
Hairy Root Cultures as a Source of Polyphenolic Antioxidants: Flavonoids, Stilbenoids and Hydrolyzable Tannins


1
Maj Institute of Pharmacology, Polish Academy of Sciences, Smętna Street 12, 31-343 Kraków, Poland
2
National Technical University, Kharkiv Polytechnic Institute, Kyrpychova Street 2, 61002 Kharkiv, Ukraine
*
Author to whom correspondence should be addressed.
Plants 2022, 11(15), 1950; https://doi.org/10.3390/plants11151950
Submission received: 20 June 2022
/
Revised: 20 July 2022
/
Accepted: 25 July 2022
/
Published: 27 July 2022
(This article belongs to the Special Issue Antioxidant Activity of Medicinal and Aromatic Plants)
Abstract
:Due to their chemical properties and biological activity, antioxidants of plant origin have gained interest as valuable components of the human diet, potential food preservatives and additives, ingredients of cosmetics and factors implicated in tolerance mechanisms against environmental stress. Plant polyphenols are the most prominent and extensively studied, albeit not only group of, secondary plant (specialized) metabolites manifesting antioxidative activity. Because of their potential economic importance, the productive and renewable sources of the compounds are desirable. Over thirty years of research on hairy root cultures, as both producers of secondary plant metabolites and experimental systems to investigate plant biosynthetic pathways, brought about several spectacular achievements. The present review focuses on the Rhizobium rhizogenes-transformed roots that either may be efficient sources of plant-derived antioxidants or were used to elucidate some regulatory mechanisms responsible for the enhanced accumulation of antioxidants in plant tissues.
1. Introduction
The interest in hairy root (HR) cultures as fast-growing, genetically stable axenic roots that are effective producers of both biomass and specialized plant metabolites started in the 1980s [1]. Since then, numerous culture systems have been established to produce compounds of economic value or potential economic interest to produce recombinant proteins to obtain transgenic plants of altered phenotype to obtain artificial seeds and perform metabolic engineering of plants [1,2,3,4,5,6]. The ability of Rhizobium rhizogenes (also known as Agrobacterium rhizogenes) [7] to transfer and permanently incorporate a foreign genetic material into the plant genomic DNA has also been used to study the regulatory mechanisms of plant metabolism [6,8,9]. Additionally, the enzymatic activity of HRs may be utilized for biotransformation and phytoremediation processes [10,11,12,13,14].
In contrast to other fast-growing types of plant tissue cultures, HRs are usually cultivated in nutrient media with a reduced concentration of macronutrients and without the addition of growth regulators that lower the costs of biotechnological processes. Several strategies were employed to scale up HR cultures from the laboratory to industrial applications, including various bioreactor designs [15,16], e.g., airlifts, bubble columns and nutrient mist, etc. The production of biologically active natural products and proteins by HR cultures has been a subject of numerous patents and shows potential for successful industrial applications.
Until now, the main limitations that hamper the industrial use of HR cultures are still unsatisfactory upscaling results (the need for large-volume bioreactors) and unsatisfactory cost/profit ratios.
Nearly every class of plant-specialized metabolite includes some compounds of antioxidative activity [17,18,19,20], but the most effective antioxidants, acting by direct free-radical scavenging, originate mostly from phenolics and terpenoids [21,22]. Numerous natural products known as antioxidants are also recognized as potent chemopreventive and anticancer agents, influencing various intracellular signaling pathways [23,24]. Antioxidants of plant origin may also be useful as protective factors against dysfunctions of nervous system connected with aging [25].
Plant metabolites with antioxidant activity, which are the constituents of widely grown food plants, can be isolated from post-production waste or obtained as side products in the process of food manufacturing [26,27,28,29]. Some natural antioxidant products are a common occurrence and their isolation from either cultivated or wild plants seem to be the optimal solution. The yields of these natural products may be improved by bioengineering using the transformation by R. rhizogenes. In the case of threatened or overexploited plant species, HR cultures offer an opportunity to produce rare and valuable compounds with no harm to the natural habitats of the plant.
The present review is based on the results published in journals indexed in the Web of Science and Scopus databases. Due to the large number of studies dealing with polyphenolic metabolites of HRs, we decided to limit the scope of the review to flavonoids, stilbenoids and hydrolyzable tannins.
2. Polyphenolic Antioxidants in HRs
Phenolics are one of the largest classes of plant-specialized metabolites. They all originate from the shikimate pathway leading to aromatic amino acids phenylalanine and tyrosine and also share the phenylpropanoid pathway (Figure 1). The compounds are divided into several subclasses, including flavonoids, tannins, stilbenoids, lignans, hydroxycinnamates and phenylethanoids, to mention a few of the most popular subclasses.
2.1. Flavonoids
2.1.1. Anthocyanins and Proanthocyanidins (PAs)
Numerous factors may influence anthocyanin and PA production by HRs. The most frequently studied were the effects of light, plant growth regulators, sucrose concentration and the R. rhizogenes strain used in the experiment. The overexpression or heterologous expression of genes-encoding transcription factors implicated in the regulation of flavonoid biosynthesis (especially members of myb gene family) or the overexpression of genes-encoding biosynthetic enzymes (DFR, LAR, ANR) are strategies often used to increase anthocyanins and PAs content in the HRs or transgenic plants regenerated from the roots.
Morris and Robbins [30] observed that the HRs of Lotus corniculatus L. (Fabaceae), maintained in the dark, accumulated insoluble tannins, which presented cyanin and delphinidin at a ratio like that found in the intact plant after hydrolysis. Supplementation with auxins reduced the content of tannins in the roots. On the contrary, in the HR cultures of Leontopodium alpinum Cass. (Asteraceae), the addition of 6-benzylaminopurine (BAP) to the culture medium 14 days prior to harvest increased the anthocyanins content [31]. An effect of auxin supplementation on anthocyanin contents in the HRs of Tartary buckwheat (Fagopyrum tataricum (L.) Gaertn., Polygonaceae) cultivar “Hokkai T10” was also studied [32]. Indole-3-butyric acid (IBA) added to a nutrient medium at a concentration of 4.92 µM, caused enhanced accumulation of cyanidin 3-O-glucoside and cyanidin 3-O-rutinoside in the roots (for structures, see Figure 2). The anthocyanin content reached 0.89 and 1.15 mg/g dry weight (DW), respectively, and was about three times higher than that found in the control cultures.
Nishiyama and Yamakawa investigated effects of the culture illumination, composition of nutrient medium and sucrose content on the anthocyanin production in the HRs of Ipomea batatas (L.) Lam. (Convolvulaceae) [33]. They found that light and high sucrose concentration in the medium (5%) favored anthocyanin production. The beneficial effect of light on cyanidin 3-O-rutinoside accumulation was also observed in Tartary buckwheat HRs [34]. Motomori et al. [35] studied polyphenolics accumulation in the HRs of Fragaria x ananassa Duch. cv. Reikou (Rosaceae). In optimum conditions, the root culture produced up to 8 mg/g DW of procyanidin B-3, but the roots of the field-grown plant contained three times as much of the compound (24.2 mg/g DW).
The expression of full-length cDNA encoding dihydroflavonol 4-reductase (DFR) from Antirrhinum majus L. (common snapdragon, Plantaginaceae) in the HRs of Lotus corniculatus L. led to several high-producing clones. The maximum content of condensed tannins in the control roots was estimated as 0.62 mg/g fresh weight (FW), whereas the transgenic roots produced up to 1.06 mg/g FW of the compounds [36]. The proportions of subunits (procyanidin, prodelphinidin and propelargonidin) yielded on the hydrolysis of condensed tannins were also measured, and significant differences in propelargonidin accumulation were observed in the transgenic clones in comparison with the controls. The plasmid 121.Sn carrying the maize gene Sn, which is responsible for the transactivation of the anthocyanin pathway in different tissues, was introduced into the genomes of ten different plant species using the transformation with R. rhizogenes. Over 50% of the obtained Medicago sativa L. (Fabaceae) and Lotus angustissimus L. accession CPI 113587 HR clones showed pigmentation. On the other hand, the HRs of Lotus corniculatus, L. japonicus (Regel) K. Larsen and L. angustissimus accession CPI 113590 produced no pigment. The results suggested the transcription factor Sn derived from monocotyledonous plant may function in several dicot species [37].
The HRs of Medicago truncatula Gaertn. (Fabaceae) and Vitis vinifera L. (Vitaceae) were used to investigate the mechanisms of anthocyanin biosynthesis regulation by transcription factors encoded by the members of myb gene family [38,39,40,41,42,43,44,45]. The expression of Arabidopsis TT2 (Transparent Testa 2) transcription factor from the MYB family in the HRs of M. truncatula led to the induction of the genes for flavonoid and proanthocyanidin (PA) biosynthesis pathway and enhanced accumulation of PAs in the roots. The average soluble PAs content in TT2-carrying transformants was over ten times higher than that in the control HRs transfected with an “empty”vector. Insoluble PAs content in TT2-expressing roots was over 24-fold higher [38]. The induction of genes for PAs biosynthetic pathway was also true when the transcription factors: GmTT2A, GmTT2B or GmMYB5A were ectopically expressed in soybean (Glycine max L.) HRs [46]. Expression of AtTT2 in chickpea HRs resulted in a high accumulation of soluble PAs (>1200 μg catechin equivalents (eq.)/g FW) [47]. MtPAR MYB transcription factor expressed in M. truncatula HRs dramatically increased soluble PAs content (up to 100-fold higher than that in the control roots) but did not influence insoluble PAs accumulation and diminished the anthocyanin content. Maximum content of soluble PAs reached 10 μM of (−) epicatechin eq. per g FW [40]. Li et al. [48] found that transgenic M. truncatula HRs that over-expressed MtPAR were characterized by enhanced accumulation of soluble PAs and diminished production of isoflavones. The role of MtPAR in the distribution of precursors to flavonoid, isoflavonoid, anthocyanin and PA pathways was emphasized by the same study. The HRs of M. truncatula overexpressing MYB2 demonstrated suppression of the expression of genes-encoding dihydroflavonol 4-reductase (DFR1) and anthocyanidin synthase (ANS). Anthocyanin accumulation characteristic of wild-type roots was ceased in the MYB2-overexpressing roots [43]. Wild-type HRs of M. truncatula with the ectopic expression of MtTT8 transcription factor showed enhanced expression of the flavonoid biosynthesis pathway genes and elevated production of PAs and anthocyanins. Contents of anthocyanins (as cyanidin 3-O-glucoside eq.), soluble PAs (as epicatechin eq.) and insoluble PAs (as proanthocyanidin B1 eq.) in the MtTT8-expressing roots reached 600 μg/g FW, 27 μg/g FW and 180 μg/g FW, respectively [44]. Heterologous expression of GhMYB36—a gene for TT2-type MYB transcription factor from a tetraploid Gossypium hirsutum L. (Malvaceae), in the HRs of M. truncatula made it possible to achieve 100 μg/g FW of total soluble PAs (epicatechin eq.) and over 1600 μg/g FW of total insoluble PAs (proanthocyanidin B1 eq.) [44]. VvMybPA1 and VvMybPA2 are transcription factors involved in the regulation of PA biosynthesis in V. vinifera seeds, leaves and exocarp of young berries. The ectopic expression of the genes-encoding these transcription factors in the HRs of V. vinifera caused activation of the enzymes of the flavonoid pathway and enhanced accumulation of PAs (up to 8 mg/g FW). Moreover, the VvMybPA1- and VvMybPA2-expressing transformed roots started to produce PAs with trihydroxylated ring B [39].
The overexpression of VvMYBC2-L1 in grapevine HRs negatively affected PAs biosynthesis, as it can be expected for a member of subgroup 4 MYB transcription factors [41]. A similar effect was achieved in V. vinifera HRs by an overexpression of VvMYBC2-L3 [42]. Transgenic HRs of Litchi chinensis Sonn. (Sapindaceae), overexpressing LcMYB1, contained over 3 mg/g FW of anthocyanidins and nearly 15 mg/g FW of PAs. Wild-type lychee HRs accumulated about 5 mg/g FW PAs and minute amounts of anthocyanidins [49]. Regulatory functions of PpMYB10.1 and MYB 182 transcription factors in anthocyanidin biosynthesis were also investigated using peach and poplar HR cultures, respectively [50,51]. Expression of Camellia sinensis (L.) Kuntze-derived genes-encoding leucoanthocyanidin reductase (LAR) and anthocyanidin reductase (ANR) in the HRs of M. truncatula resulted in slightly elevated content of insoluble PAs. The ectopic expression of CsLAR enhanced the accumulation of anthocyanins, whereas CsANRs expressing roots tend to accumulate more soluble PAs than the control roots [52].
HRs of Tartary buckwheat were frequently used to investigate the effects of different factors on anthocyanins accumulation. Thwe et al. [53] examined seven wild-type R. rhizogenes strains to assess transformation efficiency and anthocyanin content in regenerated hairy root clones of F. tataricum cv. “Hokkai T10”. The R1000 strain, except for the highest transformation efficiency and growth rate of roots, provided high transcript levels for most genes of the flavonoid biosynthetic pathway and the highest contents of cyanidin 3-O-glucoside (800 µg/g DW) and cyanidin 3-O-rutinoside (2410 µg/g DW). A plant growth regulator—ethephon—at the concentration of 0.5 mg/L, significantly increased cyanidin 3-O-glucoside and cyanidin 3-O-rutinoside accumulation in hairy roots of buckwheat. Moreover, upregulation of the genes involved in flavonoid biosynthesis upon ethephon treatment was observed [54]. Overexpression of FtMYB1, FtMYB2, FtMYB3 and FtMYB-like in the HRs of Tartary buckwheat up-regulated the genes of the phenylpropanoid biosynthetic pathway and enhanced anthocyanin production. FtMYB18 overexpression negatively influenced anthocyanin biosynthesis in the roots [9,55].
The black or purple carrot (Daucus carota L. ssp. sativus var. atrorubens Alef, Apiaceae) belongs to the Eastern group of domesticated carrots and is cultivated and commonly eaten in India, Pakistan, Turkey and Afghanistan. In contrast to the Western orange carrots, the roots of the plant accumulated anthocyanins. Acylated anthocyanidin triglycosides produced by the plant, due to their chemical stability, are used as food colorants [56]. The HRs of black carrot were obtained by the infection of taproot and hypocotyl explants of black carrot inbred line “43” and taproot explants of cv. “Night Bird”. The maximum total monomeric anthocyanin content, spectrophotometrically measured, exceeded 3 mg/g DW and was found in the HRs derived from hypocotyl explants, cultivated in ½ MS [57] liquid medium at a photoperiod (12/12 h, light:dark). The content was elevated by the addition of ethephon (200 mg/L) at the 10th day after the inoculation in the fresh medium. The more accurate measurement of anthocyanins content, by UHPLC-PDA, revealed that the black carrot HRs derived from roots of the inbred line “43” accumulated over 4 mg/g DW of the pigments and upon the treatment with ethephon up to 8 mg/g DW. Eight anthocyanins, derivatives of cyanidin and pelargonidin, were identified and quantified using UHPLC-PDA-TOF MS [56]. Supplementation of the nutrient medium with 60 g/L sucrose, increased dry mass and anthocyanin accumulation in the culture. Elicitation with 200 µM of H2O2, on 12th day of culture resulted in about 20% higher anthocyanin content [58].
Two gene-encoding transcription factors: a bHLH gene AmDelila and an R2R3-MYB gene AmRosea1 were concomitantly introduced into the HRs of common snapdragon and induced expression of anthocyanin-related genes in the roots. Further research demonstrated that AmRosea1 alone was enough to start anthocyanin accumulation in the root tissues. The total anthocyanin content in the roots expressing AmRosea1 reached 2 mg/g FW versus 0.3 mg/g FW in the control HRs [59].
Overexpression of PAP1 (production of anthocyanin pigment 1) gene in Panax ginseng C.A. Meyer (Araliaceae) HRs induced expression of phenylpropanoid and flavonoid biosynthetic pathway genes and led to 191- to 341-fold increase in anthocyanins production in comparison to the controls. The increase of anthocyanin accumulation was accompanied by the rise in the antioxidant and radical scavenging activity of PAP1-overexpressing roots and their improved anti-melanogenic activity [60]. Further investigation suggested improved antimicrobial and anti-elastase activities of PAP1-overexpressing ginseng HRs [61].
2.1.2. Flavonols and Flavanols
Except for the optimization of culture conditions (light, growth regulators, sucrose concentration, bacterial strain used to initiate the culture) and overexpression of genes from the myb gene family, elicitation with abiotic (UV-B irradiation) and biotic (yeast extract, chitosan, inactivated fungal preparations) elicitors was used to enhance production of flavonols and flavanols in the HRs. Plant hormones (methyl jasmonate, MeJa) and plant growth regulators (ethephon), in some studies classified as abiotic elicitors [16], were also applied to stimulate biosynthesis of the compounds.
HRs of buckwheat (Fagopyrum esculentum Moench) and Tartary buckwheat, except for anthocyanins and PAs, accumulated substantial amounts of flavonols (kaempferol, quercetin, rutin) and flavanols (catechin, epicatechin). Trotin et al. [62] estimated contents of (+)-catechin, (−)-epicatechin and (−)-epicatechin 3-O-gallate (for structures, see Figure 3) in a HR culture of buckwheat obtained by transformation with R. rhizogenes 15834. (−)-Epicatechin gallate was the major flavanol found in the roots. Its content reached 10 mg/g DW after 21 days of culture and was like that estimated in normal root culture. The HRs produced twice as much of (+)-catechin as normal roots (up to 8 mg/g DW) but the content of (−)-epicatechin in the transformed roots was significantly lower. It is worth noting, that after 21 days in culture dry weight of roots reached 720 mg DW per flask and 180 mg DW per flask for hairy and normal roots, respectively. The HRs of buckwheat accumulated higher yields of rutin (quercetin 3-O-rutinoside, 1.3 mg/g DW) than did untransformed roots (0.5 mg/g DW) [63]. Overexpression of Arabidopsis transcription factor AtMYB12 in buckwheat HRs led to the increased expression levels of flavonoid biosynthetic enzymes and enhanced rutin production in comparison with the control HRs [64]. Roots of Tartary buckwheat cv. “Hokkai T8” and “Hokkai T10”, obtained by infection of hypocotyl explants with R. rhizogenes strain R1000 were cultivated in both light and dark conditions. The roots contained higher amounts of flavonols when exposed to light. Quercetin content reached its maximum (nearly 1 mg/g DW) 5–10 days after the transfer of roots to the fresh medium and then dropped, whereas the highest content of rutin was found after 15 days of culture (59 mg/g DW) [35]. Catechin and rutin contents in the HRs of Tartary buckwheat differed depending on the R. rhizogenes strain that was used to obtain the culture. The quercetin content remained unchanged irrespective of the bacterial strain used [53]. The ethephon treatment (0.5–2.0 mg/mL) significantly increased rutin accumulation in the roots [54]. A similar effect was achieved with overexpression of FtMYB6, an SG7 R2R3-MYB transcription factor induced by light [65]. Overexpression of FtUGT73BE5, an UDP-glycosyltransferase, in the HRs of Tartary buckwheat caused increase in rutin content in the biomass from 0.77 to 1.29-fold when compared with the control (up to 80 mg/g DW) [66]. Choi et al. [67] investigated expression of 11 genes of the phenylpropanoid biosynthetic pathway in seedling roots, adventitious roots and HRs of F. tataricum. Five of the examined genes were highly expressed in the studied HR clones. The HRs accumulated significantly higher amounts of (+)-catechin, (−)-epicatechin and rutin than those found in the untransformed axenic roots and roots of the seedlings.
Tusevski and coworkers [68], by means of HPLC-PAD-ESI-MSn, identified and quantified numerous phenolic compounds produced by HRs of Hypericum perforatum L. (Hypericaceae) obtained by a transformation with R. rhizogenes A4. Flavonols (quercetin 6-C-glucoside, unidentified isorhamnetin-O-hexoside, rutin, kaempferol), flavanol (catechin) and two proanthocyanidin dimers were present in the dark grown roots in small quantities (<1 mg/g DW). Contents of epicatechin and one proanthocyanidin dimer exceeded 1 mg/g DW. The phenolic profile changed when the roots were exposed to light [69]. Selected lines of H. perforatum HRs, transformed with R. rhizogenes A4M70GUS, accumulated up to 4.7 mg/g DW catechin, negligible amounts of procyanidin dimers, and up to 5.9 mg/g DW of quercetin and kaempferol derivatives [70].
Overexpression of LcMYB1 in lychee HRs led to the twofold increase in total flavonol content, in reference to the control roots [49]. The HRs of Linum album Kotschy ex Boiss. (Linaceae) transformed with R. rhizogenes LBA9402, upon elicitation with a cell wall preparation from Piriformospora indica (1%, v/v; 48–120 h treatment), demonstrated enhanced expression of genes-encoding several enzymes of phenylpropanoid biosynthetic pathway and enhanced flavonols and flavanols accumulation. The roots accumulated up to 0.1 mg/g DW catechin, kaempferol (<3.6 µg/mg DW) and myricetin (<2.4 µg/mg DW). The total flavonoid content did not exceed 0.15 mg/g DW [71,72,73]. According to Thiruvengadam et al. [74] HRs of Polygonum multiflorum Thunb. (Polygonaceae) transformed with R. rhizogenes strain KCTC 2703 produced rutin, myricetin, quercetin and kaempferol, although in low quantities (0.35, 0.40, 0.29 and 0.11 mg/g DW, respectively). In the HR cultures of P. multiflorum transformed with R. rhizogenes KCCM 11,879 quercetin was the most abundant flavonol (up to 4.59 mg/g DW). Elicitation with MeJa (50 µM) caused a 3.83-fold increase in quercetin content [75].
A series of papers on HR cultures of Momordica charantia L., Momordica dioica Roxb. ex Willd., Cucumis anguria L. (Cucurbitaceae), Brassica rapa L. ssp. rapa and Brassica rapa L. ssp. pekinensis (Brassicaceae) [76,77,78,79,80,81] described the quantification of flavonols by an UPLC method with UV detection, using a set of commercially available standards comprising: rutin, myricetin, quercetin, kaempferol and catechin. The estimated contents of flavonols were low (<0.8 mg/g DW), except for M. charantia HRs that produced over 1 mg/g DW of catechin.
HRs of Isatis tinctoria L. (Brassicaceae), obtained by inoculation with R. rhizogenes LBA9402 and cultivated in the optimum conditions, produced rutin (94 μg/g DW), quercetin (53 μg/g DW), kaempferol (134 μg/g DW) and isorhamnetin (84 μg/g DW). Contents of the flavonols were higher than those found in the roots of two-year-old I. tinctoria field-grown plants [82]. Elicitation with 150 mg/L of chitosan, for 36 h, led to a significantly higher flavonol accumulation (rutin—812 μg/g DW; quercetin—733 μg/g DW; kaempferol—285 μg/g DW; isorhamnetin—618 μg/g DW) [83]. A co-culture of the HRs with an immobilized Aspergillus niger caused oxidative stress in the plant tissue and triggered up-regulation of the flavonoid biosynthetic pathway. As a result, under the optimum conditions (ca.104 spores/mL, 30 °C, initial pH value 7.0, 72 h treatment), higher contents of all flavonols but rutin were detected in the examined I. tinctoria HRs (quercetin—up to 1500 μg/g DW; isorhamnetin—up to 700 μg/g DW; kaempferol—up to 1000 μg/g DW) [84]. The roots exposed to UV-B radiation (108 kJ/m2) demonstrated substantial up-regulation of the chalcone synthase gene and increased accumulation of all monitored flavonols (rutin—up to 1500 μg/g DW; quercetin—up to 1700 μg/g DW; isorhamnetin—up to 2000 μg/g DW; kaempferol—up to 2200 μg/g DW) [85].
Ghimire et al. [86] found that elicitation either with yeast extract (100 mg/L) or with MeJa (100 μM) significantly increased flavonol accumulation in the HRs of Aster scaber (Asteraceae, full botanical name not provided). Upon elicitation with MeJa, biomass of the roots contained: 2.07 mg/g DW of myricetin, 0.69 mg/g DW of quercetin, 0.26 mg/g DW of kaempferol and 0.19 mg/g DW of rutin. The HRs of Ligularia fischeri Turcz. f. spiciformis (Nakai) (Asteraceae), obtained by inoculation with R. rhizogenes KCTC 2703, were found to produce myricetin (2.38 mg/g DW), quercetin (0.51 mg/g DW, kaempferol (0.12 mg/g DW) and rutin (0.14 mg/g DW) [87].
