
The Tomato Feruloyl Transferase FHT Promoter Is an Accurate Identifier of Early Development and Stress-Induced Suberization
 , , and
, , and
Abstract
:1. Introduction
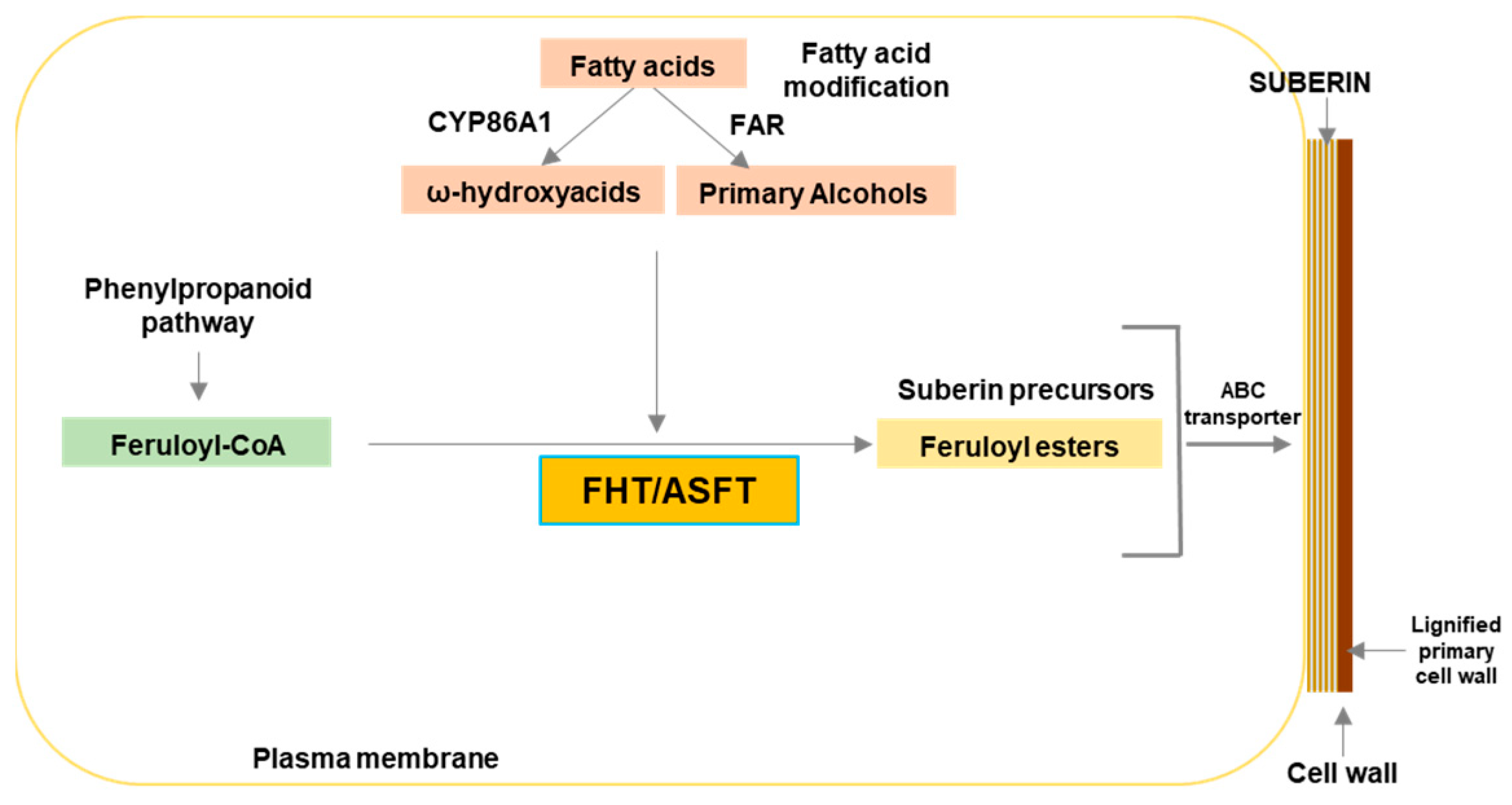
2. Results
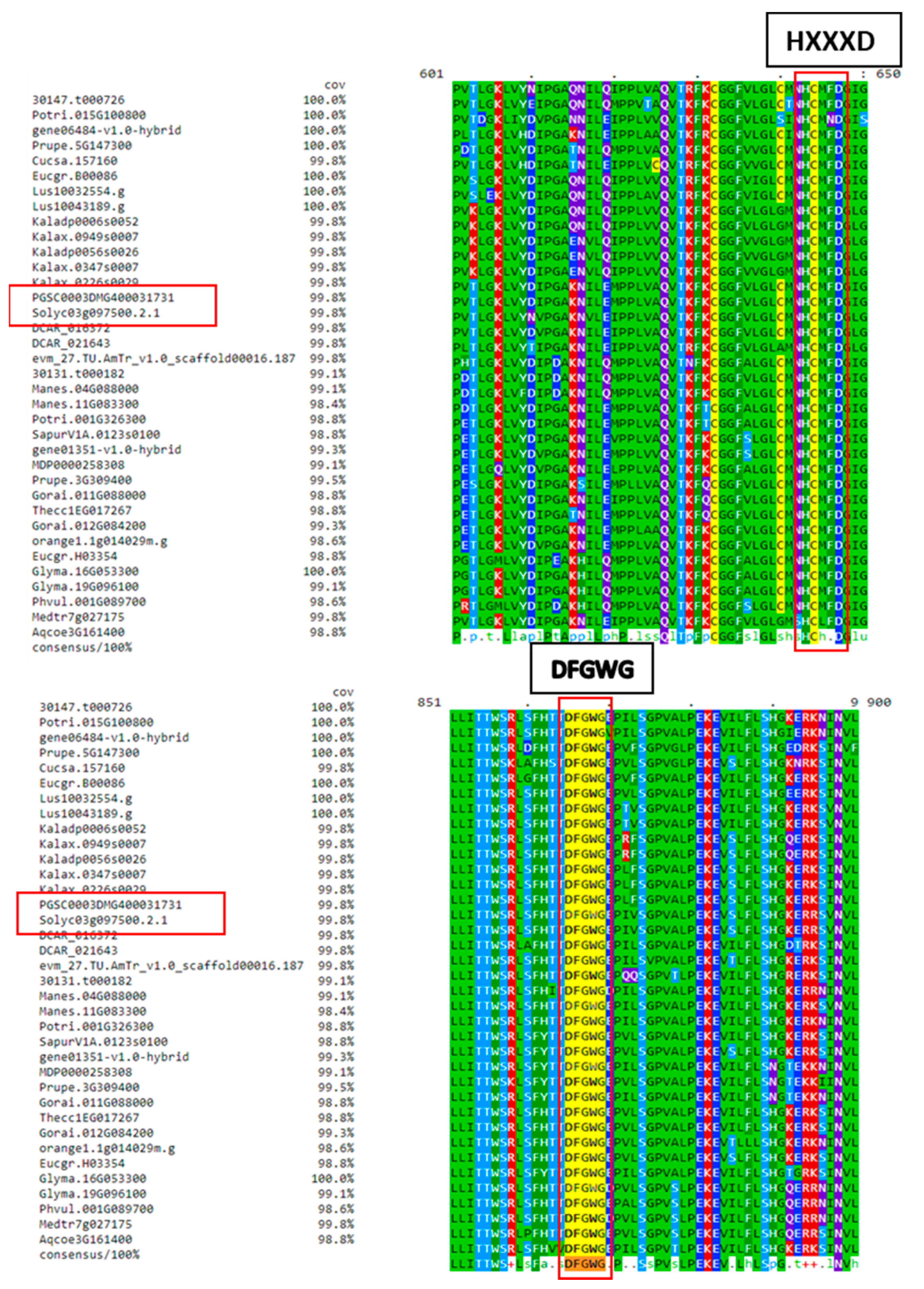
2.1. FHT Proteins of Different Plant Species Have a Conserved HxxxD Motif Involved in Catalysis and a DFGWG Motif Located at the C-Terminal End
2.2. Putative Cis Elements Found in Tomato FHT Promoter
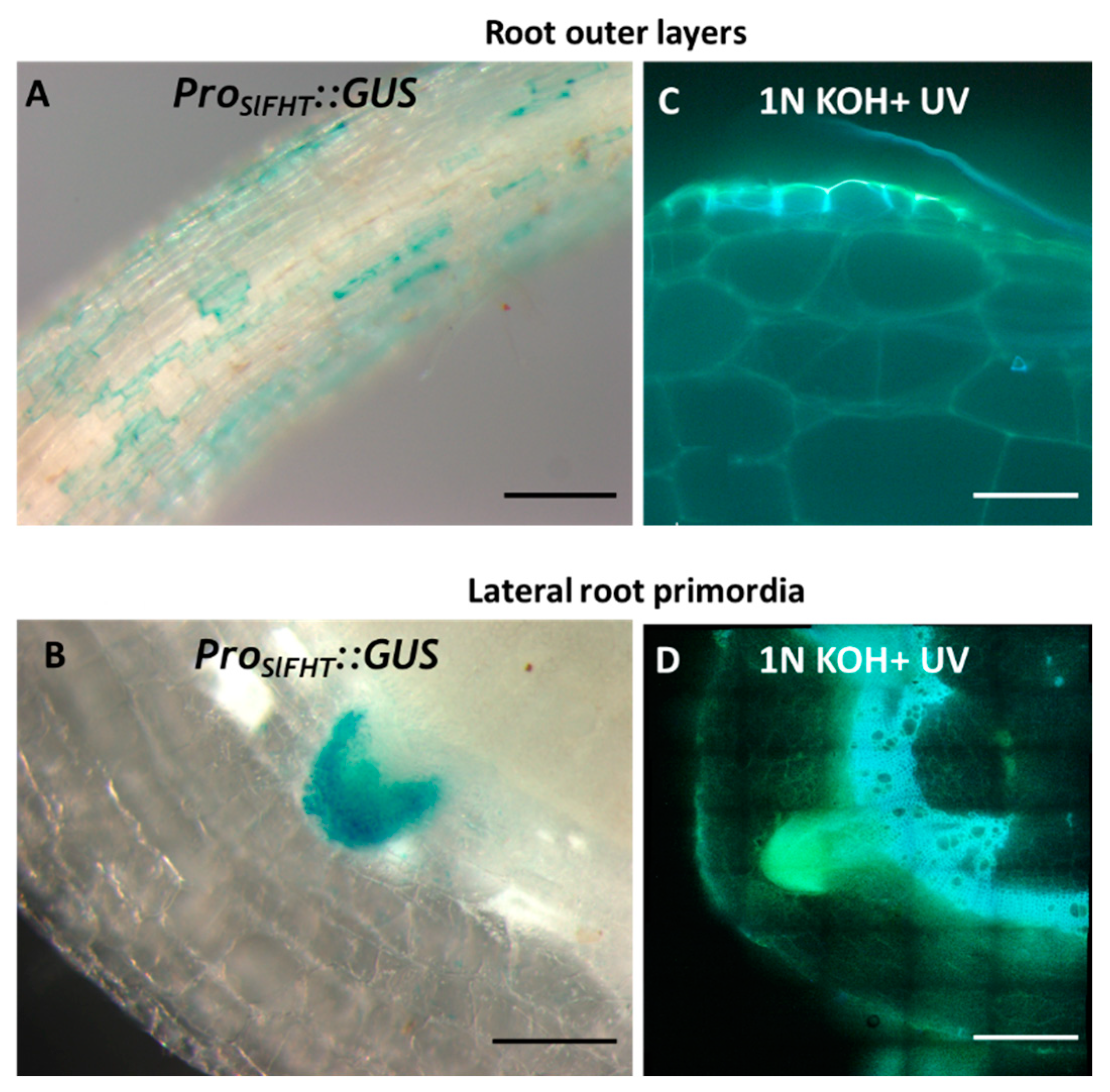
2.3. Induction of ProSlFHT:GUS in Tomato Tissues Undergoing Developmental Suberization
2.4. Induction of ProSlFHT:GUS in Tomato Tissues Undergoing Wound Healing
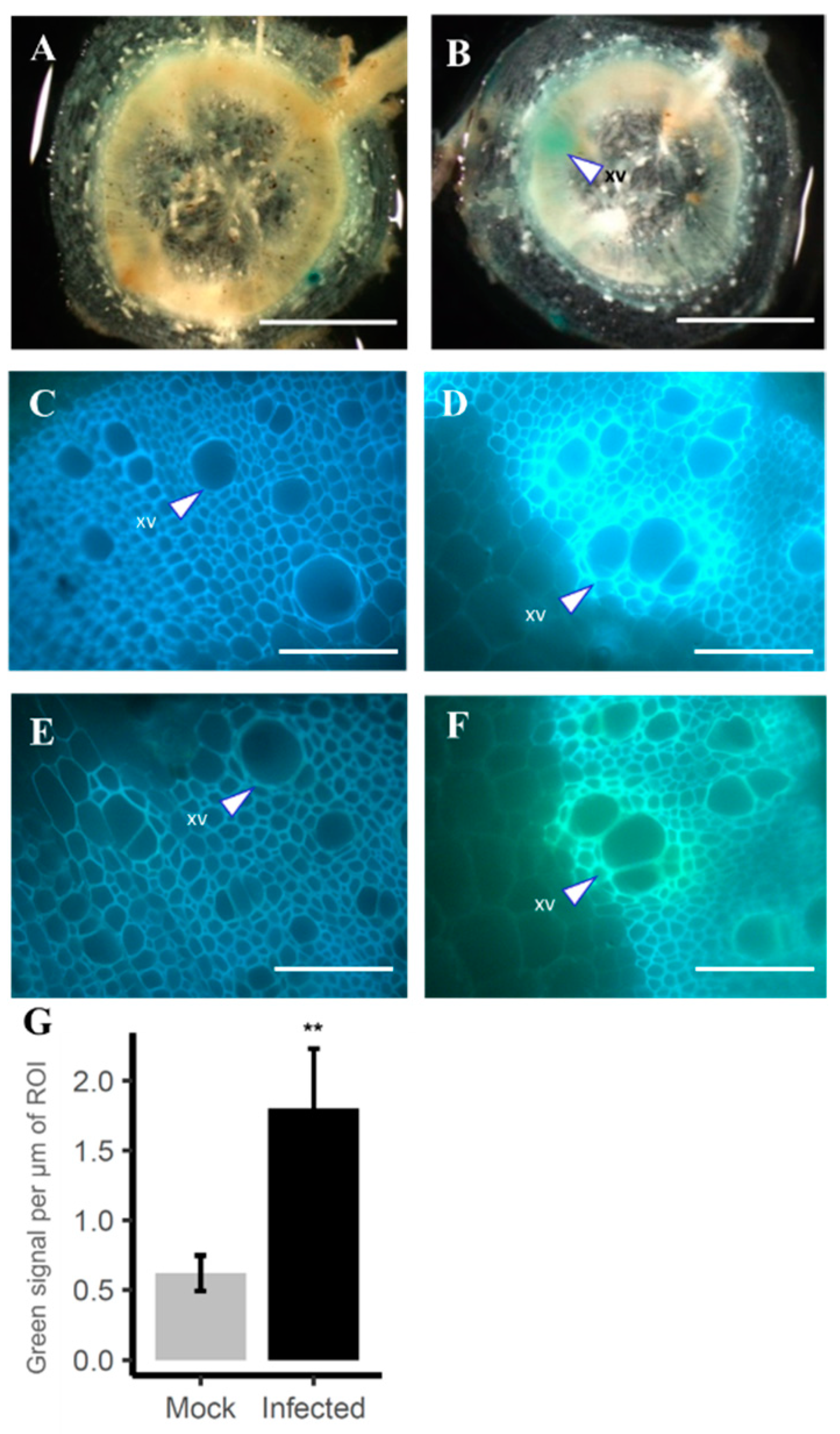
2.5. Induction of ProSlFHT::GUS in Vascular Suberization Response in Tomato against Pathogens
3. Discussion
4. Materials and Methods
4.1. Plant Material and Growth Conditions
4.2. DNA Constructs
4.3. Stable Transformation of Tomato
4.4. Detection of SlFHT Promoter Activity
4.5. Histological Methods
4.6. Wounding in Leaves
4.7. Bacterial Inoculation in Plants
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cohen, H.; Dong, Y.; Szymanski, J.; Lashbrooke, J.; Meir, S.; Almekias-Siegl, E.; Zeisler-Diehl, V.V.; Schreiber, L.; Aharoni, A. A Multilevel Study of Melon Fruit Reticulation Provides Insight into Skin Ligno-Suberization Hallmarks. Plant Physiol. 2019, 179, 1486–1501. [Google Scholar] [CrossRef] [PubMed]
- Serra, O.; Geldner, N. The Making of Suberin. New Phytol. 2022, 235, 848–866. [Google Scholar] [CrossRef] [PubMed]
- Beisson, F.; Li-Beisson, Y.; Pollard, M. Solving the Puzzles of Cutin and Suberin Polymer Biosynthesis. Curr. Opin. Plant Biol. 2012, 15, 329–337. [Google Scholar] [CrossRef] [PubMed]
- Bernards, M.A. Demystifying Suberin. Can. J. Bot. 2002, 80, 227–240. [Google Scholar] [CrossRef]
- Bernards, M.A.; Lewis, N.G. The Macromolecular Aromatic Domain in Suberized Tissue: A Changing Paradigm. Phytochemistry 1998, 47, 915–933. [Google Scholar] [CrossRef] [PubMed]
- Vishwanath, S.J.; Delude, C.; Domergue, F.; Rowland, O. Suberin: Biosynthesis, Regulation, and Polymer Assembly of a Protective Extracellular Barrier. Plant Cell Rep. 2015, 34, 573–586. [Google Scholar] [CrossRef]
