
Unveiling Plant-Based Pectins: Exploring the Interplay of Direct Effects, Fermentation, and Technological Applications in Clinical Research with a Focus on the Chemical Structure

 ,
,  ,
,  and
and 
Abstract
:1. Introduction
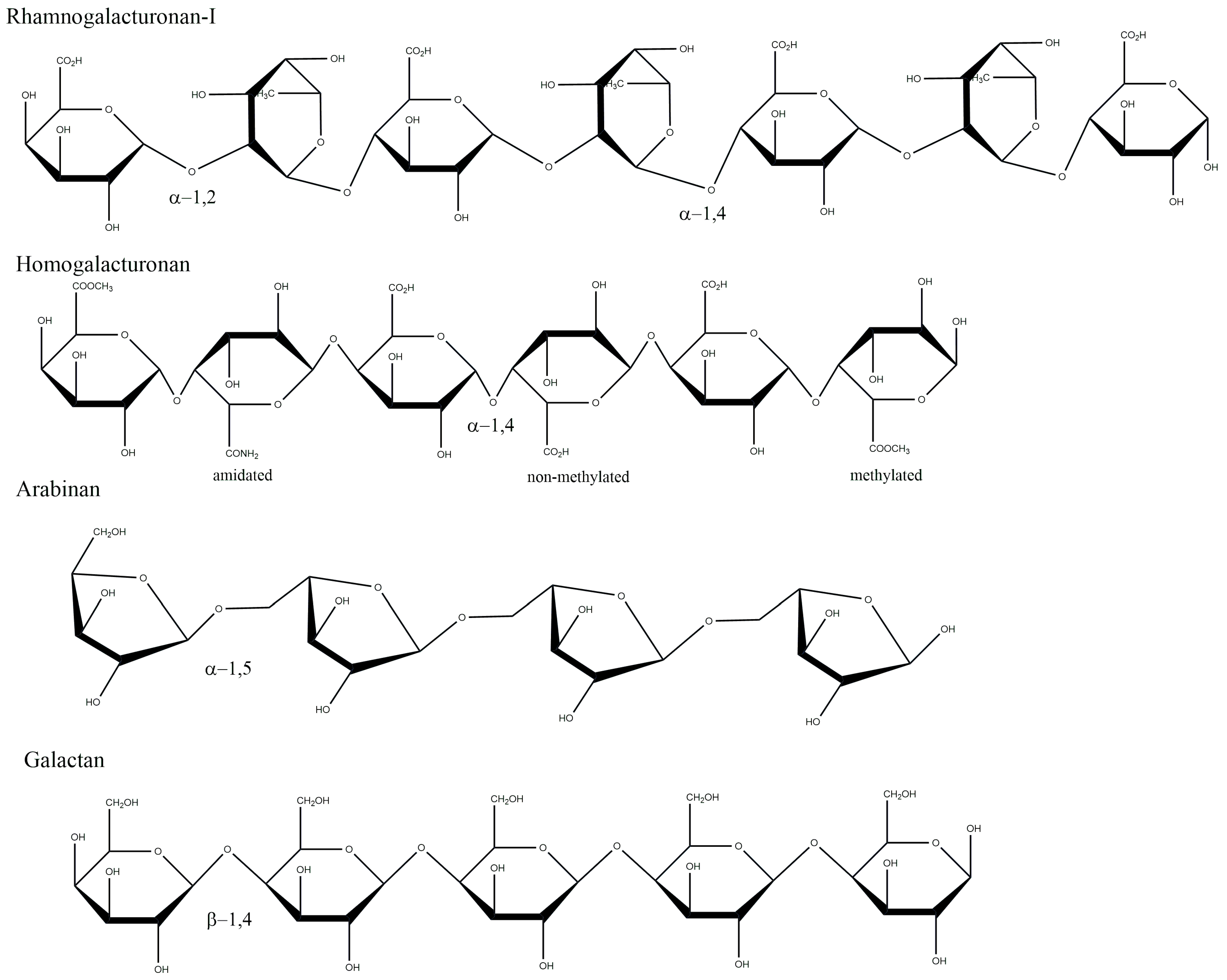
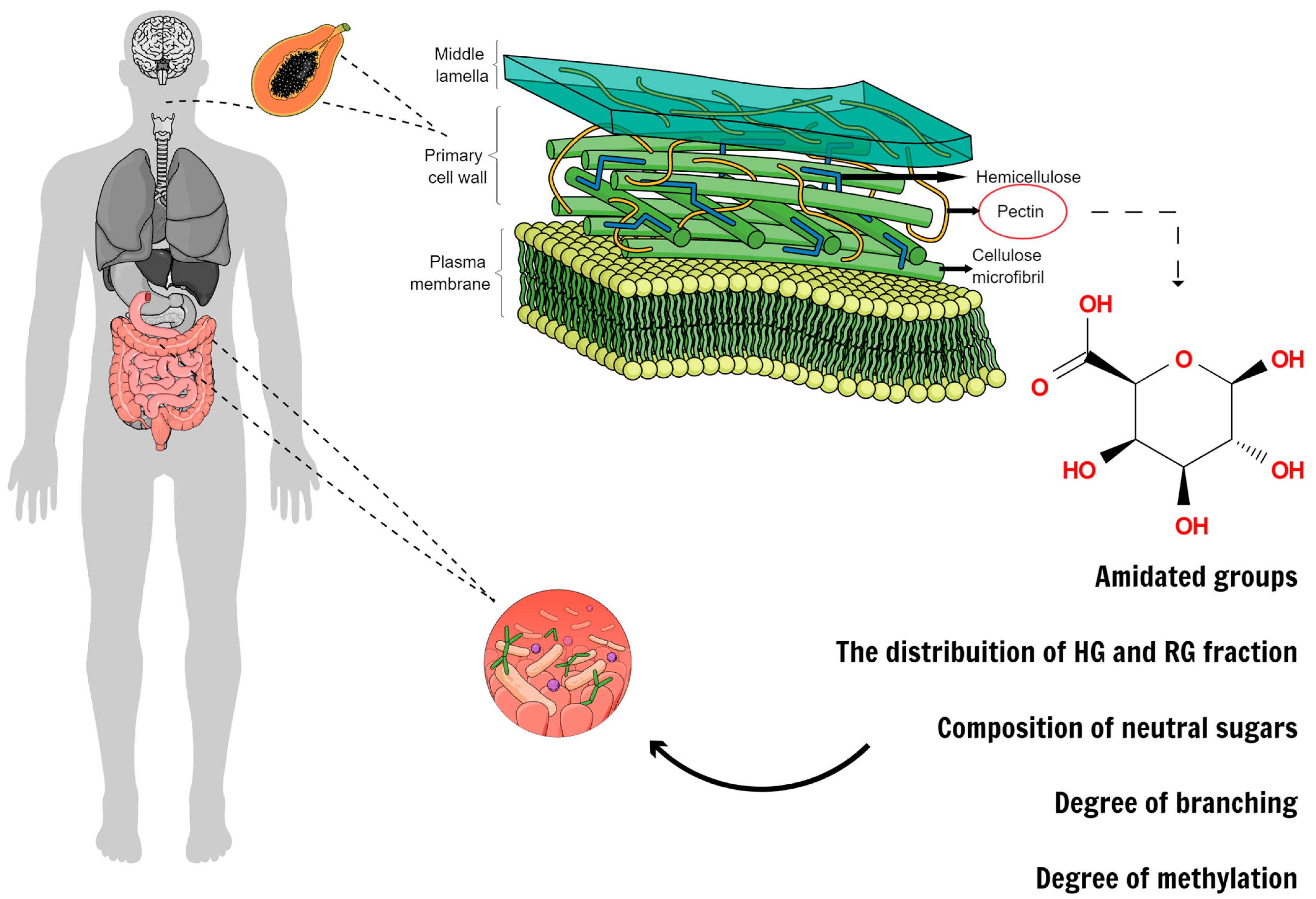
2. Chemical Structure
2.1. Backbone Structure of Pectin and Ramifications
2.2. Degree of Methyl-Esterification and Degree of Blockiness
2.3. Molecular Weight and Structure Modification Procedures
2.4. Low Molecular Weight Pectins for High-End Applications
3. In Vitro Exploration of Pectin Biological Effects
3.1. Cytotoxicity Effects of Native and Modified Pectins on Cancer Cells
3.2. Gut Microbiota Affects Pectin Fermentation
3.3. Pectin Beneficial Immunomodulation
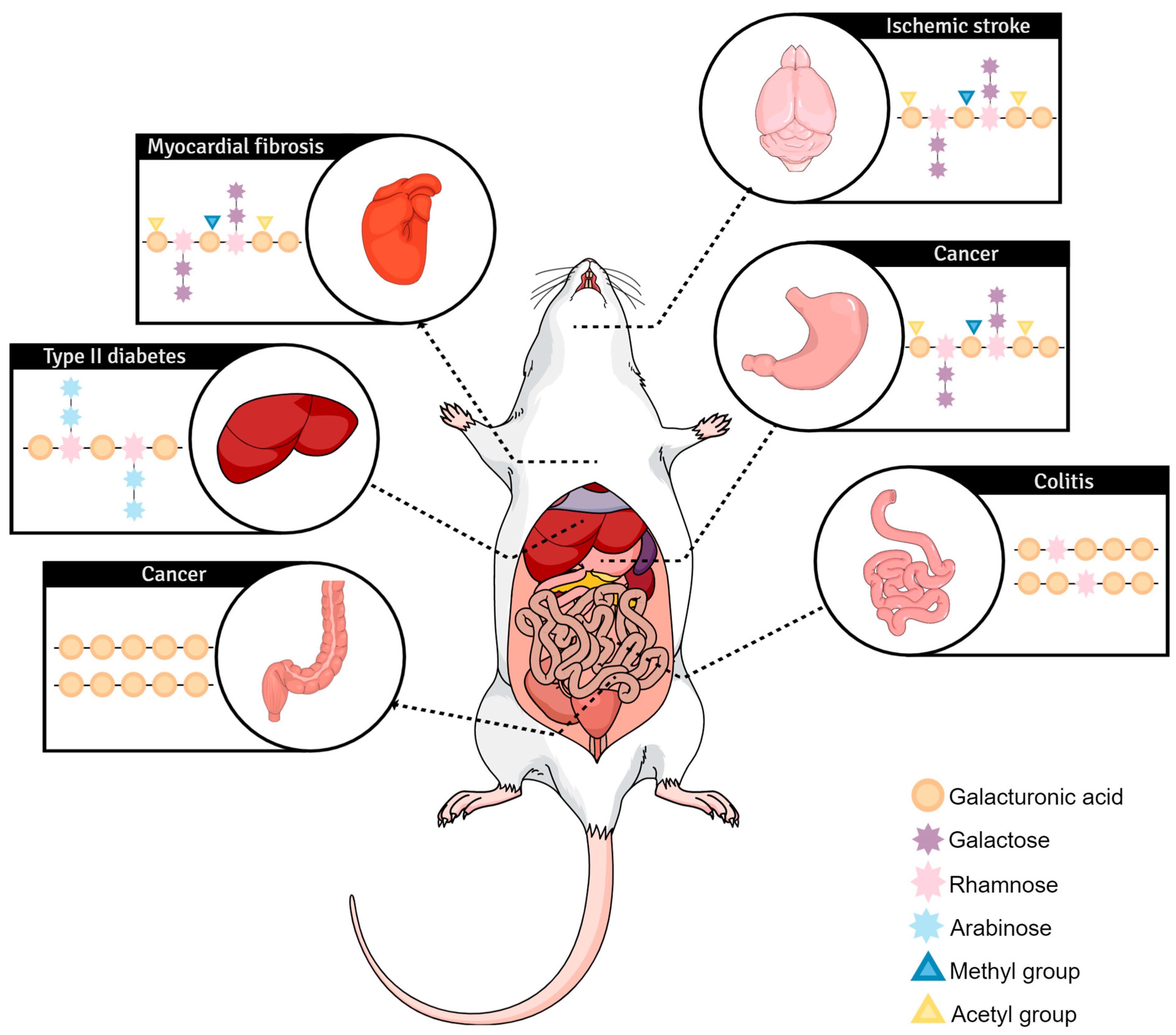
4. In Vivo Exploration of Pectin Biological Effects
5. As Close as It Gets: Clinical and Preclinical Advances to Indicate the Best Plant-Derived Pectin

{kind=link}
{kind=link}
{kind=link}
| Clinical Hypothesis | Type of Pectin Used | Characteristics of Pectin Used | Study Design | Study Population | N° of Participants | Summary of Results |
|---|---|---|---|---|---|---|
| Direct inhibition of Gal-3 may reduce subclinical cardiac fibrosis | MCP (PectaSol) by EcoNugenics, Inc. | Low molecular weight, DE under 5%, approximately 10% RG-II and enzymatic modification, | Randomized placebo-controlled | USA | 52 | Demonstrated a higher baseline Gal-3 levels in women compared with men, validated previous associations of Gal-3 with clinical factors. Did not influence surrogate measures of cardiac fibrosis, including echocardiographic [102] |
| Evaluate the effect of MCP in men with prostate cancer | MCP (PectaSol) by EcoNugenics, Inc. | Low molecular weight, DE under 5%, approximately 10% RG-II and enzymatic modification, | Prospective phase II study | USA | 59 | The results suggested that MCP in BRPC has a potential benefit, as evidenced by changes in prostate-specific antigen doubling time (PSADT) and lower-than-expected rates of disease progression compared to historical data. The exceptionally low incidence of toxicities was a promising result found as well [103]. |
| Bananas and pectin seem to be potential therapeutic agents for diarrhea | Green banana and pectin (Sigma, St. Louis, MO, USA) | Not applicable | Double-blind controlled | Bangladesh | 62 | Almost all children in the banana and pectin group had recovered from clinical diarrhea. Pectin is at least as effective as banana or has slightly better effects on stool volume and quality, although more expensive [104]. |
| The cholesterol-lowering properties of the viscous fiber pectin may depend on its physicochemical Properties | Commercial and experimental pectins (PEC1 and PEC2) * | PEC1 corresponded to 15 g/day of cellulose (control). PEC2 only corresponded to capsules that were reduced to 0.325 g of pectin if compared with PEC1. | Randomized crossover | The Netherlands | 60 | Pectin source and type affect cholesterol lowering. The efficacy of pectin to reduce cholesterol was dependent on the physicochemical properties and improved with MW (molecular weight) and DE (degree of esterification) [105]. |
| To examine and verify the clinical effects of POF | Pectin-containing oligomeric formula (POF) | Pectin content, 0.9 g. | Randomized | Japan | 198 | POF is less likely to cause enteral nutrition (EN) related events, such as diarrhea, than standard polymeric formula (SPF) [106]. |
| To investigate the effects of pectins supplementation on gastrointestinal barrier function | GENU® BETA pectin | Free-flowing powder, Particle size: Less than 1% gum on a 0.250 mm test sieve and DE typically 55% | Randomized, double-blind, placebo-controlled | The Netherlands | 97 | With 4 weeks of intervention, intestinal barrier function was not affected by pectin supplementation for both groups (healthy adults and elderly people) [107]. |
| MCP could control cancer growth and tumor metastasis | MCP | DE below 20%, pH modified by partial neutralization as Potassium/Sodium salt. | Pilot trial | Germany | 49 | The treatment with MCP did not induce antitumoral responses. However, a 50% decrease in the serum PSA level after 16 weeks of treatment, but more studies will be necessary [108]. |
6. Trending Technologies for Biomedical Applications
7. Final Thoughts
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Pedrosa, L.D.F.; Fabi, J.P. Dietary Fiber as a Wide Pillar of Colorectal Cancer Prevention and Adjuvant Therapy. Crit. Rev. Food Sci. Nutr. 2023, 1–21. [Google Scholar] [CrossRef]
- Melelli, A.; Arnould, O.; Beaugrand, J.; Bourmaud, A. The Middle Lamella of Plant Fibers Used as Composite Reinforcement: Investigation by Atomic Force Microscopy. Molecules 2020, 25, 632. [Google Scholar] [CrossRef] [Green Version]
- Do Prado, S.B.R.; Castro-Alves, V.C.; Ferreira, G.F.; Fabi, J.P. Ingestion of Non-Digestible Carbohydrates from Plant-Source Foods and Decreased Risk of Colorectal Cancer: A Review on the Biological Effects and the Mechanisms of Action. Front. Nutr. 2019, 6, 72. [Google Scholar] [CrossRef]
- Braconnot, H. Recherches Sur Un Nouvel Acide Universellement Répandu Dans Tous Les Végétaux. In Annales de Chemie et Physique; Crochard: Paris, France, 1825; pp. 173–178. ISBN 9781119130536. [Google Scholar]
