
Antioxidant Capacity of Potentilla paradoxa Nutt. and Its Beneficial Effects Related to Anti-Aging in HaCaT and B16F10 Cells

 , ,
, , 
Abstract
:1. Introduction
2. Results
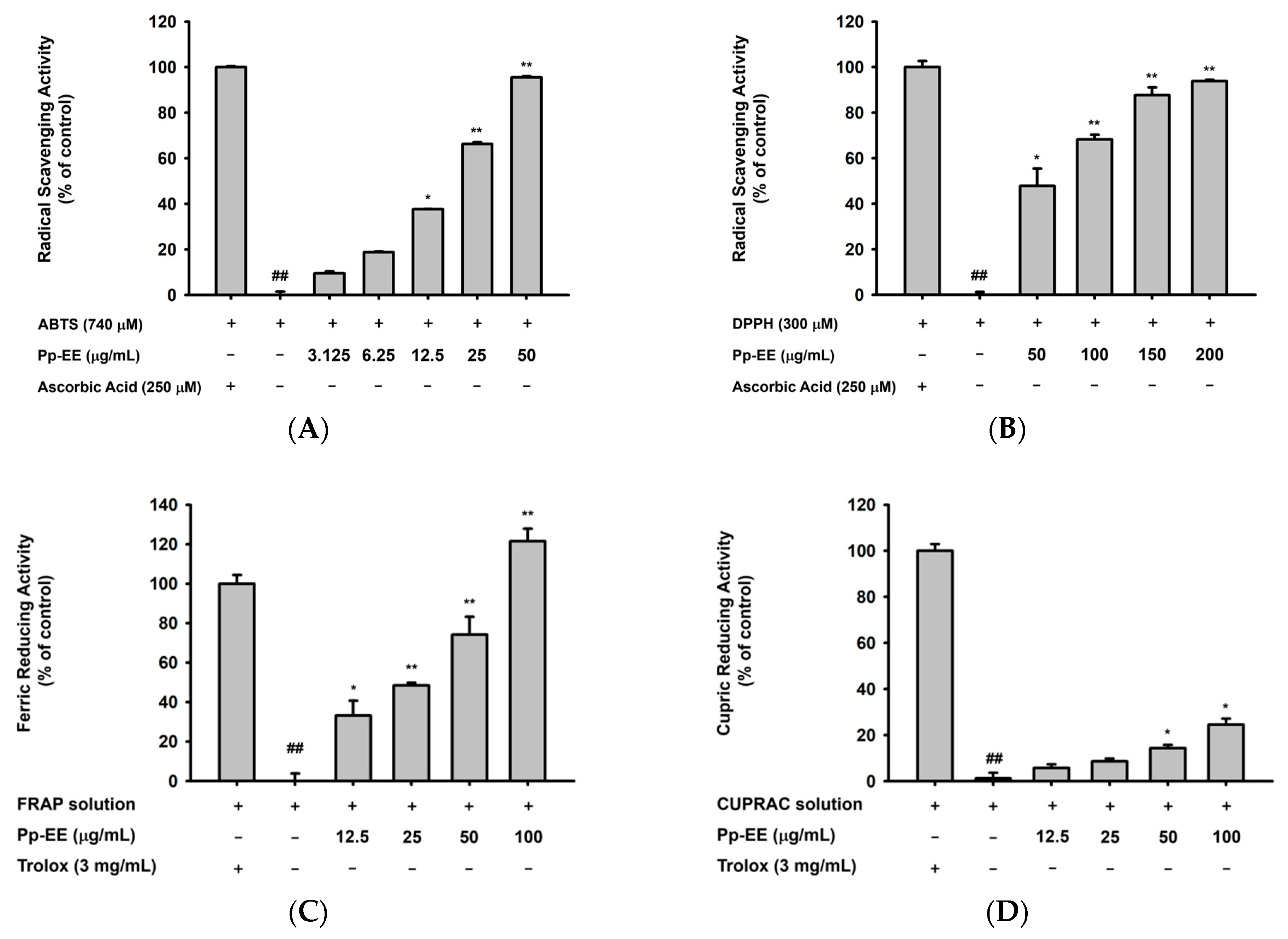
2.1. Antioxidative Capacity of Pp-EE
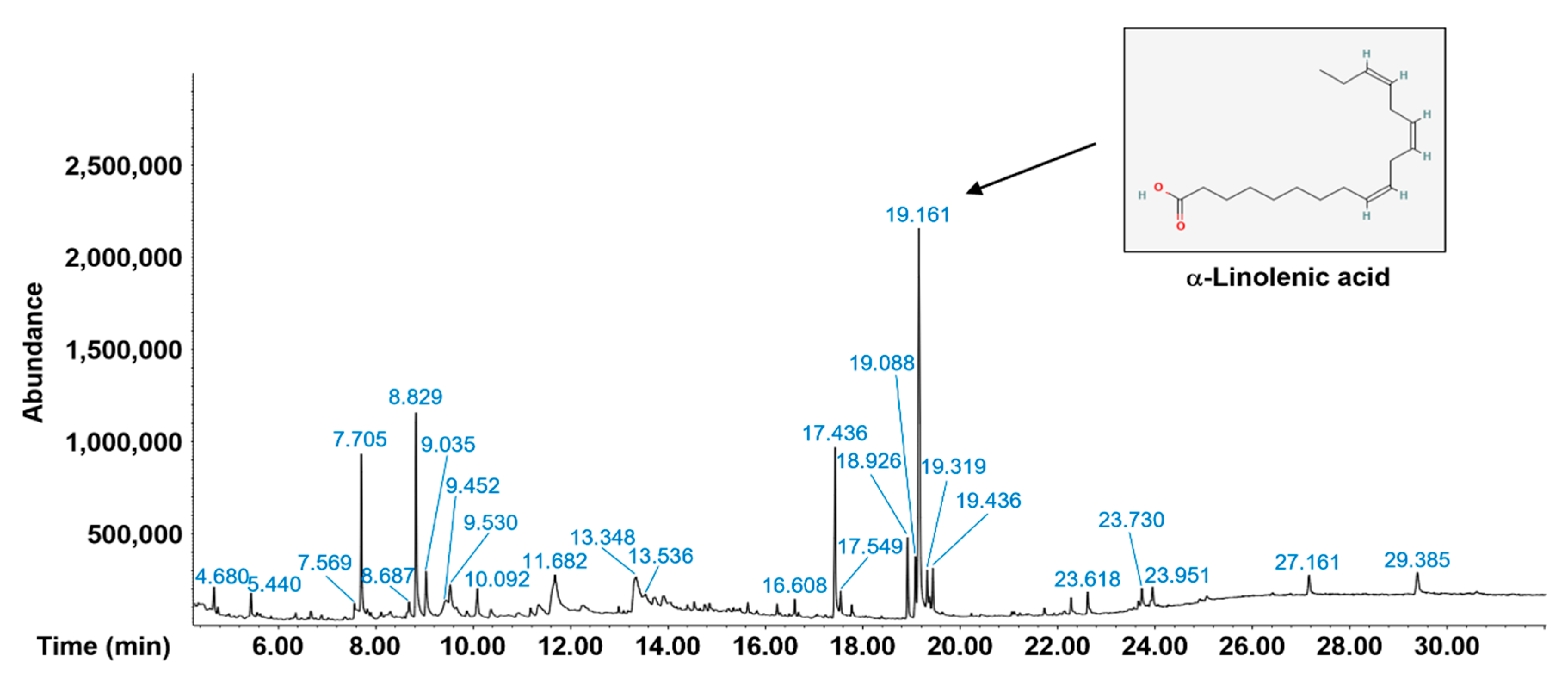
2.2. Phytochemical Components of Pp-EE
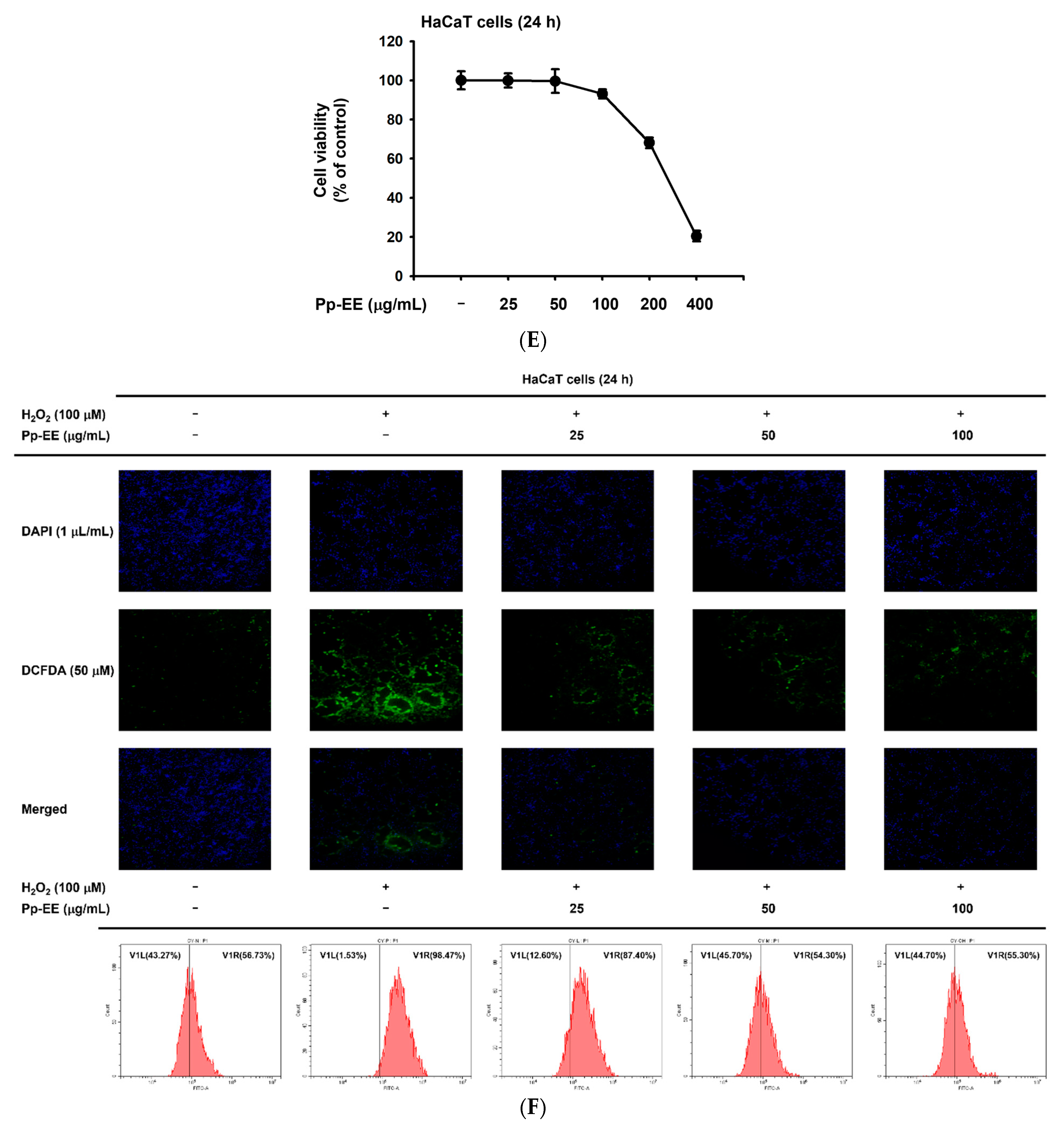
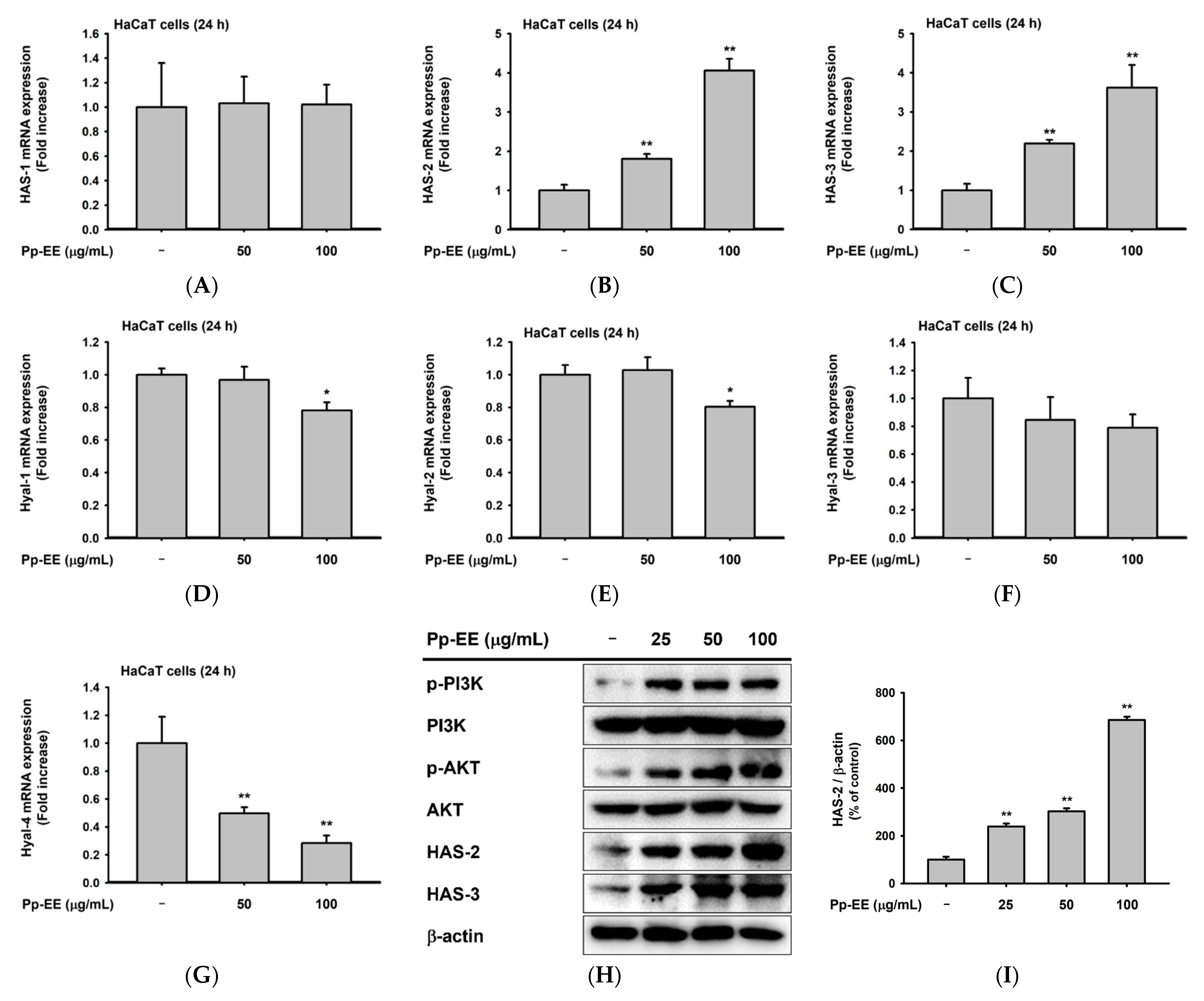
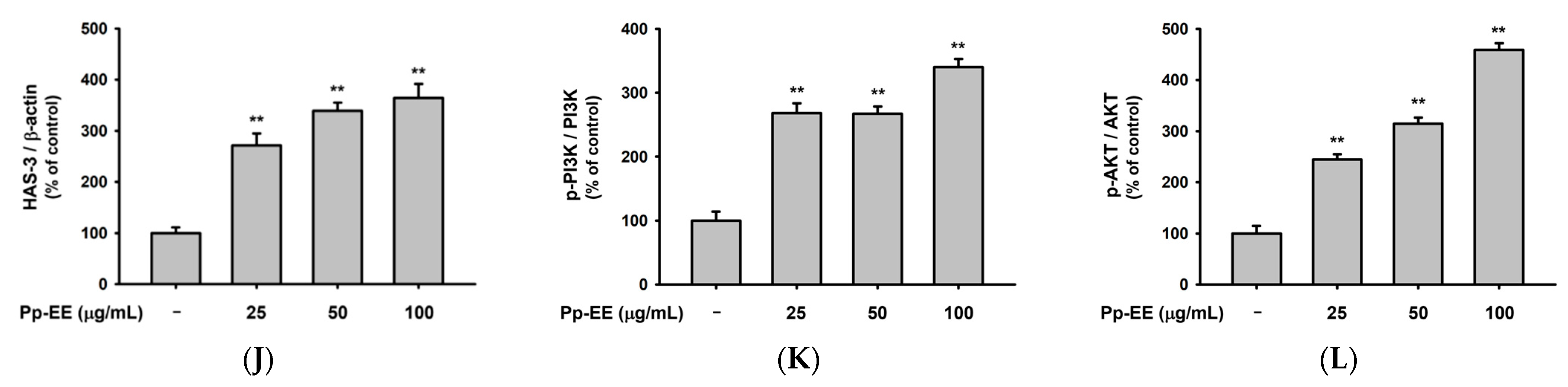
2.3. Effects of Pp-EE on Parameters That Can Affect Skin Moisture
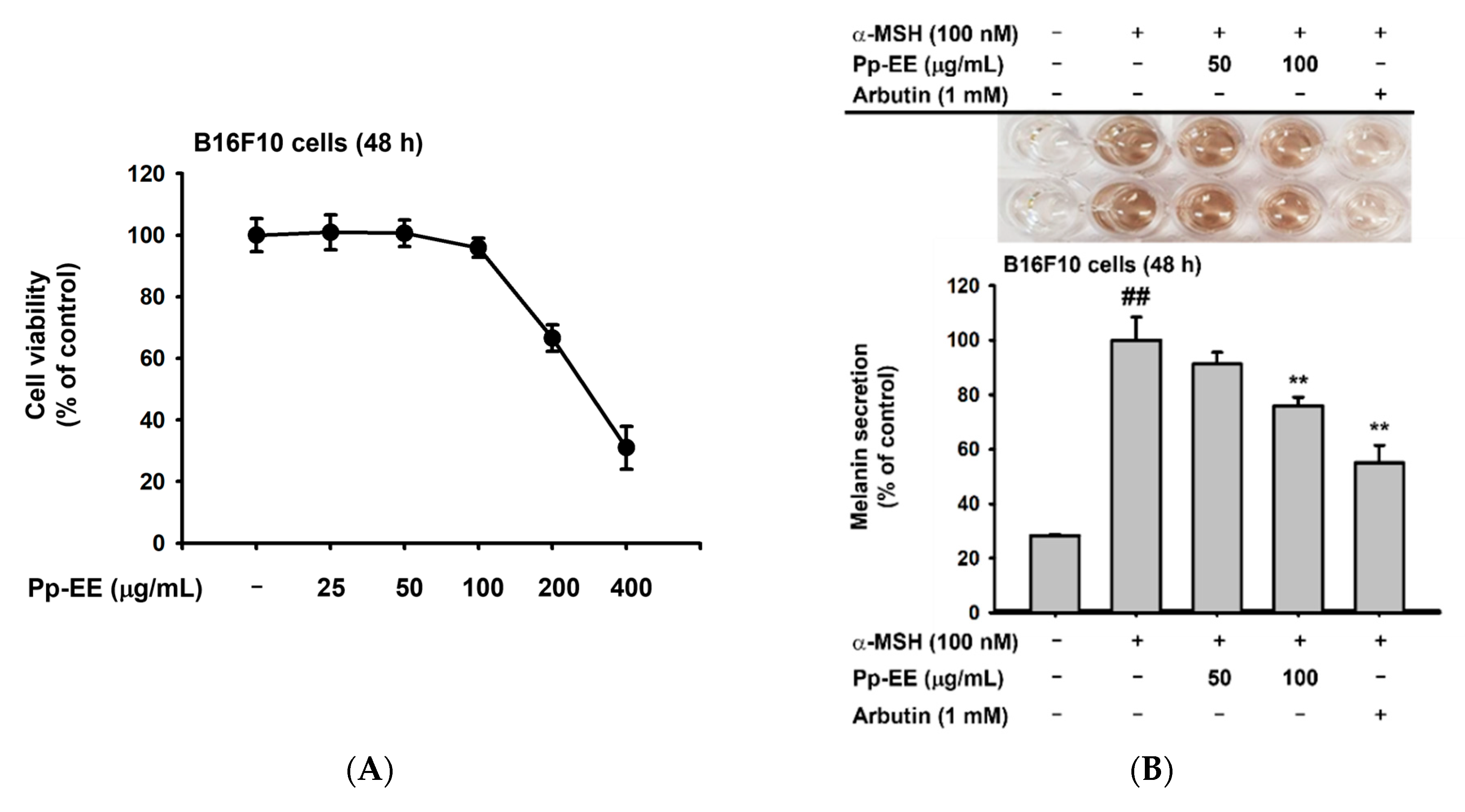
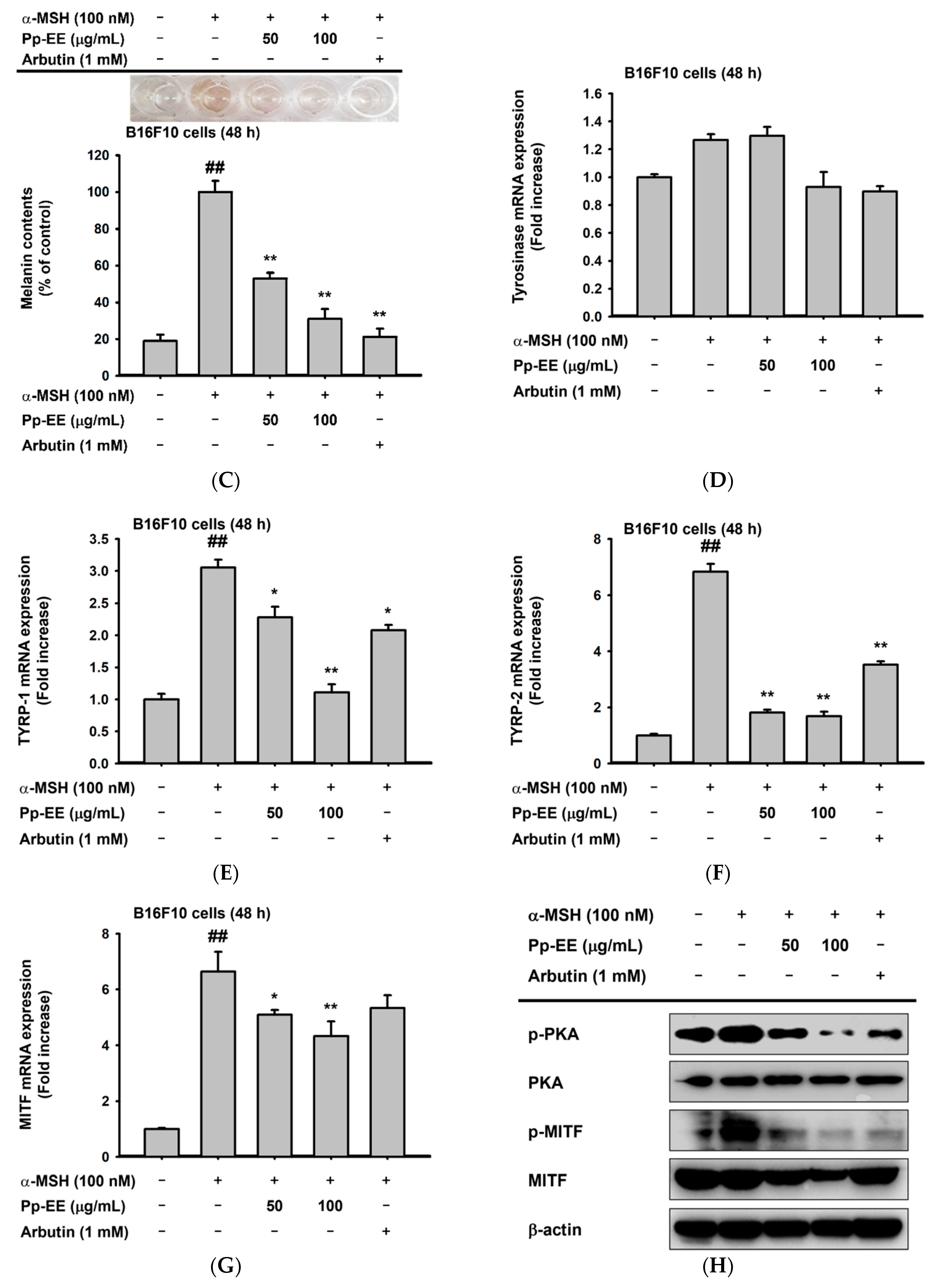
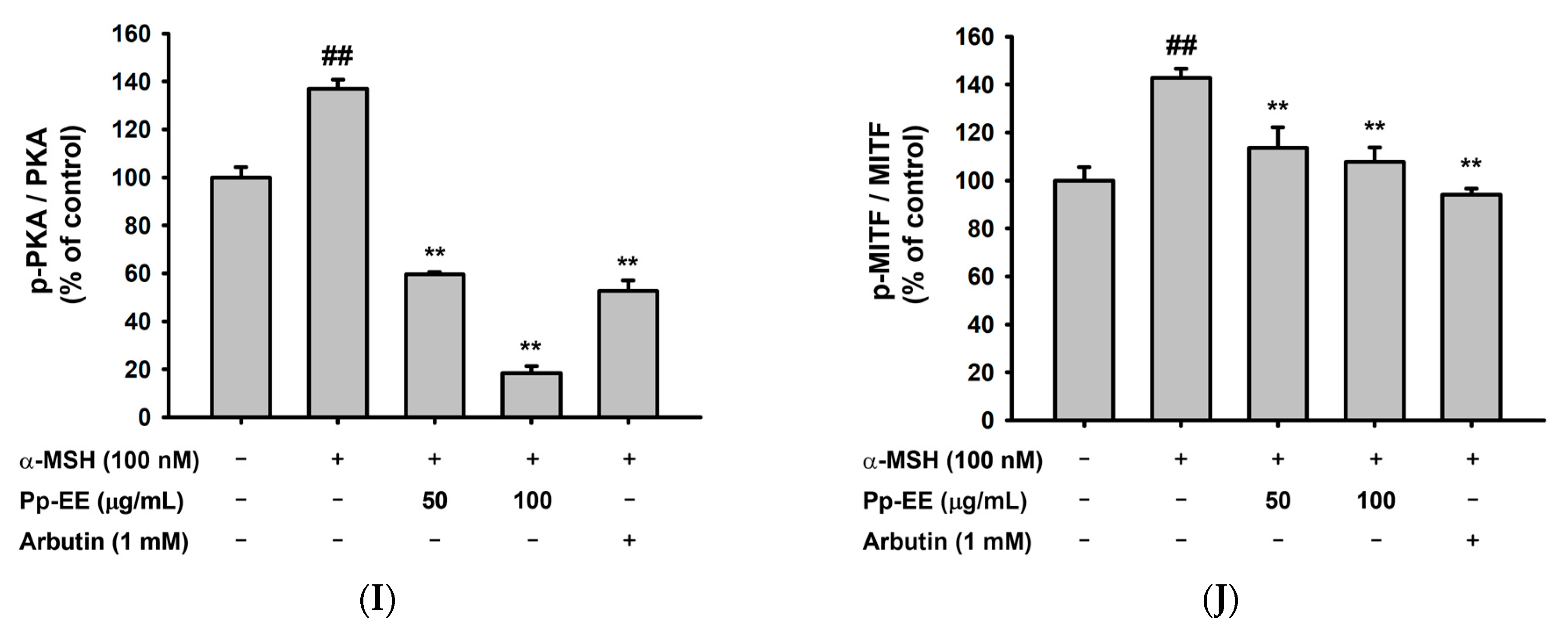
2.4. Anti-Melanogenesis Activity of Pp-EE in Melanocytes
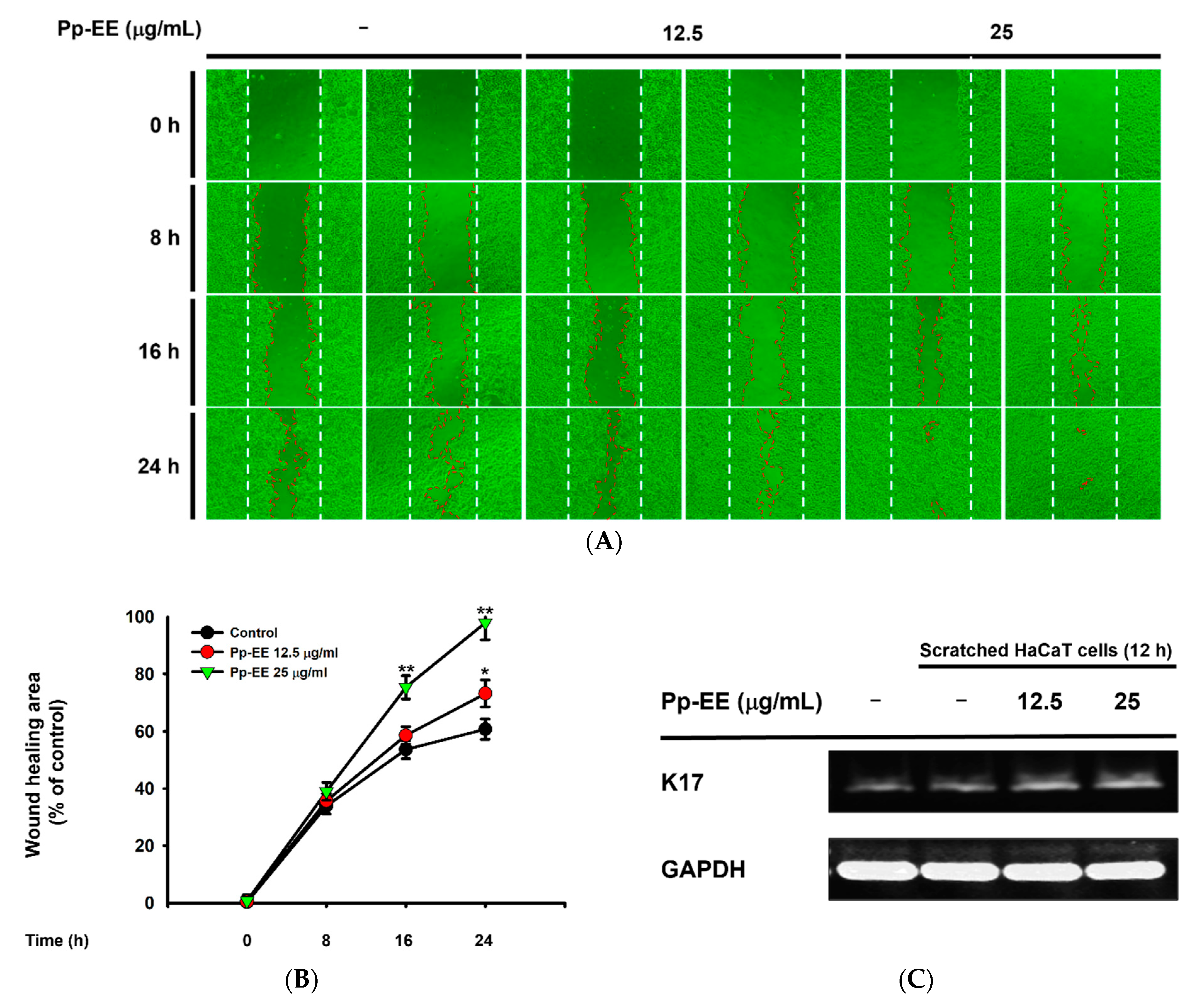
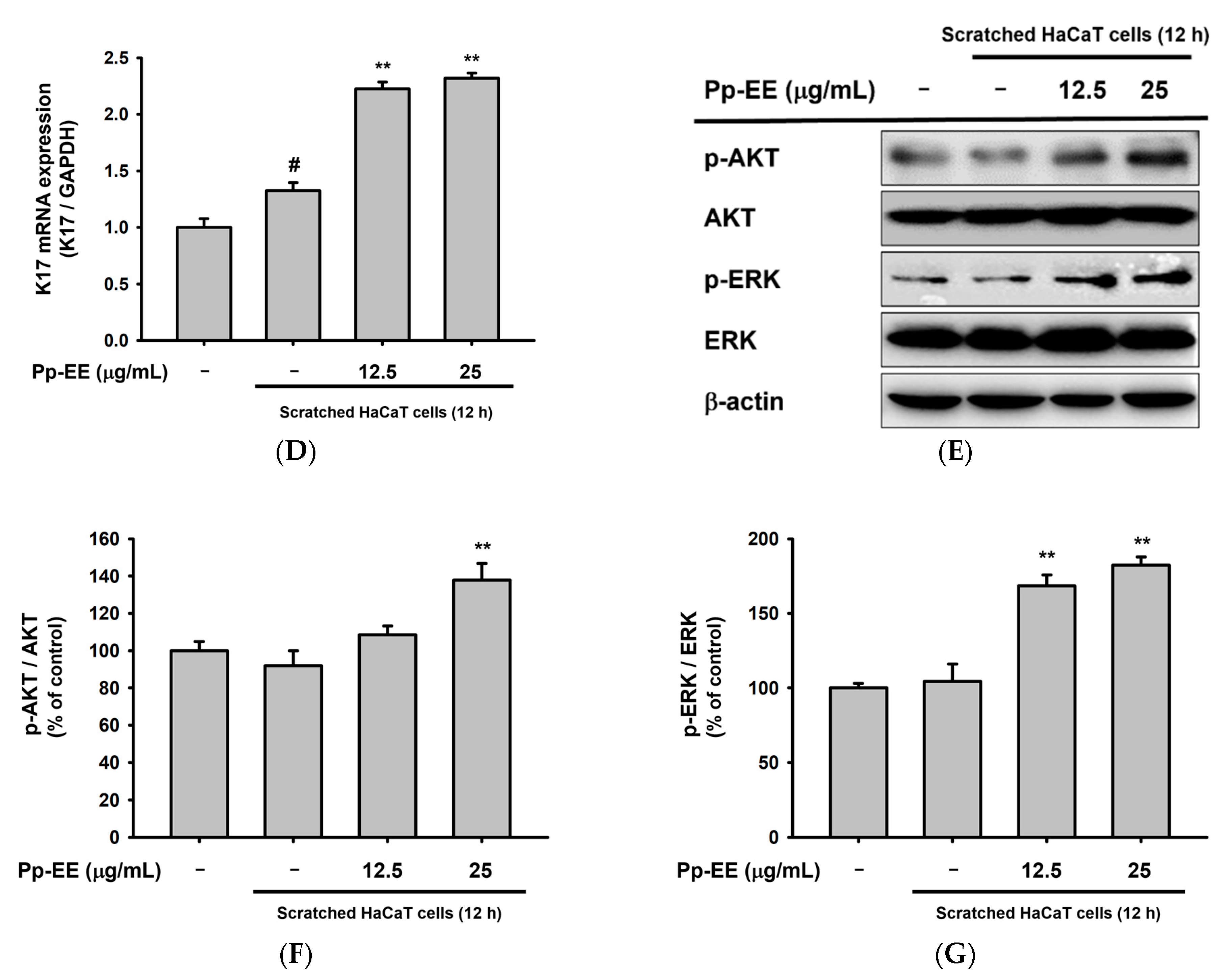
2.5. Pp-EE Promotes Wound-Healing Processes in Keratinocytes
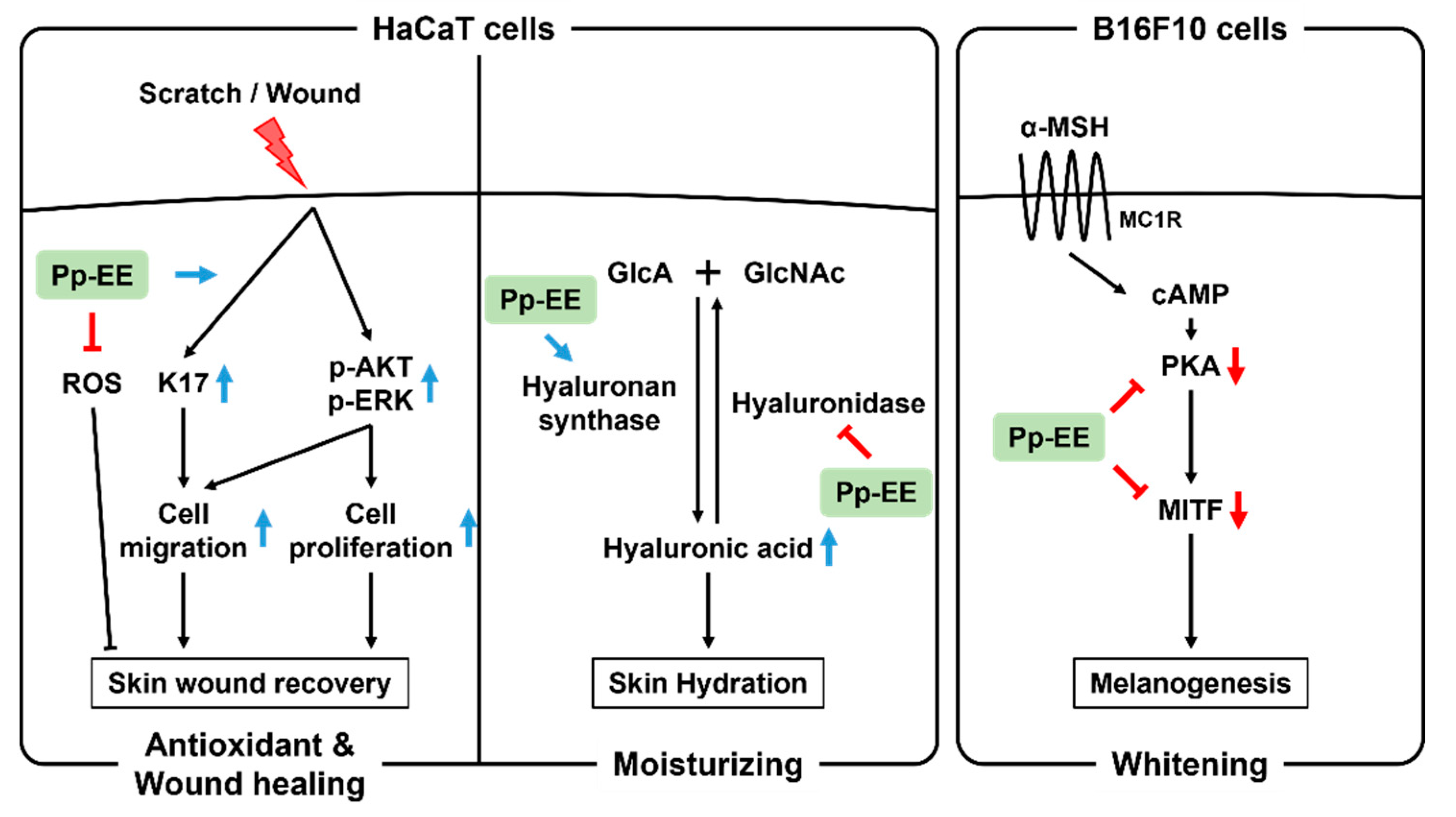
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Preparation of Pp-EE and Gas Chromatography–Mass Spectrometry
4.3. Determination of Total Phenolic Content
4.4. Determination of Total Flavonoid Content
4.5. ABTS Radical Scavenging Activity
4.6. DPPH Radical Scavenging Activity
4.7. Ferric Reducing Antioxidant Power Assay
4.8. Cupric Ion Reducing Antioxidant Capacity Assay
4.9. Cell Culture
4.10. Cell Viability Assay
4.11. ROS Generation Assay
4.12. Melanin Generation Assay
4.13. In Vitro Scratch Assay
4.14. Semi-Quantitative Reverse Transcription-PCR (RT-PCR) and Quantitative Real-Time PCR (Real-Time PCR)
4.15. Preparation of Whole Cell Lysates and Immunoblotting Analysis
4.16. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| Pp-ME | Potentilla paradoxa methanol extract |
| ROS | Reactive oxygen species |
| ECM | Extracellular matrix |
| EGFR | Epidermal growth factor receptor |
| α-MSH | α-melanocyte-stimulating hormone |
| ERK | extracellular signal-regulated kinases |
| cAMP | Cyclic adenosine monophosphate |
| PKA | protein kinase A |
| MITF | Microphthalmia-associated transcription factor |
| MC1R | Melanocortin 1 receptor |
| CREB | cAMP response element |
| RT-PCR | Reverse transcription-polymerase chain reaction |
| HPLC | High-performance liquid chromatography |
| FRAP | Ferric reducing antioxidant power |
| GC-MS | Gas chromatography–mass spectrometry |
| HA | Hyaluronic acid |
| PI3K | Phosphoinositide 3-kinase |
| AKT | Protein kinase B |
| K17 | Keratin 17 |
| CUPRAC | Cupric ion reducing antioxidant capacity |
References
- Tobin, D.J. Introduction to skin aging. J. Tissue Viability 2017, 26, 37–46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Truong, V.L.; Keum, Y.S.; Jeong, W.S. Red ginseng oil promotes hair growth and protects skin against UVC radiation. J. Ginseng Res. 2021, 45, 498–509. [Google Scholar] [CrossRef] [PubMed]
- Bouwstra, J.A.; Ponec, M. The skin barrier in healthy and diseased state. Biochim. Biophys. Acta Biomembr. 2006, 1758, 2080–2095. [Google Scholar] [CrossRef] [Green Version]
- Sturm, R.A.; Duffy, D.L. Human pigmentation genes under environmental selection. Genome Biol. 2012, 13, 248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rognoni, E.; Watt, F.M. Skin cell heterogeneity in development, wound healing, and cancer. Trends Cell Biol. 2018, 28, 709–722. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Costin, G.-E.; Hearing, V.J. Human skin pigmentation: Melanocytes modulate skin color in response to stress. FASEB J. 2007, 21, 976–994. [Google Scholar] [CrossRef]
