
Blue Light Enhances Health-Promoting Sulforaphane Accumulation in Broccoli (Brassica oleracea var. italica) Sprouts through Inhibiting Salicylic Acid Synthesis
Abstract
:1. Introduction
2. Results
2.1. Effects of Blue Light on the Growth of Broccoli Sprouts
2.2. Effects of Blue Light on the Content of SFN and MYR Activity
2.3. Effect of Blue Light on Antioxidant Enzyme Activity of Broccoli Sprouts
2.4. Effects of Blue Light on the Expression Level of Related Genes of SFN Synthesis
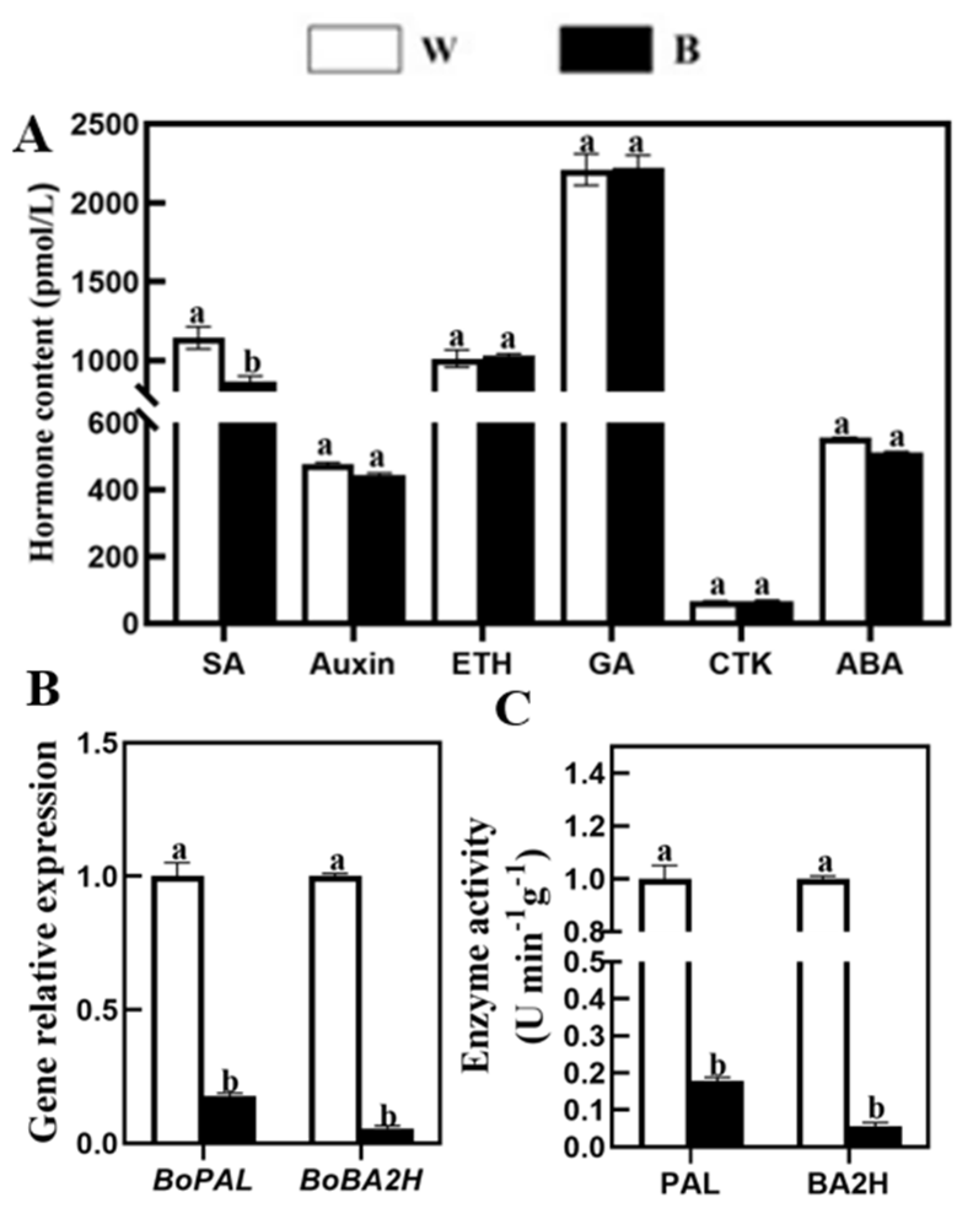
2.5. Effects of Blue Light on Endogenous Hormone Content, Expression of Key Genes and Enzymatic Activity of SA Synthesis Pathway
2.6. Effects of Exogenous SA on Content of Endogenous SA, SFN Synthesis
2.7. Effects of SA on the Expression of SFN Synthesis Genes
2.8. Verification of SA Biosynthesis-Deficient Arabidopsis Mutants
3. Materials and Methods
3.1. Plant Growth and Treatments
3.2. SA Treatments of Broccoli Sprouts
3.3. Determination of Growth Parameters
3.4. Determination of SFN Content
3.5. Enzymatic Activities Assay
3.6. Analysis of Plant Hormone Content
3.7. RNA Preparation and Quantitative Real-Time-PCR Analysis
3.8. Statistical Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Esfandiari, A.; Saei, A.; McKenzie, M.J.; Matich, A.J.; Babalar, M.; Hunter, D.A. Preferentially enhancing anti-cancer isothiocyanates over glucosinolates in broccoli sprouts How NaCl and salicylic acid af-fect their formation. Plant Physiol. Biochem. 2017, 115, 343–353. [Google Scholar]
- Miao, H.; Xia, C.; Yu, S.; Wang, J.; Zhao, Y.; Wang, Q. Enhancing health-promoting isothiocyanates in Chinese kale sprouts via manipulating BoESP. Hortic. Res. 2023, 10, uhad029. [Google Scholar]
- Cheng, A.; Shen, C.; Hung, C.; Hsu, Y. Sulforaphane Decrease of SERTAD1 Expression Triggers G1/S Arrest in Breast Cancer Cells. J. Med. Food 2019, 22, 444–450. [Google Scholar] [CrossRef]
- Martinez-Villaluenga, C.; Penas, E.; Ciska, E.; Piskula, M.K.; Kozlowska, H.; Vidal-Valverde, C.; Frias, J. Time dependence of bioactive compounds and antioxidant capacity during germination of different cultivars of broccoli and radish seeds. Food Chem. 2010, 120, 710–716. [Google Scholar] [CrossRef]
- Fix, C.; Carver-Molina, A.; Chakrabarti, M.; Azhar, M.; Carver, W. Effects of the isothiocyanate sulforaphane on TGF-β1-induced rat cardiac fibroblast activation and extracellular matrix interactions. J. Cell Physiol. 2019, 243, 13931–13942. [Google Scholar] [CrossRef]
- Farag, M.A.; Motaal, A.A.A. Sulforaphane composition, cytotoxic and antioxidant activity of crucifer vegetables. J. Adv. Res. 2010, 1, 65–70. [Google Scholar] [CrossRef]
