
The Promoter Analysis of VvPR1 Gene: A Candidate Gene Identified through Transcriptional Profiling of Methyl Jasmonate Treated Grapevine (Vitis vinifera L.)

 , ,
, ,
Abstract
:1. Introduction
2. Results
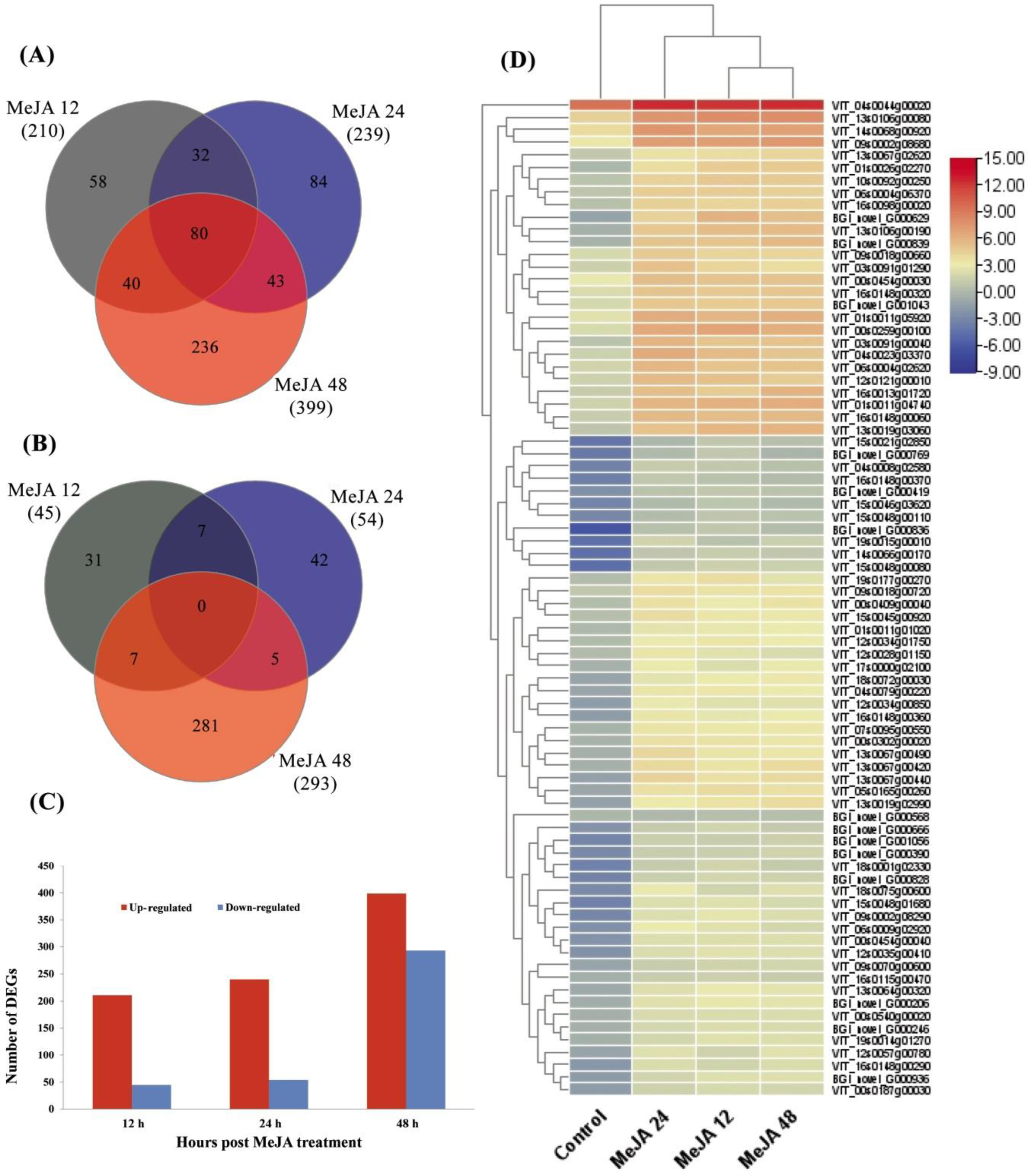
2.1. Transcriptomic Analysis of Grapevine Leaves in Response to MeJA at Different Time Points
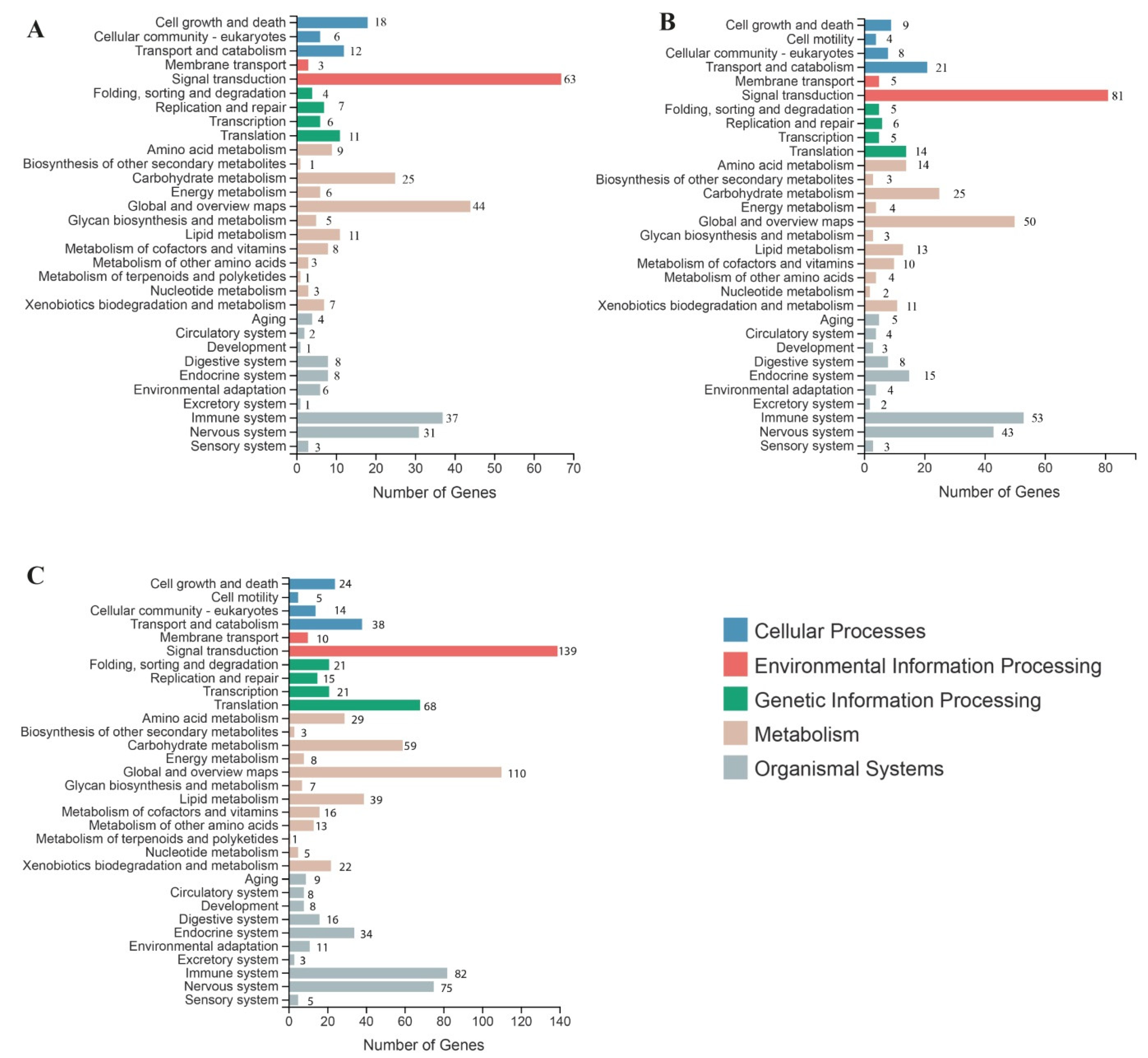
2.2. Gene Ontology (GO) Analysis of DEGs
2.3. Kyoto Encyclopedia of Genes and Genomes (KEGG) Analysis of DEGs
2.4. Transcript Levels of Genes Involved in JA Dependent Defense Pathway in Grapevine Leaves after MeJA Treatment
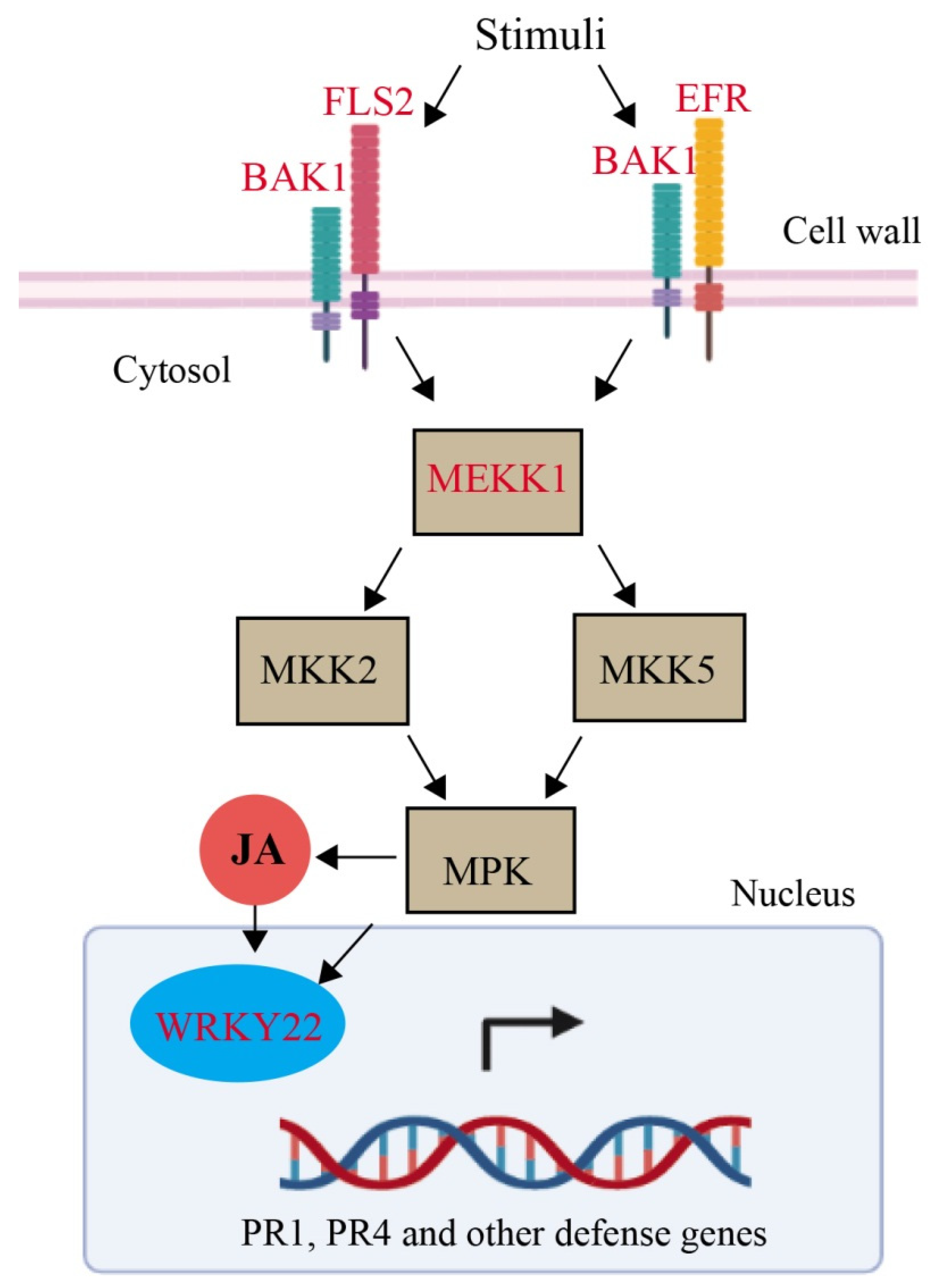
2.5. Mitogen-Activated Protein Kinase (MAPK) Pathway
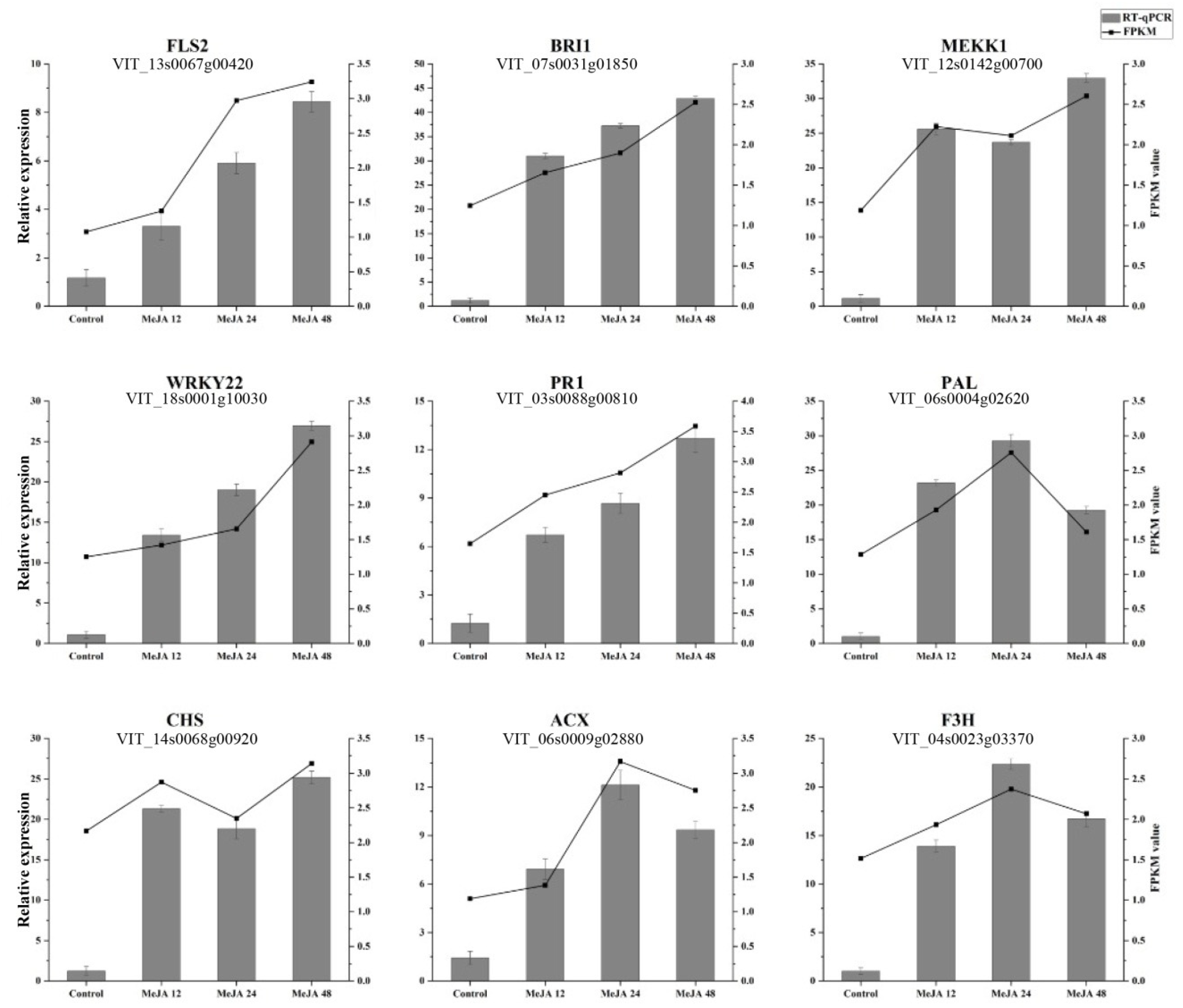
2.6. Verification of Differential Gene Expression
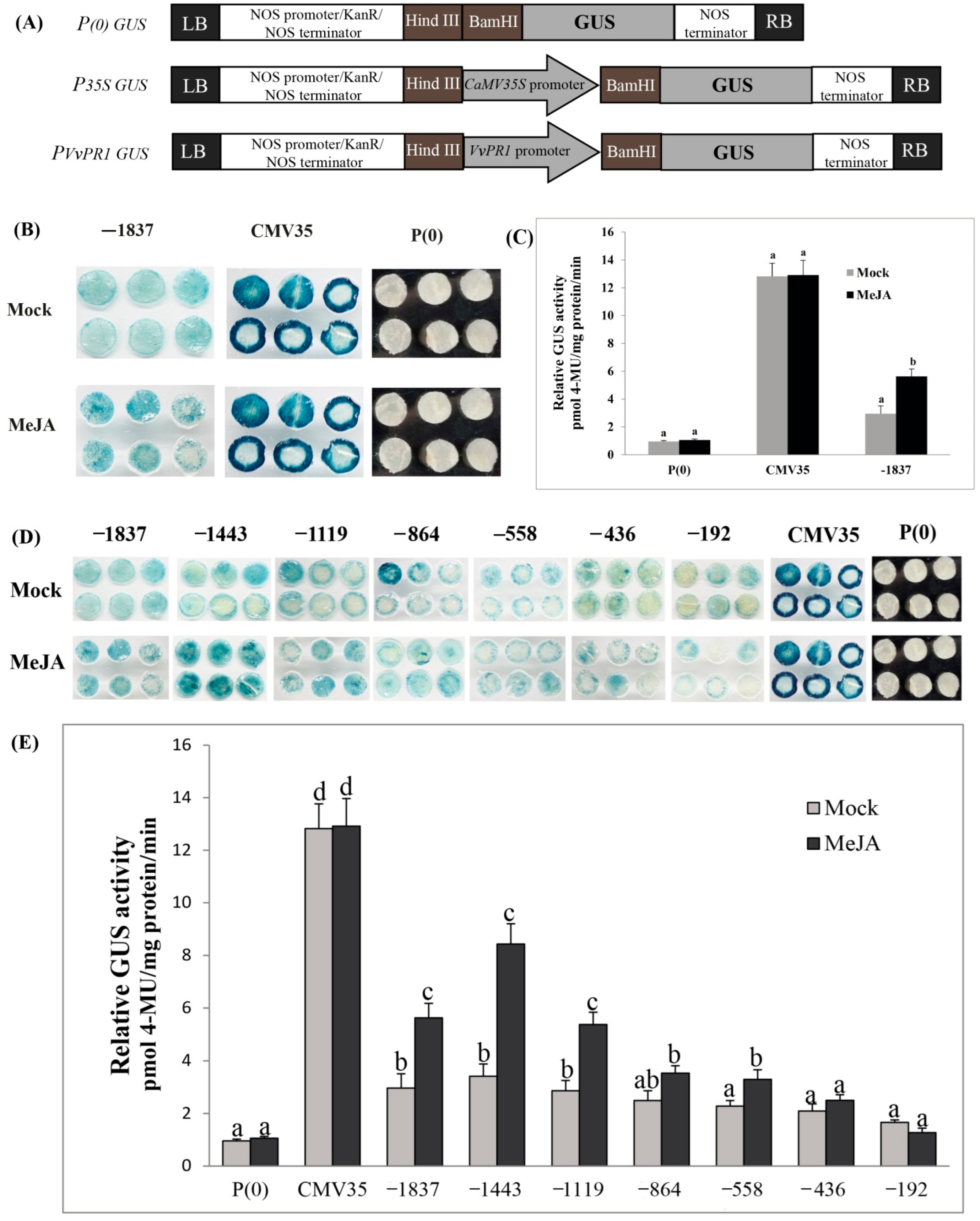
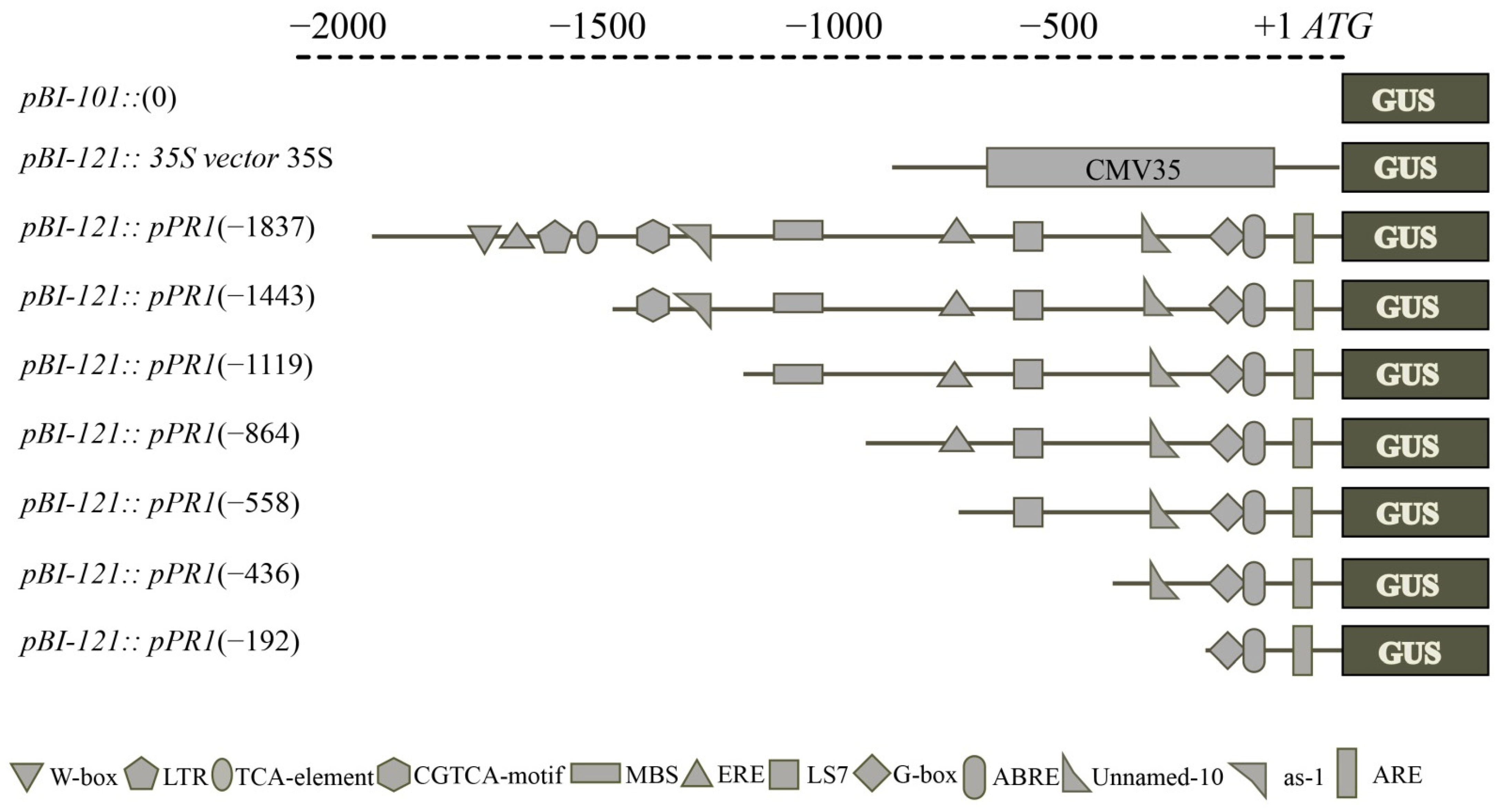
2.7. VvPR1 Promoter Isolation and Sequence Analysis
2.8. Deletion Analysis of the VvPR1 Promoter against MeJA Treatment
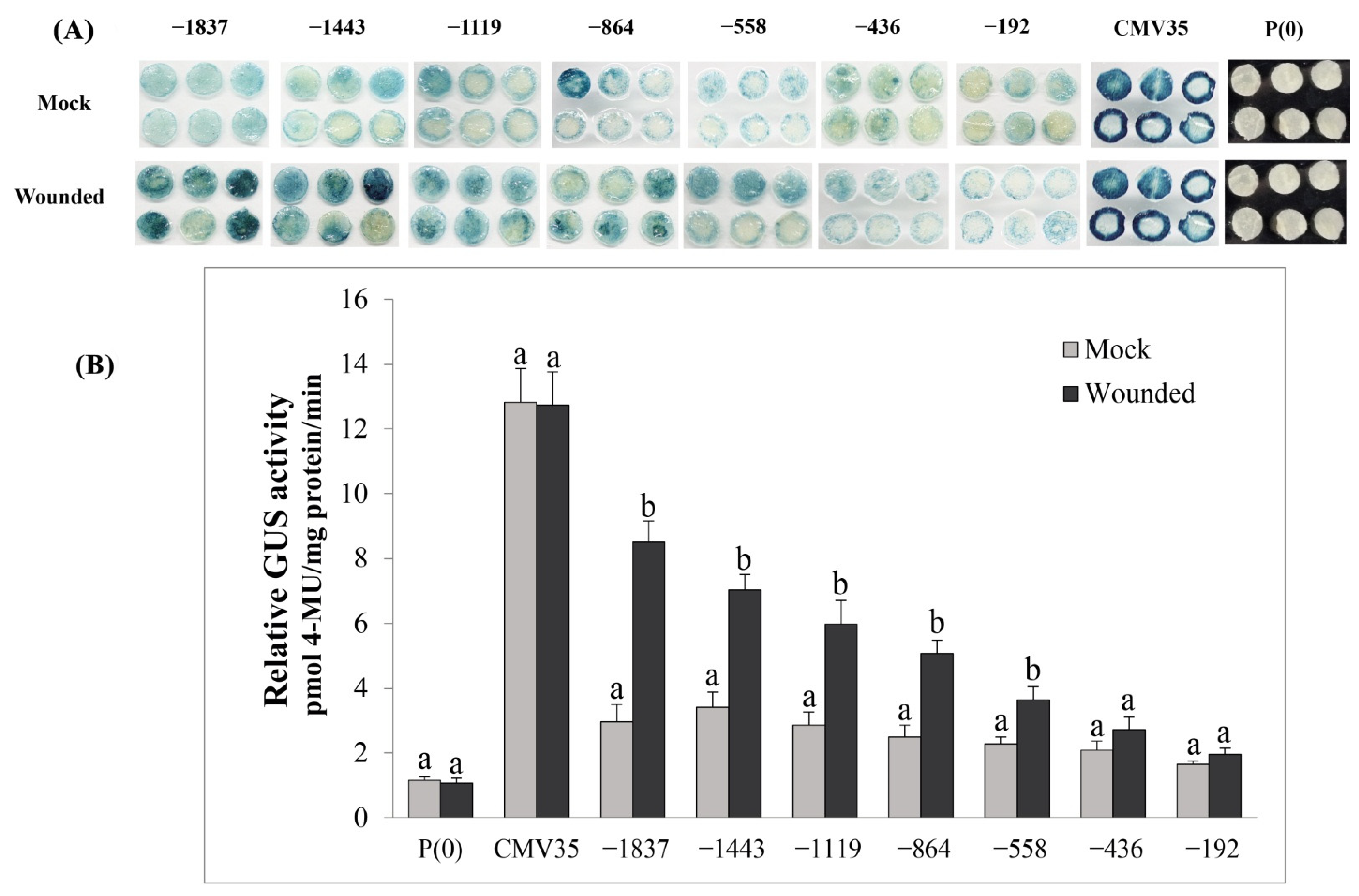
2.9. Deletion Analysis of the VvPR1 Promoter against Wound Stress
3. Discussion
4. Materials and Methods
4.1. Plant Material and Hormones Treatment
4.2. Total RNA Extraction
4.3. Library Construction of mRNA and Data Analysis
4.4. Quantitative Real-Time PCR (qRT-PCR)
4.5. Promoter Isolation of VvPR1 Gene and Sequence Analysis
4.6. Construction of Beta-Glucuronidase (GUS) Vectors
4.7. Agrobacterium-Mediated Transient Expression Assay with Abiotic Stress Treatment
4.8. GUS Activity Measurement
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Alston, J.M.; Sambucci, O. Grapes in the World Economy. In The Grape Genome; Springer: Berlin/Heidelberg, Germany, 2019; pp. 1–24. [Google Scholar]
- Ferreira, R.B.; Monteiro, S.S.; Piçarra-Pereira, M.A.; Teixeira, A.R. Engineering grapevine for increased resistance to fungal pathogens without compromising wine stability. Trends Biotechnol. 2004, 22, 168–173. [Google Scholar] [CrossRef] [PubMed]
- Pieterse, C.M.J.; Leon-Reyes, A.; Van der Ent, S.; Van Wees, S.C.M. Networking by small-molecule hormones in plant immunity. Nat. Chem. Biol. 2009, 5, 308–316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abramovitch, R.B.; Anderson, J.C.; Martin, G.B. Bacterial elicitation and evasion of plant innate immunity. Nat. Rev. Mol. Cell Biol. 2006, 7, 601–611. [Google Scholar] [CrossRef]
- Withers, J.; Dong, X. Posttranslational modifications of NPR1: A single protein playing multiple roles in plant immunity and physiology. PLoS Pathog. 2016, 12, e1005707. [Google Scholar] [CrossRef] [PubMed]
- Zarate, S.I.; Kempema, L.A.; Walling, L.L. Silverleaf whitefly induces salicylic acid defenses and suppresses effectual jasmonic acid defenses. Plant Physiol. 2007, 143, 866–875. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kawazu, K.; Mochizuki, A.; Sato, Y.; Sugeno, W.; Murata, M.; Seo, S.; Mitsuhara, I. Different expression profiles of jasmonic acid and salicylic acid inducible genes in the tomato plant against herbivores with various feeding modes. Arthropod-Plant Interact. 2012, 6, 221–230. [Google Scholar] [CrossRef]