- Pérez-Pérez, Y.; Carneros, E.; Berenguer, E.; Solís, M.T.; Bárány, I.; Pintos, B.; Gómez-Garay, A.; Risueño, M.C.; Testillano, P.S. Pectin De-Methylesterification and AGP Increase Promote Cell Wall Remodeling and Are Required during Somatic Embryogenesis of Quercus Suber. Front. Plant Sci. 2019, 9, 1915. [Google Scholar] [CrossRef] [PubMed]
- Ciriminna, R.; Fidalgo, A.; Scurria, A.; Ilharco, L.M.; Pagliaro, M. Pectin: New Science and Forthcoming Applications of the Most Valued Hydrocolloid. Food Hydrocoll. 2022, 127, 107483. [Google Scholar] [CrossRef]
- Fabi, J.P.; Broetto, S.G.; Da Silva, S.L.G.L.; Zhong, S.; Lajolo, F.M.; Do Nascimento, J.R.O. Analysis of Papaya Cell Wall-Related Genes during Fruit Ripening Indicates a Central Role of Polygalacturonases during Pulp Softening. PLoS ONE 2014, 9, e105685. [Google Scholar] [CrossRef] [Green Version]
- Wan, L.; Chen, Q.; Huang, M.; Liu, F.; Pan, S. Physiochemical, Rheological and Emulsifying Properties of Low Methoxyl Pectin Prepared by High Hydrostatic Pressure-Assisted Enzymatic, Conventional Enzymatic, and Alkaline de-Esterification: A Comparison Study. Food Hydrocoll. 2019, 93, 146–155. [Google Scholar] [CrossRef]
- Do Nascimento, G.E.; Simas-Tosin, F.F.; Iacomini, M.; Gorin, P.A.J.; Cordeiro, L.M.C. Rheological Behavior of High Methoxyl Pectin from the Pulp of Tamarillo Fruit (Solanum betaceum). Carbohydr. Polym. 2016, 139, 125–130. [Google Scholar] [CrossRef]
- Chen, J.; Cheng, H.; Zhi, Z.; Zhang, H.; Linhardt, R.J.; Zhang, F.; Chen, S.; Ye, X. Extraction Temperature Is a Decisive Factor for the Properties of Pectin. Food Hydrocoll. 2021, 112, 106160. [Google Scholar] [CrossRef]
- Da Silva, M.P.; Rosales, T.K.O.; Pedrosa, L.D.F.; Fabi, J.P. Creation of a New Proof-of-Concept Pectin/Lysozyme Nanocomplex as Potential β-Lactose Delivery Matrix: Structure and Thermal Stability Analyses. Food Hydrocoll. 2023, 134, 1081011. [Google Scholar] [CrossRef]
- Mohamed, J.M.; Alqahtani, A.; Ahmad, F.; Krishnaraju, V.; Kalpana, K. Pectin Co-Functionalized Dual Layered Solid Lipid Nanoparticle Made by Soluble Curcumin for the Targeted Potential Treatment of Colorectal Cancer. Carbohydr. Polym. 2021, 252, 117180. [Google Scholar] [CrossRef] [PubMed]
- Deshmukh, R.; Harwansh, R.K.; Das Paul, S.; Shukla, R. Controlled Release of Sulfasalazine Loaded Amidated Pectin Microparticles through Eudragit S 100 Coated Capsule for Management of Inflammatory Bowel Disease. J. Drug Deliv. Sci. Technol. 2020, 55, 101495. [Google Scholar] [CrossRef]
- Kumar, A.; Gulati, M.; Singh, S.K.; Gowthamarajan, K.; Prashar, R.; Mankotia, D.; Gupta, J.P.; Banerjee, M.; Sinha, S.; Awasthi, A.; et al. Effect of Co-Administration of Probiotics with Guar Gum, Pectin and Eudragit S100 Based Colon Targeted Mini Tablets Containing 5-Fluorouracil for Site Specific Release. J. Drug Deliv. Sci. Technol. 2020, 60, 102004. [Google Scholar] [CrossRef]
- Shahin, L.; Zhang, L.; Mohnen, D.; Urbanowicz, B.R. Insights into Pectin O-Acetylation in the Plant Cell Wall: Structure, Synthesis, and Modification. Cell Surf. 2023, 9, 100099. [Google Scholar] [CrossRef] [PubMed]
- Cui, L.; Wang, J.; Huang, R.; Tan, Y.; Zhang, F.; Zhou, Y.; Sun, L. Analysis of Pectin from Panax Ginseng Flower Buds and Their Binding Activities to Galectin-3. Int. J. Biol. Macromol. 2019, 128, 459–467. [Google Scholar] [CrossRef]
- Beukema, M.; Faas, M.M.; de Vos, P. The Effects of Different Dietary Fiber Pectin Structures on the Gastrointestinal Immune Barrier: Impact via Gut Microbiota and Direct Effects on Immune Cells. Exp. Mol. Med. 2020, 52, 1364–1376. [Google Scholar] [CrossRef]
- Pedrosa, L.D.F.; Raz, A.; Fabi, J.P. The Complex Biological Effects of Pectin: Galectin-3 Targeting as Potential Human Health Improvement? Biomolecules 2022, 12, 289. [Google Scholar] [CrossRef]
- Pabst, M.; Fischl, R.M.; Brecker, L.; Morelle, W.; Fauland, A.; Köfeler, H.; Altmann, F.; Léonard, R. Rhamnogalacturonan II Structure Shows Variation in the Side Chains Monosaccharide Composition and Methylation Status within and across Different Plant Species. Plant J. 2013, 76, 61–72. [Google Scholar] [CrossRef] [Green Version]
- Huang, Z.; Boekhorst, J.; Fogliano, V.; Capuano, E.; Wells, J.M. Distinct Effects of Fiber and Colon Segment on Microbiota-Derived Indoles and Short-Chain Fatty Acids. Food Chem. 2022, 398, 133801. [Google Scholar] [CrossRef]
- Ezzati, S.; Ayaseh, A.; Ghanbarzadeh, B.; Heshmati, M.K. Pectin from Sunflower By-Product: Optimization of Ultrasound-Assisted Extraction, Characterization, and Functional Analysis. Int. J. Biol. Macromol. 2020, 165, 776–786. [Google Scholar] [CrossRef]
- Santos, E.E.; Amaro, R.C.; Bustamante, C.C.C.; Guerra, M.H.A.; Soares, L.C.; Froes, R.E.S. Extraction of Pectin from Agroindustrial Residue with an Ecofriendly Solvent: Use of FTIR and Chemometrics to Differentiate Pectins According to Degree of Methyl Esterification. Food Hydrocoll. 2020, 107, 105921. [Google Scholar] [CrossRef]
- Kyomugasho, C.; Christiaens, S.; Shpigelman, A.; Van Loey, A.M.; Hendrickx, M.E. FT-IR Spectroscopy, a Reliable Method for Routine Analysis of the Degree of Methylesterification of Pectin in Different Fruit- and Vegetable-Based Matrices. Food Chem. 2015, 176, 82–90. [Google Scholar] [CrossRef]
- Gawkowska, D.; Cybulska, J.; Zdunek, A. Structure-Related Gelling of Pectins and Linking with Other Natural Compounds: A Review. Polymers 2018, 10, 762. [Google Scholar] [CrossRef] [Green Version]