- Salas-González, I.; Reyt, G.; Flis, P.; Custódio, V.; Gopaulchan, D.; Bakhoum, N.; Dew, T.P.; Suresh, K.; Franke, R.B.; Dangl, J.L.; et al. Coordination between Microbiota and Root Endodermis Supports Plant Mineral Nutrient Homeostasis. Science 2021, 371, eabd0695. [Google Scholar] [CrossRef] [PubMed]
- Chen, A.; Liu, T.; Wang, Z.; Chen, X. Plant Root Suberin: A Layer of Defence against Biotic and Abiotic Stresses. Front. Plant Sci. 2022, 13, 1056008. [Google Scholar] [CrossRef]
- Andersen, T.G.; Molina, D.; Kilian, J.; Franke, R.B.; Ragni, L.; Geldner, N. Tissue-Autonomous Phenylpropanoid Production Is Essential for Establishment of Root Barriers. Curr. Biol. 2021, 31, 965–977.e5. [Google Scholar] [CrossRef]
- Ranathunge, K.; Schreiber, L.; Franke, R. Suberin Research in the Genomics Era—New Interest for an Old Polymer. Plant Sci. 2011, 180, 399–413. [Google Scholar] [CrossRef]
- Höfer, R.; Briesen, I.; Beck, M.; Pinot, F.; Schreiber, L.; Franke, R. The Arabidopsis Cytochrome P450 CYP86A1 Encodes a Fatty Acid ω-Hydroxylase Involved in Suberin Monomer Biosynthesis. J. Exp. Bot. 2008, 59, 2347–2360. [Google Scholar] [CrossRef] [PubMed]
- Vishwanath, S.J.; Kosma, D.K.; Pulsifer, I.P.; Scandola, S.; Pascal, S.; Joubès, J.; Dittrich-Domergue, F.; Lessire, R.; Rowland, O.; Domergue, F. Suberin-Associated Fatty Alcohols in Arabidopsis: Distributions in Roots and Contributions to Seed Coat Barrier Properties. Plant Physiol. 2013, 163, 1118–1132. [Google Scholar] [CrossRef] [PubMed]
- Franke, R.; Höfer, R.; Briesen, I.; Emsermann, M.; Efremova, N.; Yephremov, A.; Schreiber, L. The DAISY Gene from Arabidopsis Encodes a Fatty Acid Elongase Condensing Enzyme Involved in the Biosynthesis of Aliphatic Suberin in Roots and the Chalaza-Micropyle Region of Seeds. Plant J. 2009, 57, 80–95. [Google Scholar] [CrossRef]
- Belsson, F.; Li, Y.; Bonaventura, G.; Pollard, M.; Ohlrogge, J.B. The Acyltransferase GPAT5 Is Required for the Synthesis of Suberin in Seed Coat and Root of Arabidopsis. Plant Cell 2007, 19, 351–368. [Google Scholar] [CrossRef] [PubMed]