HRs may be a source of new, previously unknown natural products. Two unusual, biologically active, derivatives of kaempferol and quercetin (4′-O-methylkaempferol-3-O-[(4′′→13′′′)-2′′′,6′′′,10′′′,14′′′-tetramethylhexadecan-13′′′-olyl]-β-D-glucopyranoside and 3′,4′-di-O-methylquercetin-7-O-[(4′′→13′′′)-2′′′,6′′′,10′′′,14′′′-tetramethylhexadec-13′′′-ol-14′′′-enyl]-β-D-glucopyranoside) were isolated from the HRs of Catharanthus roseus (L.) G. Don (Apocynaceae) [88].
2.1.3. Flavones and Flavanones
The procedures employed to enhance the production of flavones and flavanones in the HRs were similar to those described earlier for the other groups of flavonoids and include overexpression of the genes-encoding biosynthetic enzymes (CHI, PAL, C4H and 4CL), elicitation (UV-B irradiation, MeJa, yeast extract, bacterial lysates, β-cyclodextrin, iron oxide nanoparticles) and heterologous expression of transcription factors.
One of the most spectacular examples of a plant that accumulate substantial amounts of flavones is Scutellaria baicalensis Georgi (Lamiaceae). Roots of the plant contain up to 27% DW of flavones, mainly glycosidic derivatives of baicalein, wogonin and oroxylin A [89] (for structures, see Figure 4). S. baicalensis is valued for its medicinal properties [90,91], including the antioxidant activity of its major constituents [92]. First attempts to develop a HR culture system to produce pharmacologically active flavones of S. baicalensis were made in the 1990s [93,94,95]. Those studies, apart from the identification of over twenty known phenolic compounds synthesized by the roots, including flavones and phenylethanoids, led to the isolation and identification of two formerly unknown flavones. However, the yields of biologically active flavones from the cultures were unsatisfactory (up to 1.2% DW of baicalein 7-O-glucuronide). The HRs that emerged as a result of S. baicalensis transformation with R. rhizogenes LBA 9402, in optimum culture conditions produced 6.9% DW of total baicalein glycosides and 1.5% DW of total wogonin glycosides, measured by HPLC after the acidic hydrolysis of the native compounds [96]. Kuzovkina et al. [97,98], by a transformation of S. baicalensis with R. rhizogenes A4 obtained HRs that produced 5% DW of flavonoids (spectrophotometrically measured). The content increased 1.8-fold after the 72 h of treatment with 100 μM of MeJa. More accurate quantification by HPLC revealed 1.27% DW of baicalin (baicalein 7-O-glucuronide), 1.07% DW of wogonoside (wogonin 7-O-glucuronide) and 0.38% DW of their aglycones in the analyzed roots. Maximum yields of baicalin and wogonoside after MeJa treatment reached 10 mg/g DW and 24 mg/g DW, respectively [98,99]. Hirotani et al. [100] found that accumulation of mRNA for an enzyme UDP-glucose:baicalein 7-O-glucosyltransferase (UBGT) in the HRs of S. baicalensis was induced in response to wounding and salicylic acid treatment. Overexpression of chalcone isomerase gene (SbCHI) in the HRs of Baikal skullcap resulted in the increased accumulation of baicalin (42–60 mg/g DW), baicalein (8.9–12.1 mg/g DW) and wogonin (2.2–5.4 mg/g DW) in reference to GUS-control HR line. As expected, SbCHI-silenced HRs produced less flavones than the controls [101]. The HRs overexpressing phenylalanine ammonia-lyase genes (SbPAL1, SbPAL2 or SbPAL3) produced significantly higher amounts of flavone aglycones (baicalein: 11–29 mg/g DW; wogonin 2.5–6.7 mg/g DW) than the GUS-control HRs and wild type field-grown roots. Baicalin content in PAL-overexpressing roots (57–136 mg/g DW) was slightly lower than that in wild type field-grown roots (146 mg/g SbCYP82D2DW) but much higher than that found in the GUS-control HRs [102]. Also, overexpression of cinnamate 4-hydroxylase (C4H) and 4-coumaroyl CoA ligase (4CL) increased flavone production in S. baicalensis HRs [103]. Heterologous expression of the transcription factor Lc from Zea mays in the HRs of Baikal skullcap led to the enhanced accumulation of baicalin, baicalein and wogonin in the roots, but higher yields were achieved when the Arabidopsis PAP1 transcription factor was overexpressed in the same experimental system (up to 102 mg/g DW of baicalin) [104]. Some improvement in yields of flavones from the cultures was achieved as well by an optimization of the nutrient medium composition [96,105,106]. Aglycones:glucuronides ratios in the cultures may be at least partially controlled by the activities of the endogenous β-glucuronidase of Baikal skullcap (baicalinase, sGUS) and baicalein 7-O-glucuronosyltransferase (UBGAT) [107,108]. Hydroxylation of chrysin is the step in the biosynthesis of baicalein and wogonin that requires the respective CYP450 enzymes: flavone 6-hydroxylase (F6H) and flavone 8-hydroxylase (F8H). Two enzymes: SbCYP82D1.1 (F6H) and SbCYP82D2 (F8H) from S. baicalensis were described and the role of SbCYP82D2 in the wogonin biosynthesis was confirmed by its silencing in the HRs of the plant [109]. Baikal skullcap collected in the Dauria region (shared by Mongolia, Russian Federation and China) demonstrated high contents of polymethoxylated flavones. The HRs obtained from the plants retained their metabolic profile [110].
Several other species of the Scutellaria genus like S. lateriflora L., S. andrachnoides Vved., S. bornmuelleri Hausskn. ex Bornm. S. przewalskii Juz. and S. pycnoclada Juz. Bunge were also investigated in respect of their prospective use as a source of biologically active flavones [111,112,113,114,115,116,117]. American skullcap (S. lateriflora) is the second most popular species of Scutellaria used for medicinal purposes. Wilczańska-Barska et al. [111] obtained HRs of S. lateriflora by a transformation with R. rhizogenes strain A4. The roots produced a phenylethanoid glycoside—acteoside (19 mg/g DW) and flavones (scutellarin 0.6 mg/g; baicalin 14.5 mg/g; wogonoside 12.0 mg/g; wogonin 11.5 mg/g; chrysin 0.1 mg/g DW). Except for the wogonin, flavone contents were lower than those in S. baicalensis HRs [111,112]. The root culture of S. lateriflora upon elicitation with yeast extract (YE, 200 mg/L) demonstrated increased biomass accumulation and enhanced acteoside production. The optimum for flavones accumulation was 50 mg/L YE (flavone total content ca. 50 mg/g DW). Bacterial lysates applied as elicitors were less effective [111]. The HRs of the American skullcap transformed with R. rhizogenes ATCC 15834 were cultivated under continuous illumination or in the dark and elicited with β-cyclodextrin and MeJa [113]. Maximum contents of baicalein and wogonin (5.4 mg/g DW and 0.71 mg/g DW, respectively) were found in the cultures maintained in the dark, treated with 15 mM of β-cyclodextrin. Maximum contents of flavone glycosides—scutellarin (scutellarein 7-O-glucuronide) and wogonoside (ca. 0.5 mg/g DW each) were accumulated in the roots grown in the light using the same treatment. β-Cyclodextrin (15 mM) turned out to be more effective than MeJa (100 µM) as an inducer of flavone accumulation, after 24 h treatment. Better yields of the flavones (baicalin—22.5 mg/g DW; wogonin—5.4 mg/g DW) were achieved by Tuan et al. [114] after 72–96 h treatment with 100 µM MeJa. The HRs used in the experiment were obtained by the transformation with R. rhizogenes strain R1000. Stepanova et al. [115] investigated HRs of S. baicalensis, S. lateriflora, S. przewalskii and S. pycnoclada derived from the transformation of the plant material with wild-type R. rhizogenes A4. The roots of S. baicalensis and S. przewalskii showed the best growth indices when cultured in a liquid nutrient medium. Baicalin was the major flavone accumulated in all examined cultures. The HRs of S. przewalskii were characterized by a high content of flavones (ca. 33 mg/g DW in total) compared to the HRs of S. baicalensis (ca. 17 mg/g DW) and HRs of the two remaining species (about 5–13 mg/g DW). The HRs of S. andrachnoides and S. bornmuelleri produced low amounts of flavones [116,117]. The HRs of S. viscidula Bunge, S. orientalis L. and S. araxensis Grossh. have not been examined in respect of their flavone content yet [118,119].
Dracocephalum kotschyi Boiss (Lamiaceae) is a rare plant of medicinal properties that synthesizes polymethoxylated flavones and flavonols of antioxidative and anti-inflammatory activity [120]. The HRs of the plant induced by inoculation with R. rhizogenes LBA 9402 produced: apigenin, cirsimaritin, isokaempferid, penduletin, xanthomicrol and calycopterin, but the contents of individual compounds varied widely depending on the root clone. The HRs were a better source of flavonoids than the roots of the intact plant. The leaves of D. kotchyi, however, contained nearly eight times more flavonoids (1.7 mg/g DW) than the most productive HR clone [121]. A HR culture of D. kotschyi transformed with R. rhizogenes ATCC 15834 accumulated 0.19 mg/g FW of apigenin. Upon elicitation with iron oxide nanoparticles (75 mg/L, 24 h) the apigenin content increased up to 0.37 mg/g FW [122].
The genus Saussurea comprises ca. 400 plant species. Some of the plants from the genus have been traditionally used as medicines [123,124]. The HRs of S. medusa Maxim, transformed with R. rhizogenes strain R1601 and maintained in the liquid N6 medium [125], produced ca. 6.1 mg/g DW jaceosidin [126] in a 24–28-day culture. Overexpression of chalcone isomerase gene from S. medusa in the HRs of S. involucrata Kar. et Kir. ex Maxim., led to the increased accumulation of apigenin (ca. 2.6 mg/g DW) and a higher content of total flavonoids [127].
Erigeron breviscapus (Vaniot) Hand.-Mazz. (Asteraceae) synthesizes pharmacologically active flavones apigenin and scutellarein together with their glycosides. The major flavone accumulated by the plant is scutellarein. The HRs of E. breviscapus were obtained by transformation with R. tumefaciens strain C58C1 harboring pRiA4 plasmid or with the same bacterial strain modified by introducing a construct containing EbCHI gene encoding chalcone isomerase. The roots overexpressing EbCHI produced 2.2 mg/g DW of scutellarin, whereas the roots of the intact plant contained 0.21 mg/g DW of the compound (whole plant 2.6 mg/g DW). Elicitation with MeJa induced expression of several genes engaged in the flavonoid biosynthesis, including the genes-encoding chalcone synthase (CHS) and chalcone isomerase (CHI). The HRs of E. breviscapus treated with MeJa transiently accumulated up to 4.7 mg/g scutellarin [128].
HRs of Catharanthus roseus (L.) G. Don (Apocynaceae) harboring the gene encoding tryptophan feedback-resistant anthranilate synthase holoenzyme (ASαβ) produced naringin (naringenin 7-O-neohesperidoside) as the major phenolic metabolite (0.65–0.86 mg/g DW). Hesperidin (hesperetin 7-O-rutinoside) was accumulated in the roots in smaller amounts (0.11–0.23 mg/g DW) [129].
Silencing of two flavone synthase II (FNSII) genes: GmFNSII 1 and GmFNSII -2, in the HRs of soybean cultivar Hefeng 47, substantially decreased apigenin accumulation and reduced tolerance to salt stress but increased isoflavonoid production in the investigated root clones [130]. Glucose, mannitol, MeJa and NaCl significantly increased expression of GmFNSII 1 and GmFNSII -2 in the HRs of soybean. The motifs responsive to MeJa and glucose were found in the GmFNSII 1 and GmFNSII -2 promoter sequences. It was suggested that the oxidative damage induced by the salt stress may be mitigated by the flavone accumulation [131].
Components of the flavonoid fraction were analyzed in roots, HRs and a cell suspension culture derived from roots of M. truncatula [132]. M. truncatula HRs were obtained by inoculation with R. rhizogenes strain Arqua1. The HRs contained three luteolin glycosides, chrysoeriol and two flavanones: naringenin and liquiritigenin. Extracts from roots of the intact plant analyzed by the same method revealed the presence of seven flavone glycosides, derivatives of luteolin and chrysoeriol. Flavonoid aglycones, as well as flavanones, were not detected in the roots of the plant. Five flavones: orientin (luteolin 8-C-glucoside), vitexin (apigenin 8-C-glucoside), isovitexin (apigenin 6-C-glucoside), luteolin and apigenin were quantified in seeds, leaves, roots, calli and HRs of Cajanus cajan (L.) Millsp. (Fabaceae). The contents of orientin, vitexin and isovitexin in the HRs were higher than those in the roots of the intact plant but much lower than those detected in the plant leaves. Apigenin and luteolin contents in the roots of pigeon pea plants were significantly higher than those in the HRs [133]. An effect of UV-B irradiation on accumulation of phenolic constituents in C. cajan HRs was investigated by Gai et al. [134]. Contents of orientin, vitexin, isovitexin, luteolin and apigenin increased after 2–8 h UV-B treatment and were 1.27–4.44-fold higher than those in the control roots. However, the yields of the individual compounds were low. The content of the major flavone, luteolin, after 2 h of UV-B irradiation reached 19 µg/g DW.
2.1.4. Isoflavonoids
Isoflavonoids, like the remaining flavonoids, are regarded as dietary antioxidants, i.e., compounds that may protect against oxidative stress linked to inflammation and the risk of macromolecule damage by free radicals. The group of compounds includes isoflavones, isoflavanones, isoflavans, rotenoids and pterocarpans. Although they are reported from many plant families, isoflavonoids are particularly abundant in Leguminosae plants. The most known dietary source of the compounds are the yellow-skin seeds of Glycine max [135,136]. While the isoflavones are normally present in relatively low amounts in mature soybean tissues, several attempts were made to develop soybeans that accumulate much higher contents of isoflavones than those found in the wild-type seed.
To study the role of isoflavonoids in the plant resistance to fungal infection, HRs were initiated from two soybean genotypes with different susceptibility to the SDS disease (sudden death syndrome) caused by the soil-borne fungal pathogen, Fusarium solani f. sp. glycines. Daidzein derivatives predominated in the isoflavone fraction extracted both from the soybean hairy roots and intact soybean. The principal isoflavones (genistin, daidzin, their malonyl conjugates and aglycones) and isoflavonoid phytoalexins (coumestrol, coumestrol conjugates and glyceollin) (for structures, see Figure 5) were determined in extracts from the Fusarium-inoculated and non-inoculated hairy roots. Inoculation with F. solani negatively affected accumulation of all monitored isoflavonoids except for glyceollin. The compound demonstrated better antifungal activity against the investigated Fusarium species than the remaining isoflavonoids used in the assay (daidzin, daidzein, genistin, genistein, glycitin, glycitein) [137].
Simultaneous silencing of flavone synthase II genes (GmFNSII-1 and GmFSNII-2) in the HRs of soybean led to the reduction or cessation of apigenin biosynthesis. At the same time, increased accumulation of genistein was observed [130]. Synchronous silencing of FNSII and flavanone-3-hydroxylase (F3H) genes caused an increase of daidzein content in soybean HRs up to 1.55 mg/g DW (0.9 mg/g DW in control roots) [138]. Overexpression of GmMYB100, a gene encoding a R2R3 MYB transcription factor, in G. max HRs resulted in diminished expression of the genes-encoding enzymes engaged in flavonoid biosynthesis and diminished isoflavone content. Silencing of the GmMYB100 did not affect expression of the genes for the biosynthetic enzymes (GmCHS7, GmCHS8, GmCHI, GmIFS, GmF3H) but significantly increased isoflavonoid content [139]. Soybean cotyledon HRs harboring GmIFS1 showed better tolerance to salt stress and increased isoflavone content under salt stress conditions [140]. GmMYB58 and GmMYB205 are seed-specific flavonoid biosynthesis activators. Their overexpression in soybean HRs increased transcription of: GmCHS8, GmCHI, GmIFS2, GmFLS1, UGT73F2 (isoflavone UDP-glucosyltransferase gene) and IF7Mat (isoflavone 7-O-glucoside-6”-O-malonyltransferase gene). An unidentified daidzein derivative, which dominated the isoflavonoid profile of the control roots (1 mg/g DW), was excessively produced by the transgenic HRs. The daidzein derivative content reached 3.2 mg/g DW in GmMYB58-overexpressing HRs and 6.7 mg/g DW in the roots overexpressing GmMYB205 [141]. Soybean isoflavonoids are mainly glycoconjugates and their biosynthesis is catalyzed by different UDP-glycosyltransferases (UGT). Six genes for GmUGTs were overexpressed in soybean HRs. In the transgenic root lines overexpressing UGT72Z3, UGT73C20 and UGT88E19, the total content of isoflavonoids increased 1.1- to 1.6-fold in reference to the control and reached over 7 mg/g DW [142]. Increased resistance to Phytophthora sojae infection was observed in G. max HRs overexpressing GmCHI1A (chalcone isomerase gene from the soybean cv. Nannong 10-1). This observation agreed with experimentally proven induction of GmCHI1A expression and enhancement of daidzein accumulation by P. sojae [143]. Fungal infection in soybean seedlings enhance also glycitein production. The last step in the biosynthesis of this compound is methylation catalyzed by isoflavone O-methyltransferase (IOMT). IOMT, when co-expressed with the flavonoid 6-hydroxylase gene (F6H, normally not expressed in the HRs) increased the content of glycitein-related metabolites by ca. 100% compared to the control [144]. The HRs overexpressing GmMaT2 and GmMaT4, genes-encoding malonyl-CoA:flavonoid acyltransferases, produced more malonyldaidzin, malonylgenistin and malonylglucitin than the control. The major isoflavone, malonyldaidzin, content reached ca. 6.3 mg/g FW in GmMaT2 overexpressing HRs. GmMaT2 but not GmMaT4 knockdown resulted in the reduced accumulation of malonylated isoflavonoid glycosides and reduced nodule numbers. GmMaT2 is also upregulated by the rhizobial infection what implicates its participation in the nodulation process and malonylated isoflavone secretion into the rhizosphere [145]. An attempt was made to enhance isoflavonoid production in soybean HRs by elicitation with MeJa (100 μM), salicylic acid (SA, 200 μM), sonication and vacuum infiltration. Treatment with MeJa (72 h) resulted in 10.67-fold higher total isoflavonoid content (53.16 mg/g DW) than that in the untreated control. Daidzin content reached ca. 33.9 mg/g DW. Elicitation with SA, after 96 h, caused 5.78- and 65-fold increase in total isoflavonoid and genistein contents, respectively. Maximum isoflavonoid production, 75.26 mg/g DW, was achieved with 2 min sonication and subsequent 2 min vacuum infiltration of HRs on 30th day of the culture [146].
Pigeon pea HRs produced genistin (0.11 mg/g DW) and genistein (0.04 mg/g DW) [133]. Their accumulation in the HRs was enhanced by an exposition to UV-B radiation [134]. Leaves of Lotus japonicus cv. Miyakojima line MG-20 seedlings supplied with genistein produced isoprenylated isoflavonoid, wighteone. This observation led to the identification of novel prenyltransferase gene LjG6DT. The enzyme encoded by the gene worked only with genistein as a substrate and its expression was induced by the reduced glutathione (GSH), MeJa and SA. Overexpression of LjG6DT in L. japonicus HRs, together with GSH and genistein supplementation, induced enhanced wighteone production (0.18–0.24 µg/g DW) [147]. Twenty isoflavonoids, derivatives of 2ʹ-hydroxyformononetin, afrormosin, biochanin A, daidzein, formononetin, genistein and irisolidone were detected in the HRs of M. truncatula. In the roots of the intact plant 32 isoflavonoids were detected and tentatively identified. In contrast to the plant roots HRs preferably accumulated isoflavonoid aglycones [132].
Red clover (Trifolium pratense L., Fabaceae) is a forage legume producing formononetin, biochanin A, daidzein and genistein as major isoflavone constituents [148]. Kumar et al. [149] described stable production of isoflavones by HRs of T. pratense. One of the fast-growing HRs clones, displayed a high accumulation of all four pharmaceutically important isoflavones: daidzein (8.56 mg/g DW), genistein (2.45 mg/g DW), formononetin (15.23 mg/g DW) and biochanin A (1.10 mg/g DW). The HRs of T. pratense var. URS-BRS Mesclador were obtained by inoculation with R. rhizogenes strain A4TC24 and 23 isoflavonoids were tentatively identified as their metabolites. The isoflavonoids were, putatively, derivatives of: daidzein, pseudobaptigenin, pratensin, biochanin, irilone, formononetin and 3ʹ,7-di-O-methylorobol. Isoflavonoid contents were monitored in the selected clones of roots up to five months of culture. One of the examined clones (6HR) accumulated 5.5 mg/g DW biochanin A, 20.8 mg/g DW formononetin and ca. 0.7 mg/g genistein after 90 days of culture. The HRs were elicited, 7 days after subculture, using either elevated sucrose content in the medium (60 g/L) or SA (10 and 30 mg/L). Sucrose (60 g/L) increased accumulation of isoflavonoids in the HRs. After 3.5 days of treatment, roots of clone 4HR contained 12.8 mg/g DW daidzein, 0.7 mg/g DW genistein, 13.4 mg/g DW formononetin and 1.1 mg/g DW biochanin A, whereas the roots of clone 8HR produced 1.9 mg/g DW daidzein, 0.4 mg/g DW genistein, 14.8 mg/g DW formononetin and 4.1 mg/g DW biochanin A. Untreated roots (controls) accumulated 1.2–2.9 mg/g DW daidzein, 0.1–1.3 mg/g DW genistein, 2.1–3.0 mg/g DW formononetin and 0.7–4.4 mg/g DW biochanin A. Elicitation with SA was less effective than the treatment with sucrose [150].
Astragalus membranaceus (Fisch.) Bunge (proper name A. propinquus Schischkin, Fabaceae) has a long history of use in traditional Chinese medicine. Roots of the plant are also commercially available in Europe and USA as dietary supplements and functional foodstuffs. Calycosin and calycosin 7-O-glucoside were the major isoflavone components of the HRs derived from the plant. The two compounds, together with ononin (formononetin 7-O-glucoside), formononetin and astraisoflavan 7-O-glucoside were quali-quantitatively determined by LC-MS/MS. Under optimum conditions, the total isoflavonoid content in 34-day old A. membranaceus HRs reached 0.24 mg/g DW. This yield was significantly higher compared to that of three-year-old field-grown roots (0.19 mg/g DW) [151,152]. The HRs exposed to UV-B (86.4 kJ/m2) synthesized more isoflavonoids (up to 0.53 mg/g DW). All investigated genes involved in isoflavonoid biosynthesis were also up-regulated following the UV-B irradiation. PAL and C4H were found to be the key genes implicated in the control of the process [153]. Elicitation of 34 days old HRs of A. membranaceus with MeJa (283 µM, 33.75 h treatment) raised the content of isoflavonoids to 2.25 mg/g DW and upregulated the genes related to isoflavone biosynthesis [154]. A co-culture of A. membranaceus HRs with immobilized Aspergillus niger enhanced accumulation of calycosin (0.73 mg/g DW) and formononetin (1.12 mg/g DW) in the roots [155].
Psoralea corylifolia L. (Babchi, Fabaceae) is a plant used in traditional medicine of China and India. The plant accumulates coumarins, flavonoids and terpenophenols, including pharmacologically active bakuchiol, which gained some popularity as a component of cosmetics [156,157]. The first study on isoflavonoid production in the HRs of Psoralea spp. was published in 1999 [158]. HR lines derived from seven distinct species were examined in respect of their daidzein content. The examination revealed that P. leucantha F. Muell. and P. lachnostachys F. Muell HRs were the best isoflavonoid producers. One, highly productive line (daidzein 10.2 mg/g DW, coumestrol 0.48 mg/g DW) was chosen for further investigation. The roots were elicited with 30 mg/L of chitosan at the end of the exponential phase of growth (21st day). After the addition of chitosan, the content of daidzein in biomass dropped to ca. 8 mg/g DW due to the release of the compound into the culture medium. Coumestrol and genistein contents in the roots increased from ca. 0.40 to 0.52 mg/g DW and from ca. 0.03 to 0.07 mg/g DW, respectively. Coumestrol was also liberated to the medium after the elicitation with chitosan. HR cultures of P. corylifolia L., transformed with R. rhizogenes ATCC 15834, similarly to the control roots, accumulated daidzin as a major isoflavonoide. The compound was accompanied with smaller amounts of formononetin glucoside, genistin and daidzein. Bakuchiol was absent from the cultures [159]. Clones of P. corylifolia HRs, obtained by inoculation with R. rhizogenes LBA 9402, in optimum conditions produced up to 20.6 mg/g DW daidzein and up to 3.7 mg/g DW genistein [160].