- Glazebrook, J. Contrasting mechanisms of defense against biotrophic and necrotrophic pathogens. Annu. Rev. Phytopathol. 2005, 43, 205–227. [Google Scholar] [CrossRef]
- Wasternack, C.; Xie, D. The genuine ligand of a jasmonic acid receptor: Improved analysis of jasmonates is now required. Plant Signal. Behav. 2010, 5, 337–340. [Google Scholar] [CrossRef] [Green Version]
- Wasternack, C.; Strnad, M. Jasmonates: News on occurrence, biosynthesis, metabolism and action of an ancient group of signaling compounds. Int. J. Mol. Sci. 2018, 19, 2539. [Google Scholar] [CrossRef] [Green Version]
- Wasternack, C.; Parthier, B. Jasmonate-signalled plant gene expression. Trends Plant Sci. 1997, 2, 302–307. [Google Scholar] [CrossRef]
- Kępczyńska, E.; Król, P. The phytohormone methyl jasmonate as an activator of induced resistance against the necrotroph Alternaria porri f. sp. solani in tomato plants. J. Plant Interact. 2012, 7, 307–315. [Google Scholar] [CrossRef]
- Ton, J.; D’Alessandro, M.; Jourdie, V.; Jakab, G.; Karlen, D.; Held, M.; Mauch-Mani, B.; Turlings, T.C. Priming by airborne signals boosts direct and indirect resistance in maize. Plant J. 2007, 49, 16–26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.; Pang, Q.; Dai, S.; Wang, Y.; Chen, S.; Yan, X. Proteomic identification of differentially expressed proteins in Arabidopsis in response to methyl jasmonate. J. Plant Physiol. 2011, 168, 995–1008. [Google Scholar] [CrossRef]
- Desmond, O.J.; Edgar, C.I.; Manners, J.M.; Maclean, D.J.; Schenk, P.M.; Kazan, K. Methyl jasmonate induced gene expression in wheat delays symptom development by the crown rot pathogen Fusarium pseudograminearum. Physiol. Mol. Plant Pathol. 2005, 67, 171–179. [Google Scholar] [CrossRef]
- Howe, G.A.; Major, I.T.; Koo, A.J. Modularity in jasmonate signaling for multistress resilience. Annu. Rev. Plant Biol. 2018, 69, 387–415. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pauwels, L.; Goossens, A. The JAZ proteins: A crucial interface in the jasmonate signaling cascade. Plant Cell 2011, 23, 3089–3100. [Google Scholar] [CrossRef] [Green Version]
- Kloth, K.J.; Wiegers, G.L.; Busscher-Lange, J.; Van Haarst, J.C.; Kruijer, W.; Bouwmeester, H.J.; Dicke, M.; Jongsma, M.A. AtWRKY22 promotes susceptibility to aphids and modulates salicylic acid and jasmonic acid signalling. J. Exp. Bot. 2016, 67, 3383–3396. [Google Scholar] [CrossRef] [Green Version]
- Jiang, Y.; Liang, G.; Yang, S.; Yu, D. Arabidopsis WRKY57 Functions as a node of convergence for jasmonic acid– and auxin-mediated signaling in jasmonic acid–induced leaf senescence. Plant Cell 2014, 26, 230–245. [Google Scholar] [CrossRef] [Green Version]
- Gao, Q.-M.; Venugopal, S.; Navarre, D.; Kachroo, A. Low oleic acid-derived repression of jasmonic acid-inducible defense responses requires the WRKY50 and WRKY51 proteins. Plant Physiol. 2011, 155, 464–476. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Zhong, R.; Palva, E.T. WRKY70 and its homolog WRKY54 negatively modulate the cell wall-associated defenses to necrotrophic pathogens in Arabidopsis. PLoS ONE 2017, 12, e0183731. [Google Scholar] [CrossRef]