- Da Silva, M.P.; Fabi, J.P. Food Biopolymers-Derived Nanogels for Encapsulation and Delivery of Biologically Active Compounds: A Perspective Review. Food Hydrocoll. Health 2022, 2, 100079. [Google Scholar] [CrossRef]
- Einhorn-stoll, U.; Archut, A.; Eichhorn, M.; Kastner, H. Food Hydrocolloids Pectin-Plant Protein Systems and Their Application. Food Hydrocoll. 2021, 118, 106783. [Google Scholar] [CrossRef]
- Yang, W.; Zhao, P.; Li, X.; Guo, L.; Gao, W. The Potential Roles of Natural Plant Polysaccharides in Inflammatory Bowel Disease: A Review. Carbohydr. Polym. 2022, 277, 118821. [Google Scholar] [CrossRef] [PubMed]
- Beukema, M.; Ishisono, K.; de Waard, J.; Faas, M.M.; de Vos, P.; Kitaguchi, K. Pectin Limits Epithelial Barrier Disruption by Citrobacter rodentium through Anti-Microbial Effects. Food Funct. 2021, 12, 881–891. [Google Scholar] [CrossRef] [PubMed]
- Haas, K.T.; Wightman, R.; Peaucelle, A.; Höfte, H. The Role of Pectin Phase Separation in Plant Cell Wall Assembly and Growth. Cell Surf. 2021, 7, 100054. [Google Scholar] [CrossRef] [PubMed]
- Soares, C.G.; Do Prado, S.B.R.; Andrade, S.C.S.; Fabi, J.P. Systems Biology Applied to the Study of Papaya Fruit Ripening: The Influence of Ethylene on Pulp Softening. Cells 2021, 10, 2339. [Google Scholar] [CrossRef]
- Vogt, L.M.; Sahasrabudhe, N.M.; Ramasamy, U.; Meyer, D.; Pullens, G.; Faas, M.M.; Venema, K.; Schols, H.A.; de Vos, P. The Impact of Lemon Pectin Characteristics on TLR Activation and T84 Intestinal Epithelial Cell Barrier Function. J. Funct. Foods 2016, 22, 398–407. [Google Scholar] [CrossRef]
- Chylińska, M.; Szymańska-Chargot, M.; Zdunek, A. FT-IR and FT-Raman Characterization of Non-Cellulosic Polysaccharides Fractions Isolated from Plant Cell Wall. Carbohydr. Polym. 2016, 154, 48–54. [Google Scholar] [CrossRef]
- Merheb, R.; Abdel-Massih, R.M.; Karam, M.C. Immunomodulatory Effect of Natural and Modified Citrus Pectin on Cytokine Levels in the Spleen of BALB/c Mice. Int. J. Biol. Macromol. 2019, 121, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Arias, D.; Rodríguez, J.; López, B.; Méndez, P. Evaluation of the Physicochemical Properties of Pectin Extracted from Musa Paradisiaca Banana Peels at Different PH Conditions in the Formation of Nanoparticles. Heliyon 2021, 7, e06059. [Google Scholar] [CrossRef] [PubMed]
- Mao, G.; Wu, D.; Wei, C.; Tao, W.; Ye, X.; Linhardt, R.J.; Orfila, C.; Chen, S. Reconsidering Conventional and Innovative Methods for Pectin Extraction from Fruit and Vegetable Waste: Targeting Rhamnogalacturonan I. Trends Food Sci. Technol. 2019, 94, 65–78. [Google Scholar] [CrossRef]
- Gutöhrlein, F.; Drusch, S.; Schalow, S. Extraction of Low Methoxylated Pectin from Pea Hulls via RSM. Food Hydrocoll. 2020, 102, 105609. [Google Scholar] [CrossRef]
- Liang, W.L.; Liao, J.S.; Qi, J.R.; Jiang, W.X.; Yang, X.Q. Physicochemical Characteristics and Functional Properties of High Methoxyl Pectin with Different Degree of Esterification. Food Chem. 2022, 375, 131806. [Google Scholar] [CrossRef]
- Cho, E.; Jung, H.; Lee, B.; Kim, H.; Rhee, J.; Yoo, S. Green Process Development for Apple-Peel Pectin Production by Organic Acid Extraction. Carbohydr. Polym. 2018, 204, 97–103. [Google Scholar] [CrossRef]
- Wikiera, A.; Grabacka, M.; Byczyński, Ł.; Stodolak, B.; Mika, M. Enzymatically Extracted Apple Pectin Possesses Antioxidant and Antitumor Activity. Molecules 2021, 26, 1434. [Google Scholar] [CrossRef]
- Wu, D.; Chen, S.; Ye, X.; Zheng, X.; Ahmadi, S.; Hu, W.; Yu, C.; Cheng, H.; Linhardt, R.J.; Chen, J. Enzyme-Extracted Raspberry Pectin Exhibits a High-Branched Structure and Enhanced Anti-Inflammatory Properties than Hot Acid-Extracted Pectin. Food Chem. 2022, 383, 132387. [Google Scholar] [CrossRef] [PubMed]
- Freitas, C.M.P.; Coimbra, J.S.R.; Souza, V.G.L.; Sousa, R.C.S. Structure and Applications of Pectin in Food, Biomedical, and Pharmaceutical Industry: A Review. Coatings 2021, 11, 922. [Google Scholar] [CrossRef]
- Zhang, L.; Ye, X.; Ding, T.; Sun, X.; Xu, Y.; Liu, D. Ultrasound Effects on the Degradation Kinetics, Structure and Rheological Properties of Apple Pectin. Ultrason. Sonochem. 2013, 20, 222–231. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Ma, X.; Xu, Y.; Cao, Y.; Jiang, Z.; Ding, T.; Ye, X.; Liu, D. Ultrasound-Assisted Heating Extraction of Pectin from Grapefruit Peel: Optimization and Comparison with the Conventional Method. Food Chem. 2015, 178, 106–114. [Google Scholar] [CrossRef] [PubMed]
- Hu, W.; Chen, S.; Wu, D.; Zheng, J.; Ye, X. Ultrasonic-Assisted Citrus Pectin Modification in the Bicarbonate-Activated Hydrogen Peroxide System: Chemical and Microstructural Analysis. Ultrason. Sonochem. 2019, 58, 104576. [Google Scholar] [CrossRef] [PubMed]
- Hao, M.; Yuan, X.; Cheng, H.; Xue, H.; Zhang, T.; Zhou, Y.; Tai, G. Comparative Studies on the Anti-Tumor Activities of High Temperature- and PH-Modified Citrus Pectins. Food Funct. 2013, 4, 960–971. [Google Scholar] [CrossRef]
- Chen, J.; Liang, R.H.; Liu, W.; Li, T.; Liu, C.M.; Wu, S.S.; Wang, Z.J. Pectic-Oligosaccharides Prepared by Dynamic High-Pressure Microfluidization and Their in Vitro Fermentation Properties. Carbohydr. Polym. 2013, 91, 175–182. [Google Scholar] [CrossRef]
- Donadio, J.L.S.; do Prado, S.B.R.; Rogero, M.M.; Fabi, J.P. Effects of Pectins on Colorectal Cancer: Targeting Hallmarks as a Support for Future Clinical Trials. Food Funct. 2022, 13, 11438–11454. [Google Scholar] [CrossRef]