Pueraria spp. (Fabaceae) due to isoflavonoid content is traditionally used to relieve menopausal symptoms [161]. The HRs of Pueraria phaseoloides (Roxb.) Benth induced by R. rhizogenes ATCC 15834 contained ca. 1 mg/g DW puerarin (daidzein 8-C-glucoside) [162]. The culture was scaled up to a 2.5 L bioreactor and the culture medium was modified to achieve better yield. In optimum conditions, roots cultivated in the bioreactor produced ca. 5.6 mg/g DW of puerarin and the product was partially liberated to the nutrient medium [163]. A HR culture of P. candollei Wall. Ex Benth. was established using R. rhizogenes ATCC 15834. The total isoflavonoid content in the roots reached 36.48 mg/g DW. Daidzin was the major product (29.9 mg/g DW), and the puerarin content was estimated as 3.4 mg/g DW [164]. Chitosan (50, 100 and 150 mg/L), MeJa (50, 100 and 200 µM), SA (50,100 and 200 µM), YE (0.5, 1 and 2 mg/mL) and autoclaved Rhizobium culture (1, 2 and 3% v/v) were applied as elicitors to enhance isoflavonoid production in the HRs. Though all of the applied treatments increased isoflavonoid accumulation, the yeast extract addition (0.5 mg/mL, 3 days) proved to be the most effective (60.5 mg/g DW total isoflavonoids) [165]. P. candollei var. myrifica (Airy Shaw & Suvat.) Niyomdham, except for the isoflavonoids, synthesizes another phytoestrogen, deoxymiroestrol. The HRs of the plant produced up to 7 mg/g DW isoflavonoids (mainly daidzin and genistin) and up to 77 µg/g DW of deoxymiroestrol. The contents were higher than those found in the roots of the intact plant (3.5 mg/g DW and 15 µg/g DW, respectively). Chitosan, YE and MeJa were applied to study effects of elicitation on the active metabolite content. The HRs elicited with MeJa (200 µM) accumulated puerarin (0.54 mg/g DW), daidzin (8.68 mg/g DW), genistin (5.27 mg/g DW), daidzein (0.45 mg/g DW), genistein (0.16 mg/g DW), kwakhurin (1.39 mg/g DW) and deoxymiroestrol (0.25 mg/g DW), after 6 days of the treatment. The remaining elicitors also induced productivity of the roots, though to a lesser extent [166]. P. candollei Grah. ex. Benth. var. candollei was genetically transformed using two different R. rhizogenes strains (ATCC 15834 and 43,057) to obtan several clones of HRs.
The effect of the inoculum size (1 and 2% w/v) and temperature (25 or 32 °C) on the growth of the obtained HRs and on their isoflavonoid content was investigated. The maximum flavonoid accumulation (31 mg/g DW with daidzein and puerarin as major compounds) was found in the cultures started with 1% inoculum and cultivated at 32 °C. Moreover, it was found that cultivation of the HRs at higher temperature reduced browning of the tissue [167]. Kim et al. [168] obtained HRs from two different lines of P. lobata (Willd.) Ohwi (kudzu), collected in two different regions of Korea. The roots accumulated more puerarin and daidzin than the respective callus cultures, but the contents of the compounds in the tubers of the intact plants were higher. P. lobata C-glycosyltransferase (PlUGT43) uses daidzein and genistein as substrates. The overexpression of PlUGT43 in soybean hairy roots that synthesize daidzein, but not puerarin, led to the production of puerarin in the transgenic roots [169].
The transformed root cultures of Ononis spinosa L. and Ononis arvensis L. (Fabaceae) were obtained by Nóra Gampe et al. [170]. The most abundant compounds in the HRs were medicarpin, sativanone and pseudobaptigenin glucosides (16.9–28.9 mg/g DW, 1.3–11.4 mg/g DW and 0.9–2.0 mg/g DW, respectively). Formononetin and onogenin derivatives were present in smaller amounts. Two new phenolic compounds were found in the HRs (bulatlactone 2”-O-glucoside and ononilactone). The total isoflavonoid production in the cultures was comparable to that in the wild-grown O. arvensis and approximately twofold higher than that in wild-grown O. spinosa samples.
2.1.5. Miscellaneous Flavonoids
Though the HRs of Glycyrrhiza glabra L. (Fabaceae) do not produce glycyrrhizic acid, a sweet-tasting saponin responsible for the major pharmacological activity of the liquorice root (peptic ulcer healing), they synthesize vast array of biologically active polyphenols of unique structures [171,172] (see Figure 6). Two new compounds, licoagrochalcone and licoagrocarpin (chalcone- and pterocarpan-type compounds), were isolated from the HRs of licorice transformed with a Rhizobacterium strain harboring pRi15834 and pBI121 (GUS). Eight known compounds were also found in the HRs, including three chalcones, four prenylated flavanones and one prenylated flavanol [173]. Further investigation of the same plant material led to the isolation of licoagrodione, another new compound, together with five known flavonoids. The ntimicrobial activities of the isolated compounds were assessed by the disc diffusion method. Glyinflanin K was the only compound that did not show activity against the bacteria and fungi used in the experiment [174]. The continuation of this research allowed for the isolation of unusual prenylated biaurone, licoagrone, together with five known flavonoids (kanzonol D, afrormosin, odoratin, phaseol, echinatin) [175]. Li et al. [176] isolated and described a new biflavonoid, licoagrodin and another four new flavonoids (licoagrochalcone B, licoagrochalcone C, licoagrochalcone D and licoagroaurone), as well as four known flavonoids. More polar fractions separated from the HRs gave licoagroside A, ononin, calycosin 7-O-glucoside, wistin, vicenin-2, afrormosin 7-O-(6”-malonylglucoside) and isoschaftoside. Licoagroisoflavon and licoagrosides C-F were isolated as new compounds from the HRs of G. pallidiflora Maxim. The compounds were accompanied by eleven known flavonoids [177,178]. In the HRs of G. uralensis Fisch. two-fold increase in total flavonoid content (up to 30 mg/g DW) was observed following the combined 48 h of treatment with YE (0.1%) and polyethylene glycol (PEG8000, 2%) [179]. Tween 80, added to 20-day-old HRs of G. uralensis, caused enhanced production of licochalcone A accompanied by elevated mRNA levels for PAL, 4CL (4-coumarate:coenzyme A ligase) and C4H. After 15 days of the treatment, the roots yielded over 3 mg of licochalcone A per flask, and the product was almost entirely liberated to the culture medium. The control HRs accumulated up to 0.35 mg/flask of the compound in their biomass [180]. The verexpression of CHS in G. uralensis HRs resulted in the increased production of chalcones and liquiritigenin in the transgenic roots. The number of CHS copies in the examined clones of HRs was determined as 9, 10, 11, 13 and 18. The root clone with nine copies of CHS accumulated the highest contents of flavonoids [181].
The Genista tinctoria L. HRs synthesized neither isoflavone, derivatives of daidzein and genistein, nor the derivatives of apigenin and luteolin characteristic of the intact plant. Instead, the roots accumulated isoliquiritigenin (daidzein precursor, 23 mg/g DW) absent from the roots of the intact plant. Abscisic acid (ABA, 37.8 μM), added to the culture on the 42nd day of growth, induced the release of the product (80%) to the culture medium [182].
Alikaridis et al. [183] investigated roots obtained by a transformation of Silybum marianum Gaertner with R. rhizogenes ATCC 15834. The roots contained only minute amounts (0.4 μg/g DW) of silychristin and silydianin. Silybins A and B were not detected, whereas the contents of isosilybins A and B were higher than those in the untransformed roots (isosilybin A—0.01 mg/g DW). The S. marianum HRs examined by Rahnama et al. [184] contained up to 0.14 mg/g DW silychristin. The other flavonolignans were present in smaller amounts.
2.2. Stilbenoids
Stilbenoids are a class of polyphenolic plant constituents that have the general structural formula C6-C2-C6 and share an initial part of their biosynthetic pathway with flavonoids. The compounds could be found in various plant species, including peanut (Fabaceae), grapevine (Vitaceae), berries (Ericaceae), pine (Pinaceae) and tomato (Solanaceae). Their main function is the protection of the host plant against pathogen infestation and oxidative stress generated by different environmental stimuli [185]. Stilbenoids, due to their biological activity, may find application as protective agents in cardiovascular disease, diabetes, neurodegeneration, obesity and other ailments [186].
The compounds were primarily detected in elicited HRs and the procedures used to enhance the productivity of the cultured roots included elicitation (MeJa, paraqat, H2O2) with concomitant use of permeabilizing agent (methyl-β-cyclodextrin) to liberate the products into the culture medium. R2R3-MYB-type transcription factors are engaged in the up-regulation of stilbenoid biosynthesis, and this may find an application in genetic engineering.
Medina-Boliwar et al. [187] found that HRs obtained by the transformation of Arachis hypogaea L. cv. Andru II (Fabaceae) with R. rhizogenes ATCC 15834, upon elicitation with sodium acetate, produced stilbenoids that are excreted into the culture medium. Trans-resveratrol and trans-pterostilbene (for structures, see Figure 7) were identified as metabolites present in the elicited cultures. Trans-resveratrol, trans-arachidin-1 and trans-arachidin-3 were isolated from the HRs of peanut cv. Hull elicited with 10.2 mM sodium acetate. Among the three compounds, arachidin-1 was the most active as an inhibitor of lipoprotein oxidation. In the applied assay, arachidin-1 was more active than butylohydroxytoluen (BHT). In contrast to resveratrol, arachidins at a dose of 55 µM showed some cytotoxicity towards HeLa and RAW 264.7 cells [188]. The same culture was used to investigate the effects of medium optimization, and the age of the elicited roots on growth performance and root culture phenotype. The effect of elicitation on biosynthesis and liberation of stilbenoids into the culture medium was also studied. The HRs elicited on the ninth day of the culture produced trans-resveratrol (ca. 90 µg/g DW), trans-arachidin-1 (ca. 33 µg/g DW), and trans-arachidin-3 (ca. 37 µg/g DW) when cultivated in modified MS instead of Gamborg’s B5 nutrient medium. As the stilbenoids were secreted into the medium, the biomass of HRs contained only minute amounts of the compounds (ca. 1 µg/g DW, ca. 0.75 µg/g DW and ca. 0.71 µg/g DW, respectively) [189]. In a search for the optimum elicitation method, hydrogen peroxide (10 mM), MeJa (100 µM), methyl-β-cyclodextrin (9 g/L), and combination of MeJa with methyl-β-cyclodextrin were used as elicitors, instead of sodium acetate. The application of either H2O2 or cyclodextrin caused secretion of piceatannol, another stilbenoid formerly not found in the HRs of A. hypogaea cv. Hull. The contents of stilbenoids in the culture medium were monitored until 96 h after elicitation. Individual compounds reached their maximum content in the analyzed medium at different time intervals. The best results were achieved with HRs treated with the combination of MeJa and cyclodextrin. Sixty hours after elicitation, the roots produced resveratrol (ca. 5.3 mg/g DW), piceatannol (ca. 0.3 mg/g DW), arachidin-1 (ca. 4 mg/g DW) and arachidin-3 (ca. 17.1 mg/g DW) [190].
Yang et al. [191] found that the prenyl subunit of the prenylated stilbenoids like arachidins originated from the plastidic terpenoid pathway. Moreover, they purified and described membrane-bound stilbenoid-specific prenyltransferase from peanut HRs. Elicitor-treated A. hypogaea HRs were used to discover genes-encoding stilbenoid prenyltransferases. Transcripts encoding five enzymes were identified and two of the enzymes were characterized, including AhR4DT-1 that catalyzes prenylation of resveratrol at C-4 to form arachidin-2 [192]. The prenylated stilbenoids of the peanut manifest an array of interesting biological activities (e.g., anti-inflammatory, antiviral, antioxidant and cytotoxic towards human cancer cells) and have better bioavailability than resveratrol [193,194,195]. To achieve higher yields of the prenylated stilbenoids from peanut HRs, the elicitation procedure was further optimized using 125 µM MeJa (signaling molecule implicated in secondary metabolism regulation), 18 g/L cyclodextrin (permeabilizing agent that may trap the product and prevent feedback inhibition), 3 mM H2O2 (inducer of piceatannol production in the HRs of the peanut) and 1 mM MgCl2 (Mg2+ as a co-factor of resveratrol prenyltransferases) [194]. The formerly applied treatment [190] allowed for ca. 56 mg/L of arachidin-1 and ca. 148 mg/L of arachidin-3 to be obtained. With the new procedure, the yields increased to 227.4 mg/L and 370.6 mg/L, respectively. Moreover, arachidin-2 (83.1 mg/L) and arachidin-5 (68.4 mg/L) were produced by the HRs. The new optimized elicitation method was used to induce stilbenoid production in A. ipaensis and A. duranensis HRs, obtained in the same way as HRs of the peanut cv. Hull. The HRs of the wild relatives of peanut, upon elicitation synthesized less arachidin-1 and arachidin-3, but more arachidin-2 and arachidin-5 [194].
The HRs of A. hypogaea cv. Tainan9 were obtained by inoculation with R. rhizogenes strain K599 (NCPPB 2659). The production of stilbenoids in the roots was induced by the treatment of nine-day-old HRs with 100 µM MeJa in combination with 6.87 mM methyl-β-cyclodextrin. After 24 h of treatment, resveratrol (72.0 µg/g DW), arachidin-1 (179.3 µg resveratrol eq./g DW) and arachidin-3 (21 µg resveratrol eq./g DW) were found in the culture medium. MeJa applied alone did not induce the liberation of stilbenoids into the medium. Liquid chromatography with tandem mass spectrometry (LC-MS/MS) was applied to analyze the metabolites present in the spent medium from the elicited cultures. Except for the phenolic acids (hydroxybenzoic acid and caffeic acid), the following stilbenoids were detected in the medium; trans-piceatannol, trans- and cis-resveratrol, three isomers of trans-arachidin-1, 4-isopentadienyl-3,5,3’,4’-tetrahydroxystilbene, two isomers of trans-arachidin-3, two isomers of arahypin 7 and trans-3’-isopentadienyl-3,5,4’-trihydroxystilbene [196].
Another elicitation strategy was used with the HRs of A. hypogaea cv. Kalasin2 transformed with R. rhizogenes K599. Paraquat (PQ), a broad-spectrum herbicide that generates reactive oxygen species (ROS) in the plant tissue, was used in combination with MeJa and cyclodextrin. As was in the case of MeJa, PQ alone did not induce the liberation of stilbenoids from the roots. The most effective elicitation procedure that utilized 24 h of pretreatment with 500 µM PQ followed by the induction with 100 µM MeJa in combination with 6.87 mM methyl-β-cyclodextrin caused a sharp increase in stilbenoid biosynthesis. The elicited roots produced trans-resveratrol (1.3 mg/g DW, after 120 h), trans-arachidin-1 (180.1 mg/g DW, after 192 h) and trans-arachidin-3 (444.2 mg/g DW, after 120 h). LC-MS/MS analysis of the extract from the spent nutrient medium revealed the presence of hydroxybenzoic acid, two trans-piceatannol isomers, trans- and cis-resveratrol, jasmonic acid, four trans-arachidin-1 isomers, trans-arachidin-3, two isomers of arahypin 7 and arahypins 5 and 6 [197].
Wongshaya et al. [198] investigated an effect of mechanical stress (cutting) and light on stilbenoid biosynthesis in A. hypogaea cv. Tainan9 HRs. Mechanical stress increased the total phenolic content and antioxidant capacity of the cultures in both light and dark conditions. Roots cultivated in the dark showed a better response to elicitation than those grown in the light. Maximum yields of stilbenoids that were achieved using the elicitation procedure described earlier [197] were as follows: trans-resveratrol > 4 mg/g DW (uncut roots, dark, 72 h), trans-arachidin-1 ca. 240 mg/g DW (uncut roots, dark, 72 h), trans-arachidin-3 ca. 250 mg/g DW (uncut roots, dark, 72 h). The same elicitation procedure was applied to the A. hypogaea cv. Tainan9 HRs cultivated in a 5 L capacity stirred tank bioreactor or in 500 mL Erlenmayer flasks. Two densities of inoculum were evaluated: 5 g/L and 20 g/L. The optimum production of arachidins was achieved using 20 g/L inoculum grown in a 500 mL flask. Upon elicitation (72 h treatment), the HRs yielded ca. 1700 mg/L of trans-arachidin-1 and ca. 4800 mg/L of trans-arachidin-3. The roots grown in the bioreactor showed maximum productivity with 20 g/L inoculum, after 48 h treatment [199].
Three cultivars of peanut, including “Tifrunner,” “Hull,” and “Georgia Green” were transformed with R. rhizogenes ATCC 15843. The obtained HRs were treated with 125 µM MeJa, 18 g/L cyclodextrin, 3 mM H2O2 and 1 mM MgCl2 for 168 h. The extracts from the spent culture media were qualitatively and quantitatively analyzed to assess the content of stilbenoids. The HRs of the “Tifrunner” cultivar were found to be rich in arachidins 1, 2, 5 and 6, whereas the HRs of “Hull” cultivar preferably synthesized resveratrol and arachidin-3. The yields of the analyzed stilbenoids ranged from ca. 5 mg/L (arachidin-5 in “Georgia Green”) to ca. 170 mg/L (arachidin-1 in “Tifrunner”) [200].
Muscadine grape (Vitis rotundifolia Michx.), a grapevine species native to the southeastern part of North America, is a source of polyphenols that possess antioxidative and antimicrobial activity [201]. The HRs of V. rotundifolia (21 days old) were treated with 100 µM MeJa for 24 h. Extracts prepared from the root biomass and from the spent nutrient medium were subsequently analyzed in a search for stilbenoid metabolites. Resveratrol, piceid (resveratrol 3-O-glucoside) and ε-viniferin (resveratrol dehydrodimer) were present mainly in the biomass, whereas piceatannol was detected exclusively in the culture medium. Growth regulators IBA (0.05 mg/L) and BAP (0.05 mg/L) added to the culture medium did not affect yields of stilbenoids. Piceatannol and ε-viniferin were the best antioxidants among the analyzed HRs metabolites [202].
Ñopo-Olazabal et al. [203] used the HRs of muscadine grape to study the biochemical and molecular regulation of stilbenoid biosynthesis upon treatment with either 100 μM MeJA or with 10 mM H2O2, over a 96 h period. Both treatments induced the transcription of PAL, STS and resveratrol synthase gene (RS) as soon as 3 h after elicitation. Resveratrol, piceid, and ε-viniferin were identified in the control and in the elicited HRs. Except for resveratrol, the stilbenoids were accumulated in the roots. After the elicitation with MeJa, piceid content in the roots increased from 164 µg/g DW (0 h) to 337 µg/g DW at the end of experiment. The increase, however, was similar to that in the control roots. In the culture medium, piceid was undetectable until 12 h of the treatment and reached 4.7 µg/g DW at the end of the experiment. Maximum accumulation of resveratrol took place at 12 h after elicitation (106 µg/g DW) and then the content of the compound in the roots declined. The resveratrol content in the medium reached maximum at 18 h after exposition to MeJa (48 µg/g DW) and subsequently decreased to 6.4 µg/g DW at the 96 h. ε-Viniferin accumulated in the MeJa elicited roots up to 379 µg/g DW at the end of the experiment. The content was over twofold higher than that in the control roots. The culture medium contained up to 11.8 µg/g DW ε-viniferin. Hydrogen peroxide treatment was less effective as an inductor of stilbenoid biosynthesis. The piceid and resveratrol contents in the H2O2 treated roots increased to a lesser extent than it was observed for MeJa treatment. ε-Viniferin accumulated in the biomass (up to 434 µg/g DW, 96 h) and only ca. 5 µg/g DW of the compound was found in the nutrient medium. At 24 and 96 h after the treatment with H2O2, ca. 72.5 µg/g DW and 24.6 µg/g DW of resveratrol, respectively, were found in the nutrient medium. In the studied cultures, an increase in stilbenoid content correlated with an increased antioxidant capacity. The HRs of V. vinifera Pinot Noir cv. PN40024, obtained by inoculation with R. rhizogenes ATCC 15834 were cultivated in ½ SH (Schenk and Hildebrandt) medium supplemented with 2% sucrose. Piceid, resveratrol and two resveratrol dehydrodimers: ε-viniferin and δ-viniferin were constitutively present in the roots. δ-Viniferin was the major component of the stilbenoid fraction. The total stilbenoid content reached 217 µg/flask at the stationary phase of culture. The elicitation with MeJa (100 µM or 200 µM) on the 18th day of the culture (before the end of the exponential growth phase) increased an excretion rate of stilbenoids from 11% (control roots, after 10 days of the experiment) to 37% (200 µM MeJa, 10-day treatment). The control roots contained ca. 2.42 mg/g DW of stilbenoids at the end of experiment. The HRs elicited with 100 and 200 µM MeJa accumulated 6.98 mg/g DW and 4.34 mg/g DW of stilbenoids, respectively. The maximum stilbenoid content in the nutrient medium (19 mg/L) was found 10 days after the addition of MeJa (final concentration 200 µM). The concomitant use of MeJa (100 µM, added on the 18th day of culture) and methyl-β-cyclodextrins (30, 50 and 70 mM, added to the fresh culture medium before autoclaving) resulted in the increased stilbenoid contents in both root biomass and the culture medium. Maximum total stilbenoid contents, measured four days after the addition of jasmonate were 6.4 mg/g DW in the roots (30 mM cyclodextrins) and 165 mg/L in the medium (50 mM cyclodextrins). The combined treatment with MeJa and cyclodextrins caused enhanced liberation of stilbenoids (80–90%) into the nutrient medium [204]. Tisserant et al. [205], based on 13C and 1H NMR data and results of LC-MS analysis, identified major metabolites of V. vinifera Pinot Noir HRs. The main polyphenols found in the culture were stilbenoids (trans-resveratrol, trans-piceatannol, pallidol, ε-viniferin, scirpusin A, vitisin B and maackin) and flavanones (eriodictyol and naringenin).
Höll et al. [206] found that two R2R3-MYB-type transcription factors from grapevines regulated the stilbenoid biosynthetic pathway by the activation of the promoters of genes-encoding stilbene synthases (STS). One of the transcription factors, MYB15, expressed in the HRs of V. vinifera cv. Chardonnay increased levels of STS and PAL transcription. The content of trans-piceid in the MYB15 transgenic HR lines was fivefold higher than that in the control roots. Accumulation of the remaining stilbenoids was less affected.
V. vinifera subsp. sylvestris, accessions W2 and W16 and cv. Rasha, were transformed using three different strains of R. rhizogenes: ArA4, Ar318 and LBA 9402. Although all the three bacterial strains induced HRs from the grapevine explants, HR lines obtained with the strain ArA4 showed the most vigorous growth. The highest resveratrol contents, 2 to 31 times higher than that found in the control roots, were found in the HRs obtained from internodal explants. The HR cultures originated from cv. Rasha, accession W2 and accession W16 contained up to 95, 256 and 273 µg/g DW of resveratrol, respectively. The elicitation with MeJa or sodium acetate caused approximately a twofold higher production of the stilbenoid and enhanced release of the product into the culture medium [207]
Ectopic expression of the genes-encoding enzymes and transcription factors engaged in stilbenoid biosynthesis and metabolism in the HRs of tobacco led to the production of biologically active stilbenoids. V. vinifera resveratrol-O-methyltransferase (VvROMT) and human cytochrome P450 hydroxylase 1B1 (HsCYP1B1) catalyze methylation of trans-resveratrol to trans-pterostilbene and hydroxylation of trans-resveratrol to trans-piceatannol. Tobacco leaf segments were inoculated with R. rhizogenes harboring the pRiA4 plasmid alone or pRiA4 with the binary plant expression vector pK7WG2_CYP1B1 or pJCN52_ROMT for the HsCYP1B1 or VvROMT genes, respectively. The transgenic HRs expressing CYP1B1 converted exogenously added trans-resveratrol into piceatannol, especially when treated with methyl-β-cyclodextrin as the permeabilizing agent. Biotransformation rates were: 0.4% in wildtype HRs and 1.4–1.6% in the transgenic HRs lines. In contrast to the transgenic HRs, piceid was the major biotransformation product in the wild-type HRs. The yields of piceatannol from the transgenic HR cultures carrying CYP1B1 and wild type HRs were 1888 µg/L and 819 µg/L, respectively. Addition of cyclodextrin to the medium may enhance the productivity up to 7 mg/L. Transgenic HRs carrying VvROMT metabolized exogenously added trans-resveratrol into trans-pterostilbene, piceid and piceatannol. Pterostilbene was found both in the roots and the culture medium. Maximum accumulation of the compound (2.6 µg/L) was observed 24 h after resveratrol feeding [208]. Tobacco HRs carrying VvSTS (gene for stilbene synthase from V. vinifera) and AtMYB12 (gene for the transcription factor from Arabidopsis thaliana) produced over 450 µg/L stilbenoids and more flavonoids than the wild type HRs [8].