- Yadav, V.; Molina, I.; Ranathunge, K.; Castillo, I.Q.; Rothstein, S.J.; Reed, J.W. ABCG Transporters Are Required for Suberin and Pollen Wall Extracellular Barriers in Arabidopsis. Plant Cell 2014, 26, 3569–3588. [Google Scholar] [CrossRef]
- Cohen, H.; Fedyuk, V.; Wang, C.; Wu, S.; Aharoni, A. SUBERMAN Regulates Developmental Suberization of the Arabidopsis Root Endodermis. Plant J. 2020, 102, 431–447. [Google Scholar] [CrossRef]
- Kosma, D.K.; Murmu, J.; Razeq, F.M.; Santos, P.; Bourgault, R.; Molina, I.; Rowland, O. AtMYB41 Activates Ectopic Suberin Synthesis and Assembly in Multiple Plant Species and Cell Types. Plant J. 2014, 80, 216–229. [Google Scholar] [CrossRef]
- Gou, M.; Hou, G.; Yang, H.; Zhang, X.; Cai, Y.; Kai, G.; Liu, C.J. The MYB107 Transcription Factor Positively Regulates Suberin Biosynthesis. Plant Physiol. 2017, 173, 1045–1058. [Google Scholar] [CrossRef]
- Gou, J.-Y.; Yu, X.-H.; Liu, C.-J. A Hydroxycinnamoyltransferase Responsible for Synthesizing Suberin Aromatics in Arabidopsis. Proc. Natl. Acad. Sci. USA 2009, 106, 18855–18860. [Google Scholar] [CrossRef]
- Molina, I.; Li-Beisson, Y.; Beisson, F.; Ohlrogge, J.B.; Pollard, M. Identification of an Arabidopsis Feruloyl-Coenzyme A Transferase Required for Suberin Synthesis. Plant Physiol. 2009, 151, 1317–1328. [Google Scholar] [CrossRef]
- Serra, O.; Hohn, C.; Franke, R.; Prat, S.; Molinas, M.; Figueras, M. A Feruloyl Transferase Involved in the Biosynthesis of Suberin and Suberin-Associated Wax Is Required for Maturation and Sealing Properties of Potato Periderm. Plant J. 2010, 62, 277–290. [Google Scholar] [CrossRef] [PubMed]
- Shukla, V.; Han, J.P.; Cléard, F.; Lefebvre-Legendre, L.; Gully, K.; Flis, P.; Berhin, A.; Andersen, T.G.; Salt, D.E.; Nawrath, C.; et al. Suberin Plasticity to Developmental and Exogenous Cues Is Regulated by a Set of MYB Transcription Factors. Proc. Natl. Acad. Sci. USA 2021, 118, e2101730118. [Google Scholar] [CrossRef] [PubMed]
- Krishnamurthy, P.; Vishal, B.; Bhal, A.; Kumar, P.P. WRKY9 Transcription Factor Regulates Cytochrome P450 Genes CYP94B3 and CYP86B1, Leading to Increased Root Suberin and Salt Tolerance in Arabidopsis. Physiol. Plant. 2021, 172, 1673–1687. [Google Scholar] [CrossRef] [PubMed]
