
Preconditioning to Water Deficit Helps Aloe vera to Overcome Long-Term Drought during the Driest Season of Atacama Desert


Abstract
:1. Introduction
2. Results
2.1. Leaf Biomass and Biochemical Indicators of Stress
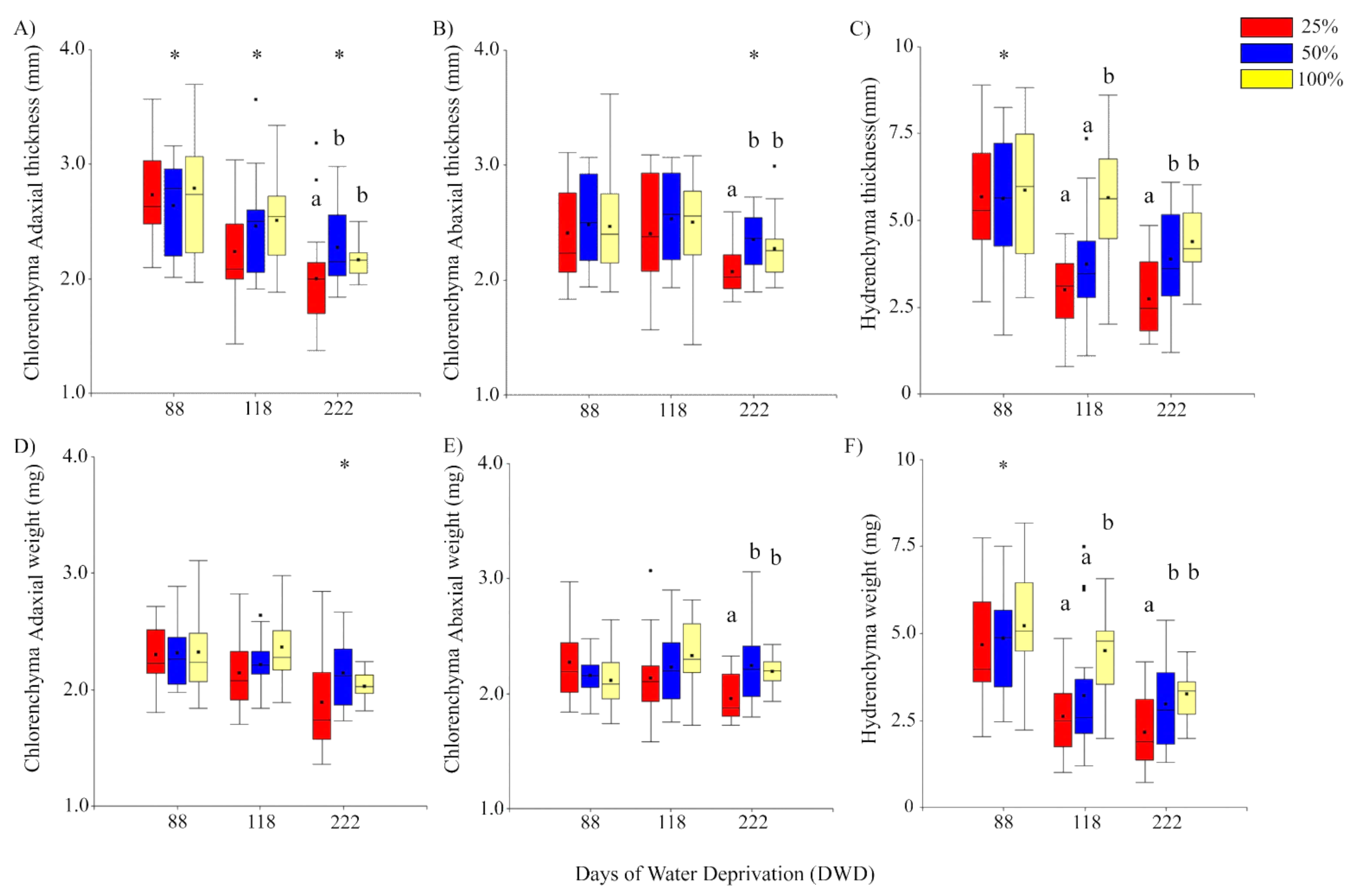
2.1.1. Chlorenchyma and Hydrenchyma Thickness
2.1.2. Chlorenchyma and Hydrenchyma Biomass
2.1.3. Proline and MDA
2.2. Photosynthetic Characteristics of Aloe vera under Prolonged Drought Stress
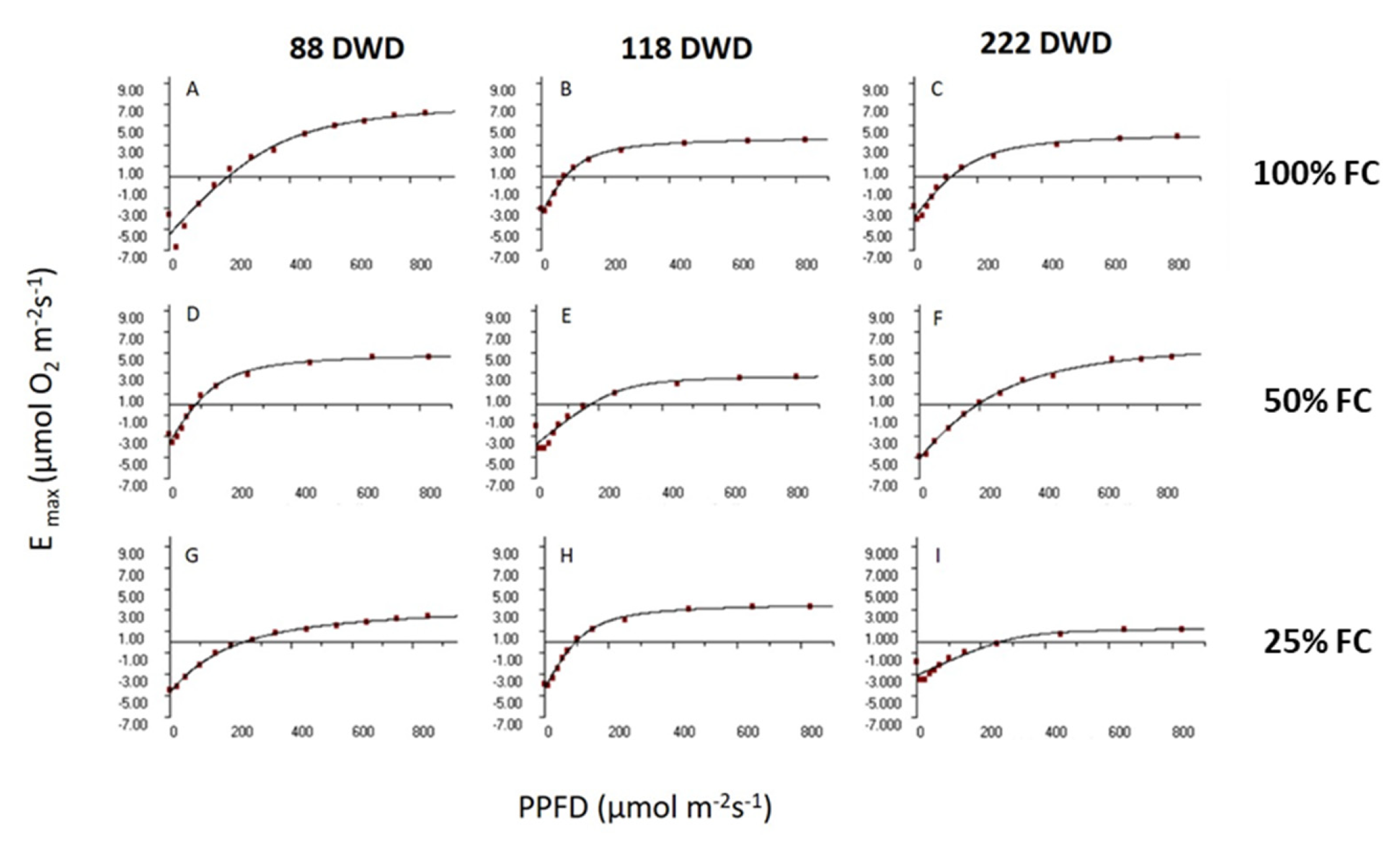
2.2.1. E/PFD Curves
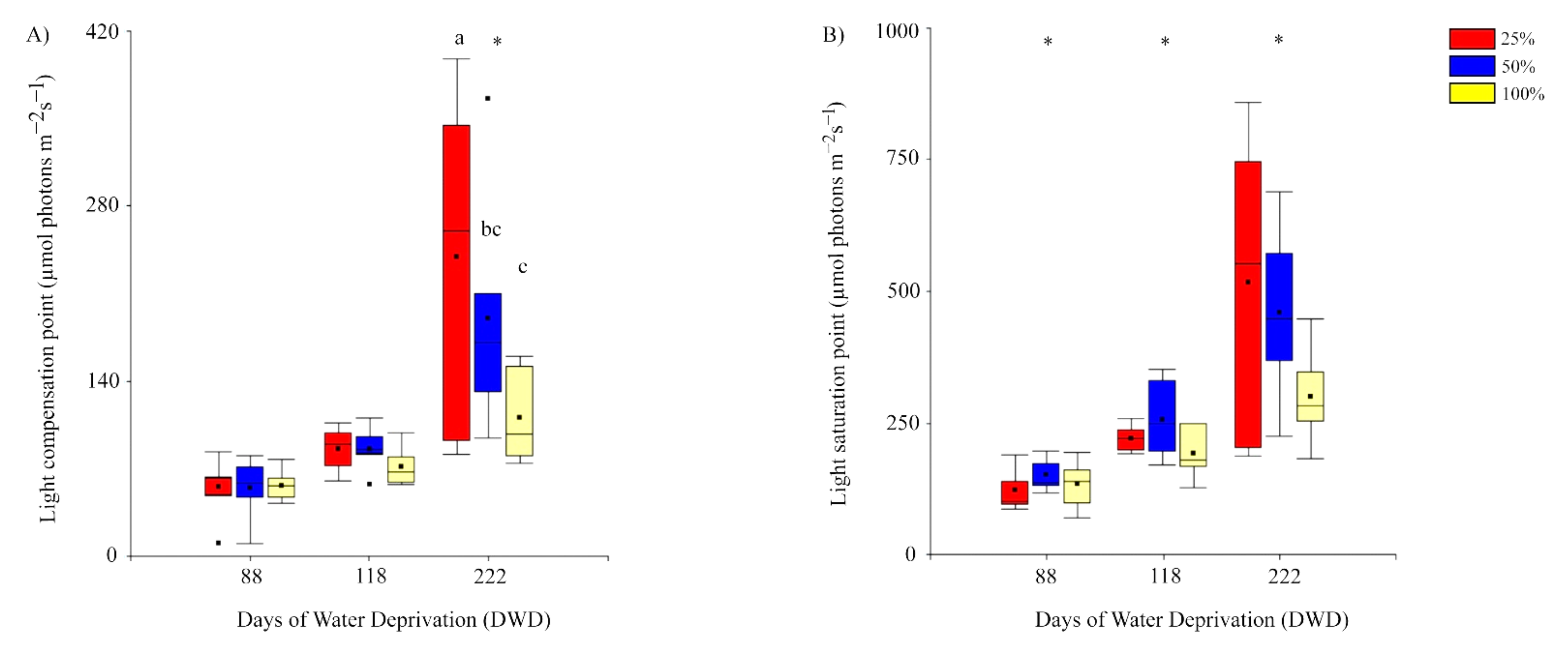
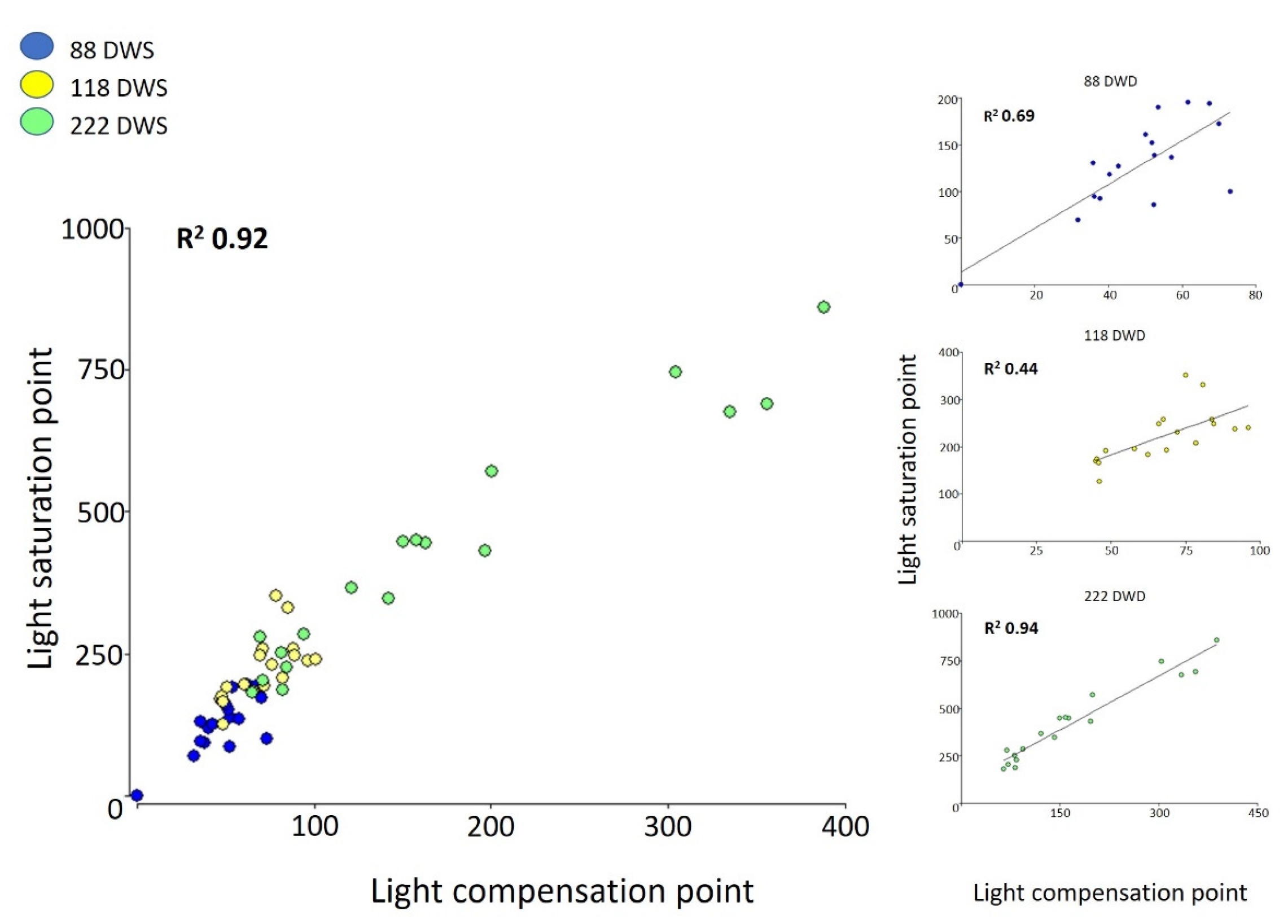
2.2.2. Light Compensation and Saturation Points
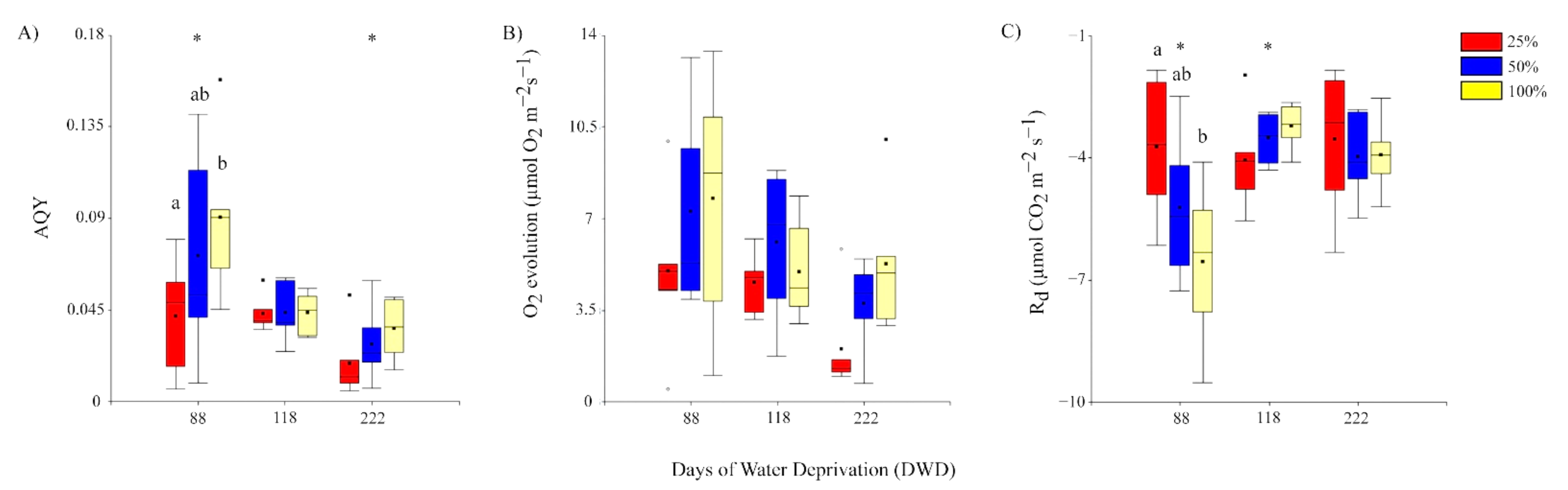
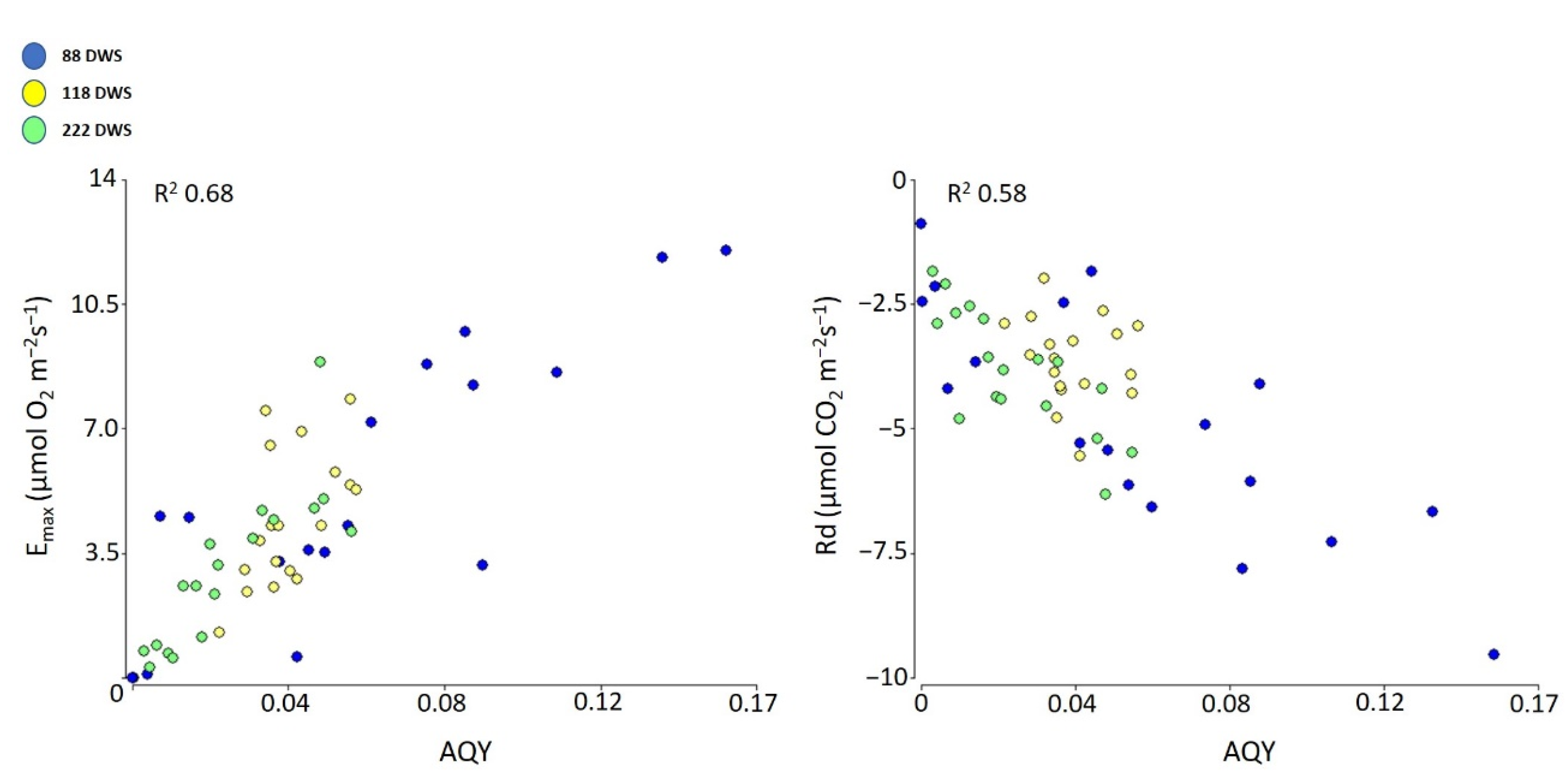
2.2.3. Apparent Quantum Yield (AQY), Photosynthetic Capacity (Emax), and Respiration (Rd)
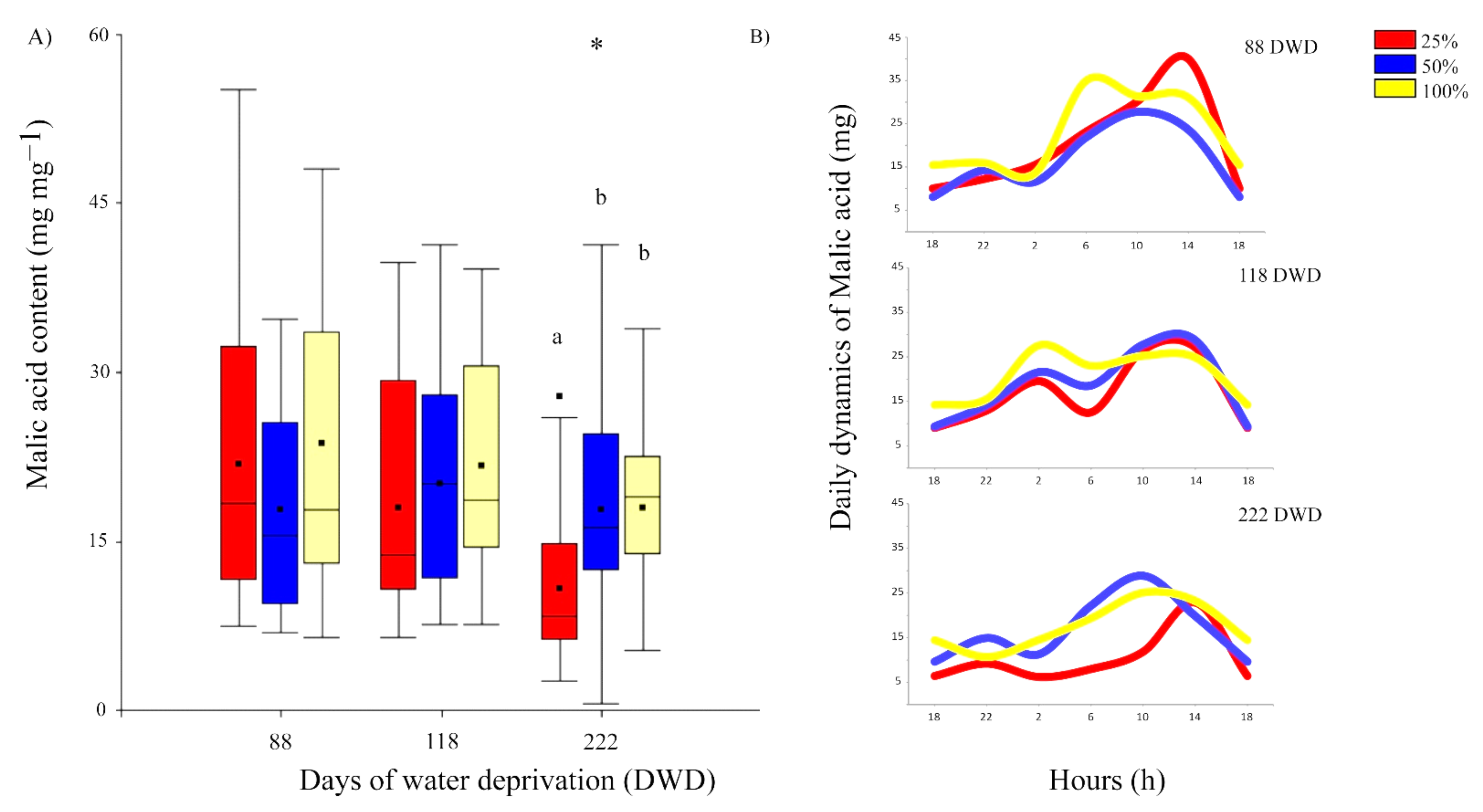
2.2.4. Malic Acid Evolution and Accumulation during Prolonged Drought Stress
3. Discussion
4. Materials and Methods
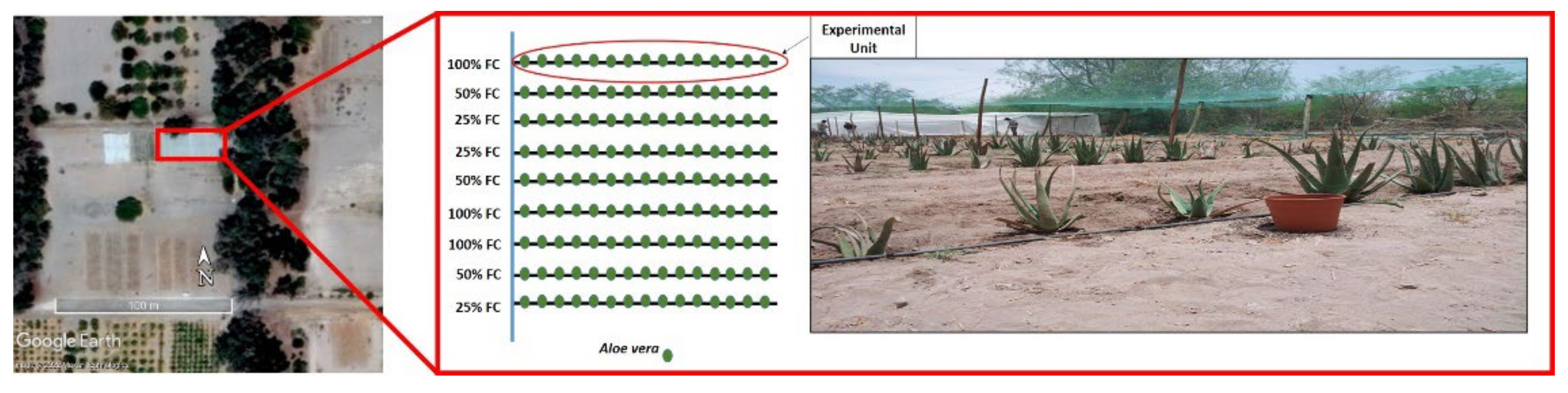
4.1. Study Site and Preconditioning Treatment
4.2. Prolonged Water Stress Experiment
4.3. Plant Biomass and Biochemical Parameters
4.4. Oxygen Evolution and Photosynthetic Parameters