- Griendling, K.K.; Touyz, R.M.; Zweier, J.L.; Dikalov, S.; Chilian, W.; Chen, Y.-R.; Harrison, D.G.; Bhatnagar, A. Measurement of reactive oxygen species, reactive nitrogen species, and redox-dependent signaling in the cardiovascular system: A scientific statement from the American Heart Association. Circ. Res. 2016, 119, e39–e75. [Google Scholar] [CrossRef]
- Di Meo, S.; Reed, T.T.; Venditti, P.; Victor, V.M. Role of ROS and RNS sources in physiological and pathological conditions. Oxid. Med. Cell. Longev. 2016, 2016, 1245049. [Google Scholar] [CrossRef]
- Shields, H.J.; Traa, A.; Van Raamsdonk, J.M. Beneficial and dtrimental effects of reactive oxygen species on lifespan: A comprehensive review of comparative and experimental studies. Front. Cell Develop. Biol. 2021, 9, 181. [Google Scholar] [CrossRef]
- Halliwell, B. Biochemistry of oxidative stress. Biochem. Soc. Trans. 2007, 35, 1147–1150. [Google Scholar] [CrossRef]
- Kasote, D.M.; Katyare, S.S.; Hegde, M.V.; Bae, H. Significance of antioxidant potential of plants and its relevance to therapeutic applications. Int. J. Biol. Sci. 2015, 11, 982. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Papakonstantinou, E.; Roth, M.; Karakiulakis, G. Hyaluronic acid: A key molecule in skin aging. Dermato-Endocrinol. 2012, 4, 253–258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakamura-Utsunomiya, A. Bone biomarkers in mucopolysaccharidoses. Int. J. Mol. Sci. 2021, 22, 12651. [Google Scholar] [CrossRef] [PubMed]
- Gupta, R.C.; Lall, R.; Srivastava, A.; Sinha, A. Hyaluronic acid: Molecular mechanisms and therapeutic trajectory. Front. Vet. Sci. 2019, 6, 192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kobayashi, T.; Chanmee, T.; Itano, N. Hyaluronan: Metabolism and function. Biomolecules 2020, 10, 1525. [Google Scholar] [CrossRef] [PubMed]
- Eghbali-Feriz, S.; Taleghani, A.; Al-Najjar, H.; Emami, S.A.; Rahimi, H.; Asili, J.; Hasanzadeh, S.; Tayarani-Najaran, Z. Anti-melanogenesis and anti-tyrosinase properties of Pistacia atlantica subsp. mutica extracts on B16F10 murine melanoma cells. Res. Pharm. Sci. 2018, 13, 533. [Google Scholar] [PubMed]
- Oh, T.-I.; Yun, J.-M.; Park, E.-J.; Kim, Y.-S.; Lee, Y.-M.; Lim, J.-H. Plumbagin suppresses α-MSH-induced melanogenesis in B16F10 mouse melanoma cells by inhibiting tyrosinase activity. Int. J. Mol. Sci. 2017, 18, 320. [Google Scholar] [CrossRef] [Green Version]
- Videira, I.F.d.S.; Moura, D.F.L.; Magina, S. Mechanisms regulating melanogenesis. An. Bras. Dermatol. 2013, 88, 76–83. [Google Scholar] [CrossRef] [Green Version]
- D’Mello, S.A.; Finlay, G.J.; Baguley, B.C.; Askarian-Amiri, M.E. Signaling pathways in melanogenesis. Int. J. Mol. Sci. 2016, 17, 1144. [Google Scholar] [CrossRef] [Green Version]
- Silveira, J.E.P.S.; Pedroso, D.M.M. UV light and skin aging. Rev. Environ. Health 2014, 29, 243–254. [Google Scholar]
- Sgonc, R.; Gruber, J. Age-related aspects of cutaneous wound healing: A mini-review. Gerontology 2013, 59, 159–164. [Google Scholar] [CrossRef] [PubMed]
