
Optimization of an Efficient Protoplast Transformation System for Transient Expression Analysis Using Leaves of Torenia fournieri



{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
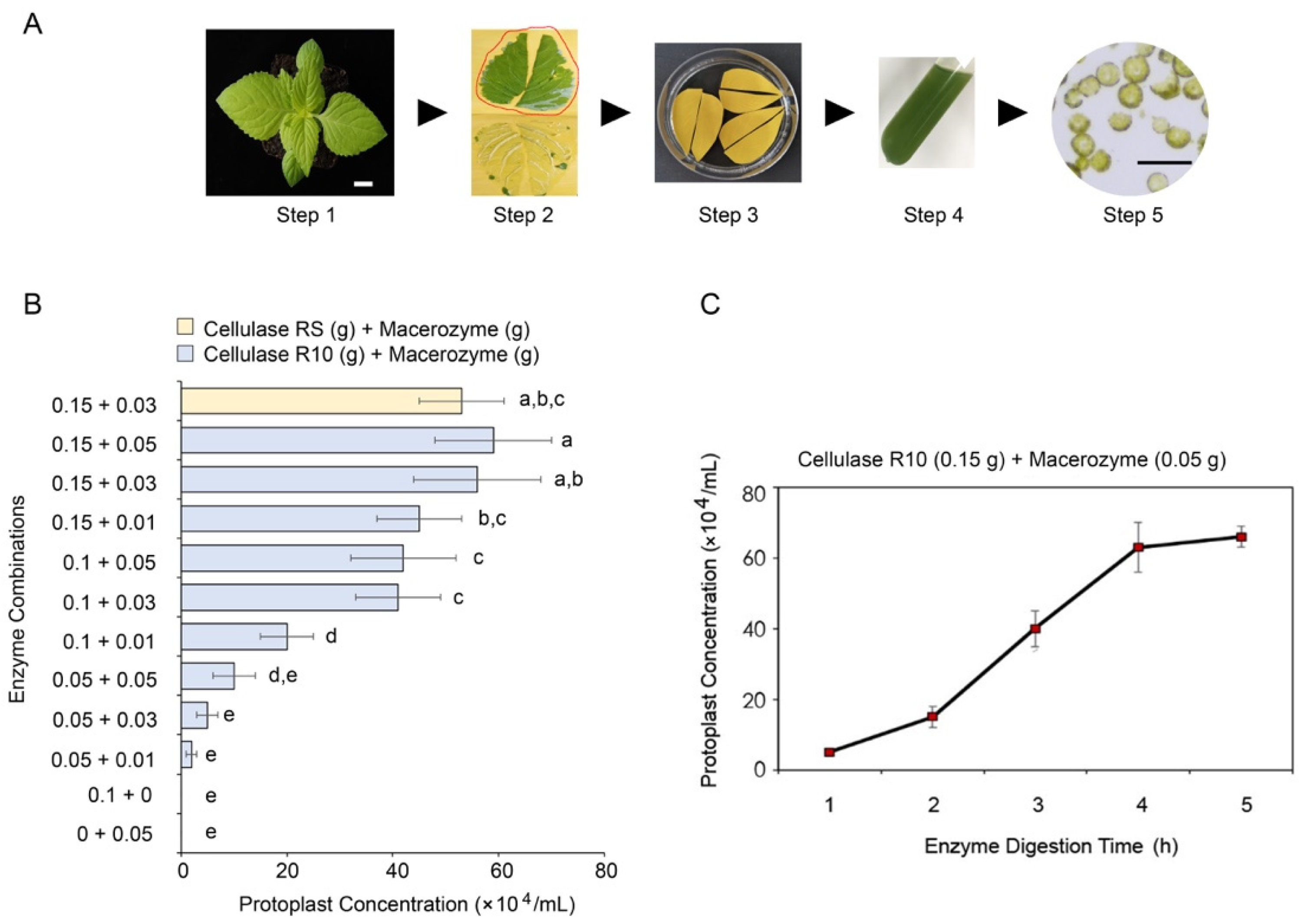
2.1. Isolation of Protoplasts from T. fournieri Leaves
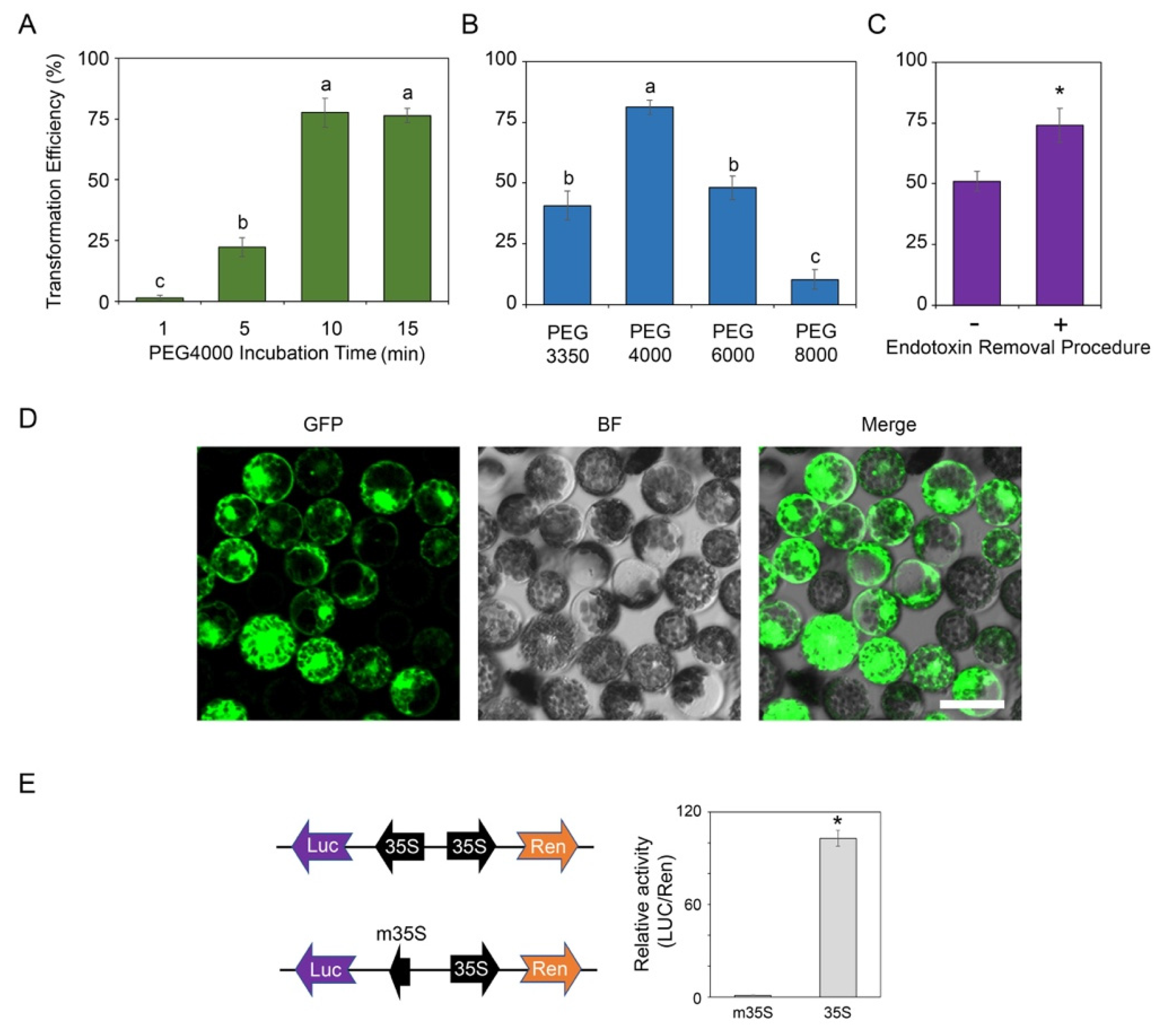
2.2. Protoplast Transformation
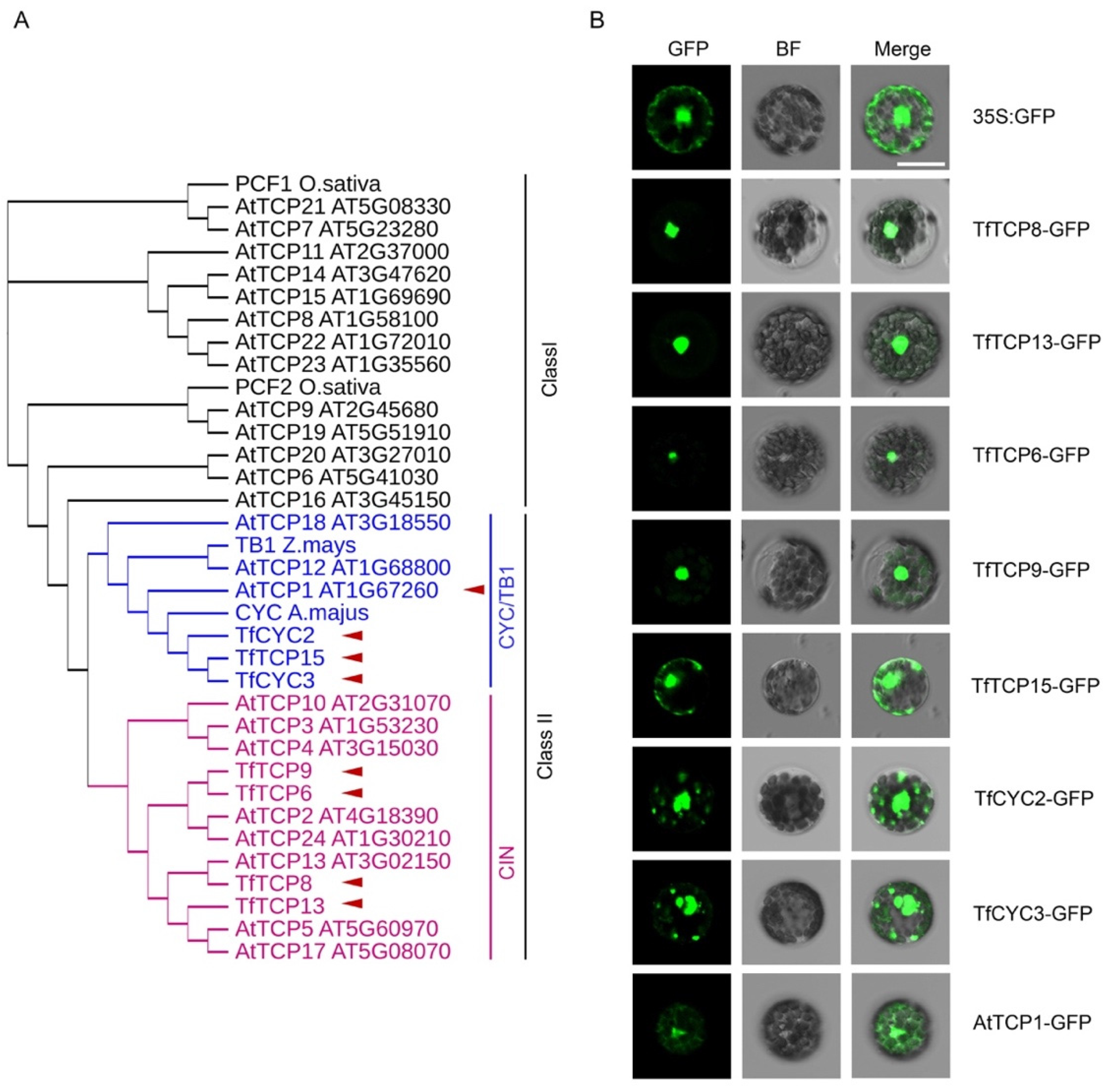
2.3. Subcellular Localization Analysis of the TCP TFs Using the Optimized T. fournieri Protoplast Transformation Protocol
3. Discussion
4. Materials and Methods
4.1. Plant Materials
4.2. Protoplast Isolation and Transformation
4.3. Luciferase Activity Measurement
4.4. Sub-Cellular Localization
4.5. Phylogenetic Tree Construction
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Aida, R. Torenia fournieri (torenia) as a model plant for transgenic studies. Plant Biotechnol-Nar. 2008, 25, 541–545. [Google Scholar] [CrossRef]
- Nishihara, M.; Shimoda, T.; Nakatsuka, T.; Arimura, G. Frontiers of torenia research: Innovative ornamental traits and study of ecological interaction networks through genetic engineering. Plant Methods 2013, 9, 23. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Zhang, D.; Gao, Q.; Luo, Y.; Zhang, H.; Ma, B.; Chen, C.; Whibley, A.; Zhang, Y.; Cao, Y.; et al. Genome structure and evolution of Antirrhinum majus L. Nat. Plants 2019, 5, 174–183. [Google Scholar] [CrossRef] [PubMed]
- Kikuchi, S.; Tanaka, H.; Shiba, T.; Mii, M.; Tsujimoto, H. Genome size, karyotype, meiosis and a novel extra chromosome in Torenia fournieri, T-baillonii and their hybrid. Chromosome Res. 2006, 14, 665–672. [Google Scholar] [CrossRef] [PubMed]