4.5. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A

Appendix B

References
- Males, J. Secrets of succulence. J. Exp. Bot. 2017, 68, 2121–2134. [Google Scholar] [CrossRef] [PubMed]
- Eggli, U.; Nyffeler, R. Living under temporally arid conditions–succulence as an adaptive strategy. Bradleya 2009, 27, 13–36. [Google Scholar] [CrossRef]
- Griffiths, H. Plant Venation: From Succulence to succulents. Curr. Biol. 2013, 23, R340. [Google Scholar] [CrossRef] [Green Version]
- Grace, O.M. Succulent plant diversity as natural capital. Plants People Planet 2019, 1, 336–345. [Google Scholar] [CrossRef]
- Griffiths, H.; Males, J. Succulent plants. Curr. Biol. 2017, 27, R853–R909. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nobel, P.; Jordan, P. Transpiration stream of desert species: Resistances and capacitances for a C3, a C4, and a CAM plants. J. Exp. Bot. 1983, 34, 1379–1391. [Google Scholar] [CrossRef]
- Grace, O.M.; Buerki, S.; Symonds, R.E.; Forest, F.; Wyk, A.E.; Smith, G.F.; Klopper, R.R.; Bjora, C.S.; Neale, S.; Demissew, S.; et al. Evolutionary history and leaf succulence as explanations for medicinal use in aloes and the global popularity of Aloe vera. BMC Evol. Biol. 2015, 15, 29. [Google Scholar] [CrossRef] [Green Version]