- Wawersik, M.J.; Mazzalupo, S.; Nguyen, D.; Coulombe, P.A. Increased levels of keratin 16 alter epithelialization potential of mouse skin keratinocytes in vivo and ex vivo. Mol. Biol. Cell 2001, 12, 3439–3450. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patel, G.K.; Wilson, C.H.; Harding, K.G.; Finlay, A.Y.; Bowden, P.E. Numerous keratinocyte subtypes involved in wound re-epithelialization. J. Investig. Dermatol. 2006, 126, 497–502. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Werner, S.; Grose, R. Regulation of wound healing by growth factors and cytokines. Physiol. Rev. 2003, 83, 835–870. [Google Scholar] [CrossRef]
- Repertinger, S.K.; Campagnaro, E.; Fuhrman, J.; El-Abaseri, T.; Yuspa, S.H.; Hansen, L.A. EGFR enhances early healing after cutaneous incisional wounding. J. Investig. Dermatol. 2004, 123, 982–989. [Google Scholar] [CrossRef] [Green Version]
- Pastore, S.; Mascia, F.; Mariani, V.; Girolomoni, G. The epidermal growth factor receptor system in skin repair and inflammation. J. Investig. Dermatol. 2008, 128, 1365–1374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.; Kim, M.S.; Jung, S.-J.; Kim, D.; Park, H.J.; Cho, D. ERK activating peptide, AES16-2M promotes wound healing through accelerating migration of keratinocytes. Sci. Rep. 2018, 8, 1–10. [Google Scholar] [CrossRef]
- Tomczyk, M.; Latté, K.P. Potentilla—A review of its phytochemical and pharmacological profile. J. Ethnopharmacol. 2009, 122, 184–204. [Google Scholar] [CrossRef]
- Uysal, S.; Zengin, G.; Locatelli, M.; Bahadori, M.B.; Mocan, A.; Bellagamba, G.; De Luca, E.; Mollica, A.; Aktumsek, A. Cytotoxic and enzyme inhibitory potential of two Potentilla species (P. speciosa L. and P. reptans Willd.) and their chemical composition. Front. Pharmacol. 2017, 8, 290. [Google Scholar] [CrossRef]
- Gairola, S.; Sharma, J.; Bedi, Y.S. A cross-cultural analysis of Jammu, Kashmir and Ladakh (India) medicinal plant use. J. Ethnopharmacol. 2014, 155, 925–986. [Google Scholar] [CrossRef]
- Lee, H.J.; Shin, J.S.; Lee, K.G.; Park, S.C.; Jang, Y.P.; Nam, J.H.; Lee, K.T. Ethanol extract of Potentilla supina Linne suppresses LPS-induced inflammatory responses through NF-κB and AP-1 inactivation in macrophages and in endotoxic mice. Phytother. Res. 2017, 31, 475–487. [Google Scholar] [CrossRef] [PubMed]
- Xiao, F.; Xu, T.; Lu, B.; Liu, R. Guidelines for antioxidant assays for food components. Food Front. 2020, 1, 60–69. [Google Scholar] [CrossRef] [Green Version]
- Gangwar, M.; Gautam, M.K.; Sharma, A.K.; Tripathi, Y.B.; Goel, R.; Nath, G. Antioxidant capacity and radical scavenging effect of polyphenol rich Mallotus philippenensis fruit extract on human erythrocytes: An in vitro study. Sci. World J. 2014, 2014, 279451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bibi Sadeer, N.; Montesano, D.; Albrizio, S.; Zengin, G.; Mahomoodally, M.F. The versatility of antioxidant assays in food science and safety—Chemistry, applications, strengths, and limitations. Antioxidants 2020, 9, 709. [Google Scholar] [CrossRef]
- Dawood, D.; Hassan, R.A.; AbdelFattah, S. antioxidant activity evaluation of methanolic extract and crude polysaccharides from Plumeria alba L. leaves. Int. J. Adv. Res. 2016, 4, 1688–1701. [Google Scholar] [CrossRef] [Green Version]
- Prior, R.L.; Wu, X.; Schaich, K. Standardized methods for the determination of antioxidant capacity and phenolics in foods and dietary supplements. J. Agric. Food Chem. 2005, 53, 4290–4302. [Google Scholar] [CrossRef] [PubMed]