- Aida, R.; Shibata, M. Agrobacterium-Mediated Transformation of Torenia (Torenia-Fournieri). Breed. Sci. 1995, 45, 71–74. [Google Scholar] [CrossRef]
- Aida, R. A protocol for transformation of Torenia. Methods Mol. Biol. 2012, 847, 267–274. [Google Scholar] [PubMed]
- Nishihara, M.; Higuchi, A.; Watanabe, A.; Tasaki, K. Application of the CRISPR/Cas9 system for modification of flower color in Torenia fournieri. BMC Plant Biol. 2018, 18, 331. [Google Scholar] [CrossRef]
- Xu, Y.; Li, R.; Luo, H.; Wang, Z.; Li, M.W.; Lam, H.M.; Huang, C. Protoplasts: Small cells with big roles in plant biology. Trends Plant Sci. 2022, 8, 828–829. [Google Scholar] [CrossRef]
- Yoo, S.D.; Cho, Y.H.; Sheen, J. Arabidopsis mesophyll protoplasts: A versatile cell system for transient gene expression analysis. Nat. Protoc. 2007, 2, 1565–1572. [Google Scholar] [CrossRef]
- Ma, R.; Zhao, J.; Du, H.C.; Tian, S.; Li, L.W. Removing endotoxin from plasmid samples by Triton X-114 isothermal extraction. Anal. Biochem. 2012, 424, 124–126. [Google Scholar] [CrossRef]
- Xiong, L.; Li, C.; Li, H.; Lyu, X.; Zhao, T.; Liu, J.; Zuo, Z.; Liu, B. A transient expression system in soybean mesophyll protoplasts reveals the formation of cytoplasmic GmCRY1 photobody-like structures. Sci. China Life Sci. 2019, 62, 1070–1077. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.H.; Jin, S.; Kim, S.Y.; Kim, W.; Ahn, J.H. A fast, efficient chromatin immunoprecipitation method for studying protein-DNA binding in Arabidopsis mesophyll protoplasts. Plant Methods 2017, 13, 42. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.W.; Fu, J.X.; Li, J.; Cheng, X.L.; Li, F.; Dong, J.F.; Liu, Z.L.; Zhuang, C.X. A Novel Co-immunoprecipitation Protocol Based on Protoplast Transient Gene Expression for Studying Protein–protein Interactions in Rice. Plant Mol. Biol. Rep. 2014, 32, 153–161. [Google Scholar] [CrossRef]
- Hellens, R.P.; Allan, A.C.; Friel, E.N.; Bolitho, K.; Grafton, K.; Templeton, M.D.; Karunairetnam, S.; Gleave, A.P.; Laing, W.A. Transient expression vectors for functional genomics, quantification of promoter activity and RNA silencing in plants. Plant Methods 2005, 1, 13. [Google Scholar] [CrossRef]
- Sparkes, I.A.; Runions, J.; Kearns, A.; Hawes, C. Rapid, transient expression of fluorescent fusion proteins in tobacco plants and generation of stably transformed plants. Nat. Protoc. 2006, 1, 2019–2025. [Google Scholar] [CrossRef]
- González-Grandío, E.; Cubas, P. Chapter 9-TCP Transcription Factors: Evolution, Structure, and Biochemical Function. In Plant Transcription Factors; Gonzalez, D.H., Ed.; Academic Press: Boston, MA, USA, 2016; pp. 139–151. [Google Scholar]
- Zhang, L.; Zhou, L.; Yung, W.S.; Su, W.; Huang, M. Ectopic expression of Torenia fournieri TCP8 and TCP13 alters the leaf and petal phenotypes in Arabidopsis thaliana. Physiol. Plant 2021, 173, 856–866. [Google Scholar] [CrossRef]
- Aguilar-Martinez, J.A.; Sinha, N. Analysis of the role of Arabidopsis class I TCP genes AtTCP7, AtTCP8, AtTCP22, and AtTCP23 in leaf development. Front. Plant Sci. 2013, 4, 406. [Google Scholar] [CrossRef]
