2018, 9, 1334. [Google Scholar] [CrossRef]
- Niture, S.K.; Jaiswal, A.K. Nrf2 protein up-regulates antiapoptotic protein Bcl-2 and prevents cellular apoptosis. J. Biol. Chem. 2012, 287, 9873–9886. [Google Scholar] [CrossRef] [PubMed]
- Borges Bubols, G.; da Rocha Vianna, D.; Medina-Remon, A.; von Poser, G.; Maria Lamuela-Raventos, R.; Lucia Eifler-Lima, V.; Cristina Garcia, S. The Antioxidant Activity of Coumarins and Flavonoids. Mini-Rev. Med. Chem. 2013, 13, 318–334. [Google Scholar] [CrossRef]
- Alia, M.; Mateos, R.; Ramos, S.; Lecumberri, E.; Bravo, L.; Goya, L. Influence of quercetin and rutin on growth and antioxidant defense system of a human hepatoma cell line (HepG2). Eur. J. Nutr. 2006, 45, 19–28. [Google Scholar] [CrossRef] [PubMed]
- Tabolacci, E.; Tringali, G.; Nobile, V.; Duca, S.; Pizzoferrato, M.; Bottoni, P.; Clementi, M.E. Rutin Protects Fibroblasts from UVA Radiation through Stimulation of Nrf2 Pathway. Antioxidants 2023, 12, 820. [Google Scholar] [CrossRef] [PubMed]
- Agarwala, S.; Mudholkar, K.; Bhuwania, R.; Satish Rao, B.S. Mangiferin, a dietary xanthone protects against mercury-induced toxicity in HepG2 cells. Environ. Toxicol. 2012, 27, 117–127. [Google Scholar] [CrossRef]
- Leite, C.F.M.; Leite, B.H.M.; de Carvalho Barros, I.M.; Gomes, S.M.; Fagg, C.W.; Simeoni, L.A.; Fonseca, Y.M. Determination of rutin in Erythroxylum suberosum extract by liquid chromatography: Applicability in standardization of herbs and stability studies. Bol. Latinoam. Caribe Plantas Med. Aromát. 2014, 13, 135–143. [Google Scholar]
- Mosmann, T. Rapid Colorimetric Assay for Cellular Growth and Survival: Application to Proliferation and Cytotoxicity Assays. J. Immunol. Methods 1983, 65, 55–63. [Google Scholar] [CrossRef]
- Hansen, M.B.; Nielsen, S.E.; Berg, K. Re-examination and further development of a precise and rapid dye method for measuring cell growth/cell kill. J. Immunol. Methods 1989, 119, 203–210. [Google Scholar] [CrossRef]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Date | Time | Place | Geographic Coordinates | Exsiccata Numbers |
|---|---|---|---|---|---|
| Cheiloclinium cognatum | 9 March 2018 | 10 a.m.–12 p.m. | (1) | 15°90′85.83″ S 47°91′36.11″ W | Mendonça, R. 4991 |
| Guazuma ulmifolia | 2 May 2018 | 17 p.m.–18 p.m. | (1) | 15°52′30.90″ S 47°57′24″ W | Fagg, C.W. 2484 |
| Hancornia speciosa | 22 February 2018 | 10 a.m.–11 a.m. | (2) | 15°45′48.21″ S 47°51′50.67″ W | Fagg, C.W. 2495 |
| Hymenaea stigonocarpa | 17 May 2018 | 14 p.m.–15 p.m. | (2) | 15°45′48.21″ S 47°51′50.67″ W | Fagg, C.W. 2491 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ribeiro, G.d.S.; Martins, D.H.N.; Gomes, J.V.D.; Davies, N.W.; Fagg, C.W.; Simeoni, L.A.; Homem-de-Mello, M.; Magalhães, P.O.; Silveira, D.; Fonseca-Bazzo, Y.M. Hepatoprotective Effects of Four Brazilian Savanna Species on Acetaminophen-Induced Hepatotoxicity in HepG2 Cells. Plants 2023, 12, 3393. https://doi.org/10.3390/plants12193393
Ribeiro GdS, Martins DHN, Gomes JVD, Davies NW, Fagg CW, Simeoni LA, Homem-de-Mello M, Magalhães PO, Silveira D, Fonseca-Bazzo YM. Hepatoprotective Effects of Four Brazilian Savanna Species on Acetaminophen-Induced Hepatotoxicity in HepG2 Cells. Plants. 2023; 12(19):3393. https://doi.org/10.3390/plants12193393
Chicago/Turabian StyleRibeiro, Gislane dos Santos, Diegue Henrique Nascimento Martins, João Victor Dutra Gomes, Noel William Davies, Christopher William Fagg, Luiz Alberto Simeoni, Mauricio Homem-de-Mello, Pérola Oliveira Magalhães, Dâmaris Silveira, and Yris Maria Fonseca-Bazzo. 2023. "Hepatoprotective Effects of Four Brazilian Savanna Species on Acetaminophen-Induced Hepatotoxicity in HepG2 Cells" Plants 12, no. 19: 3393. https://doi.org/10.3390/plants12193393