- Ruhee, R.T.; Suzuki, K. The Integrative Role of Sulforaphane in Preventing Inflammation, Oxidative Stress and Fatigue: A Review of a Potential Protective Phytochemical. Antioxidants 2020, 9, 521. [Google Scholar] [CrossRef]
- Lambrix, V.; Reichelt, M.; Mitchell-Olds, T.; Gershenzon, K.J. The Arabidopsis Epithiospecifier Protein Promotes the Hydrolysis of Glucosinolates to Nitriles and Influences Trichoplusia ni Herbivory. Plant Cell 2001, 13, 2793–2807. [Google Scholar]
- Seo, M.S.; Jin, M.; Chun, J.H.; Kim, S.J.; Park, B.S.; Shon, S.H.; Kim, J.S. Functional analysis of three BrMYB28 transcription factors controlling the biosynthesis of glucosinolates in Brassica rapa. Plant Mol. Biol. 2016, 90, 503–516. [Google Scholar] [CrossRef]
- Tian, M.; Yang, Y.; Avila, F.; William, F.; Tara, Y. Effects of Selenium Supplementation on Glucosinolate Biosynthesis in Broccoli. J. Agric. Food Chem. 2018, 66, 8036–8044. [Google Scholar] [CrossRef]
- Yang, R.; Guo, L.; Jin, X.; Shen, C.; Zhou, Y.; Gu, Z. Enhancement of glucosinolate and sulforaphane formation of broccoli sprouts by zinc sulphate via its stress effect. J. Funct. Foods 2015, 13, 345–349. [Google Scholar] [CrossRef]
- Miao, H.; Cai, C.; Wei, J.; Huang, J.; Chang, J.; Qian, H.; Zhang, X.; Zhao, Y.; Sun, B.; Wang, B.; et al. Glucose enhances indolic glucosinolate biosynthesis without reducing primary sulfur assimilation. Sci. Rep. 2016, 6, 31854. [Google Scholar]
- Riahi-Madvar, A.; Rezaee, F.; Bonyad, F.J.; Ahsaei, M.G.; Goharrizi, K.J. Effect of Salinity Stress on Enzymes’ Activity, Ions Concentration, Oxidative Stress Parameters, Biochemical Traits, Content of Sulforaphane, and CYP79F1 Gene Expression Level in Lepidium draba Plant. J. Plant Growth Regul. 2020, 39, 1075–1094. [Google Scholar]
- Amer, M.A.; Mohamed, T.R.; Rahman RA, A.; Ali, M.; Badr, A. Studies on exogenous elicitors promotion of sulforaphane content in broccoli sprouts and its effect on the MDA-MB-231 breast cancer cell line—ScienceDirect. Ann. Agric. Sci. 2021, 66, 46–52. [Google Scholar] [CrossRef]
- Kestwal, R.M.; Lin, J.C.; Bagal-Kestwal, D.; Chiang, B.H. Glucosinolates fortification of cruciferous sprouts by sulphur supplementation during cultivation to enhance anti-cancer activity. Food Chem. 2011, 126, 1164–1171. [Google Scholar] [CrossRef]
- Smith, H. Phytochrome and light signal perception by plants—An emerging synthesis. Nature 2000, 407, 585–591. [Google Scholar]
- Liu, H.; Li, S.; Ma, S.Y.; Luo, L.Y.; Liu, Y. Effects of Different LED Light Qualities on Callus Induction, Proliferation and Sulforaphane Content of Raphanus sativus L. Plant Physiol. Commun. 2010, 46, 347–350. [Google Scholar]