- Lowe, R.; Shirley, N.; Bleackley, M.; Dolan, S.; Shafee, T. Transcriptomics technologies. PLoS Comput. Biol. 2017, 13, e1005457. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chu, Y.; Corey, D.R. RNA sequencing: Platform selection, experimental design, and data interpretation. Nucleic Acid Ther. 2012, 22, 271–274. [Google Scholar] [CrossRef] [PubMed]
- González-Candelas, L.; Alamar, S.; Sánchez-Torres, P.; Zacarías, L.; Marcos, J.F. A transcriptomic approach highlights induction of secondary metabolism in citrus fruit in response to Penicillium digitatum infection. BMC Plant Biol. 2010, 10, 194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, M.; Yang, Q.; Boateng, N.A.S.; Ahima, J.; Dou, Y.; Zhang, H. Ultrastructure observation and transcriptome analysis of Penicillium expansum invasion in postharvest pears. Postharvest Biol. Technol. 2020, 165, 111198. [Google Scholar] [CrossRef]
- Ayala-Zavala, J.F.; Wang, S.Y.; Wang, C.Y.; González-Aguilar, G.A. Methyl jasmonate in conjunction with ethanol treatment increases antioxidant capacity, volatile compounds and postharvest life of strawberry fruit. Eur. Food Res. Technol. 2005, 221, 731–738. [Google Scholar] [CrossRef]
- Meng, X.; Han, J.; Wang, Q.; Tian, S. Changes in physiology and quality of peach fruits treated by methyl jasmonate under low temperature stress. Food Chem. 2009, 114, 1028–1035. [Google Scholar] [CrossRef]
- Kondo, S.; Setha, S.; Rudell, D.; Buchanan, D.; Mattheis, J. Aroma volatile biosynthesis in apples affected by 1-MCP and methyl jasmonate. Postharvest Biol. Technol. 2005, 36, 61–68. [Google Scholar] [CrossRef]
- Lalel, H.J.D.; Singh, Z.; Tan, S.C. The role of methyl jasmonate in mango ripening and biosynthesis of aroma volatile compounds. J. Hortic. Sci. Biotechnol. 2003, 78, 470–484. [Google Scholar] [CrossRef]
- Wang, K.; Liao, Y.; Kan, J.; Han, L.; Zheng, Y. Response of direct or priming defense against Botrytis cinerea to methyl jasmonate treatment at different concentrations in grape berries. Int. J. Food Microbiol. 2015, 194, 32–39. [Google Scholar] [CrossRef]
- Conconi, A.; Miquel, M.; Browse, J.A.; Ryan, C.A. Intracellular levels of free linolenic and linoleic acids increase in tomato leaves in response to wounding. Plant Physiol. 1996, 111, 797–803. [Google Scholar] [CrossRef] [Green Version]
- Maucher, H.; Hause, B.; Feussner, I.; Ziegler, J.; Wasternack, C. Allene oxide synthases of barley (Hordeum vulgare cv. Salome): Tissue specific regulation in seedling development. Plant J. 2000, 21, 199–213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, H.; Jones, A.D.; Howe, G.A. Constitutive activation of the jasmonate signaling pathway enhances the production of secondary metabolites in tomato. FEBS Lett. 2006, 580, 2540–2546. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kazan, K.; Manners, J.M. Jasmonate signaling: Toward an integrated view. Plant Physiol. 2008, 146, 1459–1468. [Google Scholar] [CrossRef] [Green Version]