- Mahmood, K.; Zeisler-Diehl, V.V.; Schreiber, L.; Bi, Y.M.; Rothstein, S.J.; Ranathunge, K. Overexpression of ANAC046 Promotes Suberin Biosynthesis in Roots of Arabidopsis thaliana. Int. J. Mol. Sci. 2019, 20, 6117. [Google Scholar] [CrossRef] [PubMed]
- Lashbrooke, J.; Cohen, H.; Levy-Samocha, D.; Tzfadia, O.; Panizel, I.; Zeisler, V.; Massalha, H.; Stern, A.; Trainotti, L.; Schreiber, L.; et al. MYB107 and MYB9 Homologs Regulate Suberin Deposition in Angiosperms. Plant Cell 2016, 28, 2097–2116. [Google Scholar] [CrossRef]
- Ayaz, A.; Huang, H.; Zheng, M.; Zaman, W.; Li, D.; Saqib, S.; Zhao, H.; Lü, S. Molecular Cloning and Functional Analysis of GmLACS2-3 Reveals Its Involvement in Cutin and Suberin Biosynthesis along with Abiotic Stress Tolerance. Int. J. Mol. Sci. 2021, 22, 9175. [Google Scholar] [CrossRef]
- Landgraf, R.; Smolka, U.; Altmann, S.; Eschen-Lippold, L.; Senning, M.; Sonnewald, S.; Weigel, B.; Frolova, N.; Strehmel, N.; Hause, G.; et al. The ABC Transporter ABCG1 Is Required for Suberin Formation in Potato Tuber Periderm. Plant Cell 2014, 26, 3403–3415. [Google Scholar] [CrossRef]
- Kashyap, A.; Jiménez-Jiménez, Á.L.; Zhang, W.; Capellades, M.; Srinivasan, S.; Laromaine, A.; Serra, O.; Figueras, M.; Rencoret, J.; Gutiérrez, A.; et al. Induced Ligno-Suberin Vascular Coating and Tyramine-Derived Hydroxycinnamic Acid Amides Restrict Ralstonia Solanacearum Colonization in Resistant Tomato. New Phytol. 2022, 234, 1411–1429. [Google Scholar] [CrossRef]
- Boher, P.; Serra, O.; Soler, M.; Molinas, M.; Figueras, M. The Potato Suberin Feruloyl Transferase FHT Which Accumulates in the Phellogen Is Induced by Wounding and Regulated by Abscisic and Salicylic Acids. J. Exp. Bot. 2013, 64, 3225–3236. [Google Scholar] [CrossRef]
- Bontpart, T.; Cheynier, V.; Ageorges, A.; Terrier, N. BAHD or SCPL Acyltransferase? What a Dilemma for Acylation in the World of Plant Phenolic Compounds. New Phytol. 2015, 208, 695–707. [Google Scholar] [CrossRef]
- Higo, K.; Ugawa, Y.; Iwamoto, M.; Korenaga, T. Plant Cis-Acting Regulatory DNA Elements (PLACE) Database: 1999. Nucleic Acids Res. 1999, 27, 297–300. [Google Scholar] [CrossRef] [PubMed]