- Kopsell, D.A.; Sams, C.E. Increases in shoot tissue pigments, glucosinolates, and mineral elements in sprouting broccoli after exposure to short-duration blue light from light emitting diodes. J. Am. Soc. Hortic. Sci. 2013, 53, 60–133. [Google Scholar] [CrossRef]
- Kopsell, D.A.; Sams, C.E.; Barickman, T.C.; Morrow, R.C. Sprouting Broccoli Accumulate Higher Concentrations of Nutritionally Important Metabolites under Narrow-band Light-emitting Diode Lighting. J. Am. Soc. Hortic. Sci. 2014, 139, 469–477. [Google Scholar] [CrossRef]
- Dougher, T.A.O.; Bugbee, B. Differences in the response of wheat, soybean and lettuce to reduced blue radiation. Photochem. Photobiol. 2010, 73, 199–207. [Google Scholar] [CrossRef]
- Zheng, Y.; Zhang, Y.; Liu, H.; LI, Y.; Liu, Y.; Hao, Y.; Lei, Y. Supplemental blue light increases growth and quality of greenhouse pak choi depending on cultivar and supplemental light intensity. J. Integr. Agr. 2018, 17, 2245–2256. [Google Scholar] [CrossRef]
- Qian, H.; Liu, T.; Deng, M.; Miao, H.; Cai, C.; Shen, W.; Wang, Q. Effects of light quality on main health-promoting compounds and antioxidant capacity of Chinese kale sprouts. Food Chem. 2016, 196, 1232–1238. [Google Scholar] [CrossRef] [PubMed]
- Abe, K.; Kido, S.; Maeda, T.; Kami, D.; Matsuura, H.; Shimura, H.; Suzuki, T. Glucosinolate profiles in Cardamine fauriei and effect of light quality on glucosinolate concentration. Sci. Hortic. 2015, 189, 12–16. [Google Scholar] [CrossRef]
- Park, C.; Kim, N.; Park, J.; Lee, S.; Lee, J.W.; Park, S. Effects of Light-Emitting Diodes on the Accumulation of Glucosinolates and Phenolic Compounds in Sprouting Canola (Brassica napus L.). Foods 2019, 8, 76. [Google Scholar] [CrossRef]
- Nawaz, F.; Shabbir, R.N.; Shahbaz, M.; Majeed, S.; Sohail, M.A. Cross Talk between Nitric Oxide and Phytohormones Regulate Plant Development during Abiotic Stresses. In Phytohormones and Their Signaling Mechanisms in Plant Development; IntechOpen: London, UK, 2017. [Google Scholar]
- Sun, B.; Yan, H.; Zhang, F.; Wang, Q. Effects of plant hormones on main health-promoting compounds and antioxidant capacity of Chinese kale. Food Res. Int. 2012, 48, 359–366. [Google Scholar] [CrossRef]
- Baenas, N.; García-Viguera, C.; Moreno, D.A. Elicitation: A Tool for Enriching the Bioactive Composition of Foods. Molecules 2014, 19, 13541–13563. [Google Scholar] [CrossRef]
- Pérez-Balibrea, S.; Moreno, D.A.; García-Viguera, C. Improving the phytochemical composition of broccoli sprouts by elicitation. Food Chem. 2011, 129, 35–44. [Google Scholar] [CrossRef]