- Chatel, G.; Montiel, G.; Pré, M.; Memelink, J.; Thiersault, M.; Saint-Pierre, B.; Doireau, P.; Gantet, P. CrMYC1, a Catharanthus roseus elicitor- and jasmonate-responsive bHLH transcription factor that binds the G-box element of the strictosidine synthase gene promoter. J. Exp. Bot. 2003, 54, 2587–2588. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, Z.-X.; Li, J.-X.; Yang, C.-Q.; Hu, W.-L.; Wang, L.-J.; Chen, X.-Y. The jasmonate-responsive AP2/ERF transcription factors AaERF1 and AaERF2 positively regulate artemisinin biosynthesis in Artemisia annua L. Mol. Plant 2012, 5, 353–365. [Google Scholar] [CrossRef] [Green Version]
- Schluttenhofer, C.; Pattanaik, S.; Patra, B.; Yuan, L. Analyses of Catharanthus roseus and Arabidopsis thaliana WRKY transcription factors reveal involvement in jasmonate signaling. BMC Genom. 2014, 15, 502. [Google Scholar] [CrossRef] [Green Version]
- Zhao, J.; Davis, L.C.; Verpoorte, R. Elicitor signal transduction leading to production of plant secondary metabolites. Biotechnol. Adv. 2005, 23, 283–333. [Google Scholar] [CrossRef]
- Van Moerkercke, A.; Galván-Ampudia, C.S.; Verdonk, J.C.; Haring, M.A.; Schuurink, R.C. Regulators of floral fragrance production and their target genes in petunia are not exclusively active in the epidermal cells of petals. J. Exp. Bot. 2012, 63, 3157–3171. [Google Scholar] [CrossRef] [Green Version]
- Falcone Ferreyra, M.L.; Rius, S.; Casati, P. Flavonoids: Biosynthesis, biological functions, and biotechnological applications. Front. Plant Sci. 2012, 3, 222. [Google Scholar] [CrossRef] [Green Version]
- Romeis, T.; Piedras, P.; Zhang, S.; Klessig, D.F.; Hirt, H.; Jones, J.D. Rapid Avr9-and Cf-9–dependent activation of MAP kinases in tobacco cell cultures and leaves: Convergence of resistance gene, elicitor, wound, and salicylate responses. Plant Cell 1999, 11, 273–287. [Google Scholar]
- Ligterink, W.; Kroj, T.; Zur Nieden, U.; Hirt, H.; Scheel, D. Receptor-mediated activation of a MAP kinase in pathogen defense of plants. Science 1997, 276, 2054–2057. [Google Scholar] [CrossRef]
- Zhang, S.; Du, H.; Klessig, D.F. Activation of the tobacco SIP kinase by both a cell wall–derived carbohydrate elicitor and purified proteinaceous elicitins from Phytophthora spp. Plant Cell 1998, 10, 435–449. [Google Scholar] [PubMed] [Green Version]
- Zhang, S.; Klessig, D.F. Resistance gene N-mediated de novo synthesis and activation of a tobacco mitogen-activated protein kinase by tobacco mosaic virus infection. Proc. Natl. Acad. Sci. USA 1998, 95, 7433–7438. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nühse, T.S.; Peck, S.C.; Hirt, H.; Boller, T. Microbial elicitors induce activation and dual phosphorylation of the Arabidopsis thaliana MAPK 6. J. Biol. Chem. 2000, 275, 7521–7526. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.; Klessig, D.F.; Nürnberger, T. A harpin binding site in tobacco plasma membranes mediates activation of the pathogenesis-related gene HIN1 independent of extracellular calcium but dependent on mitogen-activated protein kinase activity. Plant Cell 2001, 13, 1079–1093. [Google Scholar] [CrossRef] [Green Version]