- Mazur, M.J.; Spears, B.J.; Djajasaputra, A.; van der Gragt, M.; Vlachakis, G.; Beerens, B.; Gassmann, W.; van den Burg, H.A. Arabidopsis TCP Transcription Factors Interact with the SUMO Conjugating Machinery in Nuclear Foci. Front. Plant Sci. 2017, 8, 2043. [Google Scholar] [CrossRef]
- Urano, K.; Maruyama, K.; Koyama, T.; Gonzalez, N.; Inze, D.; Yamaguchi-Shinozaki, K.; Shinozaki, K. CIN-like TCP13 is essential for plant growth regulation under dehydration stress. Plant Mol. Biol. 2022, 108, 257–275. [Google Scholar] [CrossRef]
- Wei, W.; Hu, Y.; Cui, M.Y.; Han, Y.T.; Gao, K.; Feng, J.Y. Identification and Transcript Analysis of the TCP Transcription Factors in the Diploid Woodland Strawberry Fragaria vesca. Front Plant Sci. 2016, 7, 1937. [Google Scholar] [CrossRef]
- Leng, X.; Wei, H.; Xu, X.; Ghuge, S.A.; Jia, D.; Liu, G.; Wang, Y.; Yuan, Y. Genome-wide identification and transcript analysis of TCP transcription factors in grapevine. BMC Genom. 2019, 20, 786. [Google Scholar] [CrossRef] [PubMed]
- Okuda, S.; Suzuki, T.; Kanaoka, M.M.; Mori, H.; Sasaki, N.; Higashiyama, T. Acquisition of LURE-binding activity at the pollen tube tip of Torenia fournieri. Mol. Plant 2013, 6, 1074–1090. [Google Scholar] [CrossRef] [PubMed]
- Su, S.; Xiao, W.; Guo, W.; Yao, X.; Xiao, J.; Ye, Z.; Wang, N.; Jiao, K.; Lei, M.; Peng, Q.; et al. The CYCLOIDEA-RADIALIS module regulates petal shape and pigmentation, leading to bilateral corolla symmetry in Torenia fournieri (Linderniaceae). New Phytol. 2017, 215, 1582–1593. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Su, J.; Duan, S.; Ao, Y.; Dai, J.; Liu, J.; Wang, P.; Li, Y.; Liu, B.; Feng, D.; et al. A highly efficient rice green tissue protoplast system for transient gene expression and studying light/chloroplast-related processes. Plant Methods 2011, 7, 30. [Google Scholar] [CrossRef] [PubMed]