- Gigolashvili, T.; Berger, B.; Flügge, U. Specific and coordinated control of indolic and aliphatic glucosinolate biosynthesis by R2R3-MYB transcription factors in Arabidopsis thaliana. Phytochem. Rev. 2009, 8, 3–13. [Google Scholar] [CrossRef]
- Wang, T.; Zhang, D.; Yang, B.; Su, N.; Cui, J. Salicylic Acid Regulates Indole-3-Carbinol Biosynthesis Under Blue Light in Broccoli Sprouts (Brassica oleracea L.). Front. Plant Sci. 2022, 13, 848454. [Google Scholar] [CrossRef]
- Meeran, S.M.; Patel, S.N.; Tollefsbol, T.O. Sulforaphane Causes Epigenetic Repression of hTERT Expression in Human Breast Cancer Cell Lines. PLoS ONE 2010, 5, e11457. [Google Scholar] [CrossRef]
- Gao, M.; Li, G.; Yang, B.; McCombie, W.R.; Quiros, C.F. Comparative analysis of a Brassica BAC clone containing several major aliphatic glucosinolate genes with its corresponding Arabidopsis sequence. Genome 2004, 47, 666–679. [Google Scholar] [CrossRef] [PubMed]
- Yu, Z.; Li, S.; Sun, L. Progressin Studieson Genetic Engineering of Secondary Metabolitesin Plant. J. Jilin Agric. Sci. 2010, 35, 13–573. [Google Scholar]
- Mittler, R. Oxidative stress, antioxidants and stress tolerance. Trends Plant Sci. 2002, 7, 405–410. [Google Scholar] [CrossRef] [PubMed]
- Matusheski, N.V.; Jeffery, E.H. Comparison of the bioactivity of two glucoraphanin hydrolysis products found in broccoli, sulforaphane and sulforaphane nitrile. J. Agric. Food Chem. 2001, 49, 5743–5749. [Google Scholar] [CrossRef]
- Zang, Y.X.; Kim, H.U.; Kim, J.A.; Lim, M.H.; Jin, M.; Lee, S.C.; Kwon, S.J.; Lee, S.I.; Hong, J.K.; Park, T.H. Genome-wide identification of glucosinolate synthesis genes in Brassica rapa. Febs J. 2010, 276, 3559–3574. [Google Scholar] [CrossRef]
- Halkier, B.A.; Du, L. The biosynthesis of glucosinolates. Trends Plant Sci. 1997, 2, 425–431. [Google Scholar] [CrossRef]
- Golldack, D.; Su, H.; Quigley, F.; Kamasani, U.R.; Pantoja, O. Characterization of a HKT-type transporter in rice as a general alkali cation transporter. Plant J. 2010, 31, 529–542. [Google Scholar] [CrossRef]
- Wu, X.; Su, N.; Yue, X.; Fang, B.; Cui, J. IRT1 and ZIP2 were involved in exogenous hydrogen-rich water-reduced cadmium accumulation in Brassica chinensis and Arabidopsis thaliana. J. Hazard. Mater. 2021, 407, 124599. [Google Scholar] [CrossRef]
- Guo, L.; Yang, R.; Wang, Z.; Gu, Z. Effect of freezing methods on sulforaphane formation in broccoli sprouts. RSC Adv. 2015, 5, 3229–32297. [Google Scholar] [CrossRef]
- Lister, C.E.; And, J.E.L.; Walker, J.R.L. Developmental Changes in Enzymes of Flavonoid Biosynthesis in the Skins of Red and Green Apple Cultivars. J. Sci. Food Agr. 1996, 1046, 58–64. [Google Scholar] [CrossRef]