- Ren, J.; Chung, S.H. Anti-inflammatory effect of α-linolenic acid and its mode of action through the inhibition of nitric oxide production and inducible nitric oxide synthase gene expression via NF-κB and mitogen-activated protein kinase pathways. J. Agric. Food Chem. 2007, 55, 5073–5080. [Google Scholar] [CrossRef]
- Blondeau, N.; Nguemeni, C.; Debruyne, D.N.; Piens, M.; Wu, X.; Pan, H.; Hu, X.; Gandin, C.; Lipsky, R.H.; Plumier, J.-C. Subchronic alpha-linolenic acid treatment enhances brain plasticity and exerts an antidepressant effect: A versatile potential therapy for stroke. Neuropsychopharmacology 2009, 34, 2548–2559. [Google Scholar] [CrossRef] [Green Version]
- Pan, H.; Hu, X.-z.; Jacobowitz, D.M.; Chen, C.; McDonough, J.; Van Shura, K.; Lyman, M.; Marini, A.M. Alpha-linolenic acid is a potent neuroprotective agent against soman-induced neuropathology. Neurotoxicology 2012, 33, 1219–1229. [Google Scholar] [CrossRef]
- De Lorgeril, M.; Renaud, S.; Salen, P.; Monjaud, I.; Mamelle, N.; Martin, J.; Guidollet, J.; Touboul, P.; Delaye, J. Mediterranean alpha-linolenic acid-rich diet in secondary prevention of coronary heart disease. Lancet 1994, 343, 1454–1459. [Google Scholar] [CrossRef]
- Ando, H.; Ryu, A.; Hashimoto, A.; Oka, M.; Ichihashi, M. Linoleic acid and α-linolenic acid lightens ultraviolet-induced hyperpigmentation of the skin. Arch. Dermatol. Res. 1998, 290, 375–381. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.-E.; Kim, Y.-A.; Yu, S.; Park, S.Y.; Kim, K.H.; Kang, N.J. 3, 6-Anhydro-L-galactose increases hyaluronic acid production via the EGFR and AMPKα signaling pathway in HaCaT keratinocytes. J. Dermatol. Sci. 2019, 96, 90–98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kultti, A.; Kärnä, R.; Rilla, K.; Nurminen, P.; Koli, E.; Makkonen, K.M.; Si, J.; Tammi, M.I.; Tammi, R.H. Methyl-β-cyclodextrin suppresses hyaluronan synthesis by down-regulation of hyaluronan synthase 2 through inhibition of Akt. J. Biol. Chem. 2010, 285, 22901–22910. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pillaiyar, T.; Manickam, M.; Namasivayam, V. Skin whitening agents: Medicinal chemistry perspective of tyrosinase inhibitors. J. Enzyme Inhib. Med. Chem. 2017, 32, 403–425. [Google Scholar] [CrossRef] [Green Version]
- Lim, Y.-J.; Lee, E.H.; Kang, T.H.; Ha, S.K.; Oh, M.S.; Kim, S.M.; Yoon, T.-J.; Kang, C.; Park, J.-H.; Kim, S.Y. Inhibitory effects of arbutin on melanin biosynthesis of α-melanocyte stimulating hormone-induced hyperpigmentation in cultured brownish guinea pig skin tissues. Arch. Pharm. Res. 2009, 32, 367–373. [Google Scholar] [CrossRef]
- Farage, M.; Miller, K.; Elsner, P.; Maibach, H. Intrinsic and extrinsic factors in skin ageing: A review. Int. J. Cosmet. Sci. 2008, 30, 87–95. [Google Scholar] [CrossRef]
- Zhang, S.; Duan, E. Fighting against skin aging: The way from bench to bedside. Cell Transplant. 2018, 27, 729–738. [Google Scholar] [CrossRef]
- Chung, J.H.; Yano, K.; Lee, M.K.; Youn, C.S.; Seo, J.Y.; Kim, K.H.; Cho, K.H.; Eun, H.C.; Detmar, M. Differential effects of photoaging vs intrinsic aging on the vascularization of human skin. Arch. Dermatol. 2002, 138, 1437–1442. [Google Scholar] [CrossRef]