- Kemmerling, B.; Schwedt, A.; Rodriguez, P.A.; Mazzotta, S.; Frank, M.; Qamar, S.A.; Mengiste, T.; Betsuyaku, S.; Parker, J.E.; Müssig, C.; et al. The BRI1-Associated Kinase 1, BAK1, Has a Brassinolide-Independent Role in Plant Cell-Death Control. Curr. Biol. 2007, 17, 1116–1122. [Google Scholar] [CrossRef] [Green Version]
- He, K.; Gou, X.; Yuan, T.; Lin, H.; Asami, T.; Yoshida, S.; Russell, S.D.; Li, J. BAK1 and bkk1 regulate brassinosteroid-dependent growth and brassinosteroid-independent cell-death pathways. Curr. Biol. 2007, 17, 1109–1115. [Google Scholar] [CrossRef] [Green Version]
- Vert, G. Plant signaling: Brassinosteroids, immunity and effectors are BAK! Curr. Biol. 2008, 18, R963–R965. [Google Scholar] [CrossRef]
- Caño-Delgado, A.; Yin, Y.; Yu, C.; Vafeados, D.; Mora-García, S.; Cheng, J.-C.; Nam, K.H.; Li, J.; Chory, J. BRL1 and BRL3 are novel brassinosteroid receptors that function in vascular differentiation in Arabidopsis. Development 2004, 131, 5341–5351. [Google Scholar] [CrossRef] [Green Version]
- Karlova, R.; Boeren, S.; Russinova, E.; Aker, J.; Vervoort, J.; de Vries, S. The Arabidopsis somatic embryogenesis receptor-like kinase1 protein complex includes brassinosteroid-insensitive1. Plant Cell 2006, 18, 626–638. [Google Scholar] [CrossRef] [Green Version]
- Kinoshita, T.; Caño-delgado, A.I.; Seto, H.; Hiranuma, S.; Fujioka, S.; Yoshida, S.; Chory, J. Binding of brassinosteroids to the extracellular domain of plant receptor kinase BRI1. Nature 2005, 433, 167–171. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Li, X.; Meisenhelder, J.; Hunter, T.; Yoshida, S.; Asami, T.; Chory, J. Autoregulation and homodimerization are involved in the activation of the plant steroid receptor BRI1. Dev. Cell 2005, 8, 855–865. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Goshe, M.B.; Soderblom, E.J.; Phinney, B.S.; Kuchar, J.A.; Li, J.; Asami, T.; Yoshida, S.; Huber, S.C.; Clouse, S.D. Identification and Functional analysis of in vivo phosphorylation sites of the Arabidopsis brassinosteroid-insensitive1 receptor kinase. Plant Cell 2005, 17, 1685–1703. [Google Scholar] [CrossRef] [Green Version]
- Chinchilla, D.; Zipfel, C.; Robatzek, S.; Kemmerling, B.; Nürnberger, T.; Jones, J.D.G.; Felix, G.; Boller, T. A flagellin-induced complex of the receptor FLS2 and BAK1 initiates plant defence. Nature 2007, 448, 497–500. [Google Scholar] [CrossRef] [PubMed]
- Heese, A.; Hann, D.R.; Gimenez-Ibanez, S.; Jones, A.M.; He, K.; Li, J.; Schroeder, J.I.; Peck, S.C.; Rathjen, J.P. The receptor-like kinase SERK3/BAK1 is a central regulator of innate immunity in plants. Proc. Natl. Acad. Sci. USA 2007, 104, 12217–12222. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Chory, J. Brassinosteroids regulate dissociation of BKI1, a negative regulator of bri1 signaling, from the plasma membrane. Science 2006, 313, 1118–1122. [Google Scholar] [CrossRef]
- Hink, M.A.; Shah, K.; Russinova, E.; de Vries, S.C.; Visser, A.J. Fluorescence fluctuation analysis of Arabidopsis thaliana somatic embryogenesis receptor-like kinase and brassinosteroid insensitive 1 receptor oligomerization. Biophys. J. 2008, 94, 1052–1062. [Google Scholar] [CrossRef] [Green Version]