- Hęś, M.; Dziedzic, K.; Górecka, D.; Jędrusek-Golińska, A.; Gujska, E. Aloe vera (L.) Webb.: Natural sources of antioxidants—A review. Plant Food Hum. Nutr. 2019, 74, 255–265. [Google Scholar] [CrossRef] [Green Version]
- Jaiswal, S.K.; Mahajan, S.; Chakraborty, A.; Kumar, S.; Sharma, V.K. The genome sequence of Aloe vera reveals adaptive evolution of drought tolerance mechanisms. iScience 2021, 24, 102079. [Google Scholar] [CrossRef]
- Silva, H.; Sagardia, S.; Ortiz, M.; Franck, N.; Opazo, M.; Quiroz, M.; Baginsky, C.; Tapia, C. Relationships between leaf anatomy, morphology, and water use efficiency in Aloe vera (L) Burm f. as a function of watyer availability. Rev. Chil. Hist. Nat. 2014, 87, 13. [Google Scholar] [CrossRef] [Green Version]
- Delatorre-Herrera, J.; Delfino, I.; Salinas, C.; Silva, H.; Cardemil, L. Irrigation restriction effects on water use efficiency and osmotic adjustment in Aloe vera plants (Aloe barbadensis Miller). Agric. Water Manag. 2010, 97, 1564–1570. [Google Scholar] [CrossRef]