- Kim, Y.-W.; Lee, S.-H. Current status of medicinal herbs recorded from the records in Gongju city, Chungchengnam Province, Korea. Kor. Herb. Inf. 2013, 1, 77–82. [Google Scholar]
- Rinnerthaler, M.; Bischof, J.; Streubel, M.K.; Trost, A.; Richter, K. Oxidative stress in aging human skin. Biomolecules 2015, 5, 545–589. [Google Scholar] [CrossRef] [Green Version]
- Shi, P.; Du, W.; Wang, Y.; Teng, X.; Chen, X.; Ye, L. Total phenolic, flavonoid content, and antioxidant activity of bulbs, leaves, and flowers made from Eleutherine bulbosa (Mill.) Urb. Food Sci. Nutr. 2019, 7, 148–154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, H.-Y.; Agrahari, G.; Lee, M.J.; Tak, L.-J.; Ham, W.-K.; Kim, T.-Y. Low-temperature argon plasma regulates skin moisturizing and melanogenesis-regulating markers through Yes-associated protein. Int. J. Mol. Sci. 2021, 22, 1895. [Google Scholar] [CrossRef] [PubMed]
- Skoczyńska, A.; Budzisz, E.; Trznadel-Grodzka, E.; Rotsztejn, H. Melanin and lipofuscin as hallmarks of skin aging. Postepy Dermatol. Alergol. 2017, 34, 97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boo, Y.C. Arbutin as a skin depigmenting agent with antimelanogenic and antioxidant properties. Antioxidants 2021, 10, 1129. [Google Scholar] [CrossRef]
- Uchida, R.; Ishikawa, S.; Tomoda, H. Inhibition of tyrosinase activity and melanine pigmentation by 2-hydroxytyrosol. Acta Pharm. Sin. B 2014, 4, 141–145. [Google Scholar] [CrossRef] [Green Version]
- Keyes, B.E.; Liu, S.; Asare, A.; Naik, S.; Levorse, J.; Polak, L.; Lu, C.P.; Nikolova, M.; Pasolli, H.A.; Fuchs, E. Impaired epidermal to dendritic T cell signaling slows wound repair in aged skin. Cell 2016, 167, 1323–1338.e14. [Google Scholar] [CrossRef] [Green Version]
- Levine, J.M. Clinical aspects of aging skin: Considerations for the wound care practitioner. Adv. Skin Wound Care 2020, 33, 12–19. [Google Scholar] [CrossRef]
- Ashcroft, G.S.; Mills, S.J.; Ashworth, J.J. Ageing and wound healing. Biogerontology 2002, 3, 337–345. [Google Scholar] [CrossRef]
- Wong, P.; Coulombe, P.A. Loss of keratin 6 (K6) proteins reveals a function for intermediate filaments during wound repair. J. Cell Biol. 2003, 163, 327–337. [Google Scholar] [CrossRef] [Green Version]
- Lee, D.; Lim, J.; Woo, K.-C.; Kim, K.-T. Piperonylic acid stimulates keratinocyte growth and survival by activating epidermal growth factor receptor (EGFR). Sci. Rep. 2018, 8, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Dunnill, C.; Patton, T.; Brennan, J.; Barrett, J.; Dryden, M.; Cooke, J.; Leaper, D.; Georgopoulos, N.T. Reactive oxygen species (ROS) and wound healing: The functional role of ROS and emerging ROS-modulating technologies for augmentation of the healing process. Int. Wound J. 2017, 14, 89–96. [Google Scholar] [CrossRef] [PubMed]
- Jeong, D.; Qomaladewi, N.P.; Lee, J.; Park, S.H.; Cho, J.Y. The role of autophagy in skin fibroblasts, keratinocytes, melanocytes, and epidermal stem cells. J. Investig. Dermatol. 2020, 140, 1691–1697. [Google Scholar] [CrossRef] [PubMed]
- You, L.; Cho, J.Y. The regulatory role of Korean ginseng in skin cells. J. Ginseng Res. 2021, 45, 363–370. [Google Scholar] [CrossRef]
- Kowalik, K.; Paduch, R.; Strawa, J.W.; Wiater, A.; Wlizlo, K.; Wasko, A.; Wertel, I.; Pawlowska, A.; Tomczykowa, M.; Tomczyk, M. Potentilla alba extracts affect the viability and proliferation of non-cancerous and cancerous colon human epithelial cells. Molecules 2020, 25, 3080. [Google Scholar] [CrossRef]