- Lescot, M.; Déhais, P.; Thijs, G.; Marchal, K.; Moreau, Y.; Van De Peer, Y.; Rouzé, P.; Rombauts, S. PlantCARE, a Database of Plant Cis-Acting Regulatory Elements and a Portal to Tools for in Silico Analysis of Promoter Sequences. Nucleic Acids Res. 2002, 30, 325–327. [Google Scholar] [CrossRef] [PubMed]
- Woolfson, K.N.; Haggitt, M.L.; Zhang, Y.; Kachura, A.; Bjelica, A.; Rey Rincon, M.A.; Kaberi, K.M.; Bernards, M.A. Differential Induction of Polar and Non-Polar Metabolism during Wound-Induced Suberization in Potato (Solanum tuberosum L.) Tubers. Plant J. 2018, 93, 931–942. [Google Scholar] [CrossRef] [PubMed]
- Negrel, J.; Lotfy, S.; Javelle, F. Modulation of the Activity of Two Hydroxycinnamoyl Transferases in Wound-Healing Potato Tuber Discs in Response to Pectinase or Abscisic Acid. J. Plant Physiol. 1995, 146, 318–322. [Google Scholar] [CrossRef]
- Soliday, C.L.; Dean, B.B.; Kolattukudy, P.E. Suberization: Inhibition by Washing and Stimulation by Abscisic Acid in Potato Disks and Tissue Culture. Plant Physiol. 1978, 61, 170–174. [Google Scholar] [CrossRef]
- Kajala, K.; Gouran, M.; Shaar-Moshe, L.; Mason, G.A.; Rodriguez-Medina, J.; Kawa, D.; Pauluzzi, G.; Reynoso, M.; Canto-Pastor, A.; Manzano, C.; et al. Innovation, Conservation, and Repurposing of Gene Function in Root Cell Type Development. Cell 2021, 184, 3333–3348.e19. [Google Scholar] [CrossRef]
- Harris, P.J.; Trethewey, J.A.K. The Distribution of Ester-Linked Ferulic Acid in the Cell Walls of Angiosperms. Phytochem. Rev. 2010, 9, 19–33. [Google Scholar] [CrossRef]
- Donaldson, L.; Williams, N. Imaging and Spectroscopy of Natural Fluorophores in Pine Needles. Plants 2018, 7, 10. [Google Scholar] [CrossRef]
- Carnachan, S.M.; Harris, P.J. Ferulic Acid Is Bound to the Primary Cell Walls of All Gymnosperm Families. Biochem. Syst. Ecol. 2000, 28, 865–879. [Google Scholar] [CrossRef]
- Graça, J. Suberin: The Biopolyester at the Frontier of Plants. Front. Chem. 2015, 3, 62. [Google Scholar] [CrossRef]
- Kashyap, A.; Planas-marquès, M.; Valls, M. Blocking Intruders: Inducible Physico-Chemical Barriers against Plant Vascular Wilt Pathogens. J. Exp. Bot. 2021, 72, 184–198. [Google Scholar] [CrossRef] [PubMed]