The R. rhizogenes-transformed roots of pigeon pea occurred to be an excellent source of cajaninstilbene acid, superior to seeds, leaves and roots of the parent plant [209]. The elicitation of HRs by 10 h of exposure to UV-B radiation resulted in over a twofold increased accumulation of cajaninstilbene acid (up to 6.6 mg/g DW). The radiation induced oxidative stress and caused damage to the roots, which, in turn, enhanced the production of signal molecule, salicylic acid, implicated in the regulation of the secondary metabolism biosynthetic pathways [134].
The HRs of pigeon pea induced by infection with R. rhizogenes strain K599 showed good biomass increase (17.3 g DW/L was obtained in 18 days of culture). The roots were elicited using the method described earlier for peanut HRs [194]. In 12-day-old cultures, the standard nutrient medium was replaced by the elicitation medium containing 125 µM MeJA, 18 g/L methyl-β-cyclodextrin, 3 mM H2O2 and 1 mM MgCl2 and the HRs were cultivated in the dark for another 7 days. After 144 h of treatment, the total content of cajaninstilbene acid in the culture reached over 8 mg/g DW. Almost 96% of the compound was secreted into the medium; thus, 7.7 mg/g DW of the product was extracted from the medium and the remaining 0.35 mg/g DW originated from the biomass. In the control roots, content of cajaninstilbene acid was 0.03 mg/g DW and the product could not be detected in the extract from the nutrient medium [210].
2.3. Hydrolyzable Tannins
Plants that accumulate considerable amounts of hydrolysable tannins are traditionally used as anti-diarrhea medicines. They are also used topically as hemostatic, antiphlogistic and astringent agents. Hydrolyzable tannins (for structures, see Figure 8) are components of numerous food plants including the strawberry, raspberry, pomegranate and walnuts, but their bioavailability is poor. When ingested, hydrolyzable tannins are metabolized by gut microbiota to urolithins that are much better absorbed and are found in plasma mostly as glucuronides. The tannin metabolites have gained much interest as potential anticancer agents and estrogen receptor modulators of potential use in cardiovascular disorders, osteoporosis and several hormone-dependent diseases [211,212,213]. Hydrolyzable tannins production in the HRs has been studied chiefly to elucidate the regulation of the biosynthesis of these compounds.
Sanguisorba officinalis L. (Rosaceae) has been traditionally used as hemostatic and wound healing medicine. The HRs of the plant were obtained by inoculation with R. rhizogenes strain A4. Five phenolic compounds were identified in the investigated roots: gallic acid, 1,2,3,6-tetra-O-galloyl-β-D-glucose, 1,2,3,4,6-penta-O-galloyl-β-D-glucose, sanguiin H-6 and sanguiin H-11. Five lines of the studied HRs preferably synthesized sanguiin H-6 (0.217–0.569% FW) and one, showing the fastest growth, accumulated mainly 1,2,3,6-tetra-O-galloyl-β-D-glucose (0.322% FW) and sanguiin H-11 (0.221% FW). An intact plant contained 0.206% FW sanguiin H-6. The contents of hydrolysable tannins in the HRs were simililar to those found in adventitious roots; however, the biomass of HRs obtained in one four-week growth cycle was up to six times larger than that produced by adventitious roots [214].
The HRs of Geranium thunbergii Siebold ex Lindl. & Paxt. (Geraniaceae), transformed with R. rhizogenes strain A4 synthesized: gallic acid, ellagic acid, (+)-catechin, β-glucogallin (1-O-galloyl-β-D-glucopyranoside), 1,6-di-O-galloyl-β-D-glucose, 1,2,3,6-tetra-O-galloyl-β-D-glucose, 1,2,3,4,6-penta-O-galloyl-β-D-glucose, corilagin and geraniin. The HRs grown in ½ MS medium accumulated mainly 1,2,3,4,6-penta-O-galloyl-β-D-glucose whereas Gamborg’s B5 medium favored biosynthesis of geraniin [215]. Phyllanthus niruri L. (Phyllanthaceae) partly owes its medicinal properties to a high content of corilagin. The HRs of P. niruri produced neither corilagin nor geraniin but yielded several phenolic metabolites including gallic acid, (−)-epicatechin 3-O-gallate, (+)-gallocatechin and (−)-epigallocatechin 3-O-gallate [216]. Fragaria ananassa cv. Reikou infected by R. rhizogenes strain ATCC 15834 developed hairy roots that produced pedunculagin (2,3,4,6-tetra-O-galloyl-β-D-glucose). The tannin was present in the HRs grown in MS medium in the initial phase of growth (0.6% DW) and then its content declined (0.4% in 3-week culture). After 5 weeks of culture pedunculagin was not detected in the HRs. Two other nutrient media used in the study were less favorable for both biomass and pedunculagin production [35].
Lawsonia inermis L. (Lythraceae) is a source of henna, a pigment used to dye hair, skin and fingernails. A naphthoquinone, lawsone, is the compound responsible for dyeing properties as well as for bacteriostatic activity. Except for lawsone, gallic acid, flavonoids, coumarins and xanthones are known metabolites of the plant. The HRs of L. inermis were derived from the explants infected with R. rhizogenes strain NCIB 8196. The optimum growth of roots was achieved using MS medium. The HRs grown in the dark accumulated 1,2,3,6-tetra-O-galloyl-β-D-glucose (0.43% DW), 1,2,3,4,6-penta-O-galloyl-β-D-glucose (3.04% DW) and (+)-catechin. Normal roots contained up to 0.5% galloylglucoses, but up to 60 times more (+)-catechin [217].
Punica granatum L. (Lythraceae) cv. Wonderful was used to establish pomegranate HR culture. Three wild-type R. rhizogenes strains: MSU440, ATCC 15834 and A4, were transformed with a binary vector containing YFP (yellow fluorescent protein). The modified bacterial strains were used to initiate HRs. Punicalagin α and β were produced by the HRs with better efficiency than that achieved by in vitro cultured normal roots. The roots of the wild-type seedlings, however, contained more punicalagins.
The glycosylation of gallic acid to β-glucogallin is the first step in punicalagin biosynthesis. The reaction is catalyzed by a specific UDP-glycosyltransferase (UGT). Transcriptomic analysis of pomegranate fruit peel (rich in tannins) allowed for the identification of 32 putative UGTs. By comparison with the results of transcript level analyses made for other tissues, two candidate genes were identified (PgUGT1 and PgUGT13) that correlated with the tissue-specific abundance of tannins. To verify whether the two genes as well as the genes for the other UGTs are expressed in the HRs, RT-PCR was performed using RNA extracted from fruit peel, wild type roots and HRs. Expression of 26 putative UGTs was confirmed in all of the investigated tissue types [218]. Four candidate UGTs from pomegranate were cloned and biochemically characterized of which only two (UGT84A23 and UGT84A24) catalyzed formation of β-glucogallin. The overexpression or knockdown of the gene encoding one of the two UGTs in the HRs did not cause any obvious alterations in punicalagin accumulation. Double-knockdown HR lines accumulated less punicalagin but started to synthesize galloyl glucosides (ether-linked gallic acid and glucose). The new metabolites of the double-knockdown HR lines were gallic acid 3-O-glucopyranoside, gallic acid 4-O-glucopyranoside and digalloyl glucose conjugate. The results suggested that in the HRs, gallic acid may be utilized by an unknown UGTs for glucoside formation [219]. In a search for the UGTs engaged in the synthesis of gallic acid glycosides 11 candidate UGTs were identified by the comparison of UGTs expression in control root lines and double knockdown lines of pomegranate HRs. Of the 11 candidate UGTs, only one was active toward gallic acid and catalyzed the formation of a single product: gallic acid 4-O-glucoside [220].
3. Conclusions
Table 1 summarizes the most spectacular achievements of the research on highly productive hairy root cultures or genetically modified plants containing polyphenolic antioxidants from the flavonoid, stilbenoid and hydrolyzable tannin group. The high yields of the polyphenolic products were obtained using procedures like optimization of culture conditions, selection of the plant material and bacterial strain used to initiate the HRs, elicitation, permeabilization of plant tissue, adding the enzyme cofactors and genetic engineering (overexpression, heterologous expression or silencing of genes-encoding the regulatory transcription factors and biosynthetic enzymes). It should be considered, however, that high contents of polyphenols in the HRs may be not accompanied by the vigorous root growth. This can hamper applications of the cultures that accumulate products in the cells.
HR cultures provide a valuable tool for the rapid characterization of regulatory mechanisms in plant polyphenol biosynthesis, and elucidation of the roles played by the particular biosynthetic enzymes or transcription factors. The use of HRs has led to the identification and description of several enzymes implicated in polyphenol biosynthesis. Nowadays, it seems that HR culture is primarily a useful tool in the genetic engineering of plants. The implementation of the industrial process using HRs to produce antioxidant natural products looks like a distant prospect.
Author Contributions
Conceptualization, A.S.; methodology, A.S and J.M.; resources, A.S., J.M. and Y.V.Y.; data curation, A.S.; writing—original draft preparation, A.S. and Y.V.Y.; writing—review and editing, A.S., J.M., K.M. and Y.V.Y.; visualization, K.M.; supervision, A.S.; project administration, A.S.; funding acquisition, A.S. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
| 4CL | 4-coumarate:CoA ligase (4-coumaroyl CoA ligase) |
| ANR | anthocyanidin reductase |
| ANS | anthocyanidin synthase |
| BAP | 6-benzylaminopurine |
| bHLH | basic helix-loop-helix |
| BHT | butylohydroxytoluen |
| C4H | cinnamate 4-hydroxylase |
| CHI | chalcone isomerase |
| CHS | chalcone synthase |
| DFR | dihydroflavonol 4-reductase |
| DW | dry weight |
| F3H | flavone 3-hydroxylase |
| F3βOH | flavanone 3-β-hydroxylase |
| F6H | flavone 6-hydroxylase |
| F8H | flavone 8-hydroxylase |
| FNS | flavone synthase |
| FW | fresh weight |
| GUS | β-glucuronidase |
| HPLC-PAD-ESI-MSn | high-performance liquid chromatography with photodiode detection, electrospray ionization, and multi-stage mass spectrometry |
| HR | hairy roots |
| IBA | indole-3-butyric acid |
| IF7Mat | isoflavone 7-O-glucoside-6”-O-malonyltransferase |
| IFD | isoflavone dehydratase |
| IFS | isoflavone synthase |
| IOMT | isoflavone-O-methyltransferase |
| LAR | leucoanthocyanidin reductase |
| LC-MS/MS | liquid chromatography with tandem mass spectrometry |
| MaT | malonyl CoA:flavonoid acyltransferase |
| MeJa | methyl jasmonate |
| MS | Murashige and Skoog |
| PA | proanthocyanidin |
| PAL | phenylalanine ammonia lyase |
| PAP1 | production of anthocyanin pigment 1 gene |
| PQ | paraquat |
| RS | resveratrol synthase |
| SA | salicylic acid |
| SH | Schenk and Hildebrandt |
| STS | stilbene synthase |
| TOF-MS | time-of-flight mass spectroscopy |
| TT2 | transparent testa 2 transcription factor |
| UBGT | UDP-glucose:baicalein 7-O-glucosyltransferase |
| UGT | UDP-glucosyltransferase |
| UHPLC-PDA | ultra-high-performance liquid chromatography with photodiode detection |
| YE | yeast extract |
References
- Hamill, J.D.; Lidgett, A.J. Hairy root cultures–opportunities and key protocols for studies in metabolic engineering. In Hairy Roots. Culture and Applications, 1st ed.; Doran, P., Ed.; Harwood Academic Publishers: Amsterdam, The Netherlands, 1997; pp. 1–29. [Google Scholar]
- Giri, A.; Narasu, M.L. Transgenic hairy roots: Recent trends and applications. Biotechnol. Adv. 2000, 18, 1–22. [Google Scholar] [CrossRef]
- Guillon, S.; Trémouillaux-Guiller, J.; Pati, P.K.; Rideau, M.; Gantet, P. Harnessing the potential of hairy roots: Dawn of a new era. Trends Biotechnol. 2006, 24, 403–409. [Google Scholar] [CrossRef] [PubMed]
- Ono, N.N.; Tian, L. The multiplicity of hairy root cultures: Prolific possibilities. Plant Sci. 2011, 180, 439–446. [Google Scholar] [CrossRef]
- Gutierrez-Valdes, N.; Häkkinen, S.T.; Lemasson, C.; Guillet, M.; Oksman-Caldentey, K.M.; Ritala, A.; Cardon, F. Hairy root cultures–a versatile tool with multiple applications. Front. Plant Sci. 2020, 11, 33. [Google Scholar] [CrossRef]
- Shi, M.; Liao, P.; Nile, S.H.; Georgiev, M.I.; Kai, G. Biotechnological exploration of transformed root culture for value-added products. Trends Biotechnol. 2021, 39, 137–149. [Google Scholar] [CrossRef] [PubMed]
- Young, J.M.; Kuykendall, L.D.; Martínez-Romero, E.; Kerr, A.; Sawada, H. A revision of Rhizobium Frank 1889, with an emended description of the genus, and the inclusion of all species of Agrobacterium Conn 1942 and Allorhizobium undicola de Lajudie et al. 1998 as new combinations: Rhizobium radiobacter, R. rhizogenes, R. rubi, R. undicola and R. vitis. Int. J. Syst. Evol. Microbiol. 2001, 51, 89–103. [Google Scholar] [CrossRef]
- Hidalgo, D.; Georgiev, M.; Marchev, A.; Bru-Martinez, R.; Cusido, R.; Corchete, P.; Palazon, J. Tailoring tobacco hairy root metabolism for the production of stilbenes. Sci. Rep. 2017, 7, 17976. [Google Scholar] [CrossRef]
- Dong, Q.; Zhao, H.; Huang, Y.; Chen, Y.; Wan, M.; Zeng, Z.; Yao, P.; Li, C.; Wang, X.; Chen, H.; et al. FtMYB18 acts as a negative regulator of anthocyanin/proanthocyanidin biosynthesis in Tartary buckwheat. Plant Mol. Biol. 2020, 104, 309–325. [Google Scholar] [CrossRef]
- Sosa Alderete, L.G.; Talano, M.A.; Ibáñez, S.G.; Purro, S.; Agostini, E.; Milrad, S.R.; Medina, M.I. Establishment of transgenic tobacco hairy roots expressing basic peroxidases and its application for phenol removal. J. Biotechnol. 2009, 139, 273–279. [Google Scholar] [CrossRef]
- Ibáñez, S.; Talano, M.; Ontañon, O.; Suman, J.; Medina, M.I.; Macek, T.; Agostini, E. Transgenic plants and hairy roots: Exploiting the potential of plant species to remediate contaminants. New Biotechnol. 2016, 33, 625–635. [Google Scholar] [CrossRef]
- Jha, P.; Sen, R.; Jobby, R.; Sachar, S.; Bhatkalkar, S.; Desai, N. Biotransformation of xenobiotics by hairy roots. Phytochemistry 2020, 176, 112421. [Google Scholar] [CrossRef] [PubMed]
- Gantait, S.; Mukherjee, E. Hairy root culture technology: Applications, constraints and prospect. Appl. Microbiol. Biotechnol. 2021, 105, 35–53. [Google Scholar] [CrossRef] [PubMed]
- Lystvan, K.; Lystvan, V.; Shcherbak, N.; Kuchuk, M. Rhizoextraction potential of Convolvulus tricolor hairy roots for Cr6+, Ni2+, and Pb2+ removal from aqueous solutions. Appl. Biochem. Biotechnol. 2021, 193, 1215–1230. [Google Scholar] [CrossRef]
- Srikantan, C.; Srivastava, S. Bioreactor design and analysis for large-scale plant cell and hairy root cultivation. In Hairy Roots An Effective Tool of Plant Biotechnology, 1st ed.; Srivastava, V., Mehrotra, S., Mishra, S., Eds.; Springer Nature Singapore PteLtd.: Singapore, 2018; pp. 147–182. [Google Scholar]
- Hussain, M.J.; Abbas, Y.; Nazli, N.; Fatima, S.; Drouet, S.; Hano, C.; Abbasi, B.H. Root cultures, a boon for the production of valuable compounds: A comparative review. Plants 2022, 11, 439. [Google Scholar] [CrossRef] [PubMed]
- Macáková, K.; Afonso, R.; Saso, L.; Mladĕnka, P. The influence of alkaloids on oxidative stress and related signaling pathways. Free Radic. Biol. Med. 2019, 134, 429–444. [Google Scholar] [CrossRef] [PubMed]