- Choi, S.I.; Lee, J.S.; Lee, S.; Sim, W.S.; Kim, Y.C.; Lee, O.H. Potentilla rugulosa Nakai extract attenuates bisphenol A-, S- and F-induced ROS production and dfferentiation of 3T3-L1 peadipocytes in the asence of dxamethasone. Antioxidants 2020, 9, 113. [Google Scholar] [CrossRef] [Green Version]
- Walia, M.; Kumar, P.; Singh, B.; Agnihotri, V.K. UPLC-DAD quantification of chemical constituents of Potentilla atrosanguinea roots and their antioxidant activity. J. Food Sci. Technol. 2018, 55, 4337–4345. [Google Scholar] [CrossRef] [PubMed]
- Kalia, K.; Sharma, K.; Singh, H.P.; Singh, B. Effects of extraction methods on phenolic contents and antioxidant activity in aerial parts of Potentilla atrosanguinea Lodd. and quantification of its phenolic constituents by RP-HPLC. J. Agric. Food Chem. 2008, 56, 10129–10134. [Google Scholar] [CrossRef]
- Kaltalioglu, K.; Balabanli, B.; Coskun-Cevher, S. Phenolic, antioxidant, antimicrobial, and in-vivo wound healing properties of Potentilla erecta L. root extract in diabetic rats. Iran J. Pharm. Res. 2020, 19, 264–274. [Google Scholar]
- Tomovic, M.T.; Cupara, S.M.; Popovic-Milenkovic, M.T.; Ljujic, B.T.; Kostic, M.J.; Jankovic, S.M. Antioxidant and anti-inflammatory activity of Potentilla reptans L. Acta Pol. Pharm. 2015, 72, 137–145. [Google Scholar]
- Kurnia, D.; Ajiati, D.; Heliawati, L.; Sumiarsa, D. Antioxidant properties and structure-antioxidant activity relationship of Allium species leaves. Molecules 2021, 26, 7175. [Google Scholar] [CrossRef]
- Syrpas, M.; Subbarayadu, K.; Kitryte, V.; Venskutonis, P.R. High-pressure extraction of antioxidant-rich fractions from Shrubby Cinquefoil (Dasiphora fruticosa L. Rydb.) leaves: Process optimization and extract characterization. Antioxidants 2020, 9, 457. [Google Scholar] [CrossRef] [PubMed]
- Singleton, V.L.; Rossi, J.A. Colorimetry of total phenolics with phosphomolybdic-phosphotungstic acid reagents. Am. J. Enol. Vitic. 1965, 16, 144–158. [Google Scholar]
- Zhishen, J.; Mengcheng, T.; Jianming, W. The determination of flavonoid contents in mulberry and their scavenging effects on superoxide radicals. Food Chem. 1999, 64, 555–559. [Google Scholar] [CrossRef]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef]
- Choi, W.; Kim, H.S.; Park, S.H.; Kim, D.; Hong, Y.D.; Kim, J.H.; Cho, J.Y. Syringaresinol derived from Panax ginseng berry attenuates oxidative stress-induced skin aging via autophagy. J. Ginseng Res. 2021. In press. [Google Scholar] [CrossRef]
- Brand-Williams, W.; Cuvelier, M.E.; Berset, C. Use of a free radical method to evaluate antioxidant activity. Food Sci. Technol. 1995, 28, 25–30. [Google Scholar] [CrossRef]
- Hahm, K.M.; Park, S.H.; Oh, S.W.; Kim, J.H.; Yeom, H.S.; Lee, H.J.; Yang, S.; Cho, J.Y.; Park, J.O.; Lee, J. Aspergillus oryzae-fermented wheat peptone enhances the potential of proliferation and hydration of human keratinocytes through activation of p44/42 MAPK. Molecules 2021, 26, 6074. [Google Scholar] [CrossRef]
- Benzie, I.F.; Strain, J.J. The ferric reducing ability of plasma (FRAP) as a measure of “antioxidant power”: The FRAP assay. Anal. Biochem. 1996, 239, 70–76. [Google Scholar] [CrossRef] [Green Version]