- Mao, G.; Meng, X.; Liu, Y.; Zheng, Z.; Chen, Z.; Zhang, S. Phosphorylation of a WRKY transcription factor by two pathogen-responsive MAPKS drives phytoalexin biosynthesis in Arabidopsis. Plant Cell 2011, 23, 1639–1653. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Zhang, S. Phosphorylation of 1-aminocyclopropane-1-carboxylic acid synthase by MPK6, a stress-responsive mitogen-activated protein kinase, induces ethylene biosynthesis in Arabidopsis. Plant Cell 2004, 16, 3386–3399. [Google Scholar] [CrossRef] [Green Version]
- Zheng, Z.; Qamar, S.A.; Chen, Z.; Mengiste, T. Arabidopsis WRKY33 transcription factor is required for resistance to necrotrophic fungal pathogens. Plant J. 2006, 48, 592–605. [Google Scholar] [CrossRef]
- Adachi, H.; Nakano, T.; Miyagawa, N.; Ishihama, N.; Yoshioka, M.; Katou, Y.; Yaeno, T.; Shirasu, K.; Yoshioka, H. WRKY transcription factors phosphorylated by MAPK regulate a plant immune NADPH oxidase in Nicotiana benthamiana. Plant Cell 2015, 27, 2645–2663. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hussain, R.M.F.; Sheikh, A.H.; Haider, I.; Quareshy, M.; Linthorst, H.J.M. Arabidopsis WRKY50 and TGA transcription factors synergistically activate expression of PR1. Front. Plant Sci. 2018, 9, 930. [Google Scholar] [CrossRef] [PubMed]
- Tirosh, I.; Weinberger, A.; Carmi, M.; Barkai, N. A genetic signature of interspecies variations in gene expression. Nat. Genet. 2006, 38, 830–834. [Google Scholar] [CrossRef] [PubMed]
- Walther, D.; Brunnemann, R.; Selbig, J. The regulatory code for transcriptional response diversity and its relation to genome structural properties in A. thaliana. PLoS Genet. 2007, 3, e11. [Google Scholar] [CrossRef]
- Newman, J.R.S.; Ghaemmaghami, S.; Ihmels, J.; Breslow, D.K.; Noble, M.; DeRisi, J.L.; Weissman, J.S. Single-cell proteomic analysis of S. cerevisiae reveals the architecture of biological noise. Nature 2006, 441, 840–846. [Google Scholar] [CrossRef]
- Roelofs, D.; Morgan, J.; Stürzenbaum, S. The significance of genome-wide transcriptional regulation in the evolution of stress tolerance. Evol. Ecol. 2010, 24, 527–539. [Google Scholar] [CrossRef] [Green Version]
- Vos, I.A.; Emoritz, L.; Pieterse, C.M.; Van Wees, S.C.M. Impact of hormonal crosstalk on plant resistance and fitness under multi-attacker conditions. Front. Plant Sci. 2015, 6, 639. [Google Scholar] [CrossRef]
- Hong, J.K.; Lee, S.C.; Hwang, B.K. Activation of pepper basic PR-1 gene promoter during defense signaling to pathogen, abiotic and environmental stresses. Gene 2005, 356, 169–180. [Google Scholar] [CrossRef]
- Kesarwani, M.; Yoo, J.; Dong, X. Genetic Interactions of TGA transcription factors in the regulation of pathogenesis-related genes and disease resistance in Arabidopsis. Plant Physiol. 2007, 144, 336–346. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Liu, G.-J.; Yan, X.-F.; Wei, Z.-G.; Xu, Z.-R. MeJA-inducible expression of the heterologous JAZ2 promoter from Arabidopsis in Populus trichocarpa protoplasts. J. Plant Dis. Prot. 2011, 118, 69–74. [Google Scholar] [CrossRef]
- Hou, L.; Chen, L.; Wang, J.; Xu, D.; Dai, L.; Zhang, H.; Zhao, Y. Construction of stress responsive synthetic promoters and analysis of their activity in transgenic Arabidopsis thaliana. Plant Mol. Biol. Rep. 2012, 30, 1496–1506. [Google Scholar] [CrossRef]