- Maxwell, E.G.; Colquhoun, I.J.; Chau, H.K.; Hotchkiss, A.T.; Waldron, K.W.; Morris, V.J.; Belshaw, N.J. Modified Sugar Beet Pectin Induces Apoptosis of Colon Cancer Cells via an Interaction with the Neutral Sugar Side-Chains. Carbohydr. Polym. 2016, 136, 923–929. [Google Scholar] [CrossRef]
- Do Prado, S.B.R.; Shiga, T.M.; Harazono, Y.; Hogan, V.A.; Raz, A.; Carpita, N.C.; Fabi, J.P. Migration and Proliferation of Cancer Cells in Culture Are Differentially Affected by Molecular Size of Modified Citrus Pectin. Carbohydr. Polym. 2019, 211, 141–151. [Google Scholar] [CrossRef]
- Jermendi, É.; Fernández-Lainez, C.; Beukema, M.; López-Velázquez, G.; van den Berg, M.A.; de Vos, P.; Schols, H.A. TLR 2/1 Interaction of Pectin Depends on Its Chemical Structure and Conformation. Carbohydr. Polym. 2023, 303, 120444. [Google Scholar] [CrossRef]
- Emran, T.B.; Islam, F.; Mitra, S.; Paul, S.; Nath, N.; Khan, Z.; Das, R.; Chandran, D.; Sharma, R.; Mariana, C.; et al. Pectin: A Bioactive Food Polysaccharide with Cancer Preventive Potential. Molecules 2022, 27, 7405. [Google Scholar] [CrossRef]
- Do Prado, S.B.R.; Ferreira, G.F.; Harazono, Y.; Shiga, T.M.; Raz, A.; Carpita, N.C.; Fabi, J.P. Ripening-Induced Chemical Modifications of Papaya Pectin Inhibit Cancer Cell Proliferation. Sci. Rep. 2017, 7, 16564. [Google Scholar] [CrossRef] [Green Version]
- Ogutu, F.O.; Mu, T.H. Ultrasonic Degradation of Sweet Potato Pectin and Its Antioxidant Activity. Ultrason. Sonochem. 2017, 38, 726–734. [Google Scholar] [CrossRef] [PubMed]
- Palko-łabuz, A.; Maksymowicz, J.; Sobieszczańska, B.; Wikiera, A.; Skonieczna, M.; Wesołowska, O.; Środa-Pomianek, K. Newly Obtained Apple Pectin as an Adjunct to Irinotecan Therapy of Colorectal Cancer Reducing E. coli Adherence and β-Glucuronidase Activity. Cancers 2021, 13, 2952. [Google Scholar] [CrossRef] [PubMed]
- Bermúdez-Oria, A.; Rodríguez-Gutiérrez, G.; Alaiz, M.; Vioque, J.; Girón-Calle, J.; Fernández-Bolaños, J. Pectin-Rich Extracts from Olives Inhibit Proliferation of Caco-2 and THP-1 Cells. Food Funct. 2019, 10, 4844–4853. [Google Scholar] [CrossRef] [PubMed]
- Maxwell, E.G.; Colquhoun, I.J.; Chau, H.K.; Hotchkiss, A.T.; Waldron, K.W.; Morris, V.J.; Belshaw, N.J. Rhamnogalacturonan i Containing Homogalacturonan Inhibits Colon Cancer Cell Proliferation by Decreasing ICAM1 Expression. Carbohydr. Polym. 2015, 132, 546–553. [Google Scholar] [CrossRef]
- Fabi, J.P.; Cordenunsi, B.R.; Seymour, G.B.; Lajolo, F.M.; do Nascimento, J.R.O. Molecular Cloning and Characterization of a Ripening-Induced Polygalacturonase Related to Papaya Fruit Softening. Plant Physiol. Biochem. 2009, 47, 1075–1081. [Google Scholar] [CrossRef]
- Wang, S.; Li, P.; Lu, S.M.; Ling, Z.Q. Chemoprevention of Low-Molecular-Weight Citrus Pectin (Lcp) in Gastrointestinal Cancer Cells. Int. J. Biol. Sci. 2016, 12, 746–756. [Google Scholar] [CrossRef]
- Laaf, D.; Bojarová, P.; Elling, L.; Křen, V. Galectin–Carbohydrate Interactions in Biomedicine and Biotechnology. Trends Biotechnol. 2019, 37, 402–415. [Google Scholar] [CrossRef]
- Pascale, N.; Gu, F.; Larsen, N.; Jespersen, L.; Respondek, F. The Potential of Pectins to Modulate the Human Gut Microbiota Evaluated by In Vitro Fermentation: A Systematic Review. Nutrients 2022, 14, 3629. [Google Scholar] [CrossRef]
- Larsen, N.; De Souza, C.B.; Krych, L.; Cahú, T.B.; Wiese, M.; Kot, W.; Hansen, K.M.; Blennow, A.; Venema, K.; Jespersen, L. Potential of Pectins to Beneficially Modulate the Gut Microbiota Depends on Their Structural Properties. Front. Microbiol. 2019, 10, 223. [Google Scholar] [CrossRef] [Green Version]
- Cui, J.; Zhao, C.; Zhao, S.; Tian, G.; Wang, F.; Li, C.; Wang, F.; Zheng, J. Alkali + Cellulase-Extracted Citrus Pectins Exhibit Compact Conformation and Good Fermentation Properties. Food Hydrocoll. 2020, 108, 106079. [Google Scholar] [CrossRef]
- Min, B.; Kyung Koo, O.; Park, S.H.; Jarvis, N.; Ricke, S.C.; Crandall, P.G.; Lee, S.-O. Fermentation Patterns of Various Pectin Sources by Human Fecal Microbiota. Food Nutr. Sci. 2015, 6, 1103–1114. [Google Scholar] [CrossRef] [Green Version]
- Olano-Martin, E.; Gibson, G.R.; Rastall, R.A. Comparison of the in Vitro Bifidogenic Properties of Pectins and Pectic-Oligosaccharides. J. Appl. Microbiol. 2002, 93, 505–511. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di, R.; Vakkalanka, M.S.; Onumpai, C.; Chau, H.K.; White, A.; Rastall, R.A.; Yam, K.; Hotchkiss, A.T. Pectic Oligosaccharide Structure-Function Relationships: Prebiotics, Inhibitors of Escherichia Coli O157:H7 Adhesion and Reduction of Shiga Toxin Cytotoxicity in HT29 Cells. Food Chem. 2017, 227, 245–254. [Google Scholar] [CrossRef] [PubMed]
- Prado, S.B.R.; Beukema, M.; Jermendi, E.; Schols, H.A.; de Vos, P.; Fabi, J.P. Pectin Interaction with Immune Receptors Is Modulated by Ripening Process in Papayas. Sci. Rep. 2020, 10, 1629. [Google Scholar] [CrossRef] [Green Version]
- El-zayat, S.R.; Sibaii, H.; Mannaa, F.A. Toll-like Receptors Activation, Signaling, and Targeting: An Overview. Bull. Natl. Res. Cent. 2019, 43, 187. [Google Scholar] [CrossRef] [Green Version]
- Fitzgerald, K.A.; Kagan, J.C. Toll-like Receptors and the Control of Immunity. Cell 2020, 180, 1044–1066. [Google Scholar] [CrossRef]
- Tan, H.; Chen, W.; Liu, Q.; Yang, G.; Li, K. Pectin Oligosaccharides Ameliorate Colon Cancer by Regulating Oxidative Stress- and Inflammation-Activated Signaling Pathways. Front. Immunol. 2018, 9, 1504. [Google Scholar] [CrossRef] [Green Version]