- Zou, Y.; Han, C.; Wang, F.; Tan, Y.; Yang, S.; Huang, C.; Xie, S.; Xiao, X. Integrated metabolome and transcriptome analysis reveal complex molecular mechanisms underlying selenium response of Aloe vera L. J. Plant Biol. 2020, 64, 135–143. [Google Scholar] [CrossRef]
- Choudhri, P.; Rani, M.; Sangwan, R.S.; Kumar, R.; Kumar, A.; Chhokar, V. De novo sequencing, assembly and characterization of Aloe vera transcriptome and analysis of expression profiles of genes related to saponin and anthraquinone metabolism. BMB Genom. 2018, 19, 427. [Google Scholar] [CrossRef]
- Habibi, G. Effects of mild and severe drought stress on the biomass, phenolic compounds production and photochemical activity of Aloe vera (L.) Burm.f. Acta Agric. Scand. 2018, 111, 463–476. [Google Scholar] [CrossRef] [Green Version]
- Habibi, G. Changes in crassulacean acid metabolism expression, chloroplast ultrastructure, photochemical and antioxidant activity in the Aloe vera during acclimation to combined drought and salt stress. Funct. Plant Biol. 2021, 49, 40–53. [Google Scholar] [CrossRef]
- Ahl, L.I.; Mravec, J.; Jorgensen, B.; Rudall, P.J.; Ronsted, N.; Grace, O.M. Dynamics of intracellular mannan and cell wall folding in the drought responses of succulent Aloe species. Plant Cell Environ. 2019, 42, 2458–2471. [Google Scholar] [CrossRef] [Green Version]