- Williamson, G.; Faulkner, K.; Plumb, G.W. Glucosinolates and phenolics as antioxidants from plant foods. Eur. J. Cancer Prev. 1998, 7, 17–21. [Google Scholar]
- Cabello-Hurtado, F.; Gicquel, M.; Esnault, M.-A. Evaluation of the antioxidant potential of cauliflower (Brassica oleracea) from a glucosinolate content perspective. Food Chem. 2012, 132, 1003–1009. [Google Scholar] [CrossRef]
- Wang, X.; Liu, R.; Yang, Y.; Zhang, M. Isolation, purification and identification of antioxidants in an aqueous aged garlic extract. Food Chem. 2015, 187, 37–43. [Google Scholar] [CrossRef]
- Brewer, M.S. Natural antioxidants: Sources, compounds, mechanisms of action, and potential applications. Compr. Rev. Food Sci. Food Saf. 2011, 10, 221–247. [Google Scholar] [CrossRef]
- Krishnajah, D.; Sarbatly, R.; Nithyanandam, R. A review of the antioxidant potential of medicinal plant species. Food Bioprod Process. 2011, 89, 217–233. [Google Scholar] [CrossRef]
- Wink, M.; Ashour, M.L.; El-Readi, M.Z. Secondary metabolites from plants inhibiting ABC transporters and reversing resistance of cancer cells and microbes to cytotoxic and antimicrobial agents. Front. Microbiol. 2012, 3, 130. [Google Scholar] [CrossRef] [Green Version]
- Choudhari, A.S.; Mandave, P.C.; Deshpande, M.; Ranjekar, P.; Prakash, O. Phytochemicals in cancer treatment: From preclinical studies to clinical practice. Front. Pharmacol. 2020, 10, 1614. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cui, X.; Lin, Q.; Liang, Y. Plant-derived antioxidants protect the nervous system from aging by inhibiting oxidative stress. Front. Aging Neurosci. 2020, 12, 209. [Google Scholar] [CrossRef] [PubMed]
- Duba, K.S.; Casazza, A.A.; Mohamed, H.B.; Perego, P.; Fiori, L. Extraction of polyphenols from grape skins and defatted grape seeds using subcritical water: Experiments and modeling. Food Bioprod. Process. 2015, 94, 29–38. [Google Scholar] [CrossRef]
- Sorrenti, V.; Randazzo, C.L.; Caggia, C.; Ballistreri, G.; Romeo, F.V.; Fabroni, S.; Timpanaro, N.; Raffaele, M.; Vanella, L. Beneficial effects of pomegranate peel extract and probiotics on pre-adipocyte differentiation. Front. Microbiol. 2019, 10, 660. [Google Scholar] [CrossRef]
- Cova, C.M.; Boffa, L.; Pistocchi, M.; Giorgini, S.; Luque, R.; Cravotto, G. Technology and process design for phenols recovery from industrial chicory (Chicorium intybus) leftovers. Molecules 2019, 24, 2681. [Google Scholar] [CrossRef] [Green Version]
- Khalili, F.; Dinani, S.T. Extraction of phenolic compounds from olive-waste cake using ultrasonic process. J. Food Meas. Charact. 2018, 12, 974–981. [Google Scholar] [CrossRef]
- Morris, P.; Robbins, M.P. Condensed tannin formation by Agrobacterium rhizogenes transformed root and shoot organ cultures of Lotus corniculatus. J. Exp. Bot. 1992, 43, 221–231. [Google Scholar] [CrossRef]
- Hook, I. Secondary metabolites in hairy root cultures of Leontopodium alpinum Cass. (Edelweiss). Plant Cell Tiss. Org. Cult. 1994, 38, 321–326. [Google Scholar] [CrossRef]
- Park, C.H.; Thwe, A.A.; Kim, S.J.; Park, J.S.; Arasu, M.V.; Al-Dhabi, N.A.; Park, N.I.; Park, S.U. Effect of auxins on anthocyanin accumulation in hairy root cultures of Tartary buckwheat cultivar Hokkai T10. Nat. Prod. Commun. 2016, 11, 1283–1286. [Google Scholar] [CrossRef] [Green Version]
- Nishiyama, Y.; Yamakawa, T. Effect of medium composition on the production of anthocyanins by hairy root cultures of Ipomea batatas. Plant Biotechnol. 2004, 21, 411–414. [Google Scholar] [CrossRef] [Green Version]
- Thwe, A.A.; Kim, Y.; Li, X.; Kim, Y.B.; Park, N.I.; Kim, H.H.; Kim, S.-J.; Park, S.U. Accumulation of phenylpropanoids and correlated gene expression in hairy roots of Tartary buckwheat under light and dark conditions. Appl. Biochem. Biotechnol. 2014, 174, 2537–2547. [Google Scholar] [CrossRef] [PubMed]
- Motomori, Y.; Shimomura, K.; Mori, K.; Kunitake, H.; Nakashima, T.; Tanaka, M.; Miyazaki, S.; Ishimaru, K. Polyphenol production in hairy root cultures of Fragaria x ananassa. Phytochemistry 1995, 40, 1425–1428. [Google Scholar] [CrossRef]
- Bavage, A.D.; Davies, I.G.; Robbins, M.P.; Morris, P. Expression of an Antirrhinum dihydroflavonol reductase gene results in changes in condensed tannin structure and accumulation in root cultures of Lotus corniculatus (bird’s foot trefoil). Plant Mol. Biol. 1997, 35, 443–458. [Google Scholar] [CrossRef]
- Damiani, F.; Paolocci, F.; Consonni, G.; Crea, F.; Tonelli, C.; Arcioni, S. A maize anthocyanin transactivator induces pigmentation in hairy roots of dicotyledonous species. Plant Cell Rep. 1998, 17, 339–344. [Google Scholar] [CrossRef]
- Pang, Y.; Peel, G.J.; Sharma, S.B.; Tang, Y.; Dixon, R.A. A transcript profiling approach reveals an epicatechin-specific glucosyltransferase expressed in the seed coat of Medicago truncatula. Proc. Natl. Acad. Sci. USA 2008, 105, 14210–14215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Terrier, N.; Torregrosa, L.; Ageorges, A.; Vialet, S.; Verriès, C.; Cheynier, V.; Romieu, C. Ectopic expression of VvMybPA2 promotes proanthocyanidin biosynthesis in grapevine and suggests additional targets in the pathway. Plant Physiol. 2009, 149, 1028–1041. [Google Scholar] [CrossRef] [Green Version]
- Verdier, J.; Zhao, J.; Torres-Jerez, I.; Ge, S.; Liu, C.; He, X.; Mysore, K.S.; Dixon, R.A.; Udvardi, M.K. MtPAR MYB transcription factor acts as an on switch for proanthocyanidin biosynthesis in Medicago truncatula. Proc. Natl. Acad. Sci. USA 2012, 109, 1766–1771. [Google Scholar] [CrossRef] [Green Version]
- Huang, Y.-F.; Vialet, S.; Guiraud, J.-L.; Torregrosa, L.; Bertrand, Y.; Cheynier, V.; This, P.; Terrier, N. A negative MYB regulator of proanthocyanidin accumulation, identified through expression quantitative locus mapping in the grape berry. New Phytol. 2014, 201, 795–809. [Google Scholar] [CrossRef]
- Cavallini, E.; Matus, J.T.; Finezzo, L.; Zenoni, S.; Loyola, R.; Guzzo, F.; Schlechter, R.; Ageorges, A.; Arce-Johnson, P.; Tornielli, G.B. The phenylpropanoid pathway is controlled at different branches by a set of R2R3-MYB C2 repressors in grapevine. Plant Physiol. 2015, 167, 1448–1470. [Google Scholar] [CrossRef]
- Jun, J.H.; Liu, C.; Xiao, X.; Dixon, R.A. The transcriptional repressor MYB2 regulates both spatial and temporal patterns of proanthocyanidin and anthocyanin pigmentation in Medicago truncatula. Plant Cell 2015, 27, 2860–2879. [Google Scholar] [PubMed] [Green Version]
- Li, P.; Chen, B.; Zhang, G.; Chen, L.; Dong, Q.; Wen, J.; Mysore, K.S.; Zhao, J. Regulation of anthocyanin and proanthocyanidin biosynthesis by Medicago truncatula bHLH transcription factor MtTT8. New Phytol. 2016, 210, 905–921. [Google Scholar] [CrossRef] [PubMed]
- Lu, N.; Roldan, M.; Dixon, R.A. Characterization of two TT2-type MYB transcription factors regulating proanthocyanidin biosynthesis in tetraploid cotton, Gossypium hirsutum. Planta 2017, 246, 323–335. [Google Scholar] [CrossRef] [PubMed]
- Lu, N.; Rao, X.; Li, Y.; Jun, J.H.; Dixon, R.A. Dissecting the transcriptional regulation of proanthocyanidin and anthocyanin biosynthesis in soybean (Glycine max). Plant Biotechnol. J. 2021, 19, 1429–1442. [Google Scholar] [CrossRef]
- Aggarwal, P.R.; Nag, P.; Choudhary, P.; Chakraborty, N.; Chakraborty, S. Genotype-independent Agrobacterium rhizogenes-mediated root transformation of chickpea: A rapid and efficient method for reverse genetics studies. Plant Methods 2018, 14, 55. [Google Scholar] [CrossRef] [Green Version]
- Li, P.; Dong, Q.; Ge, S.; He, X.; Verdier, J.; Li, D.; Zhao, J. Metabolic engineering of proanthocyanidin production by repressing the isoflavone pathways and redirecting anthocyanidin precursor flux in legume. Plant Biotechnol. J. 2016, 14, 1604–1618. [Google Scholar] [CrossRef] [Green Version]
- Qin, Y.; Wang, D.; Fu, J.; Zhang, Z.; Qin, Y.; Hu, G.; Zhao, J. Agrobacterium rhizogenes-mediated hairy root transformation as an efficient system for gene function analysis in Litchi chinensis. Plant Methods 2021, 17, 103. [Google Scholar] [CrossRef]
- Xu, S.; Lai, E.; Zhao, L.; Cai, Y.; Ogutu, C.; Cherono, S.; Han, Y.; Zheng, B. Development of a fast and efficient root transgenic system for functional genomics and genetic engineering in peach. Sci. Rep. 2020, 10, 2836. [Google Scholar] [CrossRef] [Green Version]
- Yoshida, K.; Ma, D.; Constabel, C.P. The MYB182 protein down-regulates proanthocyanidin biosynthesis in poplar by repressing both structural and regulatory flavonoid genes. Plant Physiol. 2015, 167, 693–710. [Google Scholar] [CrossRef] [Green Version]
- Pang, Y.; Abeysinghe, I.S.B.; He, J.; He, X.; Huhman, D.; Mewan, K.M.; Sumner, L.W.; Yun, J.; Dixon, R.A. Functional characterization of proanthocyanidin pathway enzymes from tea and their application for metabolic engineering. Plant Physiol. 2013, 161, 1103–1116. [Google Scholar] [CrossRef] [Green Version]
- Thwe, A.; Arasu, M.V.; Li, X.; Park, C.H.; Kim, S.J.; Al-Dhabi, N.A.; Park, S.U. Effect of different Agrobacterium rhizogenes strains on hairy root induction and phenylpropanoid biosynthesis in Tartary buckwheat (Fagopyrum tataricum Gaertn). Front. Microbiol. 2016, 7, 318. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Thwe, A.A.; Park, C.H.; Kim, S.J.; Arasu, M.V.; Al-Dhabi, N.A.; Lee, S.Y.; Park, S.U. Ethephon-induced phenylpropanoid accumulation and related gene expression in tartary buckwheat (Fagopyrum tataricum (L.) Gaertn.) hairy root. Biotechnol. Biotechnol. Equip. 2017, 31, 304–311. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Sathasivam, R.; Park, N.I.; Wu, Q.; Park, S.U. Enhancement of phenylpropanoid accumulation in Tartary buckwheat hairy roots by overexpression of MYB transcription factors. Ind. Crop. Prod. 2020, 156, 112887. [Google Scholar] [CrossRef]
- Barba-Espin, G.; Chen, S.-T.; Agnolet, S.; Nymark Hegelund, J.; Stanstrup, J.; Christensen, J.H.; Müller, R.; Lütken, H. Ethephon-induced changes in antioxidants and phenolic compounds in anthocyanin-producing black carrot hairy root cultures. J. Exp. Bot. 2020, 71, 7030–7045. [Google Scholar] [CrossRef] [PubMed]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bioassays with tobacco tissue cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Barba-Espin, G.; Martínez-Jiménez, C.; Izquierdo-Martínez, A.; Acosta-Motos, J.R.; Hernández, J.A.; Díaz-Vivancos, P. H2O2-elicitation of black carrot hairy roots induces a controlled oxidative burst leading to increased anthocyanin production. Plants 2021, 10, 2753. [Google Scholar] [CrossRef]
- Piao, C.; Wu, J.; Cui, M.-L. The combination of R2R3-MYB gene AmRosea1 and hairy root culture is a useful tool for rapidly induction and production of anthocyanins in Antirrhinum majus L. AMB Expr. 2021, 11, 128. [Google Scholar] [CrossRef]
- Jin, S.; Hyun, T.K. Ectopic expression of Production of Anthocyanin Pigment 1 (PAP1) improves the antioxidant and anti-melanogenic properties of ginseng (Panax ginseng C.A. Meyer) hairy roots. Antioxidants 2020, 9, 922. [Google Scholar] [CrossRef]
- Jin, S.; Bang, S.; Ahn, M.-A.; Lee, K.; Kim, K.; Hyun, T.K. Overproduction of anthocyanin in ginseng hairy roots enhances their antioxidant, antimicrobial, and anti-elastase activities. J. Plant Biotechnol. 2021, 48, 100–105. [Google Scholar] [CrossRef]
- Trotin, F.; Moumou, Y.; Vasseur, J. Flavanol production by Fagopyrum esculentum hairy and normal root cultures. Phytochemistry 1993, 32, 929–931. [Google Scholar] [CrossRef]
- Kim, Y.K.; Xu, H.; Park, W.T.; Park, N.I.; Lee, S.Y.; Park, S.U. Genetic transformation of buckwheat (Fagopyrum esculentum M.) with Agrobacterium rhizogenes and production of rutin in transformed root cultures. Aust. J. Crop Sci. 2010, 4, 485–490. [Google Scholar]
- Park, N.I.; Li, X.; Thwe, A.A.; Lee, S.Y.; Kim, S.G.; Wu, Q.; Park, S.U. Enhancement of rutin in Fagopyrum esculentum hairy root cultures by the Arabidopsis transcription factor AtMYB12. Biotechnol. Lett. 2012, 34, 577–583. [Google Scholar] [CrossRef] [PubMed]
- Yao, P.; Huang, Y.; Dong, Q.; Wan, M.; Wang, A.; Chen, Y.; Li, C.; Wu, Q.; Chen, H.; Zhao, H. FtMYB6, a light-induced SG7 R2R3-MYB transcription factor, promotes flavonol biosynthesis in Tartary buckwheat (Fagopyrum tataricum). J. Food Agricul. Chem. 2020, 68, 13685–13696. [Google Scholar] [CrossRef]
- Yin, Q.; Han, X.; Han, Z.; Chen, Q.; Shi, Y.; Gao, H.; Zhang, T.; Dong, G.; Xiong, C.; Song, C.; et al. Genome-wide analyses reveals a glucosyltransferase involved in rutin and emodin glucoside biosynthesis in tartary buckwheat. Food Chem. 2020, 318, 126478. [Google Scholar] [CrossRef] [PubMed]
- Choi, M.; Sathasivam, R.; Nguyen, B.V.; Park, N.I.; Woo, S.-H.; Park, S.U. Expression analysis of phenylpropanoid pathway genes and metabolomic analysis of phenylpropanoid compounds in adventitious, hairy, and seedling roots of Tartary buckwheat. Plants 2021, 11, 90. [Google Scholar] [CrossRef]
- Tusevski, O.; Petreska Stanoeva, J.; Stefova, M.; Kungulovski, D.; Pancevska, N.A.; Sekulovski, N.; Panov, S.; Gadzovska Simic, S. Hairy roots of Hypericum perforatum L.: A promising system for xanthone production. Cent. Eur. J. Biol. 2013, 8, 1010–1022. [Google Scholar] [CrossRef]
- Tusevski, O.; Petreska Stanoeva, J.; Stefova, M.; Gadzovska Simic, S. Phenolic profile of dark-grown and photoperiod-exposed Hypericum perforatum L hairy root cultures. Sci. World J. 2013, 602752. [Google Scholar] [CrossRef]
- Tusevski, O.; Vinterhalter, B.; Kristić Milošević, D.; Soković, M.; Ćirić, A.; Vinterhalter, D.; Zdravković Korać, S.; Petreska Stanoeva, J.; Gadzovska Simic, S. Production of phenolic compounds, antioxidant and antimicrobial activities in hairy root and shoot cultures of Hypericum perforatum L. Plant Cell Tissue Organ Cult. 2017, 128, 589–605. [Google Scholar] [CrossRef]
- Tashackori, H.; Sharifi, M.; Chashmi, N.A.; Safaie, N.; Behmanesh, M. Induced-differential changes on lignan and phenolic acid compounds in Linum album hairy roots by fungal extract of Piriformospora indica. Plant Cell Tissue Organ Cult. 2016, 127, 187–194. [Google Scholar] [CrossRef]
- Tashackori, H.; Sharifi, M.; Chashmi, N.A.; Behmanesh, M.; Safaie, N. Piriformospora indica cell wall modulates gene expression and metabolite profile in Linum album hairy roots. Planta 2018, 248, 1289–1306. [Google Scholar] [CrossRef]
- Tashackori, H.; Sharifi, M.; Chashmi, N.A.; Behmanesh, M.; Safaie, N.; Sagharyan, M. Physiological, biochemical, and molecular responses of Linum album to digested cell wall of Piriformospora indica. Physiol. Mol. Biol. Plants 2021, 27, 2695–2708. [Google Scholar] [CrossRef]
- Thiruvengadam, M.; Praveen, N.; Kim, E.-H.; Kim, S.-H.; Chung, I.-M. Production of antraquinones, phenolic compounds and biological activities from hairy root cultures of Polygonum multiflorum Thunb. Protoplasma 2014, 251, 555–566. [Google Scholar] [CrossRef] [PubMed]
- Ho, T.-T.; Lee, J.-D.; Ahn, M.-S.; Kim, S.-W.; Park, S.-Y. Enhanced production of phenolic compounds in hairy root cultures of Polygonum multiflorum and its metabolite discrimination using HPLC and FT-IR methods. Appl. Microbiol. Biotechnol. 2018, 102, 9563–9575. [Google Scholar] [CrossRef]
- Thiruvengadam, M.; Praveen, N.; John, K.M.M.; Yang, Y.-S.; Kim, S.-H.; Chung, I.-M. Establishment of Momordica charantia hairy root cultures for the production of phenolic compounds and determination of their biological activities. Plant Cell Tissue Organ Cult. 2014, 118, 545–557. [Google Scholar] [CrossRef]