- Brundrett, M.C.; Kendrick, B.; Peterson, C.A. Efficient Lipid Staining in Plant Material with Sudan Red 7b or Fluoral Yellow 088 in Polyethylene Glycol-Glycerol. Biotech. Histochem. 1991, 66, 111–116. [Google Scholar] [CrossRef] [PubMed]
- Naseer, S.; Lee, Y.; Lapierre, C.; Franke, R.; Nawrath, C.; Geldner, N. Casparian Strip Diffusion Barrier in Arabidopsis Is Made of a Lignin Polymer without Suberin. Proc. Natl. Acad. Sci. USA 2012, 109, 10101–10106. [Google Scholar] [CrossRef] [PubMed]
- Wahrenburg, Z.; Benesch, E.; Lowe, C.; Jimenez, J.; Vulavala, V.K.R.; Shiyou, L.; Hammerschmidt, R.; Douches, D.; Yim, W.C.; Santos, P.; et al. Transcriptional Regulation of Wound Suberin Deposition in Potato Cultivars with Differential Wound Healing Capacity. Plant J. 2021, 107, 77–99. [Google Scholar] [CrossRef] [PubMed]
- Lulai, E.C.; Corsini, D.L. Differential Deposition of Suberin Phenolic and Aliphatic Domains and Their Roles in Resistance to Infection during Potato Tuber (Solanum tuberosum L.) Wound-Healing. Physiol. Mol. Plant Pathol. 1998, 53, 209–222. [Google Scholar] [CrossRef]
- Bernards, M.A.; Lewis, N.G. Alkyl Ferulates in Wound Healing Potato Tubers. Phytochemistry 1992, 31, 3409–3412. [Google Scholar] [CrossRef] [PubMed]
- Berhin, A.; de Bellis, D.; Franke, R.B.; Buono, R.A.; Nowack, M.K.; Nawrath, C. The Root Cap Cuticle: A Cell Wall Structure for Seedling Establishment and Lateral Root Formation. Cell 2019, 176, 1367–1378.e8. [Google Scholar] [CrossRef]
- Philippe, G.; Sørensen, I.; Jiao, C.; Sun, X.; Fei, Z.; Domozych, D.S.; Rose, J.K. Cutin and Suberin: Assembly and Origins of Specialized Lipidic Cell Wall Scaffolds. Curr. Opin. Plant Biol. 2020, 55, 11–20. [Google Scholar] [CrossRef]
- Mazier, M.; Flamain, F.; Nicolaï, M.; Sarnette, V.; Caranta, C. Knock-down of Both EIF4E1 and EIF4E2 Genes Confers Broad-Spectrum Resistance against Potyviruses in Tomato. PLoS ONE 2011, 6, e29595. [Google Scholar] [CrossRef]
- Planas-Marquès, M.; Kressin, J.P.; Kashyap, A.; Panthee, D.R.; Louws, F.J.; Coll, N.S.; Valls, M. Four Bottlenecks Restrict Colonization and Invasion by the Pathogen Ralstonia Solanacearum in Resistant Tomato. J. Exp. Bot. 2019, 71, 2157–2171. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Motif Name | Sequence | Motif | Function |