- Hino, S.; Sonoyama, K.; Bito, H.; Kawagishi, H.; Aoe, S.; Morita, T. Low-Methoxyl Pectin Stimulates Small Intestinal Mucin Secretion Irrespective of Goblet Cell Proliferation and Is Characterized by Jejunum Muc2 Upregulation in Rats. J. Nutr. 2013, 143, 34–40. [Google Scholar] [CrossRef] [Green Version]
- Popov, S.V.; Markov, P.A.; Nikitina, I.R.; Petrishev, S.; Smirnov, V.; Ovodov, Y.S. Preventive Effect of a Peptic Polysaccharide of the Common Cranberry Vaccinium oxycoccos L. on Acetic Acid-Induced Colitis in Mice. World J. Gastroenterol. 2006, 12, 6646–6651. [Google Scholar] [CrossRef]
- De Freitas Pedrosa, L.; Lopes, R.G.; Fabi, J.P. The Acid and Neutral Fractions of Pectins Isolated from Ripe and Overripe Papayas Differentially Affect Galectin-3 Inhibition and Colon Cancer Cell Growth. Int. J. Biol. Macromol. 2020, 164, 2681–2690. [Google Scholar] [CrossRef] [PubMed]
- Do Nascimento, R.S.; de Freitas Pedrosa, F.; Diethelm, L.T.H.; Souza, T.; Shiga, T.M.; Fabi, J.P. The Purification of Pectin from Commercial Fruit Flours Results in a Jaboticaba Fraction That Inhibits Galectin-3 and Colon Cancer Cell Growth. Food Res. Int. 2020, 137, 109747. [Google Scholar] [CrossRef] [PubMed]
- Sahasrabudhe, N.M.; Beukema, M.; Tian, L.; Troost, B.; Scholte, J.; Bruininx, E.; Bruggeman, G.; van den Berg, M.; Scheurink, A.; Schols, H.A.; et al. Dietary Fiber Pectin Directly Blocks Toll-like Receptor 2-1 and Prevents Doxorubicin-Induced Ileitis. Front. Immunol. 2018, 9, 383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, S.; Kuwabara, R.; Beukema, M.; Ferrari, M.; de Haan, B.J.; Walvoort, M.T.C.; de Vos, P.; Smink, A.M. Low Methyl-Esterified Pectin Protects Pancreatic β-Cells against Diabetes-Induced Oxidative and Inflammatory Stress via Galectin-3. Carbohydr. Polym. 2020, 249, 116863. [Google Scholar] [CrossRef] [PubMed]
- Guan, Y.; Zhang, Z.; Yu, X.; Yan, J.; Zhou, Y.; Cheng, H.; Tai, G. Components of Heat-Treated Helianthus annuus L. Pectin Inhibit Tumor Growth and Promote Immunity in a Mouse CT26 Tumor Model. J. Funct. Foods 2018, 48, 190–199. [Google Scholar] [CrossRef]
- Cui, Y.; Zhang, N.N.; Wang, D.; Meng, W.H.; Chen, H.S. Modified Citrus Pectin Alleviates Cerebral Ischemia/Reperfusion Injury by Inhibiting NLRP3 Inflammasome Activation via TLR4/NF-ĸB Signaling Pathway in Microglia. J. Inflamm. Res. 2022, 15, 3369–3385. [Google Scholar] [CrossRef]
- Gehlken, C.P.g.; Rogier van der Velde, A.; Meijers, W.C.; Silljé, H.H.W.; Muntendam, P.; Dokter, M.M.; van Gilst, W.H.; Schols, H.A.; de Boer, R.A. Pectins from Various Sources Inhibit Galectin-3-Related Cardiac Fibrosis. Curr. Res. Transl. Med. 2022, 70, 103321. [Google Scholar] [CrossRef]
- Zhang, W.; Xu, P.; Zhang, H. Pectin in Cancer Therapy: A Review. Trends Food Sci. Technol. 2015, 44, 258–271. [Google Scholar] [CrossRef]
- Wang, L.; Zhao, L.; Gong, F.; Sun, C.; Du, D.D.; Yang, X.X.; Guo, X. Modified Citrus Pectin Inhibits Breast Cancer Development in Mice by Targeting Tumor-Associated Macrophage Survival and Polarization in Hypoxic Microenvironment. Acta Pharmacol. Sin. 2022, 43, 1556–1567. [Google Scholar] [CrossRef]
- Pienta, K.J.; Nailk, H.; Akhtar, A.; Yamazaki, K.; Replogle, T.S.; Lehr, J.; Donat, T.L.; Tait, L.; Hogan, V.; Raz, A. Inhibition of Spontaneous Metastasis in a Rat Prostate Cancer Model by Oral Administration of Modified Citrus Pectin. J. Natl. Cancer Inst. 1995, 87, 348–353. [Google Scholar] [CrossRef]
- Li, Y.; Liu, L.; Niu, Y.; Feng, J.; Sun, Y.; Kong, X.; Chen, Y.; Chen, X.; Gan, H.; Cao, S.; et al. Modified Apple Polysaccharide Prevents against Tumorigenesis in a Mouse Model of Colitis-Associated Colon Cancer: Role of Galectin-3 and Apoptosis in Cancer Prevention. Eur. J. Nutr. 2012, 51, 107–117. [Google Scholar] [CrossRef] [PubMed]
- Sabater, C.; Molina-Tijeras, J.A.; Vezza, T.; Corzo, N.; Montilla, A.; Utrilla, P. Intestinal Anti-Inflammatory Effects of Artichoke Pectin and Modified Pectin Fractions in the Dextran Sulfate Sodium Model of Mice Colitis. Artificial Neural Network Modelling of Inflammatory Markers. Food Funct. 2019, 10, 7793–7805. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ren, T.; Liu, F.; Wang, D.; Li, B.; Jiang, P.; Li, J.; Li, H.; Chen, C.; Wu, W.; Jiao, L. Rhamnogalacturonan-I Enriched Pectin from Steamed Ginseng Ameliorates Lipid Metabolism in Type 2 Diabetic Rats via Gut Microbiota and AMPK Pathway. J. Ethnopharmacol. 2023, 301, 115862. [Google Scholar] [CrossRef]
- Zhu, R.G.; Sun, Y.D.; Li, T.P.; Chen, G.; Peng, X.; Duan, W.B.; Zheng, Z.Z.; Shi, S.L.; Xu, J.G.; Liu, Y.H.; et al. Comparative Effects of Hawthorn (Crataegus pinnatifida Bunge) Pectin and Pectin Hydrolyzates on the Cholesterol Homeostasis of Hamsters Fed High-Cholesterol Diets. Chem. Biol. Interact. 2015, 238, 42–47. [Google Scholar] [CrossRef]
- Ismail, M.F.; Gad, M.Z.; Hamdy, M.A. Study of the Hypolipidemic Properties of Pectin, Garlic and Ginseng in Hypercholesterolemic Rabbits. Pharmacol. Res. 1999, 39, 157–166. [Google Scholar] [CrossRef]
- Munoz-Almagro, N.; Montilla, A.; Villamiel, M. Role of Pectin in the Current Trends towards Low-Glycaemic Food Consumption. Food Res. Int. 2021, 140, 109851. [Google Scholar] [CrossRef] [PubMed]
- Bai, Y.; Wu, P.; Wang, K.; Li, C.; Li, E.; Gilbert, R.G. Food Hydrocolloids Effects of Pectin on Molecular Structural Changes in Starch during Digestion. Food Hydrocoll. 2017, 69, 10–18. [Google Scholar] [CrossRef] [Green Version]