- Thiruvengadam, M.; Rekha, K.; Chung, I.-M. Induction of hairy roots by Agrobacterium rhizogenes-mediated transformation of spine gourd (Momordica dioica Roxb. ex. Willd) for the assessment of phenolic compounds and biological activities. Sci. Hortic. 2016, 198, 132–141. [Google Scholar] [CrossRef]
- Yoon, J.-Y.; Chung, I.-M.; Thiruvengadam, M. Evaluation of phenolic compounds, antioxidant and antimicrobial activities from transgenic hairy root cultures of gherkin (Cucumis anguria L.). S. Afr. J. Bot. 2015, 100, 80–86. [Google Scholar] [CrossRef]
- Chung, I.-M.; Rekha, K.; Rajakumar, G.; Thiruvengadam, M. Production of glucosinolates, phenolic compounds and associated gene expression profiles of hairy root cultures in turnip (Brassica rapa ssp. rapa). 3 Biotech 2016, 6, 175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chung, I.-M.; Rekha, K.; Rajakumar, G.; Thiruvengadam, M. Production of bioactive compounds and gene expression alterations in hairy root cultures of chinese cabbage elicited by copper oxide nanoparticles. Plant Cell Tissue Organ Cult. 2018, 134, 95–106. [Google Scholar] [CrossRef]
- Chung, I.-M.; Rekha, K.; Rajakumar, G.; Thiruvengadam, M. Influence of silver nanoparticles on the enhancement and transcriptional changes of glucosinolates and phenolic compounds in genetically transformed root cultures of Brassica rapa ssp. rapa. Bioprocess Biosyst. Eng. 2018, 41, 1665–1677. [Google Scholar] [CrossRef]
- Gai, Q.-Y.; Jiao, J.; Luo, M.; Wei, Z.-F.; Zu, Y.-G.; Ma, W.; Fu, Y.-J. Establishment of hairy root cultures by Agrobacterium rhizogenes mediated transformation of Isatis tinctoria L. for the efficient production of flavonoids and evaluation of antioxidant activities. PLoS ONE 2015, 10, e0119022. [Google Scholar] [CrossRef]
- Jiao, J.; Gai, Q.-Y.; Wang, X.; Qin, Q.-P.; Wang, Z.-Y.; Liu, J.; Fu, Y.-J. Chitosan elicitation of Isatis tinctoria L. hairy root cultures for enhancing flavonoid productivity and gene expression and related antioxidant activity. Ind. Crops Prod. 2018, 124, 28–35. [Google Scholar] [CrossRef] [PubMed]
- Jiao, J.; Gai, Q.-Y.; Wang, W.; Zang, Y.-P.; Niu, L.-L.; Fu, Y.-J.; Wang, X. Remarkable enhancement of flavonoid production in a co-cultivation system of Isatis tinctoria L. hairy root cultures and immobilized Aspergillus niger. Ind. Crops Prod. 2018, 112, 252–261. [Google Scholar] [CrossRef] [PubMed]
- Jiao, J.; Gai, Q.-Y.; Yao, L.-P.; Niu, L.-L.; Zang, Y.-P.; Fu, Y.-J. Ultraviolet radiation for flavonoid augmentation in Isatis tinctoria L. hairy root cultures mediated by oxidative stress and biosynthetic gene expression. Ind. Crops Prod. 2018, 118, 347–354. [Google Scholar] [CrossRef]
- Ghimire, B.K.; Thiruvengadam, M.; Chung, I.-M. Identification of elicitors enhances the polyphenolic compounds and pharmacological potential in hairy root cultures of Aster scaber. S. Afr. J. Bot. 2019, 125, 92–101. [Google Scholar] [CrossRef]
- Ansari, M.A.; Chung, I.-M.; Rajakumar, G.; Alzohairy, M.A.; Almatroudi, A.; Khanna, V.G.; Thiruvengadam, M. Evaluation of polyphenolic compounds and pharmacological activities in hairy root cultures of Ligularia fischeri Turcz. f. spiciformis (Nakai). Molecules 2019, 24, 1586. [Google Scholar] [CrossRef] [Green Version]
- Chung, I.-M.; Ahmad, A.; Ali, M.; Lee, O.-K.; Kim, M.-Y.; Kim, J.-H.; Yoon, D.-Y.; Peebles, C.A.M.; San, K.Y. Flavonoid glucosides from the hairy roots of Catharanthus roseus. J. Nat. Prod. 2009, 72, 613–620. [Google Scholar] [CrossRef] [PubMed]
- Tomimori, T.; Jin, H.; Miyaichi, Y.; Toyofuku, S.; Namba, T. Studies on the constituents of Scutellaria species. VI. On the flavonoid constituents of the root of Scutellaria baicalensis Georgi (5). Quantitative analysis of flavonoids in Scutellaria roots by high-performance liquid chromatography. Yakugaku Zasshi 1985, 105, 148–155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shang, X.; He, X.; He, X.; Li, M.; Zhang, R.; Fan, P.; Zhang, Q.; Jia, Z. The genus Scutellaria an ethnopharmacological and phytochemical review. J. Ethnopharmacol. 2010, 128, 279–313. [Google Scholar] [CrossRef]
- Dinda, B.; Dinda, S.; DasSharma, S.; Banik, R.; Chakraborty, A.; Dinda, M. Therapeutic potentials of baicalin and its aglycone, baicalein against inflammatory disorders. Eur. J. Med. Chem. 2017, 131, 68–80. [Google Scholar] [CrossRef]
- Gao, Z.; Huang, K.; Yang, X.; Xu, H. Free radical scavenging and antioxidant activities of flavonoids extracted from the radix of Scutellaria baicalensis Georgi. Biochim. Biophys. Acta Gen. Subj. 1999, 1472, 643–650. [Google Scholar] [CrossRef]
- Zhou, Y.; Hirotani, M.; Yoshikawa, T.; Furuya, T. Flavonoids and phenylrthanoids from hairy root cultures of Scutellaria baicalensis. Phytochemistry 1997, 44, 83–87. [Google Scholar] [CrossRef]
- Nishikawa, K.; Ishimaru, K. Flavonoids in root cultures of Scutellaria baicalensis. J. Plant Physiol. 1997, 151, 633–636. [Google Scholar] [CrossRef]
- Nishikawa, K.; Furukawa, H.; Fujioka, T.; Fujii, H.; Mihashi, K.; Shimomura, K.; Ishimaru, K. Flavone production in transformed root cultures of Scutellaria baicalensis Georgi. Phytochemistry 1999, 52, 885–890. [Google Scholar] [CrossRef]
- Stojakowska, A.; Malarz, J. Flavonoid production in transformed root cultures of Scutellaria baicalensis. J. Plant Physiol. 2000, 156, 121–125. [Google Scholar] [CrossRef]
- Kuzovkina, I.N.; Guseva, A.V.; Alterman, I.E.; Karnachuk, N.A. Flavonoid production in transformed Scutellaria baicalensis roots and ways of its regulation. Russ. J. Plant. Physiol. 2001, 48, 448–452. [Google Scholar] [CrossRef]
- Kuzovkina, I.N.; Guseva, A.V.; Kovács, D.; Szöke, É.; Vdovitchenko, M.Y. Flavones in genetically transformed Scutellaria baicalensis roots and induction of their synthesis by elicitation with methyl jasmonate. Russ. J. Plant Physiol. 2005, 52, 77–82. [Google Scholar] [CrossRef]
- Kovács, G.; Kuzovkina, I.N.; Szöke, É.; Kursinszki, L. HPLC Determination of flavonoids in hairy-root cultures of Scutellaria baicalensis Georgi. Chromatographia 2004, 60, S81–S85. [Google Scholar] [CrossRef]
- Hirotani, M.; Kuroda, R.; Suzuki, H.; Yoshikawa, T. Cloning and expression of UDP-glucose:flavonoid 7-O-glucosyltransferase from hairy root cultures of Scutellaria baicalensis. Planta 2000, 210, 1006–1013. [Google Scholar] [CrossRef]
- Park, N.I.; Xu, H.; Li, X.; Kim, S.-J.; Park, S.U. Enhancement of flavone levels through overexpression of chalcone isomerase in hairy root cultures of Scutellaria baicalensis. Funct. Integr. Genom. 2011, 11, 491–496. [Google Scholar] [CrossRef]
- Park, N.I.; Xu, H.; Li, X.; Kim, Y.S.; Lee, M.Y.; Park, S.U. Overexpression of phenylalanine ammonia-lyase improves flavones production in transgenic hairy root cultures of Scutellaria baicalensis. Process Biochem. 2012, 47, 2575–2580. [Google Scholar] [CrossRef]
- Kim, Y.S.; Kim, Y.B.; Kim, Y.J.; Lee, M.Y.; Park, S.U. Overexpression of cinnamate 4-hydroxylase and 4-coumaroyl CoA ligase prompted flavone accumulation in Scutellaria baicalensis hairy roots. Nat. Prod. Commun. 2014, 9, 803–807. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, C.H.; Xu, H.; Yeo, H.J.; Park, Y.E.; Hwang, G.-S.; Park, N.I.; Park, S.U. Enhancement of the flavone contents of Scutellaria baicalensis hairy roots via metabolic engineering using maize Lc and Arabidopsis PAP1 transcription factors. Metab. Eng. 2021, 64, 64–73. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.S.; Li, X.; Park, W.T.; Uddin, M.R.; Park, N.I.; Kim, Y.B.; Lee, M.Y.; Park, S.U. Influence of media and auxins on growth and falvone production in hairy root cultures of baikal skullcap, Scutellaria baicalensis. Plant Omics J. 2012, 5, 24–27. [Google Scholar]
- Park, C.H.; Kim, Y.S.; Li, X.; Kim, H.H.; Arasu, M.V.; Al-Dhabi, N.A.; Lee, S.Y.; Park, S.U. Influence of different carbohydrates on flavonoid accumulation in hairy root cultures of Scutellaria baicalensis. Nat. Prod. Commun. 2016, 11, 799–802. [Google Scholar] [CrossRef] [Green Version]
- Dikaya, V.S.; Solovyeva, A.I.; Sidorov, R.A.; Solovyev, P.A.; Stepanova, A.Y. The relationship between endogenous b-glucuronidase activity and biologically active flavones-aglycone contents in hairy roots of Baikal skullcap. Chem. Biodivers. 2018, 15, e1700409. [Google Scholar] [CrossRef] [PubMed]
- Solov’eva, A.I.; Evsyukov, S.V.; Sidorov, R.A.; Stepanova, A.Y. Correlation of endogenous β-glucuronidase activity with differentiation of in vitro cultures of Scutellaria baicalensis. Acta Physiol. Plant. 2020, 42, 169. [Google Scholar] [CrossRef]
- Zhao, Q.; Cui, M.-Y.; Levsh, O.; Yang, D.; Liu, J.; Li, J.; Hill, L.; Yang, L.; Hu, Y.; Weng, J.-K.; et al. Two CYP82D enzymes function as flavone hydroxylases in the biosynthesis of root-specific 4’-deoxyflavones in Scutellaria baicalensis. Mol. Plant 2018, 11, 135–148. [Google Scholar] [CrossRef] [Green Version]
- Elkin, Y.N.; Kulesh, N.I.; Stepanova, A.Y.; Solovieva, A.I.; Kargin, V.M.; Manyakhin, A.Y. Methylated flavones of the hairy root culture Scutellaria baicalensis. J. Plant Physiol. 2018, 231, 277–280. [Google Scholar] [CrossRef]
- Wilczańska-Barska, A.; Królicka, A.; Głód, D.; Majdan, M.; Kawiak, A.; Krauze-Baranowska, M. Enhanced accumulation of secondary metabolites in hairy root cultures of Scutellaria lateriflora following elicitation. Biotechnol. Lett. 2012, 34, 1757–1763. [Google Scholar] [CrossRef]
- Kim, J.K.; Kim, Y.S.; Kim, Y.J.; Uddin, M.R.; Kim, Y.B.; Kim, H.H.; Park, S.Y.; Lee, M.Y.; Chung, S.O.; Park, S.U. Comparative analysis of flavonoids and polar metabolites from hairy roots of Scutellaria baicalensis and Scutellaria lateriflora. World J. Microbiol. Biotechnol. 2014, 30, 887–892. [Google Scholar] [CrossRef]
- Marsh, Z.; Yang, T.; Nopo-Olazabal, L.; Wu, S.; Ingle, T.; Joshee, N.; Medina-Bolivar, F. Effect of light, methyl jasmonate and cyclodextrin on production of phenolic compounds in hairy root cultures of Scutellaria lateriflora. Phytochemistry 2014, 107, 50–60. [Google Scholar] [CrossRef] [PubMed]
- Tuan, P.A.; Kim, Y.S.; Kim, Y.; Thwe, A.A.; Li, X.; Park, C.H.; Lee, S.Y.; Park, S.U. Molecular characterization of flavonoid biosynthetic genes and accumulation of baicalin, baicalein, and wogonin in plant and hairy root of Scutellaria lateriflora. Saudi J. Biol. Sci. 2018, 25, 1639–1647. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stepanova, A.Y.; Solov’eva, A.I.; Malunova, M.V.; Salamaikina, S.A.; Panov, Y.M.; Lelishentsev, A.A. Hairy roots of Scutellaria spp. (Lamiaceae) as promising producers of antiviral flavones. Molecules 2021, 26, 3927. [Google Scholar] [CrossRef]
- Kuzovkina, I.N.; Prokof’eva, M.Y.; Umralina, A.R.; Chernysheva, T.P. Morphological and biochemical characteristics of genetically transformed roots of Scutellaria andrachnoides. Russ. J. Plant Physiol. 2014, 61, 697–706. [Google Scholar] [CrossRef]
- Gharari, Z.; Bagheri, K.; Danafar, H.; Sharafi, A. Enhanced flavonoid production in hairy root cultures of Scutellaria bornmuelleri by elicitor induced over-expression of MYB7 and FNSП2 genes. Plant Physiol. Biochem. 2020, 148, 35–44. [Google Scholar] [CrossRef]
- Lei, W.; Tang, S.-H.; Luo, K.-M.; Sun, M. Molecular cloning and expression profiling of a chalcone synthase gene from hairy root cultures of Scutellaria viscidula Bunge. Genet. Mol. Biol. 2010, 33, 285–291. [Google Scholar] [CrossRef] [PubMed]
- Gharari, Z.; Bagheri, K.; Sharafi, A. Establishment of in vitro genetically engineered cultures in Scutellaria orientalis and S. araxensis. Biologia 2020, 75, 2383–2393. [Google Scholar] [CrossRef]
- Heydari, P.; Yavari, M.; Adibi, P.; Asghari, G.; Ghanadian, S.-M.; Dida, G.O.; Khamesipour, F. Medicinal Properties and Active Constituents of Dracocephalum kotschyi and Its Significance in Iran: A Systematic Review. Evid.-Based Complement. Altern. Med. 2019, 2019, 9465309. [Google Scholar] [CrossRef] [Green Version]
- Fattahi, M.; Nazeri, V.; Torras-Claveria, L.; Sefidkon, F.; Cusido, R.M.; Zamani, Z.; Palazon, J. A new biotechnological source of rosmarinic acid and surface flavonoids: Hairy root cultures of Dracocephalum kotschyi Boiss. Ind. Crops Prod. 2013, 50, 256–263. [Google Scholar] [CrossRef]
- Nourozi, E.; Hosseini, B.; Maleki, R.; Mandoulakani, B.A. Iron oxide nanoparticles: A novel elicitor to enhance anticancer flavonoid production and gene expression in Dracocephalum kotschyi hairy-root cultures. J. Sci. Food Agric. 2019, 99, 6418–6430. [Google Scholar] [CrossRef]
- Wang, Y.-F.; Ni, Z.-Y.; Dong, M.; Cong, B.; Shi, Q.-W.; Gu, Y.-C.; Kiyota, H. Secondary metabolites of plants from the genus Saussurea: Chemistry and biological activity. Chem. Biodiv. 2010, 7, 2623–2659. [Google Scholar] [CrossRef] [PubMed]
- Zhao, T.; Li, S.-J.; Zhang, Z.-X.; Zhang, M.-L.; Shi, Q.-W.; Gu, Y.-C.; Dong, M.; Kiyota, H. Chemical constituents from the genus Saussurea and their biological activities. Heterocycl. Commun. 2017, 23, 331–358. [Google Scholar] [CrossRef]
- Chu, C.; Wang, C.; Sun, C.; Hsu, C.; Yin, C.; Chu, C.; Bi, F. Establishment of an efficient medium for anther culture of rice through comparative experiments on the nitrogen sources. Sci. Sin. 1975, 18, 223–231. [Google Scholar]
- Zhao, D.; Fu, C.; Chen, Y.; Ma, F. Transformation of Saussurea medusa for hairy roots and jaceosidin production. Plant Cell Rep. 2004, 23, 468–474. [Google Scholar] [CrossRef]
- Li, F.-X.; Jin, Z.-P.; Zhao, D.-X.; Cheng, L.-Q.; Fu, C.-X.; Ma, F. Overexpression of the Saussurea medusa chalcone isomerase gene in S. involucrata hairy root cultures enhances their biosynthesis of apigenin. Phytochemistry 2006, 67, 553–560. [Google Scholar] [CrossRef]
- Chen, R.; Chen, X.; Zhu, T.; Liu, J.; Xiang, X.; Yu, J.; Tan, H.; Gao, S.; Li, Q.; Fang, Y.; et al. Integrated transcript and metabolite profiles reveal that EbCHI plays an important role in scutellarin accumulation in Erigeron breviscapus hairy roots. Front. Plant Sci. 2018, 9, 789. [Google Scholar] [CrossRef]
- Chung, I.-M.; Hong, S.-B.; Peebles, C.A.M.; Kim, J.-A.; San, K.Y. Effect of the engineered indole pathway on accumulation of phenolic compounds in Catharanthus roseus hairy roots. Biotechnol. Prog. 2007, 23, 327–332. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.N.; Wang, B.; Li, H.; Yao, L.M.; Wu, T.L. Flavonoid production is effectively regulated by RNAi interference of two flavone synthase genes from Glycine max. J. Plant Biol. 2010, 53, 425–432. [Google Scholar] [CrossRef]
- Yan, J.; Wang, B.; Jiang, Y.; Cheng, L.; Wu, T. GmFNSII-Controlled soybean flavone metabolism responds to abiotic stresses and regulates plant salt tolerance. Plant Cell Physiol. 2014, 55, 74–86. [Google Scholar] [CrossRef] [Green Version]
- Staszków, A.; Swarcewicz, B.; Banasiak, J.; Muth, D.; Jasiński, M.; Stobiecki, M. LC/MS profiling of flavonoid glycoconjugates isolated from hairy roots, suspension root cell cultures and seedling roots of Medicago truncatula. Metabolomics 2011, 7, 604–613. [Google Scholar] [CrossRef] [Green Version]
- Gai, Q.-Y.; Jiao, J.; Wang, X.; Fu, Y.-J.; Lu, Y.; Liu, J.; Wang, Z.-Y.; Xu, X.-J. Simultaneous quantification of eleven bioactive phenolic compounds in pigeon pea natural resources and in vitro cultures by ultra-high performance liquid chromatography coupled with triple quadrupole mass spectrometry (UPLC-QqQ-MS/MS). Food Chem. 2021, 335, 127602. [Google Scholar] [CrossRef] [PubMed]