- Apak, R.; Güçlü, K.; Özyürek, M.; Karademir, S.E. Novel total antioxidant capacity index for dietary polyphenols and vitamins C and E, using their cupric ion reducing capability in the presence of neocuproine: CUPRAC method. J. Agric. Food Chem. 2004, 52, 7970–7981. [Google Scholar] [CrossRef]
- Lee, J.O.; Hwang, S.H.; Shen, T.; Kim, J.H.; You, L.; Hu, W.; Cho, J.Y. Enhancement of skin barrier and hydration-related molecules by protopanaxatriol in human keratinocytes. J. Ginseng Res. 2021, 45, 354–360. [Google Scholar] [CrossRef]
- Jeong, D.; Lee, J.; Park, S.H.; Kim, Y.A.; Park, B.J.; Oh, J.; Sung, G.-H.; Aravinthan, A.; Kim, J.-H.; Kang, H.; et al. Antiphotoaging and antimelanogenic effects of Penthorum chinense Pursh ethanol extract due to antioxidant- and autophagy-Inducing properties. Oxid. Med. Cell. Longev. 2019, 2019, 9679731. [Google Scholar] [CrossRef] [PubMed]
- Park, S.H.; Kim, D.S.; Kim, S.; Lorz, L.R.; Choi, E.; Lim, H.Y.; Hossain, M.A.; Jang, S.; Choi, Y.I.; Park, K.J.; et al. Loliolide presents antiapoptosis and antiscratching effects in human keratinocytes. Int. J. Mol. Sci. 2019, 20, 651. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lorz, L.R.; Yoo, B.C.; Kim, M.-Y.; Cho, J.Y. Anti-wrinkling and anti-melanogenic effect of Pradosia mutisii methanol extract. Int. J. Mol. Sci. 2019, 20, 1043. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aziz, N.; Son, Y.-J.; Cho, J.Y. Thymoquinone suppresses IRF-3-mediated expression of type I interferons via suppression of TBK1. Int. J. Mol. Sci. 2018, 19, 1355. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.O.; Kim, J.H.; Kim, S.; Kim, M.Y.; Hong, Y.H.; Kim, H.G.; Cho, J.Y. Gastroprotective effects of the nonsaponin fraction of Korean Red Ginseng through cyclooxygenase-1 upregulation. J. Ginseng Res. 2020, 44, 655–663. [Google Scholar] [CrossRef]
- You, L.; Kim, M.-Y.; Cho, J.Y. Protective Effect of Potentilla glabra in UVB-Induced Photoaging Process. Molecules 2021, 26, 5408. [Google Scholar] [CrossRef]
- Kim, J.K.; Shin, K.K.; Kim, H.; Hong, Y.H.; Choi, W.; Kwak, Y.S.; Han, C.K.; Hyun, S.H.; Cho, J.Y. Korean Red Ginseng exerts anti-inflammatory and autophagy-promoting activities in aged mice. J. Ginseng Res. 2021, 45, 717–725. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Extract | TPC 1 | TFC 2 |
|---|---|---|
| Pp-EE | 273.26 ± 2.28 | 102.08 ± 2.95 |
| Peak No. | RT 1 | Name of the Compound | Type of the Compound | Peak Area % |
|---|---|---|---|---|
| 1 | 4.680 | Dimethyl sulfone | Sulfoxide | 1.140 |
| 2 | 5.440 | 4H-Pyran-4-one, 2,3-dihydro-3,5-dihydroxy-6-methyl- | Pyrone | 1.333 |
| 3 | 7.569 | 2-Propanamine, N-methyl-N-nitroso- | - | 1.292 |
| 4 | 7.705 | 4H-Pyran-4-one, 2,3-dihydro-3,5-dihydroxy-6-methyl- | Pyrone | 6.864 |
| 5 | 8.687 | Benzofuran, 2,3-dihydro- | Benzofuran | 1.332 |
| 6 | 8.829 | 5-Hydroxymethylfurfural | Alcohol | 10.547 |
| 7 | 9.035 | 1,2,3-Propanetriol, 1-acetate | Glyceride | 3.347 |
| 8 | 9.452 | Methyl 4-(methylthio)butyrate | Fatty ester | 2.007 |
| 9 | 9.530 | 2,4-Dihydroxy-2,5-dimethyl-3(2H)-furan-3-one | - | 2.249 |
| 10 | 10.092 | 2-Methoxy-4-vinylphenol | Phenol | 2.244 |
| 11 | 11.682 | Benzaldehyde, 3-methoxy- | Benzaldehyde | 8.468 |
| 12 | 13.348 | Methyl-.beta.-D-thiogalactoside | Glucoside | 6.937 |