- Dang, G.; Wang, W.; Zhong, R.; Wu, W.; Chen, L.; Zhang, H. Pectin Supplement Alleviates Gut Injury Potentially through Improving Gut Microbiota Community in Piglets. Front. Microbiol. 2022, 13, 1069694. [Google Scholar] [CrossRef]
- Wen, X.; Zhong, R.; Dang, G.; Xia, B.; Wu, W.; Tang, S.; Tang, L.; Liu, L.; Liu, Z.; Chen, L.; et al. Pectin Supplementation Ameliorates Intestinal Epithelial Barrier Function Damage by Modulating Intestinal Microbiota in Lipopolysaccharide-Challenged Piglets. J. Nutr. Biochem. 2022, 109, 109107. [Google Scholar] [CrossRef]
- Li, J.; Wang, L.; Yang, K.; Zhang, G.; Li, S.; Gong, H.; Liu, M.; Dai, X. Structure Characteristics of Low Molecular Weight Pectic Polysaccharide and Its Anti-Aging Capability by Modulating the Intestinal Homeostasis. Carbohydr. Polym. 2023, 303, 120467. [Google Scholar] [CrossRef]
- Zhang, S.; Mao, Y.; Zhang, Z.; Li, Z.; Kong, C. Theranostics Pectin Supplement Significantly Enhanced the Anti-PD-1 Efficacy in Tumor-Bearing Mice Humanized with Gut Microbiota from Patients with Colorectal Cancer. Theranostics 2021, 11, 4155–4170. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Y.; Liu, Y.; He, Y.; Zhang, B.; Zhao, L.; Tian, S.; Wang, Q. Biomaterials Intestinal-Targeted Nanotubes-in-Microgels Composite Carriers for Capsaicin Delivery and Their Effect for Alleviation of Salmonella Induced Enteritis. Biomaterials 2022, 287, 121613. [Google Scholar] [CrossRef] [PubMed]
- Tanes, C.; Bittinger, K.; Gao, Y.; Bushman, F.D.; Lewis, J.D.; Wu, G.D. Article Role of Dietary Fiber in the Recovery of the Human Gut Microbiome and Its Metabolome Role of Dietary Fiber in the Recovery of the Human Gut Microbiome and Its Metabolome. Cell Host Microbe 2021, 29, 394–407.e5. [Google Scholar] [CrossRef]
- Reynolds, A.; Mann, J.; Cummings, J.; Winter, N.; Mete, E.; Morenga, L. Te Carbohydrate Quality and Human Health: A Series of Systematic Reviews and Meta-Analyses. Lancet 2019, 393, 434–445. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naqash, F.; Masoodi, F.A.; Rather, S.A.; Wani, S.M.; Gani, A. Emerging Concepts in the Nutraceutical and Functional Properties of Pectin—A Review. Carbohydr. Polym. 2017, 168, 227–239. [Google Scholar] [CrossRef]
- Kumar, A.; Chauhan, G.S. Extraction and Characterization of Pectin from Apple Pomace and Its Evaluation as Lipase (Steapsin) Inhibitor. Carbohydr. Polym. 2010, 82, 454–459. [Google Scholar] [CrossRef]
- Eliaz, I.; Weil, E.; Wilk, B. Integrative Medicine and the Role of Modified Citrus Pectin/Alginates in Heavy Metal Chelation and Detoxification—Five Case Reports. Forsch Komplementärmed 2007, 14, 358–364. [Google Scholar] [CrossRef]
- Eliaz, I.; Raz, A. Pleiotropic E Ff Ects of Modified Citrus Pectin. Nutrients 2019, 11, 2619. [Google Scholar] [CrossRef] [Green Version]
- Eliaz, I.; Hotchkiss, A.T.; Fishman, M.L.; Rode, D. The Effect of Modified Citrus Pectin on Urinary Excretion of Toxic Elements. Phyther. Res. 2006, 864, 859–864. [Google Scholar] [CrossRef]
- Zhao, Z.Y.; Liang, L.; Fan, X.; Yu, Z.; Hotchkiss, A.T.; Wilk, B.J.; Eliaz, I. The Role of Modified Citrus Pectin as an Effective Chelator of Lead in Children Hospitalized with Toxic Lead Levels. Altern. Ther. 2008, 14, 34–38. [Google Scholar]
- Lau, E.S.; Liu, E.; Paniagua, S.M.; Sarma, A.A.; Zampierollo, G.; López, B.; Díez, J.; Wang, T.J.; Ho, J.E. Galectin-3 Inhibition with Modified Citrus Pectin in Hypertension. JACC Basic Transl. Sci. 2021, 6, 12–21. [Google Scholar] [CrossRef]
- Keizman, D.; Frenkel, M.; Peer, A.; Kushnir, I.; Rosenbaum, E.; Sarid, D.; Leibovitch, I.; Mano, R.; Yossepowitch, O.; Margel, D.; et al. Modified Citrus Pectin Treatment in Non-Metastatic Biochemically Relapsed Prostate Cancer: Results of a Prospective Phase II Study. Nutrients 2021, 13, 4295. [Google Scholar] [CrossRef] [PubMed]
- Rabbani, G.H.; Teka, T.; Zaman, B.; Majid, N.; Khatun, M.; Fuchs, G.J. Clinical Studies in Persistent Diarrhea: Dietary Management with Green Banana or Pectin in Bangladeshi Children. Gastroenterology 2001, 121, 554–560. [Google Scholar] [CrossRef] [PubMed]
- Brouns, F.; Theuwissen, E.; Adam, A.; Bell, M.; Berger, A.; Mensink, R.P. Cholesterol-Lowering Properties of Different Pectin Types in Mildly Hyper-Cholesterolemic Men and Women. Eur. J. Clin. Nutr. 2012, 66, 591–599. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maruyama, M.; Goshi, S.; Kashima, Y.; Mizuhara, A.; HIgashiguchi, T. Clinical Effects of a Pectin-Containing Oligomeric Formula in Tube Feeding Patients: A Multicenter Randomized. Nutr. Clin. Pract. 2019, 35, 3–6. [Google Scholar] [CrossRef] [PubMed]
- Wilms, E.; Jonkers, D.M.A.E.; Savelkoul, H.F.J.; Elizalde, M.; Tischmann, L.; de Vos, P.; Masclee, A.A.M.; Troost, F.J. The Impact of Pectin Supplementation on Intestinal Barrier Function in Healthy Young Adults and Healthy Elderly. Nutrients 2019, 11, 1554. [Google Scholar] [CrossRef] [Green Version]
- Azémar, M.; Hildenbrand, B.; Haering, B.; Heim, M.E.; Unger, C. Clinical Benefit in Patients with Advanced Solid Tumors Treated with Modi Fi Ed Citrus Pectin: A Prospective Pilot Study. Clin. Med. 2007, 1, 73–80. [Google Scholar]
- Kedir, W.M.; Deresa, E.M.; Diriba, T.F. Heliyon Pharmaceutical and Drug Delivery Applications of Pectin and Its Modi Fi Ed Nanocomposites. Heliyon 2022, 8, e10654. [Google Scholar] [CrossRef]
- Mercadante, S.; Prestia, G.; Adile, C.; Casuccio, A.; Care, I.; Relief, P.; Care, S.; Maddalena, L. Intranasal Fentanyl Versus Fentanyl Pectin Nasal Spray for the Management of Breakthrough Cancer Pain in Doses Proportional to Basal Opioid Regimen. J. Pain 2014, 15, 602–607. [Google Scholar] [CrossRef] [Green Version]