- Gai, Q.-Y.; Lu, Y.; Jiao, J.; Fu, J.-X.; Xu, X.-J.; Yao, L.; Fu, Y.-J. Application of UV-B radiation for enhancing the accumulation of bioactive phenolic compounds in pigeon pea [Cajanus cajan (L.) Millsp.] hairy root cultures. J. Photochem. Photobiol. B Biol. 2022, 228, 112406. [Google Scholar] [CrossRef] [PubMed]
- Miadoková, E. Isoflavonoids-an overview of their biological activities and potential health benefits. Interdiscip. Toxicol. 2009, 2, 211–218. [Google Scholar] [CrossRef] [PubMed]
- Sohn, S.I.; Pandian, S.; Oh, Y.J.; Kang, H.J.; Cho, W.S.; Cho, Y.S. Metabolic engineering of isoflavones: An updated overview. Front. Plant Sci. 2021, 12, 670103. [Google Scholar] [CrossRef] [PubMed]
- Lozovaya, V.V.; Lygin, A.V.; Zernova, O.V.; Li, S.; Hartman, G.L.; Widholm, J.M. Isoflavonoid accumulation in soybean hairy roots upon treatment with Fusarium solani. Plant Physiol. Biochem. 2004, 42, 671–679. [Google Scholar] [CrossRef]
- Jiang, Y.; Hu, Y.; Wang, B.; Wu, T. Bivalent RNA interference to increase isoflavone biosynthesis in soybean (Glycine max). Braz. Arch. Biol. Technol. 2014, 57, 163–170. [Google Scholar] [CrossRef] [Green Version]
- Yan, J.; Wang, B.; Zhong, Y.; Yao, L.; Cheng, L.; Wu, T. The soybean R2R3 MYB transcription factor GmMYB100 negatively regulates plant flavonoid biosynthesis. Plant Mol. Biol. 2015, 89, 35–48. [Google Scholar] [CrossRef]
- Jia, T.; An, J.; Liu, Z.; Yu, B.; Chen, J. Salt stress induced soybean GmIFS1 expression and isoflavone accumulation and salt tolerance in transgenic soybean cotyledon hairy roots and tobacco. Plant Cell Tissue Organ Cult. 2017, 128, 469–477. [Google Scholar] [CrossRef]
- Han, X.; Yin, Q.; Liu, J.; Jiang, W.; Di, S.; Pang, Y. GmMYB58 and GmMYB205 are seed-specific activators for isoflavonoid biosynthesis in Glycine max. Plant Cell Rep. 2017, 36, 1889–1902. [Google Scholar] [CrossRef]
- Yin, Q.; Shen, G.; Di, S.; Fan, C.; Chang, Z.; Pang, Y. Genome-wide identification and functional characterization of UDP-glucosyltransferase genes involved in flavonoid biosynthesis in Glycine max. Plant Cell Physiol. 2017, 58, 1558–1572. [Google Scholar] [CrossRef]
- Zhou, Y.; Huang, J.; Zhang, X.; Zhu, L.; Wang, X.; Guo, N.; Zhao, J.; Xing, H. Overexpression of chalcone isomerase (CHI) increases resistance against Phytophthora sojae in soybean. J. Plant Biol. 2018, 61, 309–319. [Google Scholar] [CrossRef]
- Uchida, K.; Sawada, Y.; Ochiai, K.; Sato, M.; Inaba, J.; Hirai, M.Y. Identification of a unique type of isoflavone O-methyltransferase, GmIOMT1, based on multi-omics analysis of soybean under biotic stress. Plant Cell Physiol. 2020, 61, 1974–1985. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, M.Z.; Zhang, Y.; Zeng, X.; Li, P.; Wang, X.; Benedito, V.A.; Zhao, J. Isoflavone malonyl-CoA acyltransferase GmMaT2 is involved in nodulation of soybean by modifying synthesis and secretion of isoflavones. J. Exp. Bot. 2021, 72, 1349–1369. [Google Scholar] [CrossRef]
- Theboral, J.; Sivanandhan, G.; Subramanyam, K.; Arun, M.; Selvaraj, N.; Manickavasagam, M.; Ganapathi, A. Enhanced production of isoflavones by elicitation in hairy root cultures of Soybean. Plant Cell Tissue Organ Cult. 2014, 117, 477–481. [Google Scholar] [CrossRef]
- Liu, J.; Jiang, W.; Xia, Y.; Wang, X.; Shen, G.; Pang, Y. Genistein-specific G6DT gene for the inducible production of wighteone in Lotus japonicus. Plant Cell Physiol. 2018, 59, 128–141. [Google Scholar] [CrossRef]
- Saviranta, N.M.; Julkunen-Tiitto, R.; Oksanen, E.; Karjalainen, R.O. Red clover (Trifolium pratense L.) isoflavones: Root phenolic compounds affected by biotic and abiotic stress factors. J. Sci. Food Agric. 2010, 90, 418–423. [Google Scholar] [CrossRef]
- Kumar, M.A.; Pammi, S.S.S.; Sukanya, M.S.; Giri, A. Enhanced production of pharmaceutically important isoflavones from hairy root rhizoclones of Trifolium pratense L. In Vitro Cell Dev. Biol. 2018, 54, 94–103. [Google Scholar] [CrossRef]
- Reis, A.; Boutet-Mercey, S.; Massot, S.; Ratet, P.; Zuanazzi, J.A.S. Isoflavone production in hairy root cultures and plantlets of Trifolium pratense. Biotechnol. Lett. 2019, 41, 427–442. [Google Scholar] [CrossRef] [PubMed]
- Park, Y.J.; Thwe, A.A.; Li, X.; Kim, Y.J.; Kim, J.K.; Arasu, M.V.; Al-Dhabi, N.A.; Park, S.U. Triterpene and flavonoid biosynthesis and metabolic profiling of hairy roots, adventitious roots, and seedling roots of Astragalus membranaceus. J. Agric. Food Chem. 2015, 63, 8862–8869. [Google Scholar] [CrossRef]
- Jiao, J.; Gai, Q.-Y.; Luo, M.; Peng, X.; Zhao, C.-J.; Fu, Y.-J.; Ma, W. Direct determination of astragalosides and isoflavonoids from fresh Astragalus membranaceus hairy root cultures by high speed homogenization coupled with cavitation-accelerated extraction followed by liquid chromatography-tandem mass spectrometry. RSC Adv. 2015, 5, 34672–34681. [Google Scholar] [CrossRef]
- Jiao, J.; Gai, Q.-Y.; Wang, W.; Luo, M.; Gu, C.-B.; Fu, Y.-J.; Ma, W. Ultraviolet radiation-elicited enhancement of isoflavonoid accumulation, biosynthetic gene expression, and antioxidant activity in Astragalus membranaceus hairy root cultures. J. Agric. Food Chem. 2015, 63, 8216–8224. [Google Scholar] [CrossRef] [PubMed]
- Gai, Q.-Y.; Jiao, J.; Luo, M.; Wang, W.; Gu, C.-B.; Fu, Y.-J.; Ma, W. Tremendous enhancements of isoflavonoid biosynthesis, associated gene expression and antioxidant capacity in Astragalus membranaceus hairy root cultures elicited by methyl jasmonate. Process Biochem. 2016, 51, 642–649. [Google Scholar] [CrossRef]
- Jiao, J.; Gai, Q.-Y.; Niu, L.-L.; Wang, X.-Q.; Guo, N.; Zang, Y.-P.; Fu, Y.-J. Enhanced production of two bioactive isoflavone aglycones in Astragalus membranaceus hairy root cultures by combining deglycosylation and elicitation of immobilized edible Aspergillus niger. J. Agricul. Food Chem. 2017, 65, 9078–9086. [Google Scholar] [CrossRef]
- Cui, Y.; Taniguchi, S.; Kuroda, T.; Hatano, T. Constituents of Psoralea corylifolia fruits and their effects on methicillin-resistant Staphylococcus aureus. Molecules 2015, 20, 12500–12511. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, Y.; Liu, X.; Wu, Y.; Li, Y.; Guo, F. Meroterpenes from Psoralea corylifolia against Pyricularia oryzae. Planta Med. 2014, 80, 1298–1303. [Google Scholar] [CrossRef] [PubMed]
- Bourgaud, F.; Bouque, V.; Guckert, A. Production of flavonoids by Psoralea hairy root cultures. Plant Cell Tissue Organ Cult. 1999, 56, 97–104. [Google Scholar] [CrossRef]
- Abhyankar, G.; Reddy, V.D.; Giri, C.C.; Rao, K.V.; Lakshmi, V.V.S.; Prabhakar, S.; Vairamani, M.; Thippeswamy, B.S.; Bhattacharya, P.S. Amplified fragment length polymorphism and metabolomic profiles of hairy roots of Psoralea corylifolia L. Phytochemistry 2005, 66, 2441–2457. [Google Scholar] [CrossRef]
- Shinde, A.N.; Malpathak, N.; Fulzele, D.P. Impact of nutrient components on production of the phytoestrogens daidzein and genistein by hairy roots of Psoralea corylifolia. J. Nat. Med. 2010, 64, 346–353. [Google Scholar] [CrossRef]
- Cherdshewasart, W.; Sutjit, W. Correlation of antioxidant activity and major isoflavonoid contents of the phytoestrogen-rich Pueraria mirifica and Pueraria lobata tubers. Phytomedicine 2008, 15, 38–43. [Google Scholar] [CrossRef]
- Shi, H.-P.; Kintzios, S. Genetic transformation of Pueraria phaseoloides with Agrobacterium rhizogenes and puerarin production in hairy roots. Plant Cell Rep. 2003, 21, 1103–1107. [Google Scholar] [CrossRef]
- Kintzios, S.; Makri, O.; Pistola, E.; Matakiadis, T.; Shi, H.P.; Economou, A. Scale-up production of puerarin from hairy roots of Pueraria phaseoloides in an airlift bioreactor. Biotechnol. Lett. 2004, 26, 1057–1059. [Google Scholar] [CrossRef] [PubMed]
- Udomsuk, L.; Jarukamjorn, K.; Tanaka, H.; Putalun, W. Isoflavonoid Production in a Hairy Roots Culture of Pueraria candollei. Z. Naturforsch. 2009, 64, 687–691. [Google Scholar] [CrossRef] [PubMed]
- Udomsuk, L.; Jarukamjorn, K.; Tanaka, H.; Putalun, W. Improved isoflavonoid production in Pueraria candollei hairy root cultures using elicitation. Biotechnol. Lett. 2011, 33, 369–374. [Google Scholar] [CrossRef] [PubMed]
- Udomsin, O.; Yusakul, G.; Kraithong, W.; Udomsuk, L.; Kitisripanya, T.; Juengwatanatrakul, T.; Putalun, W. Enhanced accumulation of high-value deoxymiroestrol and isoflavonoids using hairy root as a sustainable source of Pueraria candollei var. mirifica. Plant Cell Tissue Organ Cult. 2019, 136, 141–151. [Google Scholar] [CrossRef]
- Danphitsanuparn, P.; Boonsnongcheep, P.; Boriboonkaset, T.; Chintapakorn, Y.; Prathanturarug, S. Effects of Agrobacterium rhizogenes strains and other parameters on production of isoflavonoids in hairy roots of Pueraria candollei Grah. ex Benth. var. candollei. Plant Cell Tissue Organ Cult. 2012, 111, 315–322. [Google Scholar] [CrossRef]
- Kim, S.; Cha, M.-S.; Lee, E.; Kim, I.; Kwon, J.E.; Kang, S.C.; Park, T.-H. In vitro induction of hairy root from isoflavones-producing Korean wild arrowroot Pueraria lobata. J. Plant Biotechnol. 2012, 39, 205–211. [Google Scholar] [CrossRef]
- Wang, X.; Li, C.; Zhou, C.; Li, J.; Zhang, Y. Molecular characterization of the C-glucosylation for puerarin biosynthesis in Pueraria lobata. Plant J. 2017, 90, 535–546. [Google Scholar] [CrossRef] [Green Version]
- Gampe, N.; Szakács, Z.; Darcsi, A.; Boldizsár, I.; Szőke, É.; Kuzovkina, I.; Kursinszki, L.; Béni, S. Qualitative and quantitative phytochemical analysis of Ononis hairy root cultures. Front. Plant Sci. 2021, 11, 622585. [Google Scholar] [CrossRef]
- Lee, M.R. Liquorice (Glycyrrhiza glabra): The journey of the sweet root from Mesopotamia to England. J. R. Coll. Physicians Edinb. 2018, 48, 378–382. [Google Scholar] [CrossRef]
- Asl, M.N.; Hosseinzadeh, H. Review of pharmacological effects of Glycyrrhiza sp. and its bioactive compounds. Phytother. Res. 2008, 22, 709–724. [Google Scholar] [CrossRef]
- Asada, Y.; Li, W.; Yoshikawa, T. Isoprenylated flavonoids from hairy root cultures of Glycyrrhiza glabra. Phytochemistry 1998, 47, 389–392. [Google Scholar] [CrossRef]
- Li, W.; Asada, Y.; Yoshikawa, T. Antimicrobial flavonoids from Glycyrrhiza glabra hairy root cultures. Planta Med. 1998, 64, 746–747. [Google Scholar] [CrossRef] [PubMed]
- Asada, Y.; Li, W.; Yoshikawa, T. The first prenylated biaurone, licoagrone from hairy root cultures of Glycyrrhiza glabra. Phytochemistry 1999, 50, 1015–1019. [Google Scholar] [CrossRef]
- Li, W.; Asada, Y.; Yoshikawa, T. Flavonoid constituents from Glycyrrhiza glabra hairy root cultures. Phytochemistry 2000, 55, 447–456. [Google Scholar] [CrossRef]
- Li, W.; Asada, Y.; Koike, K.; Hirotani, M.; Rui, H.; Yoshikawa, T.; Nikaido, T. Flavonoids from Glycyrrhiza pallidiflora hairy root cultures. Phytochemistry 2001, 58, 595–598. [Google Scholar] [CrossRef]
- Li, W.; Koike, K.; Asada, Y.; Hirotani, M.; Rui, H.; Yoshikawa, T.; Nikaido, T. Flavonoids from Glycyrrhiza pallidiflora hairy root cultures. Phytochemistry 2002, 60, 351–355. [Google Scholar] [CrossRef]
- Zhang, H.-C.; Liu, J.-M.; Lu, H.-Y.; Gao, S.-L. Enhanced flavonoid production in hairy root cultures of Glycyrrhiza uralensis Fisch by combining the over-expression of chalcone isomerase gene with the elicitation treatment. Plant Cell Rep. 2009, 28, 1205–1213. [Google Scholar] [CrossRef]
- Zhang, H.-C.; Liu, J.-M.; Chen, H.-M.; Gao, C.-C.; Lu, H.-Y.; Zhou, H.; Li, Y.; Gao, S.-L. Up-regulation of licochalcone A biosynthesis and secretion by Tween 80 in hairy root cultures of Glycyrrhiza uralensis Fisch. Mol. Biotechnol. 2011, 47, 50–56. [Google Scholar] [CrossRef]
- Yin, Y.-C.; Hou, J.-M.; Tian, S.-K.; Yang, L.; Zhang, Z.-X.; Li, W.-D.; Liu, Y. Overexpressing chalcone synthase (CHS) gene enhanced flavonoids accumulation in Glycyrrhiza uralensis hairy roots. Bot. Lett. 2020, 167, 219–231. [Google Scholar] [CrossRef]
- Łuczkiewicz, M.; Kokotkiewicz, A. Genista tinctoria hairy root cultures for selective production of isoliquiritigenin. Z. Naturforsch. 2005, 60, 867–875. [Google Scholar] [CrossRef]
- Alikaridis, F.; Papadakis, D.; Pantelia, K.; Kephalas, T. Flavonolignan production from Silybum marianum transformed and untransformed root cultures. Fitoterapia 2000, 71, 379–384. [Google Scholar] [CrossRef]
- Rahnama, H.; Hasanloo, T.; Shams, M.R.; Sepehrifar, R. Silymarin production by hairy root culture of Silybum marianum (L.) Gaertn. Iran. J. Biotechnol. 2008, 6, 113–118. [Google Scholar]
- Chong, J.; Poutaraud, A.; Hugueney, P. Metabolism and roles of stilbenes in plants. Plant Sci. 2009, 177, 143–155. [Google Scholar] [CrossRef]
- Akinwumi, B.C.; Bordun, K.-A.M.; Anderson, H.D. Biological activities of stilbenoids. Int. J. Mol. Sci. 2018, 19, 792. [Google Scholar] [CrossRef] [Green Version]
- Medina-Bolivar, F.; Condori, J.; Rimando, A.M.; Hubstenberger, J.; Shelton, K.; O’Keefe, S.F.; Bennett, S.; Dolan, M.C. Production and secretion of resveratrol in hairy root cultures of peanut. Phytochemistry 2007, 68, 1992–2003. [Google Scholar] [CrossRef] [PubMed]
- Abbott, J.A.; Medina-Bolivar, F.; Martin, E.M.; Engelberth, A.S.; Villagarcia, H.; Clausen, E.C.; Carrier, D.J. Purification of resveratrol, arachidin-1, and arachidin-3 from hairy root cultures of peanut (Arachis hypogaea) and determination of their antioxidant activity and cytotoxicity. Biotechnol. Prog. 2010, 26, 1344–1351. [Google Scholar] [CrossRef] [PubMed]
- Condori, J.; Sivakumar, G.; Hubstenberger, J.; Dolan, M.C.; Sobolev, V.S.; Medina-Bolivar, F. Induced biosynthesis of resveratrol and the prenylated stilbenoids arachidin-1 and arachidin-3 in hairy root cultures of peanut: Effects of culture medium and growth stage. Plant Physiol. Biochem. 2010, 48, 310–318. [Google Scholar] [CrossRef]
- Yang, T.; Fang, L.; Nopo-Olazabal, C.; Condori, J.; Nopo-Olazabal, L.; Balmaceda, C.; Medina-Bolivar, F. Enhanced production of resveratrol, piceatannol, arachidin-1, and arachidin-3 in hairy root cultures of peanut co-treated with methyl jasmonate and cyclodextrin. J. Agric. Food Chem. 2015, 63, 3942–3950. [Google Scholar] [CrossRef]
- Yang, T.; Fang, L.; Rimando, A.M.; Sobolev, V.; Mockaitis, K.; Medina-Bolivar, F. A Stilbenoid-specific prenyltransferase utilizes dimethylallyl pyrophosphate from the plastidic terpenoid pathway. Plant Physiol. 2016, 171, 2483–2498. [Google Scholar] [CrossRef] [Green Version]
- Yang, T.; Fang, L.; Sanders, S.; Jayanthi, S.; Rajan, G.; Podicheti, R.; Thallapuranam, S.K.; Mockaitis, K.; Medina-Bolivar, F. Stilbenoid prenyltransferases define key steps in the diversification of peanut phytoalexins. J. Biol. Chem. 2018, 293, 28–46. [Google Scholar] [CrossRef] [Green Version]
- Mohammadhosseinpour, S.; Ho, L.-C.; Fang, L.; Xu, J.; Medina-Bolivar, F. Arachidin-1, a prenylated stilbenoid from peanut, induces apoptosis in triple-negative breast cancer cells. Int. J. Mol. Sci. 2022, 23, 1139. [Google Scholar] [CrossRef] [PubMed]
- Fang, L.; Yang, T.; Medina-Bolivar, F. Production of prenylated stilbenoids in hairy root cultures of peanut (Arachis hypogaea) and its wild relatives A. ipaensis and A. duranensis via an optimized elicitation procedure. Molecules 2020, 25, 509. [Google Scholar] [CrossRef] [Green Version]