| 13 | 13.536 | D-erythro-Pentose, 2-deoxy- | Carbohydrate | 2.028 |
| 14 | 16.608 | 1,2-Benzenedicarboxylic acid, bis(2-methylpropyl) ester | Organic acid | 0.761 |
| 15 | 17.436 | n-Hexadecanoic acid | Fatty Acid | 8.383 |
| 16 | 17.549 | Dibutyl phthalate | Organic acid | 1.411 |
| 17 | 18.926 | Phytol | Diterpenoid | 3.241 |
| 18 | 19.088 | 9,12-Octadecadienoic acid (Z,Z)- | Fatty acid | 2.894 |
| 19 | 19.161 | 9,12,15-Octadecatrienoic acid, (Z,Z,Z)- | Fatty acid | 20.397 |
| 20 | 19.319 | Octadecanoic acid | Fatty acid | 3.276 |
| 21 | 19.436 | 9,12,15-Octadecatrienoic acid, ethyl ester, (Z,Z,Z)- | Fatty acid | 2.564 |
| 22 | 22.618 | Bis(2-ethylhexyl) phthalate | Organic acid | 1.225 |
| 23 | 23.730 | Linolenic acid, 2-hydroxy-1-(hydroxymethyl)ethyl ester (Z,Z,Z)- | Glyceride | 1.369 |
| 24 | 23.951 | 1,2,4-Triazol-3-amine, 5-(1,3,5-trimethyl-4-pyrazolyl)amino- | Triazole | 1.352 |
| 25 | 27.161 | Vitamin E | Tocopherol | 1.424 |
| 26 | 29.385 | Hexamethylcyclotrisiloxane | Siloxane | 1.913 |
| PCR Type | Gene Name | Sequence (5′–3′) | |
|---|---|---|---|
| RT-PCR | K17 | Forward | CATGCAGGCCTTGGAGATAGA |
| Reverse | CACGCAGTAGCGGTTCTCTGT | ||
| GAPDH | Forward | CACTCACGGCAAATTCAACGGCAC | |
| Reverse | GACTCCACGACATACTCAGCAC | ||
| Real-time PCR | HAS-1 | Forward | TGTATCCTGCATCAGCGGTC |
| Reverse | GCCGGTCATCCCCAAAAGTA | ||
| HAS-2 | Forward | GTGGATGACCTACGAAGCGA | |
| Reverse | TACCCCGGTAGAAGAGCTGG | ||
| HAS-3 | Forward | TTGGCCTCATTCCTGTGTCC | |
| Reverse | CTGGCAATAAGCTGTGTAGGC | ||
| Hyal-1 | Forward | TGTGGACGTGGATGTCAGTG | |
| Reverse | GTAGTAGGGGTAGGTGCCCA | ||
| Hyal-2 | Forward | ATGTGCAGAACTGGGAGAGC | |
| Reverse | GGAAGCAAGTGTCTCGTCCA | ||
| Hyal-3 | Forward | TCTGGGCATCATAGCCAACC | |
| Reverse | AGAGGCCGAGTTGGTTCTTG | ||
| Hyal-4 | Forward | AACTGCATCCAAGGCCAACT | |
| Reverse | CTCAGCAGCTCTGGTCACAT | ||
| MITF | Forward | TCCGTTTCTTCTGCGCTCAT | |
| Reverse | CTGATGGACGATGCCCTCTC | ||
| Tyrosinase | Forward | GTCCACTCACAGGGATAGCAG | |
| Reverse | AGAGTCTCTGTTATGGCCGA | ||
| TYRP-1 | Forward | ATGGAACGGGAGGACAAACC | |
| Reverse | TCCTGACCTGGCCATTGAAC | ||
| TYRP-2 | Forward | CAGTTTCCCCGAGTCTGCAT | |
| Reverse | GTCTAAGGCGCCCAAGAACT | ||
| GAPDH (Human) | Forward | GACAGTCAGCCGCATCTTCT | |
| Reverse | GCGCCAATACGACCAAATC | ||
| GAPDH (Mouse) | Forward | TGTGAACGGATTTGGCCGTA | |
| Reverse | ACTGTGCCGTTGAATTTGCC | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, H.P.; Kim, D.S.; Park, S.H.; Shin, C.Y.; Woo, J.J.; Kim, J.W.; An, R.-B.; Lee, C.; Cho, J.Y. Antioxidant Capacity of Potentilla paradoxa Nutt. and Its Beneficial Effects Related to Anti-Aging in HaCaT and B16F10 Cells. Plants 2022, 11, 873. https://doi.org/10.3390/plants11070873
Lee HP, Kim DS, Park SH, Shin CY, Woo JJ, Kim JW, An R-B, Lee C, Cho JY. Antioxidant Capacity of Potentilla paradoxa Nutt. and Its Beneficial Effects Related to Anti-Aging in HaCaT and B16F10 Cells. Plants. 2022; 11(7):873. https://doi.org/10.3390/plants11070873
Chicago/Turabian StyleLee, Hwa Pyoung, Dong Seon Kim, Sang Hee Park, Chae Yun Shin, Jin Joo Woo, Ji Won Kim, Ren-Bo An, Changyoung Lee, and Jae Youl Cho. 2022. "Antioxidant Capacity of Potentilla paradoxa Nutt. and Its Beneficial Effects Related to Anti-Aging in HaCaT and B16F10 Cells" Plants 11, no. 7: 873. https://doi.org/10.3390/plants11070873