- Portenoy, R.K.; Burton, A.W.; Gabrail, N.; Taylor, D.; Pectin, F. Fentanyl Pectin Nasal Spray (FPNS) in the Treatment of Breakthrough Cancer Pain. Pain 2010, 151, 617–624. [Google Scholar] [CrossRef]
- Watts, P.; Smith, A. PecSys: In Situ Gelling System for Optimised Nasal Drug Delivery. Expert Opin. Drug Deliv. 2009, 6, 543–552. [Google Scholar] [CrossRef] [PubMed]
- Rosales, T.K.O.; Fabi, J.P. Pectin-Based Nanoencapsulation Strategy to Improve the Bioavailability of Bioactive Compounds. Int. J. Biol. Macromol. 2023, 229, 11–21. [Google Scholar] [CrossRef] [PubMed]
- Rosales, T.K.O.; da Silva, M.P.; Lourenço, F.R.; Hassimotto, N.M.A.; Fabi, J.P. Nanoencapsulation of Anthocyanins from Blackberry (Rubus spp.) through Pectin and Lysozyme Self-Assembling. Food Hydrocoll. 2021, 114, 106563. [Google Scholar] [CrossRef]
- Morello, G.; Quarta, A.; Gaballo, A.; Moroni, L.; Gigli, G.; Polini, A.; Gervaso, F. A Thermo-Sensitive Chitosan/Pectin Hydrogel for Long-Term Tumor Spheroid Culture. Carbohydr. Polym. 2021, 274, 118633. [Google Scholar] [CrossRef] [PubMed]
- Zanoni, M.; Cortesi, M.; Zamagni, A.; Arienti, C.; Pignatta, S.; Tesei, A. Modeling Neoplastic Disease with Spheroids and Organoids. J. Hematol. Oncol. 2020, 13, 97. [Google Scholar] [CrossRef] [PubMed]



| Plant Material | Extraction/Modification Procedure | Resulted Structure | Reference |
|---|---|---|---|
| Tomato plants | Only water (WSP); different concentration imidazole (ISP); sodium carbonate and borohydride (DASP); potassium hydroxide (KOH) | WSP, ISP, and DASP all consisted of pectins, with DASP having RGI fragments; the KOH fraction was composed of xyloglucans, arabinoxylans, and galactans | [32] |
| Banana peel | Acetic, citric, hydrochloric (HCl), and nitric (HNO3) acids at pH 2 or 3 | Acetic, hydrochloric, and nitric acid resulted in LMPs with shorter chains, while citric acid resulted in HMs with longer chains at both pHs | [34] |
| Pea hulls | HNO3 or citric acid, under variable pH, temperature, and time conditions | DM ranged between 36 to 49%, with no differences between acids; higher pH (2) yielded more neutral sugars and less uronic acids, proportionally, for both acids | [36] |
| Commercial citrus pectin with 73% DM | De-esterification with variable NaOH concentrations, with hydrochloric acid pH adjustment | Besides lower DM, de-esterified pectin had lower molecular weights, zeta-potential, polydispersity index (PDI), and HG proportion while having higher RGI | [37] |
| Fuji apple-peel waste | Tartaric, malic, and citric acids, at 0.1 or 1.0 M | Malic and citric acids yielded higher DM pectins than tartaric (65–67% compared to 59%); higher concentrations seem to indicate a tendency of reduced DMs; Mw varied from 26 to 110 kDa | [38] |
| Dried apple-pomace | H2SO4, or endo-β-1,4-xylanase (Xyl) and endo-cellulase (Cel) | Acid-treated had the lowest Mw, GalA content, and DM; Cel + Xyl treatment resulted in a close Mw, with much higher GalA (74.7% × 59.9%) and DM (67.5 × 56.1%); Acid treatment also led to higher Glc and Ara, while Cel + Xyl presented more Gal, Rha, and Man | [39] |
| Raspberry | HCl or enzymatic hydrolysis (α-amylase, papain, cellulose), following step-wise alcohol precipitation | HCl-modified pectins had much lower Mw, PDI, and Xyl, with absent Man residues, while also having higher amounts of Gal and GalA. Other neutral sugars, such as Rha and Glu, were also higher in enzyme-extracted pectin | [40] |
| Commercial apple pectin (74%) | Ultrasound with amplitudes from 20 to 60% for 60 min at 25 °C and temperatures ranging from 5 to 45 °C for 30% sonication amplitude | Temperature variation seemed to decrease the Mw reduction (299 kDa × 361 kDa, comparing 5 to 45 °C); the lowest Mw and PDI values were obtained at 60 min; DM and RGI proportion was reduced by ultrasound intensity and time | [42] |
| Grapefruit peel | 50% Ultrasound amplitude in 2-s cycles (UP), with sporadic agitation; other fraction stood 90 min under 80 °C without ultrasound (CP) | Mw was slightly lower for the UP (109.54 × 132.01 kDa), as well as GalA, Glc, and Man residues; DM, as well as Rha, Gal, and Xyl residues, were comparable; Ara was much higher in UP (12.72 × 4.43 mol%) | [43] |
| Citrus peel | NaHCO3 in different concentrations and 30% H2O2 in water with ultrasound; compared to the same medium without ultrasound | The chemical + ultrasound group showed a 30-fold Mw reduction (33 × 1088 kDa), and the chemical group a 10-fold reduction (103 kDa); every monosaccharide proportion was comparable, besides a higher proportion of Fuc (1.81 × 0.52 mol%) and a lower GalA proportion (54.39 × 57.23 mol%) between groups; both had higher Gal contents than control | [44] |
| Commercial citrus pectin | Chemical modification through saponification with NaOH, acidification to pH 3.0 with HCl, and neutralization with NaOH (MCP); high temperature through heating in an autoclave for 121 °C for 30 min two times, on two different days (HTCP) | Acid treatment removed completely all methylations; the Mw of both were comparable (61 and 9 kDa peaks for MCP; 24 kDa for HTCP); all pectins had the same monosaccharide profile, mostly composed by GalA, with Rha, Gal, and Ara as secondary components | [45] |