- Xing, D.; Mao, R.; Li, Z.; Wu, Y.; Qin, X.; Fu, W. Leaf intracellular water transport rate based on physiological impedance: A possible role of leaf internal retained water in photosynthesis and growth of tomatoes. Front. Plant Sci. 2022, 13, 845628. [Google Scholar] [CrossRef]
- Khajeeyan, R.; Salehi, A.; Dehnavi, M.M.; Farajee, H.; Kohanmoo, M.A. Physiological and yield responses of Aloe vera plant to biofertilizers under different irrigation regimes. Agric. Water Manag. 2019, 225, 105768. [Google Scholar] [CrossRef]
- Nobel, P. Parenchyma-Chlorenchyma water movement during drought for the hemiepiphytic cactus Hylocereus undatus. Ann. Bot. 2006, 97, 469–474. [Google Scholar] [CrossRef] [Green Version]
- Silva, H.; Sagardia, S.; Seguel, O.; Torres, C.; Tapia, C.; Franck, N.; Cardemil, L. Effect of water availability on growth and water use efficiency for biomass and gel production in Aloe vera (Aloe barbadensis M.). Ind. Crop. Prod. 2010, 31, 20–27. [Google Scholar] [CrossRef]
- Manzoni, S.; Vico, G.; Katul, G.; Palmroth, S.; Porporato, A. Optimal plant water-use strategies under stochastic rainfall. Water Resour. Res. 2014, 50, 5379–5394. [Google Scholar] [CrossRef]
- Males, J.; Griffiths, H. Stomatal biology of CAM plants. Plant Physiol. 2017, 174, 550–560. [Google Scholar] [CrossRef] [Green Version]
- Lucini, L.; Pellizzoni, M.; Pellegrino, R.; Molinari, G.P.; Colla, G. Phytochemical constituents and in vitro radical scavenging activity of different Aloe species. Food Chem. 2015, 170, 501–507. [Google Scholar] [CrossRef]
- Pinheiro, H.A.; DaMatta, F.M.; Chaves, A.R.M.; Fontes, E.P.B.; Loureiro, M.E. Drought tolerance in relation to protection against oxidative stress in clones of Coffea canephora subjected to long term drought. Plant Sci. 2004, 167, 1307–1314. [Google Scholar] [CrossRef]
- Silva-Lima, C.; Ferreira-Silva, S.L.; Leite-Carvalho, F.E.; Lima-Neto, M.C.; Magalhaes-Aragao, R.; Nascimento-Silva, E.; Sousa, R.M.J.; Gomes-Silveira, J.A. Antioxidant protection and PSII regulation mitigate photo-oxidative stress induced by drought followed by high light in cashew plants. Environ. Exp. Bot. 2018, 149, 59–69. [Google Scholar] [CrossRef] [Green Version]
