
Unexpected Discovery of Thelypteris palustris (Thelypteridaceae) in Sicily (Italy): Morphological, Ecological Analysis and Habitat Characterization

 ,
,  and
and
Abstract
:1. Introduction
2. Results and Discussion
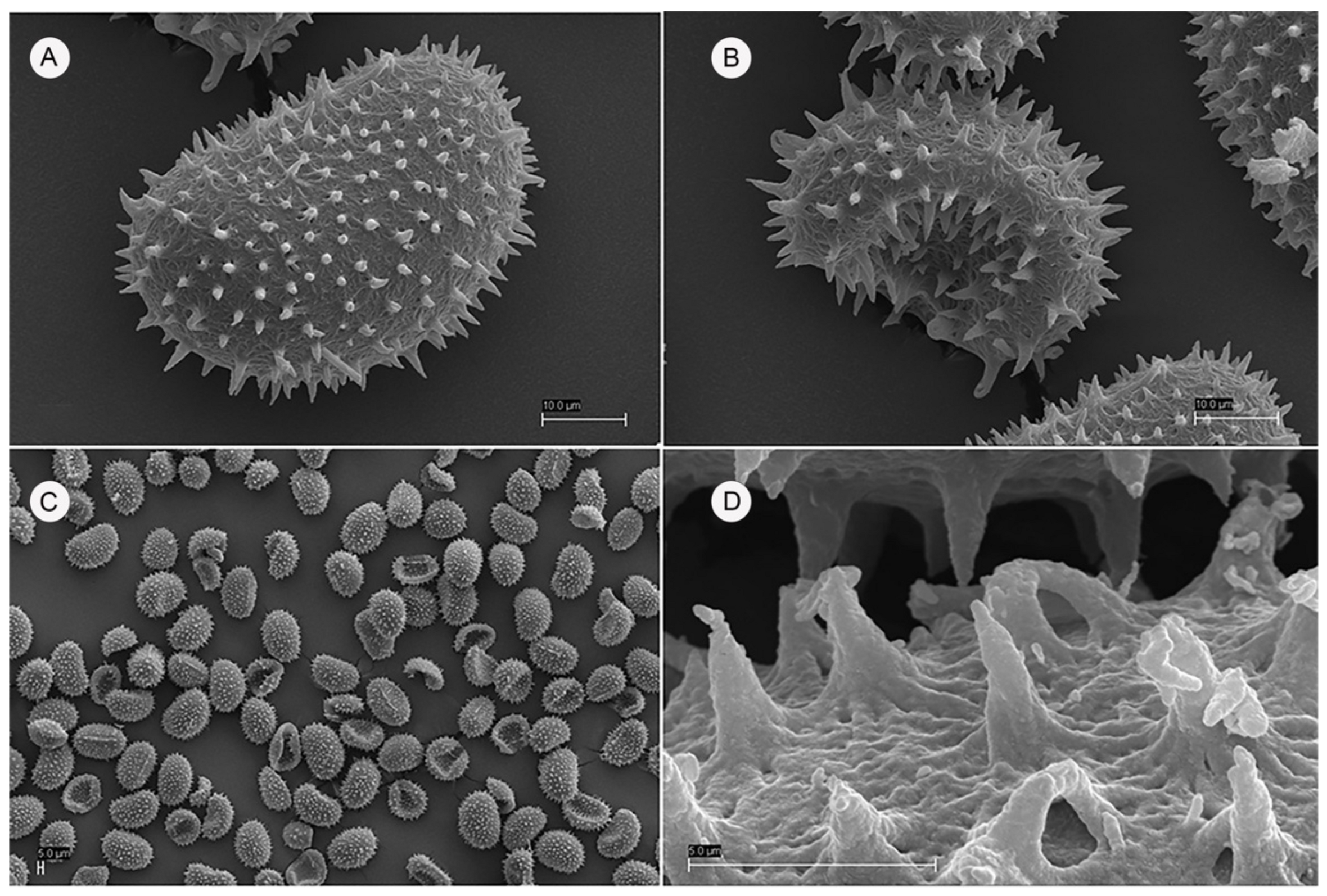
2.1. Description of the Species (Based on the New Population)
2.2. Distribution and Conservation Status in Italy
2.3. Plant Communities with T. palustris in Italy
2.4. Vegetation Ecology and Habitat
2.5. Floristic Composition and Phytosociological Insights of the Thelypteris palustris Population in Sicily
3. Materials and Methods
3.1. Study Area
3.2. Data Sets and Data Processing
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A. Phytosociological Survey of Aquatic and Marsh Vegetation Syntaxonomical Scheme
- LEMNETEA O. de Bolos et Masclans 1955
- LEMNETALIA MINORIS O. de Bolos et Masclans 1955
- LEMNION MINORIS O. de Bolos et Masclans 1955
- Lemnetum minoris von Soó 1927
- Wolffietum arrhizae Myawaki & J.Tx. 1960
- POTAMOGETONETEA Klika in Klika et Novak 1941
- POTAMOGETONETALIA Koch 1926
- POTAMION (Koch 1926) Libbert 1931
- Myriophylletum verticillati Lemnée 1937
- NINPHAEION ALBAE Oberd. 1957
- Potamogetono natantis-Polygonetum natantis Knapp et Stoffers 1962
- Potametum pusilli von Soó 1927
- Potametum natantis von Soó 1927
- UTRICULARIETALIA Den Hartog & Segal 1964
- UTRICULARION VULGARIS Passarge 1964
- Utricularietum australis Müller & Görs 1960
- CERATOPHYLLION DEMERSI Den Hartog & Segal ex Passarge 1996
- Potamogetono-Ceratophylletum submersi Pop 1962
- RANUNCULION AQUATILIS Géhu 1961
- Ranunculetum aquatilis Géhu 1961
- RANUNCULION OMIOPHYLLO-HEDERACEI Rivas-Martínez et al. 2002
- Ranunculetum omiophylli Br.-Bl. & Tüxen ex Pizarro 1995
- PHRAGMITO-MAGNOCARICETEA Klika in Klika et Novak 1941
- PHRAGMITETALIA Koch 1926
- PHRAGMITION COMMUNIS Koch 1926
- Sparganietum erecti Philippi 1973
- Scirpetum lacustris Schmale 1939
- Typhetum domingensis Brullo, Minissale & Spamp. 1994
- MAGNOCARICETALIA Pignatti 1953
- MAGNOCARICION ELATAE Koch 1926
- Galio palustris-Juncetum inflexi Venanzoni et Gigante 2000
- Eleocharitetum palustris Savic 1926
- Iridetum pseudacori Krywanski 1974
- CARICION GRACILIS Géhu 1961
- Thelypterido palustris-Caricetum paniculatae ass. nova hoc loco
- OENANTHETALIA AQUATICAE Hejny ex Balatova-Tulackova et al. 1993
- ALOPECURO-GLYCERION SPICATAE Brullo, Minissale, Spamp. 1994
- Glycerio spicatae-Oenanthetum aquaticae Brullo, Minissale & Spamp. 1994
- ISOËTO-NANOJUNCETEA Br.-Bl. et Tx. in Br.-Bl. et al. 1952
- Nanocyperetalia K1ika 1935
- Nanocyperionflavescentis W. Koch ex Libbert 1932
- Lythrum portula comm.
References
- Fernald, M.L. A study of Thelypteris palustris. Rhodora 1929, 31, 27–36. [Google Scholar]