| Commercial apple pectin | Solution pH was adjusted to H2SO4, and the solution was incubated in a microfluidizer; optimal variables were 155 MPa and 6 cycles | Almost 33% of the pectin content was converted into pectic oligosaccharides (POS); GalA content was greatly decreased in POS (29.56 × 71.68 wt%); Ara, Gal, Rha, Glc, and Xyl had a 3-fold increase in the POS sample | [46] |
| Pectin/Fragment Used | In Vitro Model | Main Results Found | Authors |
|---|---|---|---|
| Modified sugar beet pectin fragments | HT29 and DLD1 cell culture | Pectin stimulated apoptosis and detachment of HT29 cells; the galactan fragment was most efficient in lowering cell proliferation | [48] |
| Size-fractioned oligo- and polysaccharides from MCP | HCT116, HT29, and PC3 cell lines | MCP30/10 and 10/3 stimulated cytotoxicity through necroptosis and necrosis on the HCT116 cell line; smaller fragments significantly reduced homotypic aggregation and cell migration | [49] |
| Papaya pectin from different ripening points | HCT116, HT29, and PC3 cell line | Pectins extracted from the third day after harvesting inhibited cancer cell aggregation, migration, and further reduced viability while enhancing cytotoxicity; they also increased pAkt, pErk1/2, p21, and caspase-3 protein expression | [52] |
| Ultrasound-fragmented Sweet Potato pectin with high GalA content and low DM | Oxygen radical absorbance capacity and Ferric reducing antioxidant power assay (ORAC and FRAP) | The most fragmented pectin performed better in both antioxidant assays, starting in lower doses, compared to the other pectins | [53] |
| Enzymatically extracted apple pectin | HCT116 and Caco-2 cell culture | Pectins were able to synergize with Irinotecan increasing colorectal cancer cell cytotoxicity through apoptosis induction compared to drug-only controls; ROS levels increased; reduced IL-6 and COX-2 in LPS-induced HCT116 cells; pectins also reduced E. coli adherence to colorectal cancer cells | [54] |
| Acid and heat-treated pectic extracts from olives | Caco-2 and THP-1 cell culture | Higher doses (3.33 and 10 mg/mL) were able to cease cell proliferation completely up to seven days; pectin extracts successfully induced caspase-3 activity, compared to MCP and control | [55] |
| Varied potato pectins, citrus polygalacturonic acid, larch AG | HT29, DLD1, HCT116, LoVo; and Caco-2 cell culture | Potato RGI was the most effective in reducing cell proliferation in a dose-response manner; authors detected ICAM-1 downregulation and proposed reduction of proliferation was due to cell detachment | [56] |
| Low-molecular-weight citrus pectin | AGS and SW-480 cell culture | LCP lowered cell viability, proliferation, and cell cycle progression; the pectin had comparable values of Cyclin B1 and Bcl-xL downregulation to 5-FU | [58] |
| Papaya pectin from different ripening points | HEK-TLR reporter cell lines | The ripe-fruits pectins activated TLR3, 5, and 9; all pectin samples activated TLR2 and 4; only unripe-fruit pectins inhibited TLR3 and TLR9 | [66] |
| Acid and neutral fractions of papaya pectins from different ripening points | HCT116 and HT29 cell culture, recombinant human Gal-3 hemagglutination | The acid fraction from four days after harvest was able to inhibit Gal-3 hemagglutination from the second-lowest dose onward; cell viability was reduced mostly by full water-soluble fractions or acid fractions | [72] |
| Oligosaccharides from jaboticaba, plum, and papaya flours | HCT116 cell culture and recombinant human Gal-3 hemagglutination | Only jaboticaba oligosaccharides were able to inhibit Gal-3 hemagglutination and colorectal cancer cell viability | [73] |
| Lemon pectins with varying DMs | HEK-TLR reporter cell lines | Low-DM pectins inhibited TLR2 activity way more than higher-DM ones; specific TLR2-TLR1 heterodimer inhibition was observed | [74] |
| Lemon pectins with varying DMs | Human pancreatic islets and MIN6 cells | Low-DM pectins showed protective effects on human islets against streptozotocin and IL1b + IFN-y + TNF-a inflammatory stress through a Gal-3 binding-dependent way | [75] |
| High-temperature sunflower pectin and polygalacturonic acid | CT26 colorectal cancer cell line | Pectins were able to upregulate JNK, ERK, and p38 phosphorylation while down-regulating Akt time-dependently; dose-dependent stimulation of apoptosis was also observed in vitro | [76] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pedrosa, L.d.F.; Nascimento, K.R.; Soares, C.G.; Oliveira, D.P.d.; de Vos, P.; Fabi, J.P. Unveiling Plant-Based Pectins: Exploring the Interplay of Direct Effects, Fermentation, and Technological Applications in Clinical Research with a Focus on the Chemical Structure. Plants 2023, 12, 2750. https://doi.org/10.3390/plants12142750
Pedrosa LdF, Nascimento KR, Soares CG, Oliveira DPd, de Vos P, Fabi JP. Unveiling Plant-Based Pectins: Exploring the Interplay of Direct Effects, Fermentation, and Technological Applications in Clinical Research with a Focus on the Chemical Structure. Plants. 2023; 12(14):2750. https://doi.org/10.3390/plants12142750
Chicago/Turabian StylePedrosa, Lucas de Freitas, Karen Rebouças Nascimento, Caroline Giacomelli Soares, Débora Preceliano de Oliveira, Paul de Vos, and João Paulo Fabi. 2023. "Unveiling Plant-Based Pectins: Exploring the Interplay of Direct Effects, Fermentation, and Technological Applications in Clinical Research with a Focus on the Chemical Structure" Plants 12, no. 14: 2750. https://doi.org/10.3390/plants12142750