- Smith, J.A.C.; Lüttge, U. Day-night changes in leaf water relations associated with the rhythm of crassulacean acid metabolism in Kalanchoe dagremontiana. Planta 1985, 163, 272–282. [Google Scholar] [CrossRef]
- Lütge, U. Ecophysiology of CAM photosynthesis. In Terrestrial Photosynthesis in A Changing Environment, 1st ed.; Flexas, J., Loretto, F., Medrano, H., Eds.; Cambridge University Press: Cambridge, UK, 2015; pp. 71–84. [Google Scholar]
- Lüttge, U. Stem CAM in arborescent succulents. Tree 2008, 22, 139–148. [Google Scholar] [CrossRef]
- Vitale, L.; Vitale, E.; Costanzo, G.; De Maio, A.; Arena, C. Photo-protective mechanisms and the role of poly (ADP-Ribose) polymerase activity in a facultative CAM plant exposed to long-term water deprivation. Plants 2020, 9, 1192. [Google Scholar] [CrossRef] [PubMed]
- Oyarce, A.; Delatorre-Herrera, J.; Delatorre-Castillo, J.P.; Sepúlveda-Soto, I.; González-Castro, R.; Low, C.; Pavon-Djavid, G.; Ostria-Gallardo, E.; Cardemil, L. Influence of water stress and radiation, on the fructan production and its relationship with the hydrenquima water content and the photosynthetic activity in Aloe vera (L) Burm f. plants. in preparation.