- Gao, L.; Shen, G.; Zhang, L.; Qi, J.; Zhang, C.; Ma, C.; Li, J.; Wang, L.; Malook, S.U.; Wu, J. An efficient system composed of maize protoplast transfection and HPLC-MS for studying the biosynthesis and regulation of maize benzoxazinoids. Plant Methods 2019, 15, 144. [Google Scholar] [CrossRef]
- Wu, F.; Hanzawa, Y. A Simple Method for Isolation of Soybean Protoplasts and Application to Transient Gene Expression Analyses. J. Vis. Exp. 2018, 131, e57258. [Google Scholar] [CrossRef]
- Fischer, R.; Hain, R. Tobacco protoplast transformation and use for functional analysis of newly isolated genes and gene constructs. Methods Cell Biol. 1995, 50, 401–410. [Google Scholar]
- Sivanandhan, G.; Bae, S.; Sung, C.; Choi, S.R.; Lee, G.J.; Lim, Y.P. Optimization of Protoplast Isolation from Leaf Mesophylls of Chinese Cabbage (Brassica rapa ssp. pekinensis) and Subsequent Transfection with a Binary Vector. Plants 2021, 10, 2636. [Google Scholar] [CrossRef]
- Gou, Y.-J.; Li, Y.-L.; Bi, P.-P.; Wang, D.-J.; Ma, Y.-Y.; Hu, Y.; Zhou, H.-C.; Wen, Y.-Q.; Feng, J.-Y. Optimization of the protoplast transient expression system for gene functional studies in strawberry (Fragaria vesca). Plant Cell Tissue Organ Cult. (PCTOC) 2020, 141, 41–53. [Google Scholar] [CrossRef]
- Wang, J.; Wang, Y.; Lu, T.; Yang, X.; Liu, J.; Dong, Y.; Wang, Y. An Efficient and Universal Protoplast Isolation Protocol Suitable for Transient Gene Expression Analysis and Single-Cell RNA Sequencing. Int. J. Mol. Sci. 2022, 23, 3419. [Google Scholar] [CrossRef]