|---|---|---|---|---|
| Nº | ||||
| Stress | ||||
| WBOXNTCHN48 | CTGACY | 1 | “W box.” Elicitor-responsive element | |
| WBOXNTERF3 | TGACY | 5 | “W box.” Response to wounding | |
| OSE1ROOTNODULE | AAAGAT | 5 | Organ-specific elements (OSE) in infected cells of root nodules | |
| OSE2ROOTNODULE | CTCTT | 2 | Organ-specific elements (OSE) in infected cells of root nodules | |
| PREATPRODH | ACTCAT | 1 | Pro- or hypo-osmolarity-responsive element | |
| RAV1AAT | CAACA | 5 | RAV1 binding site, cold responsiveness | |
| MYB1AT | WAACCA | 3 | Element involved in dehydration responsiveness | |
| MYBCORE | CNGTTR | 2 | Element involved in response to water stress | |
| GT1GMSCAM4 | GAAAAA | 9 | Pathogenesis and salt-induced element | |
| CURECORECR | GTAC | 10 | Copper-response element | |
| CCAATBOX1 | CCAAT | 5 | HSE (heat shock element) | |
| ANAERO1CONSENSUS | AAACAAA | 2 | Response to anaerobiosis | |
| ACGTATERD1 | ACGT | 4 | Involved in etiolation-induced dehydration | |
| Hormones | ||||
| ABA | WBBOXPCWRKY1 | TTTGACY | 1 | WRKY proteins binding site, responsive to ABA |
| MYB2CONSENSUSAT | YAACKG | 1 | MYB recognition site involved in dehydration and ABA response | |
| MYCATERD1 | CATGTG | 1 | MYC binding site involved in response to dehydration and ABA | |
| MYCATRD22 | CACATG | 1 | MYC binding site involved in response to dehydration and ABA | |
| MYCCONSENSUSAT | CANNTG | 6 | MYC binding site involved in response to dehydration and ABA | |
| SORLIP1AT | GCCAC | 4 | Light-inducible, root-specific, and ABA-responsive element | |
| Salicylic acid | WBOXATNPR1 | TTGAC | 3 | “W-box”; binding site for SA-induced WRKY transcription factor |
| Ethylene | ERELEE4 CGTCA-motif | AWTTCAAA | 1 | Ethylene-responsive element |
| Auxin | CATATGGMSAUR | CATATG | 2 | Element involved in auxin responsiveness |
| Gibberellin | GARE2OSREP1 | TAACGTA | 1 | Gibberellin-responsive element (GARE) |
| GAREAT | TAACAAR | 2 | GARE (GA-responsive element) | |
| MYBGAHV | TAACAAA | 2 | Central element of gibberellin (GA) response complex (GARC) | |