- Leon, J.; Yalpani, N.; Raskin, I.; Lawton, M.A. Induction of Benzoic Acid 2-Hydroxylase in Virus-Inoculated Tobacco. Plant Physiol. 1993, 23, 286. [Google Scholar] [CrossRef] [PubMed]
- Sudhakar, C.; Lakshmi, A.; Giridarakumar, S. Changes in the antioxidant enzyme efficacy in two high yielding genotypes of mulberry (Morus alba L.) under NaCl salinity. Plant Sci. 2001, 161, 613–619. [Google Scholar] [CrossRef]
- Singh, V.P.; Kumar, J.; Singh, S.; Prasad, S.M. Dimethoate modifies enhanced UV-B effects on growth, photosynthesis and oxidative stress in mung bean (Vigna radiata L.) seedlings: Implication of salicylic acid. Pestic. Biochem. Phys. 2014, 116, 13–23. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Niu, J.; Xiao, M.; Cai, Z. Graphene oxide-SiO2 nanocomposite as the adsorbent for extraction and preconcentration of plant hormones for HPLC analysis. J. Chromatogr. B 2017, 1046, 58–64. [Google Scholar] [CrossRef] [PubMed]
- Flatschacher, D.; Speckbacher, V.; Zeilinger, S. qRAT: An R-based stand-alone application for relative expression analysis of RT-qPCR data. Bmc Bioinform. 2022, 23, 286. [Google Scholar] [CrossRef]
- Zhou, C.; Zhu, Y.; Luo, Y. Effects of Sulfur Fertilization on the Accumulation of Health-Promoting Phytochemicals in Radish Sprouts. J. Agr. Food Chem. 2013, 61, 7552–7559. [Google Scholar] [CrossRef]
- Nestle, M. Broccoli sprouts as inducers of carcinogen-detoxifying enzyme systems: Clinical, dietary, and policy implications. Proc. Natl. Acad. Sci. USA 1997, 94, 11149–11151. [Google Scholar] [CrossRef]
- Ji, H.; Tang, W.; Zhou, X.; Wu, Y. Combined Effects of Blue and Ultraviolet Lights on the Accumulation of Flavonoids in Tartary Buckwheat Sprouts. Pol. J. Food Nutr. Sci. 2016, 66, 93–98. [Google Scholar] [CrossRef]
- Christie, J.M.; Briggs, W.R. Blue Light Sensing in Higher Plants. J. Biol. Chem. 2001, 276, 11457–11460. [Google Scholar] [CrossRef]
- Liu, Y.F.; Qi, M.F.; Li, T.L. Photosynthesis, photoinhibition, and antioxidant system in tomato leaves stressed by low night temperature and their subsequent recovery. Plant Sci. 2012, 196, 8–17. [Google Scholar] [CrossRef]
- Yamada, K.; Hasegawa, T.; Minami, E.; Shibuya, N.; Kosemura, S.; Yamamura, S.; Hasegawa, K. Induction of myrosinase gene expression and myrosinase activity in radish hypocotyls by phototropic stimulation. J. Plant Physiol. 2003, 160, 255–259. [Google Scholar] [CrossRef] [PubMed]
- Stine, H.; Anna, K.; Bok-Rye, L.; Shikha, S.; Richard, M.; Anne-Berit, W.; Bengtsson, G.B.; Stanislav, K. Diurnal and light regulation of sulphur assimilation and glucosinolate biosynthesis in Arabidopsis. J. Exp. Bot. 2013, 64, 1039–1048. [Google Scholar]