- Huang, M.; Zhang, L.; Zhou, L.; Wang, M.; Yung, W.-S.; Wang, Z.; Duan, S.; Xiao, Z.; Wang, Q.; Wang, X.; et al. An expedient survey and characterization of the soybean JAGGED 1 (GmJAG1) transcription factor binding preference in the soybean genome by modified ChIPmentation on soybean protoplasts. Genomics 2021, 113, 344–355. [Google Scholar] [CrossRef] [PubMed]
- Schweiger, R.; Schwenkert, S. Protein-protein interactions visualized by bimolecular fluorescence complementation in tobacco protoplasts and leaves. J. Vis. Exp. JoVE 2014, 85, 51327. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Yu, H.; Lin, S.; Gao, Y. Molecular Characterization of FT and FD Homologs from Eriobotrya deflexa Nakai forma koshunensis. Front. Plant Sci. 2016, 7, 8. [Google Scholar] [CrossRef] [PubMed]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, L.; Yung, W.-S.; Wang, Z.; Li, M.-W.; Huang, M. Optimization of an Efficient Protoplast Transformation System for Transient Expression Analysis Using Leaves of Torenia fournieri. Plants 2022, 11, 2106. https://doi.org/10.3390/plants11162106
Zhang L, Yung W-S, Wang Z, Li M-W, Huang M. Optimization of an Efficient Protoplast Transformation System for Transient Expression Analysis Using Leaves of Torenia fournieri. Plants. 2022; 11(16):2106. https://doi.org/10.3390/plants11162106
Chicago/Turabian StyleZhang, Ling, Wai-Shing Yung, Zhili Wang, Man-Wah Li, and Mingkun Huang. 2022. "Optimization of an Efficient Protoplast Transformation System for Transient Expression Analysis Using Leaves of Torenia fournieri" Plants 11, no. 16: 2106. https://doi.org/10.3390/plants11162106