- Brents, L.K.; Medina-Bolivar, F.; Seely, K.A.; Nair, V.; Bratton, S.M.; Ñopo-Olazabal, L.; Patel, R.Y.; Liu, H.; Doerksen, R.J.; Prather, P.L.; et al. Natural prenylated resveratrol analogs arachidin-1 and -3 demonstrate improved glucuronidation profiles and have affinity for cannabinoid receptors. Xenobiotica 2012, 42, 139–156. [Google Scholar] [CrossRef] [Green Version]
- Pilaisangsuree, V.; Somboon, T.; Tonglairoum, P.; Keawracha, P.; Wongsa, T.; Kongbangkerd, A.; Limmongkon, A. Enhancement of stilbene compounds and anti-inflammatory activity of methyl jasmonate and cyclodextrin elicited peanut hairy root. Plant Cell Tissue Organ Cult. 2018, 132, 165–179. [Google Scholar] [CrossRef]
- Somboon, T.; Chayjarung, P.; Pilaisangsuree, V.; Keawracha, P.; Tonglairoum, P.; Kongbangkerd, A.; Wongkrajang, K.; Limmongkon, A. Methyl jasmonate and cyclodextrin-mediated defense mechanism and protective effect in response to paraquat-induced stress in peanut hairy root. Phytochemistry 2019, 163, 11–22. [Google Scholar] [CrossRef] [PubMed]
- Wongshaya, P.; Chayjarung, P.; Tothong, C.; Pilaisangsuree, V.; Somboon, T.; Kongbangkerd, A.; Limmongkon, A. Effect of light and mechanical stress in combination with chemical elicitors on the production of stilbene compounds and defensive responses in peanut hairy root culture. Plant Physiol. Biochem. 2020, 157, 93–104. [Google Scholar] [CrossRef]
- Eungsuwan, N.; Chayjarung, P.; Pankam, J.; Pilaisangsuree, V.; Wongshaya, P.; Kongbangkerd, A.; Sriphannam, C.; Limmongkon, A. Production and antimicrobial activity of trans-resveratrol, trans-arachidin-1 and trans-arachidin-3 from elicited peanut hairy root cultures in shake flasks compared with bioreactors. J. Biotechnol. 2021, 326, 28–36. [Google Scholar] [CrossRef] [PubMed]
- Gajurel, G.; Hasan, R.; Medina-Bolivar, F. Antioxidant assessment of prenylated stilbenoid-rich extracts from elicited hairy root cultures of three cultivars of peanut (Arachis hypogaea). Molecules 2021, 26, 6778. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.; Yagiz, Y.; Hsu, W.-Y.; Simonne, A.; Lu, J.; Marshall, M.R. Antioxidant, antibacterial, and antibiofilm properties of polyphenols from muscadine grape (Vitis rotundifolia Michx.) pomace against selected foodborne pathogens. J. Agricul. Food Chem. 2014, 62, 6640–6649. [Google Scholar] [CrossRef]
- Ñopo-Olazabal, C.; Hubstenberger, J.; Ñopo-Olazabal, L.; Medina-Bolivar, F. Antioxidant activity of selected stilbenoids and their bioproduction in hairy root cultures of muscadine grape (Vitis rotundifolia Michx.). J. Agric. Food Chem. 2013, 61, 11744–11758. [Google Scholar] [CrossRef]
- Ñopo-Olazabal, C.; Condori, J.; Ñopo-Olazabal, L.; Medina-Bolivar, F. Differential induction of antioxidant stilbenoids in hairy roots of Vitis rotundifolia treated with methyl jasmonate and hydrogen peroxide. Plant Physiol. Biochem. 2014, 74, 50–69. [Google Scholar] [CrossRef] [PubMed]
- Tisserant, L.-P.; Aziz, A.; Jullian, N.; Jeandet, P.; Clément, C.; Courot, E.; Boitel-Conti, M. Enhanced stilbene production and excretion in Vitis vinifera cv Pinot Noir hairy root cultures. Molecules 2016, 21, 1703. [Google Scholar] [CrossRef]
- Tisserant, L.-P.; Hubert, J.; Lequart, M.; Borie, N.; Maurin, N.; Pilard, S.; Jeandet, P.; Aziz, A.; Renault, J.-H.; Nuzillard, J.-M.; et al. 13C NMR and LC-MS profiling of stilbenes from elicited grapevine hairy root cultures. J. Nat. Prod. 2016, 79, 2846–2855. [Google Scholar] [CrossRef] [PubMed]
- Höll, J.; Vannozzi, A.; Czemmel, S.; D’Onofrio, C.; Walker, A.R.; Rausch, T.; Lucchin, M.; Boss, P.K.; Dry, I.B.; Bogs, J. The R2R3-MYB transcription factors MYB14 and MYB15 regulate stilbene biosynthesis in Vitis vinifera. Plant Cell 2013, 25, 4135–4149. [Google Scholar] [CrossRef] [Green Version]
- Hosseini, S.M.; Bahramnejad, B.; Baneh, H.D.; Emamifar, A.; Goodwin, P.H. Hairy root culture optimization and resveratrol production from Vitis vinifera subsp. sylvesteris. World J. Microbiol. Biotechnol. 2017, 33, 67. [Google Scholar] [CrossRef] [PubMed]
- Hidalgo, D.; Martinez-Márquez, A.; Moyano, E.; Bru-Martinez, R.; Corchete, P.; Palazon, J. Bioconversion of stilbenes in genetically engineered root and cell cultures of tobacco. Sci. Rep. 2017, 7, 45331. [Google Scholar] [CrossRef] [Green Version]
- Jiao, J.; Gai, Q.-Y.; Wang, X.; Liu, J.; Lu, Y.; Wang, Z.-Y.; Xu, X.-J.; Fu, Y.-J. Effective Production of Phenolic Compounds with Health Benefits in Pigeon Pea [Cajanus cajan (L.) Millsp.] Hairy Root Cultures. J. Agric. Food Chem. 2020, 68, 8350–8361. [Google Scholar] [CrossRef]
- Gajurel, G.; Ñopo-Olazabal, l.; Hendrix, E.; Medina-Bolivar, F. Production and secretion of cajaninstilbene acid in hairy root cultures of pigeon pea (Cajanus cajan) co-treated with multiple elicitors. Planta 2022, 11, 834. [Google Scholar] [CrossRef]
- Espín, J.C.; Larrosa, M.; García-Conesa, M.T.; Thomás-Barberán, F. Biological significance of urolithins, the gut microbial ellagic acid-derived metabolites: The evidence so far. Evid.-Based Complement. Altern. Med. 2013, 2013, 270418. [Google Scholar] [CrossRef] [Green Version]
- Al-Harbi, S.A.; Abdulrahman, A.O.; Zamzami, M.A.; Khan, M.I. Urolithins: The gut based polyphenol metabolites of ellagitannins in cancer prevention, a review. Front. Nutr. 2021, 8, 647582. [Google Scholar] [CrossRef]
- Vini, R.; Azeez, J.M.; Remadevi, V.; Susmi, T.R.; Ayswarya, R.S.; Sujatha, A.S.; Muraleedharan, P.; Lathika, L.M.; Sreeharshan, S. Urolithins: The colon microbiota metabolites as endocrine modulators: Prospects and perspectives. Front. Nutr. 2022, 8, 800990. [Google Scholar] [CrossRef] [PubMed]
- Ishimaru, K.; Hirose, M.; Takahashi, K.; Koyama, K.; Shimomura, K. Tannin production in hairy root cultures of Sanguisorba officinalis L. Plant Tissue Cult. Lett. 1991, 8, 114–117. [Google Scholar] [CrossRef]
- Ishimaru, K.; Shimomura, K. Tannin production in hairy root culture of Geranium thunbergii. Phytochemistry 1991, 30, 825–828. [Google Scholar] [CrossRef]
- Ishimaru, K.; Yoshimatsu, K.; Yamakawa, T.; Kamada, H.; Shimomura, K. Phenolic constituents in tissue cultures of Phyllanthus niruri. Phytochemistry 1992, 31, 2015–2018. [Google Scholar] [CrossRef]
- Bakkali, A.-T.; Jaziri, M.; Ishimaru, K.; Tanaka, N.; Shimomura, K.; Yoshimatsu, K.; Homes, J.; Vanhaelen, M. Tannin production in hairy root cultures of Lawsonia inermis. J. Plant. Physiol. 1997, 151, 505–508. [Google Scholar] [CrossRef]
- Ono, N.N.; Bandaranayake, P.C.G.; Tian, L. Establishment of pomegranate (Punica granatum) hairy root cultures for genetic interrogation of the hydrolyzable tannin biosynthetic pathway. Planta 2012, 236, 931–941. [Google Scholar] [CrossRef] [PubMed]
- Ono, N.N.; Qin, X.; Wilson, A.E.; Li, G.; Tian, L. Two UGT84 family glycosyltransferases catalyze a critical reaction of hydrolyzable tannin biosynthesis in pomegranate (Punica granatum). PLoS ONE 2016, 11, e0156319. [Google Scholar] [CrossRef]
- Chang, L.; Wu, S.; Tian, L. Effective genome editing and identification of a regiospecific gallic acid 4-O-glycosyltransferase in pomegranate (Punica granatum L.). Hortic. Res. 2019, 6, 123. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
Simplified scheme of biosynthesis of selected plant polyphenols and positions of biosynthetic enzymes mentioned in the review (PAL—phenylalanine ammonia lyase; C4H—cinnamate 4-hydroxylase; 4CL—4-coumarate:CoA ligase; STS—stilbene synthase; RS—resveratrol synthase; CHS—chalcone synthase; CHI—chalcone isomerase; FNS—flavone synthase; IFS—isoflavone synthase; F3βOH—flavanone 3-β-hydroxylase; F3H—flavone 3-hydroxylase; F6H—flavone 6-hydroxylase; F8H—flavone 8-hydroxylase; IFD—isoflavone dehydratase; DFR—dihydroflavonol 4-reductase; ANS—anthocyanidin synthase; ANR—anthocyanidin reductase; LAR—leucoanthocyanidin reductase). Hydroxybenzoic acids group include gallic acid, a precursor of hydrolyzable tannins; PAs—proanthocyanidins).
Figure 1.
Simplified scheme of biosynthesis of selected plant polyphenols and positions of biosynthetic enzymes mentioned in the review (PAL—phenylalanine ammonia lyase; C4H—cinnamate 4-hydroxylase; 4CL—4-coumarate:CoA ligase; STS—stilbene synthase; RS—resveratrol synthase; CHS—chalcone synthase; CHI—chalcone isomerase; FNS—flavone synthase; IFS—isoflavone synthase; F3βOH—flavanone 3-β-hydroxylase; F3H—flavone 3-hydroxylase; F6H—flavone 6-hydroxylase; F8H—flavone 8-hydroxylase; IFD—isoflavone dehydratase; DFR—dihydroflavonol 4-reductase; ANS—anthocyanidin synthase; ANR—anthocyanidin reductase; LAR—leucoanthocyanidin reductase). Hydroxybenzoic acids group include gallic acid, a precursor of hydrolyzable tannins; PAs—proanthocyanidins).

Figure 2.
Chemical structures of selected anthocyanins and procyanidin B1.

Figure 3.
Chemical structures of selected flavonols (upper row) and flavanols.

Figure 4.
Chemical structures of selected flavones and flavanones.

Figure 5.
Chemical structures of selected isoflavonoids.

Figure 6.
Chemical structures of selected rare and atypical flavonoids found in HRs.


Figure 7.
Chemical structures of selected stilbenoids produced by HRs.

Figure 8.
Chemical structures of gallic acid, ellagic acid and selected hydrolyzable tannins.



{kind=link}
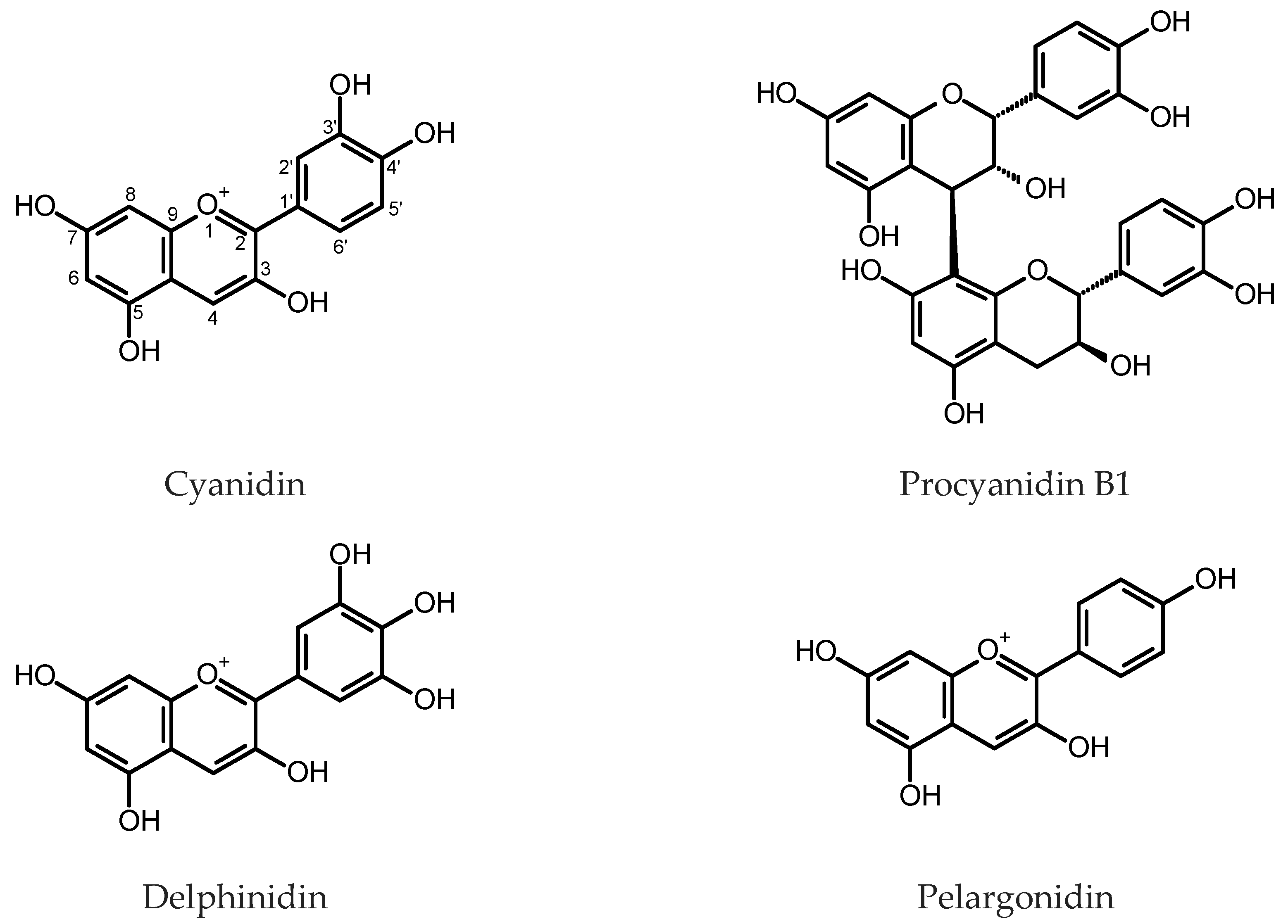{kind=link}
{kind=link}
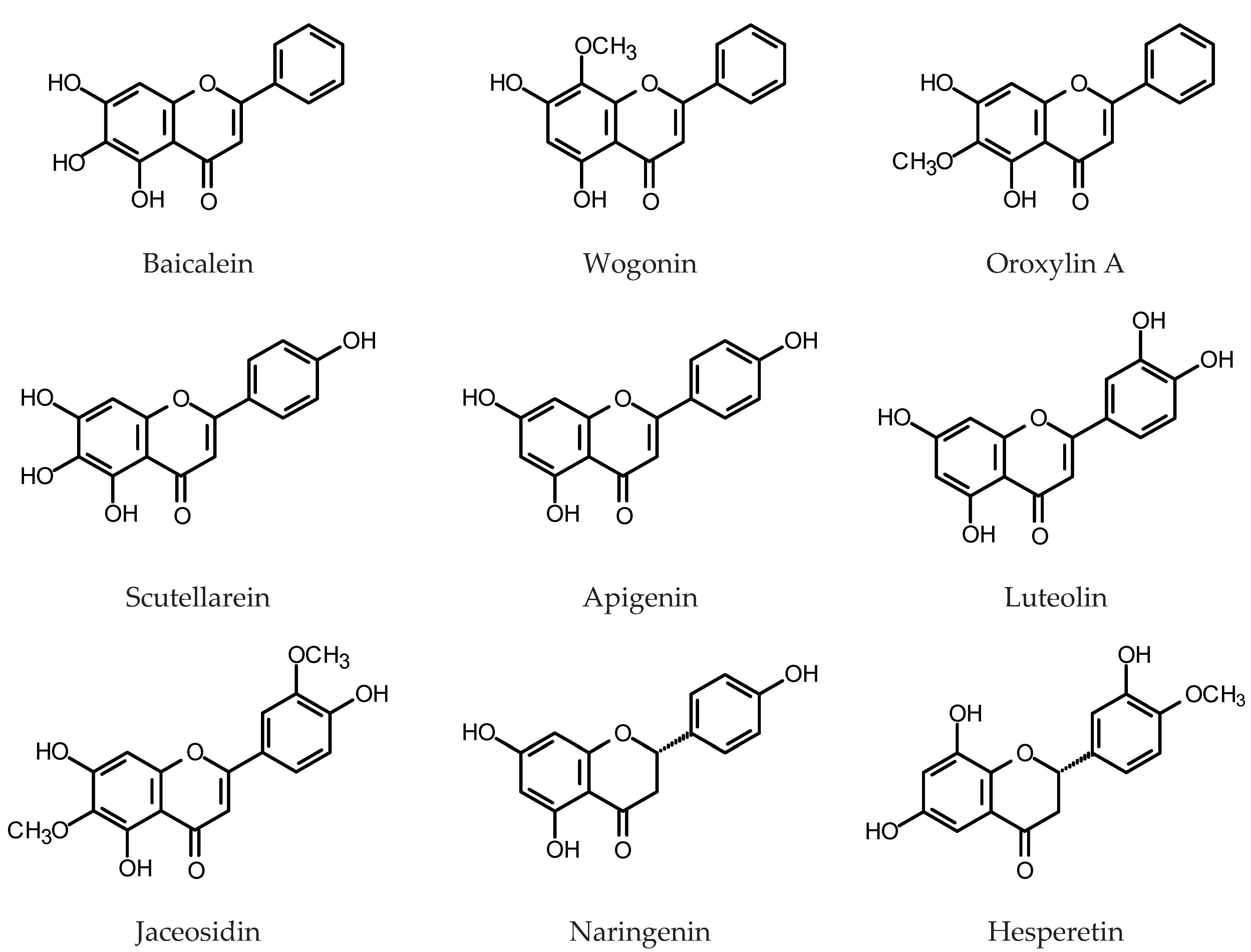{kind=link}
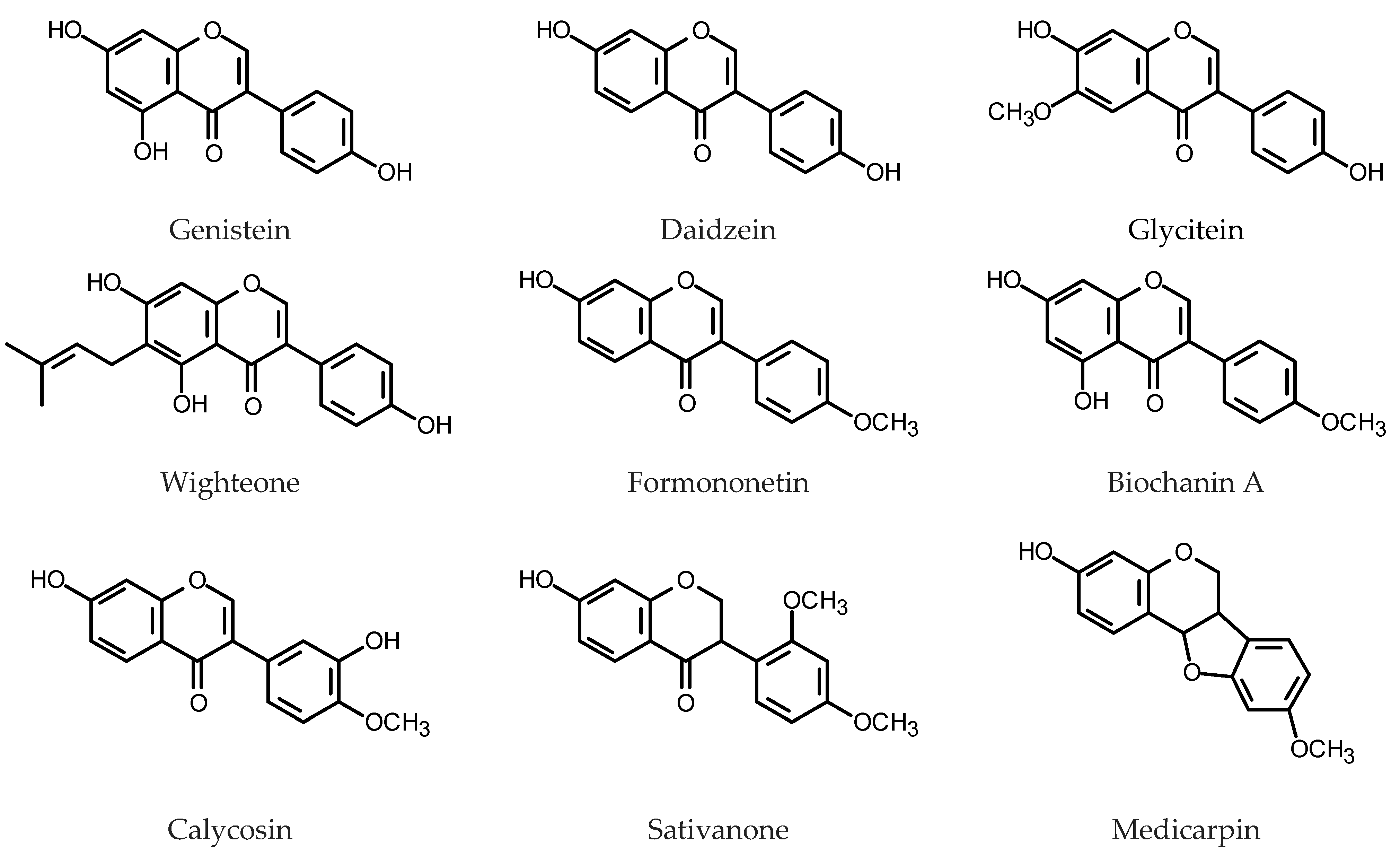{kind=link}
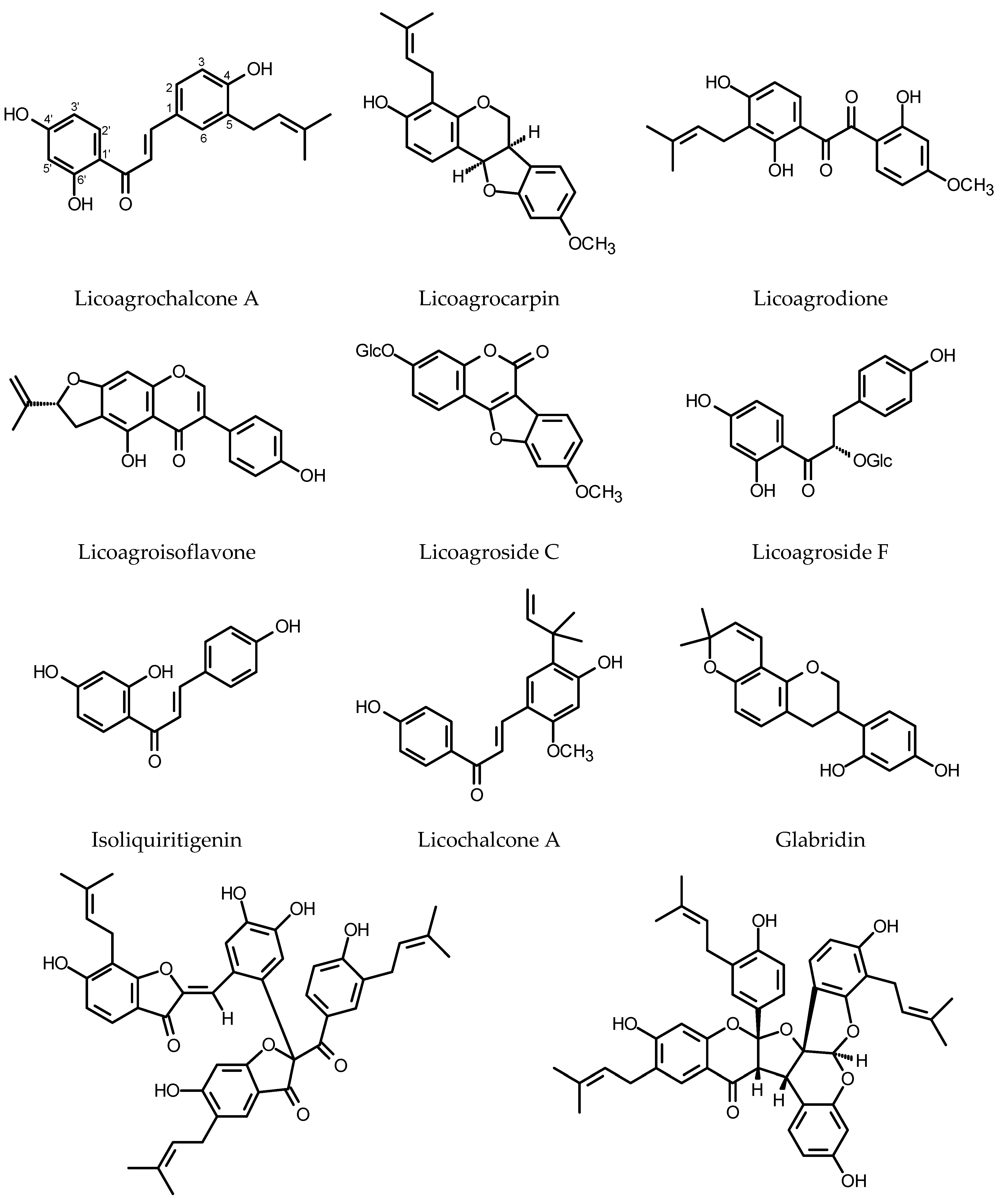{kind=link}
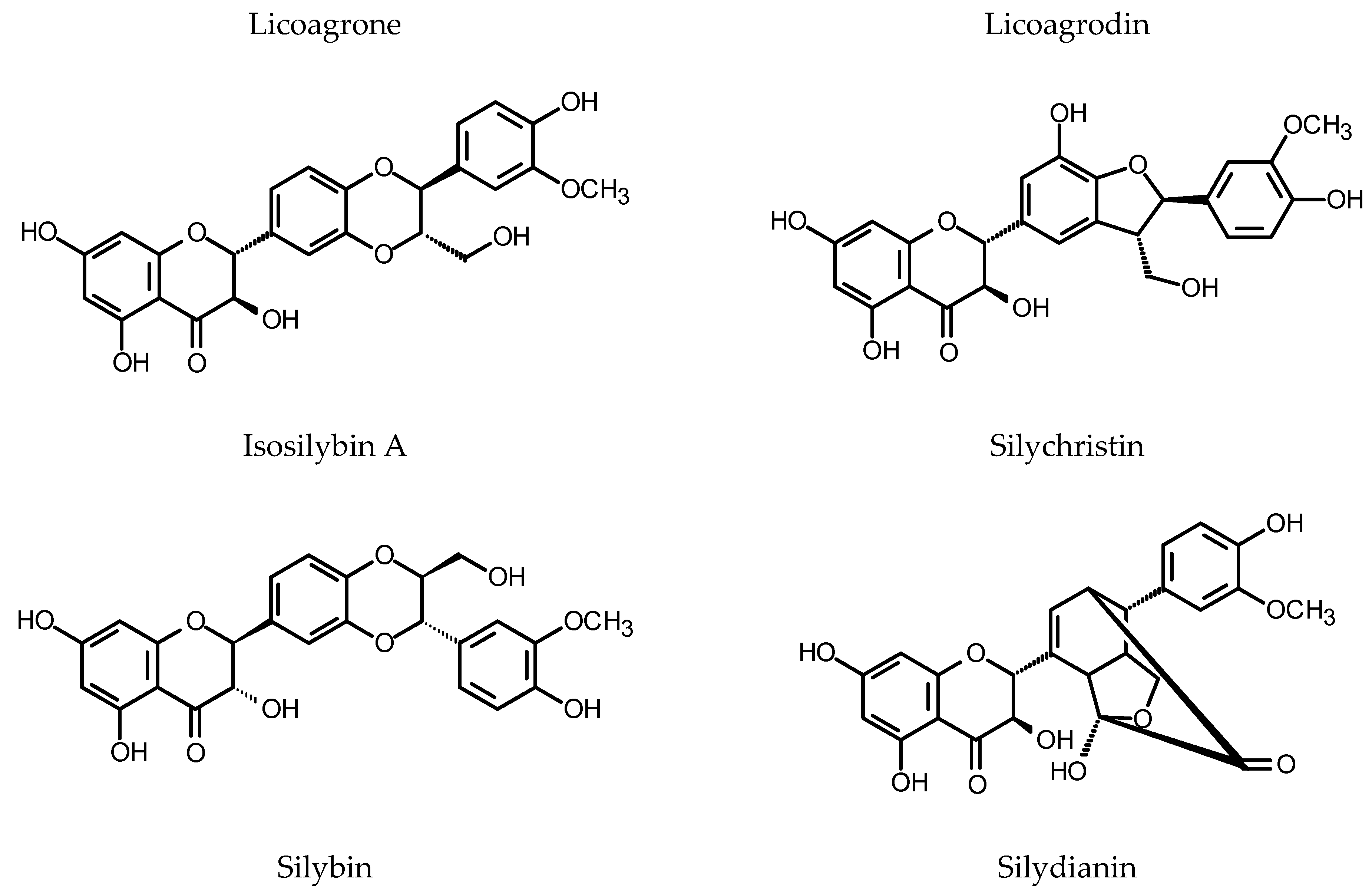{kind=link}
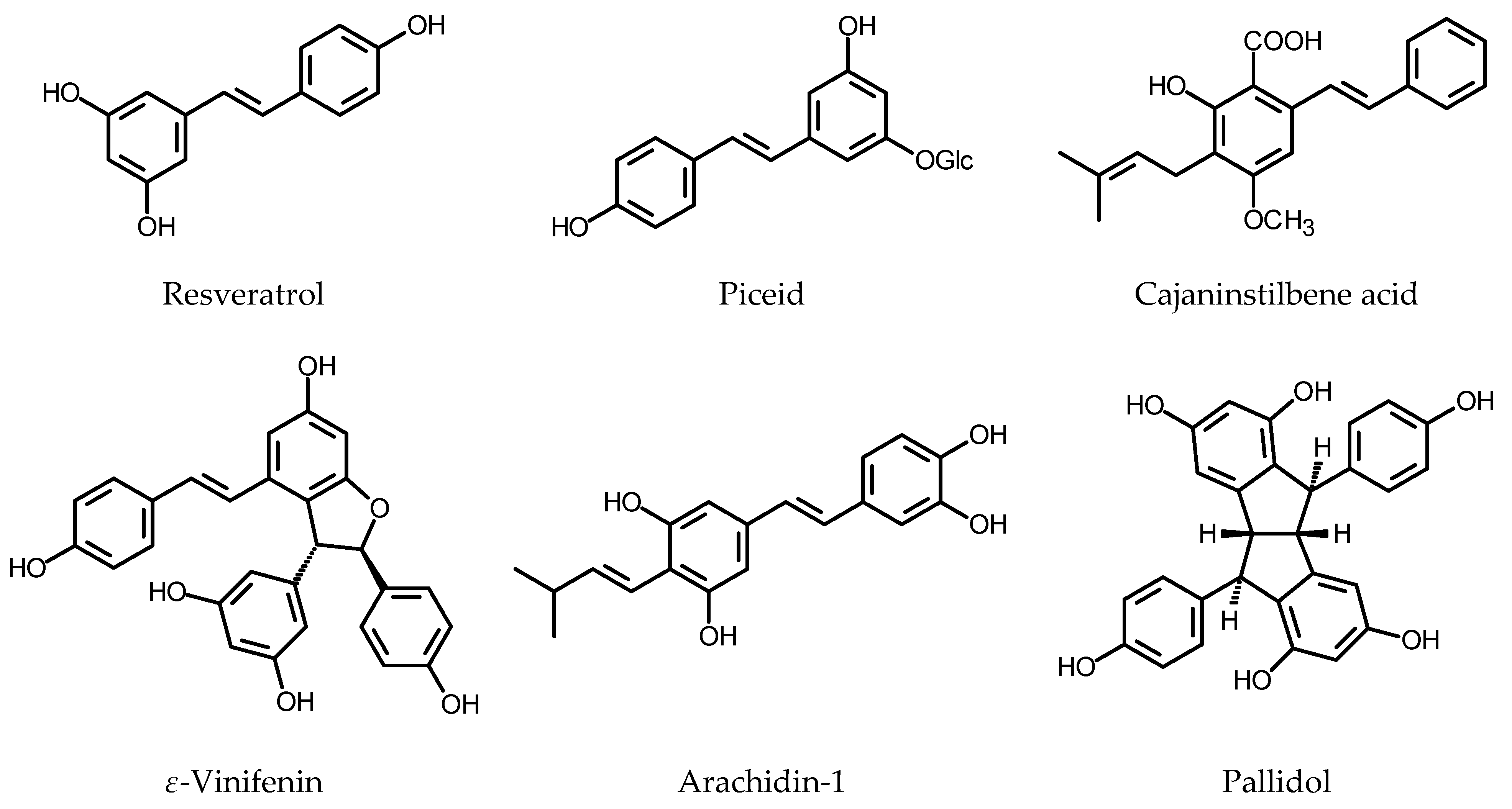{kind=link}
{kind=link}
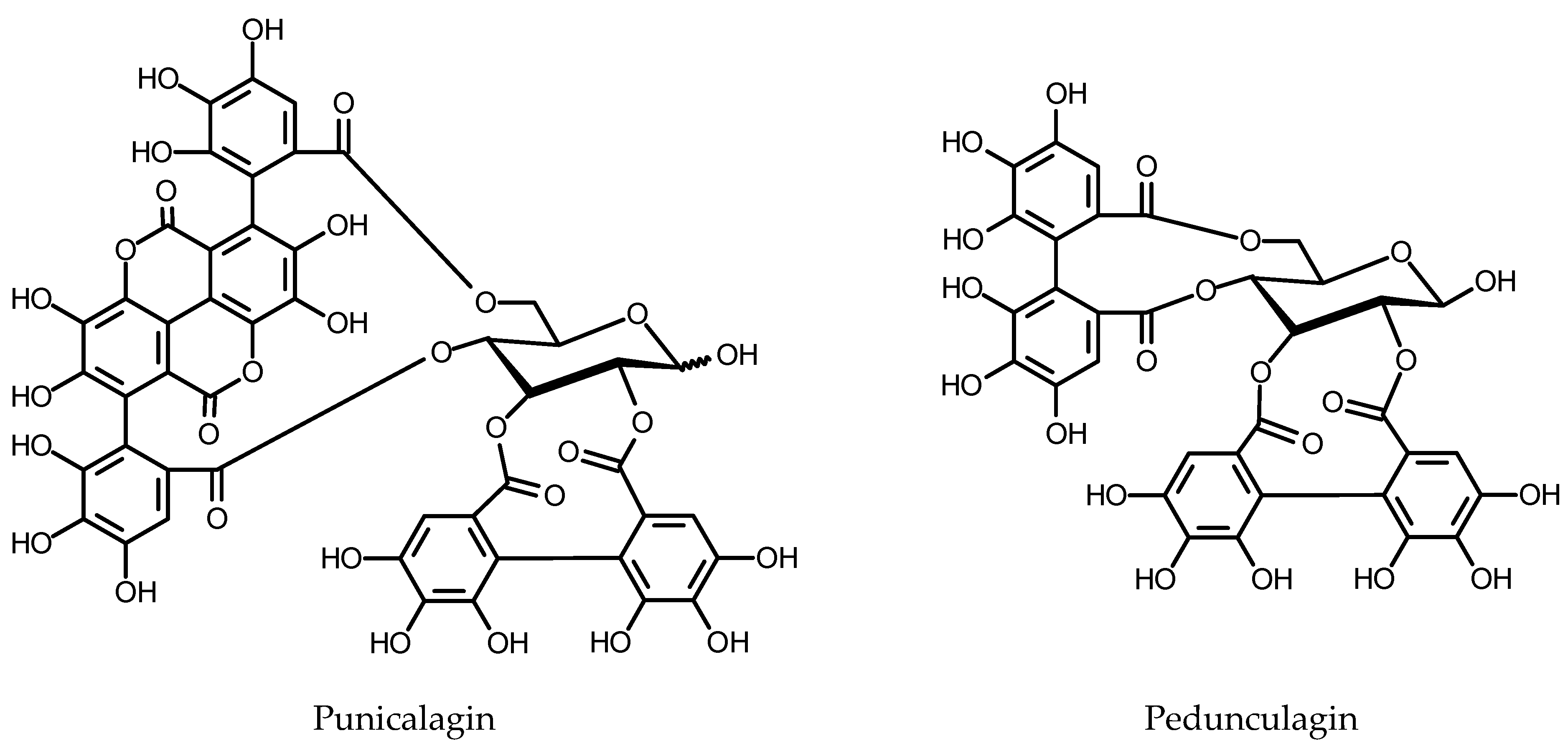{kind=link}
Table 1.
Hairy root cultures that produced high yields of antioxidative natural products.
| Plant Species | Hairy Roots | Natural Product | Maximum Content in the Biomass | Lit. |
|---|---|---|---|---|
| Fragaria x ananassa Duch cv. Reikou | Wild type | Procyanidin B-3 | 8 mg/g DW | [35] |
| Litchi chinensis Sonn. | Overexpressing LcMYB1 | Anthocyanins Proanthocyanidins | 3 mg/g FW 15 mg/g FW | [48] |
| Vitis vinifera L. | Overexpressing VvMybP1, VvMybP2 | Proanthocyanidins | 8 mg/g FW | [38] |
| Fagopyrum tataricum (L.) Moench. cv. Hokkai T10 | Wild type | Cyanidin 3-O-glucoside Cyanidin 3-O-rutinoside | 0.8 mg/g DW 2.4 mg/g DW | [52] |
| Daucus carota L. ssp. sativus var. atrorubens Alef | Wild type, treated with ethephon | Anthocyanins | 8 mg/g DW | [9] |
| Antirrhinum majus L. | Overexpressing AmRosea1 | Anthocyanins | 2 mg/g FW | [58] |
| Fagopyrum esculentum Moench | Wild type | (−)-Epicatechin 3-O-gallate (+)-Catechin | 10 mg/g DW 8 mg/g DW | [61] |
| Fagopyrum tataricum (L.) Moench cv. Hokkai T10 | Wild type | Rutin | 59 mg/g DW | [34] |
| Fagopyrum tataricum | Overexpressing FtUGT73BE5 | Rutin | Up to 80 mg/g DW | [65] |
| Polygonum multiflorum Thunb. | Wild type, elicited with MJ (50 µM) | Quercetin | Up to 14 mg/g DW | [74] |
| Isatis tinctoria L. | Wild type, exposed to UV-B | Rutin Quercetin Isorhamnetin Kaempferol | Up to 1.5 mg/g DW Up to 1.7 mg/g DW Up to 2.0 mg/g DW Up to 2.2 mg/g DW | [84] |
| Ligularia fischeri (Ledeb.) Turcz. | Wild type | Myricetin | 2.4 mg/g DW | [86] |
| Scutellaria baicalensis Georgi | Wild type | Baicalein derivatives Wogonin derivatives Oroxylin A derivatives | 68.7 mg/g DW 15.1 mg/g DW 11.9 mg/g DW | [96] |
| Scutellaria baicalensis Georgi | Overexpressing SbPAL | Baicalin Baicalein Wogonin | 136 mg/g DW 29 mg/g DW 6.7 mg/g DW | [102] |
| Scutellaria lateriflora L. | Wild type | Baicalin Wogonoside Wogonin | 14.5 mg/g DW 12.0 mg/g DW 11.5 mg/g DW | [111] |
| Saussurea medusa Maxim | Wild type | Jaceosidin | 6.1 mg/g DW | [126] |
| Saussurea involucrata Kar. et Kir. ex Maxim | Overexpressing SmCHI | Apigenin | 2.6 mg/g DW | [127] |
| Erigeron breviscapus (Vaniot) Hand.-Mazz. | Overexpressing EbCHI | Scutellarin | 2.2 mg/g DW | [128] |
| Glycine max (L.) Merr. | Overexpressing GmMYB205 | Daidzein derivative | 6.7 mg/g DW | [141] |
| Glycine max (L.) Merr. | Overexpressing GmMaT2 | Malonyldaidzin | 6.3 mg/g FW | [145] |
| Glycine max (L.) Merr. | Wild type, elicited with MeJa (100 µM) | Daidzin | 33.9 mg/g DW | [146] |
| Trifolium pratense L. | Wild type | Daidzein Genistein Formononetin Biochanin A | 8.6 mg/g DW 2.5 mg/g DW 15.2 mg/g DW 1.1 mg/g DW | [149] |
| Astragalus membranaceus (Fisch.) Bunge | Wild type, co-cultivated with the immobilized Aspergillus niger | Calycosin Formononetin | 0.7 mg/g DW 1.1 mg/g DW | [155] |
| Psoralea lachnostachys F. Muell | Wild type | Daidzein Coumestrol | 10.2 mg/g DW 0.5 mg/g DW | [158] |
| Psoralea corylifolia L. | Wild type | Daidzin | Over 10 mg/g DW | [159] |
| Pueraria candollei Wall. Ex Benth | Wild type | Daidzin Puerarin | Up to 29.9 mg/g DW Up to 3.4 mg/g DW | [164] |
| Ononis spinosa L. | Wild type | Medicarpin glucoside Sativanone glucoside Pseudobaptigenin glucoside | 22.3–28.9 mg/g DW 5.6–11.4 mg/g DW 1.2–2.0 mg/g DW | [170] |
| Arachis hypogaea L. cv. Hull | Wild type, elicited with MeJa (100 µM) and cyclodextrin (9 g/L) | Resveratrol Piceatannol Arachidin-1 Arachidin-3 | 5.3 mg/g DW 0.3 mg/g DW 4.0 mg/g DW (56 mg/L) 17.1 mg/g DW (148 mg/L) | [190] |
| Arachis hypogaea L. cv. Hull | Wild type, elicited with MeJa (125 µM), cyclodextrin (18 g/L), H2O2 (3 mM), and MgCl2 (1 mM) | Arachidin-1 Arachidin-2 Arachidin-3 Arachidin-5 | 227.4 mg/L 83.1 mg/L 370.6 mg/L 68.4 mg/L | [194] |
| Arachis hypogaea L. cv. Kalasin2 | Wild type, elicited with paraquat (500 µM), MeJa (100 µM) and cyclodextrin (6.87 mM) | Resveratrol Arachidin-1 Arachidin-3 | 1.3 mg/g DW 180.1 mg/g DW 444.2 mg/g DW | [197] |
| Arachis hypogaea L. cv. Tainan9 | Wild type, elicited with paraquat (500 µM), MeJa (100 µM) and cyclodextrin (6.87 mM) | Arachidin-1 Arachidin-3 | 1700 mg/L 4800 mg/L | [199] |
| Vitis rotundifolia Michx. | Wild type, elicited with MeJa (100 µM) | Piceid ε-Viniferin | 0.34 mg/g DW 0.38 mg/g DW | [203] |
| Vitis rotundifolia Michx | Wild type, elicited with H2O2 (10 mM) | ε-Viniferin | 0.43 mg/g DW | [203] |
| Vitis vinifera Pinot Noir cv. PN40024 | Wild type, elicited with MeJa (100 µM) | Total stilbenoids | 6.98 mg/g DW | [204] |
| Vitis vinifera Pinot Noir cv. PN40024 | Wild type, elicited with MeJa (100 µM) and cyclodextrin (50 mM) | Total stilbenoids | 165 mg/L (medium) 6.4 mg/g DW | [204] |
| Vitis vinifera subsp. sylvestris acc. W16 | Wild type | Resveratrol | 0.27 mg/g DW | [207] |
| Cajanus cajan (L.) Millsp. | Wild type, irradiated with UV-B | Cajaninstilbene acid | Up to 6.6 mg/g DW | [134] |
| Cajanus cajan (L.) Millsp. | Wild type, elicited with MeJa (125 µM), cyclodextrin (18 g/L), H2O2 (3 mM), and MgCl2 (1 mM) | Cajaninstilbene acid | Over 8 mg/g DW | [210] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Malarz, J.; Michalska, K.; Yudina, Y.V.; Stojakowska, A. Hairy Root Cultures as a Source of Polyphenolic Antioxidants: Flavonoids, Stilbenoids and Hydrolyzable Tannins. Plants 2022, 11, 1950. https://doi.org/10.3390/plants11151950
AMA Style
Malarz J, Michalska K, Yudina YV, Stojakowska A. Hairy Root Cultures as a Source of Polyphenolic Antioxidants: Flavonoids, Stilbenoids and Hydrolyzable Tannins. Plants. 2022; 11(15):1950. https://doi.org/10.3390/plants11151950
Chicago/Turabian StyleMalarz, Janusz, Klaudia Michalska, Yulia V. Yudina, and Anna Stojakowska. 2022. "Hairy Root Cultures as a Source of Polyphenolic Antioxidants: Flavonoids, Stilbenoids and Hydrolyzable Tannins" Plants 11, no. 15: 1950. https://doi.org/10.3390/plants11151950
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.