- Maxwell, C.; Griffiths, H.; Borland, A.M.; Broadmeadow, J.S.J.; McDavid, C.R. Photoinhibitory responses of the epiphytic bromeliad Guzmania monostachia during the dry season in Trinidad maintain photochemical integrity under adverse conditions. Plant Cell Environ. 1992, 15, 37–47. [Google Scholar] [CrossRef]
- Maxwell, C.; Griffiths, H.; Young, A.J. Photosynthetic acclimation to light regime and water stress by the C3-CAM epiphyte Guzmania monostachia: Gas-exchange characteristics, photochemical efficiency and the xanthophyll cycle. Funct. Ecol. 1994, 6, 746–754. [Google Scholar] [CrossRef]
- Merzlyak, M.; Solovchenko, A.; Pogosyan, S. Optical properties of rhodoxanthin accumulated in Aloe arborescens Mill. leaves under high-light stress with special reference to its photoprotective function. Photochem. Photobiol. Sci. 2005, 4, 333–340. [Google Scholar] [CrossRef]
- Flexas, J.; Bota, J.; Loreto, F.; Cornic, G.; Sharkey, T.D. Diffusive and metabolic limitations to photosynthesis under drought and salinity in C3 plants. Plant Biol. 2004, 6, 269–279. [Google Scholar] [CrossRef]
- Lüttge, U.; Ball, E.; Fetene, M. Flexibility of crassulacean acid metabolism in Kalanchoe pinnata (Lam.) Pers. II. Light-use characteristics of plants grown in low or high light. J. Plant Physiol. 1991, 137, 268–272. [Google Scholar] [CrossRef]
- delRío, C.; Lobos, F.; Siegmund, A.; Tejos, C.; Osses, P.; Huaman, Z.; Meneses, J.P.; García, J.-L. GOFOS, ground optical fog observation system for monitoring the vertical stratocumulus-fog cloud distribution in the coast of the Atacama Desert, Chile. J. Hydrol. 2021, 597, 126190. [Google Scholar] [CrossRef]
- Latorre, C.; González, A.L.; Quade, J.; Fariña, J.M.; Pinto, R.; Marquet, P. Establishment and formation of fog-dependent Tillandsia landbeckii dunes in the Atacama Desert: Evidence from radiocarbon and stable isotopes. J. Geophys. Res. 2011, 116, G03033. [Google Scholar]
- Poblete, I.; Lanino, M.; Rojas, G. El descubrimiento de una sobreviviente: Cepa vinífera Tamarugal. Idesia 2020, 38, 47–58. [Google Scholar] [CrossRef]
- Nachabe, M.H. Refining the definition of field capacity in the literature. J. Irrig. Drain. Eng. 1998, 124, 230–232. [Google Scholar] [CrossRef]
- Rivero, R.M.; Ruiz, J.M.; Romero, L.M. Importance of N source on heat stress tolerance due to the accyumulation of proline and quaternary ammonium compounds in tomato plants. Plant Biol. 2004, 6, 702–707. [Google Scholar] [CrossRef]
- Ortega-Villasante, C.; Rellán-Álvarez, R.; Del Campo, F.F.; Carpena-Ruiz, R.O.; Hernández, L.E. Cellular damage induced by cadmium and mercury in Medicago sativa. J. Exp. Bot. 2005, 56, 2239–2251. [Google Scholar] [CrossRef]
- Pastenes, C.; Horton, P. Effect of high temperature on photosynthesis in bean. I. Oxygen evolution and chlorophyll fluorescence. Plant Physiol. 1996, 112, 1245–1251. [Google Scholar] [CrossRef] [Green Version]
- Di Rienzo, J.A.; Casanoves, F.; Balzarini, M.G.; González, L.; Tablada, M.; Robledo, W. Infostat, Versión 2008; Grupo Infostat, FCA, Universidad Nacional de Córdoba: Córdoba, Argentina, 2008. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment FC | Mean Leaf N° | Leaf Thickness (cm) | Volume Leaf Disc (cm3) | Plant Diameter (cm) | Plant Height (cm) |
|---|---|---|---|---|---|
| 25% | 15.3 | 0.825 | 8.25 | 50.07 | 45.68 |
| 50% | 15.4 | 0.794 | 7.94 | 49.00 | 48.10 |
| 100% | 14.7 | 0.839 | 8.4 | 48.53 | 47.74 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Delatorre-Castillo, J.P.; Delatorre-Herrera, J.; Lay, K.S.; Arenas-Charlín, J.; Sepúlveda-Soto, I.; Cardemil, L.; Ostria-Gallardo, E. Preconditioning to Water Deficit Helps Aloe vera to Overcome Long-Term Drought during the Driest Season of Atacama Desert. Plants 2022, 11, 1523. https://doi.org/10.3390/plants11111523
Delatorre-Castillo JP, Delatorre-Herrera J, Lay KS, Arenas-Charlín J, Sepúlveda-Soto I, Cardemil L, Ostria-Gallardo E. Preconditioning to Water Deficit Helps Aloe vera to Overcome Long-Term Drought during the Driest Season of Atacama Desert. Plants. 2022; 11(11):1523. https://doi.org/10.3390/plants11111523
Chicago/Turabian StyleDelatorre-Castillo, José P., José Delatorre-Herrera, Kung Sang Lay, Jorge Arenas-Charlín, Isabel Sepúlveda-Soto, Liliana Cardemil, and Enrique Ostria-Gallardo. 2022. "Preconditioning to Water Deficit Helps Aloe vera to Overcome Long-Term Drought during the Driest Season of Atacama Desert" Plants 11, no. 11: 1523. https://doi.org/10.3390/plants11111523