- Kim, D.; Langmead, B.; Salzberg, S.L. HISAT: A fast spliced aligner with low memory requirements. Nat. Methods 2015, 12, 357–360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Langmead, B.; Salzberg, S.L. Fast gapped-read alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, B.; Dewey, C.N. RSEM: Accurate transcript quantification from RNA-Seq data with or without a reference genome. BMC Bioinform. 2011, 12, 323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Lescot, M.; Déhais, P.; Thijs, G.; Marchal, K.; Moreau, Y.; Van de Peer, Y.; Rouzé, P.; Rombauts, S. PlantCARE, a database of plant cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences. Nucleic Acids Res. 2002, 30, 325–327. [Google Scholar] [CrossRef]
- Yang, Y.; Li, R.; Qi, M. In vivo analysis of plant promoters and transcription factors by agroinfiltration of tobacco leaves. Plant J. 2000, 22, 543–551. [Google Scholar] [CrossRef]
- Jefferson, R.A.; Kavanagh, T.A.; Bevan, M.W. GUS fusions: Beta-glucuronidase as a sensitive and versatile gene fusion marker in higher plants. EMBO J. 1987, 6, 3901–3907. [Google Scholar] [CrossRef]
- Yu, Y.; Xu, W.; Wang, J.; Wang, L.; Yao, W.; Xu, Y.; Ding, J.; Wang, Y. A core functional region of the RFP1 promoter from Chinese wild grapevine is activated by powdery mildew pathogen and heat stress. Planta 2013, 237, 293–303. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Samples | Total Raw Reads (M) | Total Clean Reads (M) | Clean Reads Q20 (%) | Clean Reads Ratio (%) | Total Mapping (%) | DEGs |
|---|---|---|---|---|---|---|
| CR | 44.40 | 42.46 | 97.07 | 95.63 | 88.17 | |
| MeJA12 | 43.82 | 42.59 | 97.13 | 97.18 | 89.01 | 256 |
| MeJA24 | 43.82 | 42.56 | 97.16 | 97.12 | 87.39 | 294 |
| MeJA48 | 43.82 | 42.39 | 97.14 | 96.74 | 86.62 | 692 |
| Total | 175.86 | 170.00 | 388.50 | 386.67 | 351.19 | 1242 |
| Name | MeJA 12 | MeJA 24 | MeJA 48 | ||||
|---|---|---|---|---|---|---|---|
| Down | UP | Down | UP | Down | UP | ||
| Biological Process | Biological regulation | 28 | 5 | 35 | 42 | 58 | |
| Cellular process | 68 | 14 | 83 | 130 | 225 | ||
| Developmental process | 1 | 9 | 2 | 10 | 7 | 11 | |
| Growth | 0 | 2 | |||||
| Immune system process | 0 | 4 | 0 | 4 | 4 | 9 | |
| Interspecies interaction between organisms | 0 | 5 | 0 | 5 | 1 | 10 | |
| Localization | 7 | 14 | 5 | 13 | 24 | 32 | |
| Locomotion | 0 | 3 | 1 | 3 | 9 | 8 | |
| Metabolic process | 14 | 61 | 15 | 81 | 112 | 107 | |
| Multi-organism process | 0 | 2 | 0 | 2 | 8 | 3 | |
| Multicellular organismal process | 1 | 11 | 1 | 11 | 10 | 13 | |
| Reproduction | 0 | 2 | 0 | 2 | 8 | 6 | |
| Reproductive process | 0 | 2 | 0 | 2 | 1 | 13 | |
| Response to stimulus | 3 | 33 | 9 | 38 | 27 | 57 | |
| Rhythmic process | 0 | 1 | |||||
| Signaling | 0 | 18 | 1 | 21 | 18 | 34 | |
| Cellular Component | Cellular anatomical entity | 32 | 131 | 194 | 225 | ||
| Intracellular | 8 | 56 | 112 | 111 | |||
| Protein-containing complex | 0 | 8 | 41 | 28 | |||
| Molecular Function | Antioxidant activity | 0 | 5 | 0 | 4 | ||