| PYRIMIDINEBOXOSRAMY1A | CCTTTT | 2 | Gibberellin-response cis element of GARE and pyrimidine | |
| TATCCACHVAL21 | TATCCAC | 1 | Conserved cis-acting response complex (GARC) | |
| PYRIMIDINEBOXHVEPB1 | TTTTTTCC | 2 | “Pyrimidine box”. Required for GA induction | |
| Organ-/Tissue-/Cell type-specific | ||||
| ROOTMOTIFTAPOX1 | ATATT | 12 | Root specific element from rolD gene | |
| SP8BFIBSP8BIB | TACTATT | 3 | Tuberous root specific element | |
| RHERPATEXPA7 | KCACGW | 1 | Right part of RHEs (root-hair-specific cis-elements) | |
| EBOXBNNAPA | CANNTG | 6 | RRE element. Tissue-specific activation of phenylpropanoids biosynthesis genes | |
| RYREPEATBNNAPA | CATGCA | 1 | Required for seed-specific expression | |
| POLLEN1LELAT52 | AGAAA | 12 | Element required for pollen-specific expression | |
| CACTFTPPCA1 | YACT | 28 | Element related to mesophyll expression | |
| BOXIINTPATPB | ATAGAA | 3 | Conserved element in promoters of plastids genes. | |
| TAAAGSTKST1 | TAAAG | 12 | Dof1 protein controlling guard cell-specific gene expression | |
| Others | ||||
| MYBPZM | CCWACC | 3 | Core of consensus myb homolog binding site | |
| MYBST1 | GGATA | 2 | Core motif of MybSt1 (a potato MYB homolog) binding site | |
| RYREPEATLEGUMINBOX | CATGCAY | 1 | RY repeat found in seed-storage protein genes |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kashyap, A.; Jiménez-Jiménez, Á.; Figueras, M.; Serra, O.; Valls, M.; Coll, N.S. The Tomato Feruloyl Transferase FHT Promoter Is an Accurate Identifier of Early Development and Stress-Induced Suberization. Plants 2023, 12, 1890. https://doi.org/10.3390/plants12091890
Kashyap A, Jiménez-Jiménez Á, Figueras M, Serra O, Valls M, Coll NS. The Tomato Feruloyl Transferase FHT Promoter Is an Accurate Identifier of Early Development and Stress-Induced Suberization. Plants. 2023; 12(9):1890. https://doi.org/10.3390/plants12091890
Chicago/Turabian StyleKashyap, Anurag, Álvaro Jiménez-Jiménez, Mercè Figueras, Olga Serra, Marc Valls, and Nuria S. Coll. 2023. "The Tomato Feruloyl Transferase FHT Promoter Is an Accurate Identifier of Early Development and Stress-Induced Suberization" Plants 12, no. 9: 1890. https://doi.org/10.3390/plants12091890