- Go-Eun, Y.; Arif, R.; Kiwoung, Y.; Jong-In, P.; Byung, H.; Ill-Sup, N. Exogenous Methyl Jasmonate and Salicylic Acid Induce Subspecies-Specific Patterns of Glucosinolate Accumulation and Gene Expression in Brassica oleracea L. Molecules 2016, 2016, 1417. [Google Scholar]
- Pucci, A.; Picarella, M.E.; Mazzucato, A. Phenotypic, genetic and molecular characterization of 7B-1, a conditional male-sterile mutant in tomato. Theor. Appl. Genet. 2017, 130, 2361–2374. [Google Scholar] [CrossRef] [PubMed]
- Kurepin, L.V.; Walton, L.J.; Reid, D.M.; Chinnappa, C.C. Light regulation of endogenous salicylic acid levels in hypocotyls of Helianthus annuus seedlings. Botany 2010, 88, 904–910. [Google Scholar] [CrossRef]
- Pérez-Balibrea, S.; Fernández, D.A.M.; Viguera, C.G. Influence of light on health-promoting phytochemicals of broccoli sprouts. J. Sci. Food Agric. 2010, 88, 904–910. [Google Scholar] [CrossRef]
- Frerigmann, H.; Gigolashvili, T. MYB34, MYB51, and MYB122 distinctly regulate indolic glucosinolate biosynthesis in Arabidopsis thaliana. Mol. Plant 2014, 7, 814–828. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer Sequence (5′ to 3′) | |
|---|---|---|
| Forward | Reverse | |
| Actin | CTGTTCCAATCTACGAGGGTTTCT | GCTCGGCTGTGGTGGTGAA |
| BoElong | AAGGTCGTCTGAAAGAGTTGGG | TGATTTCGTTGTCGTTAGTGCC |
| BoCYP83A1 | CAAACGCTACAAACTGCC | GTGGGTGAGAACAAGTGG |
| BoUGT74B1 | CAAAGACGATAAAGGCTACGGC | TCCCAAAGGAACCAAACGAA |
| BoST5b | CTGATACAGCCTCGCATTG | GAGGGTTTGTGGAATCGTTG |
| BoMYR | ACACGAGATGGCAGAAAG | GACCTCCTTGGTTCACTC |
| BoESP | AAGAGGGAGGACCCGAGGCT | TCCTTTGCTCACTCCACC |
| BoPAL | AGCAGCGGAACAGATGAA | ACTCCCTTTCATCTGTTCC |
| BoBA2H | GCCCTTGTGGTAGCGAAAT | TTGCTCAACACCAGGAAG |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guo, Y.; Gong, C.; Cao, B.; Di, T.; Xu, X.; Dong, J.; Zhao, K.; Gao, K.; Su, N. Blue Light Enhances Health-Promoting Sulforaphane Accumulation in Broccoli (Brassica oleracea var. italica) Sprouts through Inhibiting Salicylic Acid Synthesis. Plants 2023, 12, 3151. https://doi.org/10.3390/plants12173151
Guo Y, Gong C, Cao B, Di T, Xu X, Dong J, Zhao K, Gao K, Su N. Blue Light Enhances Health-Promoting Sulforaphane Accumulation in Broccoli (Brassica oleracea var. italica) Sprouts through Inhibiting Salicylic Acid Synthesis. Plants. 2023; 12(17):3151. https://doi.org/10.3390/plants12173151
Chicago/Turabian StyleGuo, Youyou, Chunyan Gong, Beier Cao, Tiantian Di, Xinxin Xu, Jingran Dong, Keying Zhao, Kai Gao, and Nana Su. 2023. "Blue Light Enhances Health-Promoting Sulforaphane Accumulation in Broccoli (Brassica oleracea var. italica) Sprouts through Inhibiting Salicylic Acid Synthesis" Plants 12, no. 17: 3151. https://doi.org/10.3390/plants12173151