| Binding | 14 | 119 | 113 | 202 | |||
| Catalytic activity | 25 | 127 | 128 | 193 | |||
| Molecular function regulator | 0 | 5 | 1 | 10 | |||
| Molecular transducer activity | 0 | 1 | |||||
| Protein-folding chaperone | 0 | 1 | |||||
| Structural molecule activity | 0 | 3 | 35 | 5 | |||
| Transcription regulator activity | 0 | 5 | 6 | 2 | |||
| Translation regulator activity | 2 | 2 | |||||
| Transporter activity | 5 | 10 | 16 | 16 | |||
| Gene Description | Gene ID | Log2 Fold Change | ||
|---|---|---|---|---|
| MeJA 12 | MeJA 24 | MeJA 48 | ||
| PLD | VIT_05s0020g00200 | 1.37 | 1.13 | 1.57 |
| LOX | VIT_01s0010g02750 | 0.53 | 0.27 | 0.88 |
| LOX | VIT_13s0064g01480 | 2.25 | 1.90 | 3.11 |
| ACX | VIT_06s0009g02970 | 1.57 | 2.39 | 1.52 |
| ACX | VIT_06s0009g02880 | 1.98 | 3.35 | 2.88 |
| PAL | VIT_06s0004g02620 | 2.46 | 3.27 | 2.98 |
| CHS | VIT_05s0136g00260 | 1.70 | 2.29 | 2.19 |
| CHS | VIT_14s0068g00920 | 2.16 | 2.99 | 2.77 |
| F3H | VIT_04s0023g03370 | 3.01 | 3.95 | 3.20 |
| F3H | VIT_18s0001g14310 | 1.03 | 1.63 | 1.52 |
| MYB | VIT_15s0046g00170 | 1.56 | 2.04 | 2.59 |
| MYB | VIT_18s0001g09850 | 0.12 | 0.82 | 1.11 |
| WRKY2 | VIT_01s0011g00220 | 0.21 | 0.27 | 0.89 |
| WRKY22 | VIT_18s0001g10030 | 0.41 | 0.62 | 0.98 |
| PR1 | VIT_03s0088g00810 | 1.64 | 1.38 | 2.49 |
| PR4 | VIT_03s0088g00780 | 0.80 | 0.90 | 1.12 |
| bHLH | VIT_02s0025g03220 | 0.40 | 0.40 | 1.20 |
| Gene Description | Gene ID | Log2-fold Change | ||
|---|---|---|---|---|
| MeJA 12 | MeJA 24 | MeJA 48 | ||
| FLS2 | VIT_09s0002g07680 | 0.58 | 1.37 | 1.70 |
| FLS2 | VIT_09s0002g07750 | 1.32 | 2.15 | 2.29 |
| FLS2 | VIT_09s0018g00830 | 0.97 | 1.04 | 2.89 |
| FLS2 | VIT_09s0054g00100 | 0.81 | 2.05 | 1.99 |
| FLS2 | VIT_09s0070g00620 | 0.25 | 1.02 | 1.07 |
| FLS2 | VIT_11s0118g00160 | 1.50 | 2.37 | 2.02 |
| FLS2 | VIT_13s0067g00420 | 3.01 | 3.40 | 3.21 |
| EFR | VIT_08s0007g00850 | 2.61 | 2.16 | 2.89 |
| EFR | VIT_18s0166g00050 | 2.41 | 2.60 | 3.39 |
| BAK1 | BGI_novel_G000939 | 2.54 | 4.98 | 4.72 |
| BRI1 | VIT_07s0031g01850 | 0.67 | 0.60 | 0.94 |
| MEKK1 | VIT_12s0142g00700 | 3.54 | 2.06 | 3.99 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rahman, F.U.; Zhang, Y.; Khan, I.A.; Liu, R.; Sun, L.; Wu, Y.; Jiang, J.; Fan, X.; Liu, C. The Promoter Analysis of VvPR1 Gene: A Candidate Gene Identified through Transcriptional Profiling of Methyl Jasmonate Treated Grapevine (Vitis vinifera L.). Plants 2022, 11, 1540. https://doi.org/10.3390/plants11121540
Rahman FU, Zhang Y, Khan IA, Liu R, Sun L, Wu Y, Jiang J, Fan X, Liu C. The Promoter Analysis of VvPR1 Gene: A Candidate Gene Identified through Transcriptional Profiling of Methyl Jasmonate Treated Grapevine (Vitis vinifera L.). Plants. 2022; 11(12):1540. https://doi.org/10.3390/plants11121540
Chicago/Turabian StyleRahman, Faiz Ur, Ying Zhang, Irshad Ahmad Khan, Ruitao Liu, Lei Sun, Yandi Wu, Jianfu Jiang, Xiucai Fan, and Chonghuai Liu. 2022. "The Promoter Analysis of VvPR1 Gene: A Candidate Gene Identified through Transcriptional Profiling of Methyl Jasmonate Treated Grapevine (Vitis vinifera L.)" Plants 11, no. 12: 1540. https://doi.org/10.3390/plants11121540