- Tryon, A.F. Structure and variation in spores of Thelypteris palustris. Rhodora 1971, 73, 433–460. [Google Scholar]
- Tryon, A.F.; Tryon, R.; Badré, F. Classification, spores, and nomenclature of the marsh fern. Rhodora 1980, 82, 461–474. [Google Scholar]
- Fraser-Jenkins, C.R. New Species Syndrome in Indian Pteridology and the Ferns of Nepal; International Book Distributors: Dehra Dun, India, 1997. [Google Scholar]
- Christenhusz, M.; von Raab-Straube, E. Polypodiopsida. In Euro+Med Plantbase—The Information Resource for Euro-Mediterranean plant Diversity. 2013. Available online: http://ww2.bgbm.org/EuroPlusMed/ (accessed on 9 January 2021).
- Conti, F.; Bartolucci, F.; Iocchi, M.; Tinti, D. Atlas of the pteridological knowledge of Abruzzo (Central Italy). Webbia 2011, 66, 251–305. [Google Scholar] [CrossRef]
- Bovio, M. Flora Vascolare della Valle d’Aosta. In Repertorio Commentato e Stato Delle Conoscenze; Testolin Editore: Sarre, Italy, 2014. [Google Scholar]
- Bartolucci, F.; Peruzzi, L.; Galasso, G.; Albano, A.; Alessandrini, A.; Ardenghi, N.M.G.; Astuti, G.; Bacchetta, G.; Ballelli, S.; Banfi, E.; et al. An updated checklist of the vascular flora native to Italy. Plant Biosyst. 2018, 152, 179–303. [Google Scholar] [CrossRef]
- García Criado, M.; Väre, H.; Nieto, A.; Bento Elias, R.; Dyer, R.; Ivanenko, Y.; Ivanova, D.; Lansdown, R.; Molina, J.A.; Rouhan, G.; et al. European Red List of Lycopods and Ferns; IUCN: Brussels, Belgium, 2017; pp. iv + 59. [Google Scholar]
- Rodwell, J.S. British Plant Communities. In Woodlands and Scrub; Cambridge University Press: Cambridge, UK, 1991. [Google Scholar]
- Angiolini, C.; Ferretti, G.; Foggi, B.; Frignani, F.; Gestri, G.; Landi, M.; Lastrucci, L.; Monacci, F.; Peruzzi, L.; Sani, A.; et al. Segnalazione 51. In Contributi Alla Flora Vascolare di Toscana I (1–85); Peruzzi, L., Viciani, D., Bedini, G., Eds.; Atti. Soc. Tosc. Sci. Nat. Mem. Serie B: Italy, 2009; Volume 116, pp. 33–44. Available online: https://www.researchgate.net/publication/264273119_Contributi_per_una_flora_vascolare_di_Toscana_I_1-85 (accessed on 1 June 2021).
- Lastrucci, L.; Dell’Olmo, L.; Foggi, B.; Massi, L.; Nuccio, C.; Vicenti, C.; Viciani, D. Contribution to the knowledge of the vegetation of the Lake Massaciuccoli (northern Tuscany, Italy). Plant Sociol. 2017, 54, 67–87. [Google Scholar]
- Spampinato, G.; Sciandrello, S.; Giusso del Galdo, G.; Puglisi, M.; Tomaselli, V.; Cannavò, S.; Musarella, C.M. Contribution to the knowledge of Mediterranean wetland biodiversity: Plant communities of the Aquila Lake (Calabria, Southern Italy). Plant Sociol. 2019, 56, 53–68. [Google Scholar]
- Pedrotti, F. Nota sulla flora e vegetazione del Lago di Madrano (Trentino). Inf. Bot. Ital. 1991, 22, 182–193. [Google Scholar]
- Pedrotti, F. Nota sulla vegetazione degli ambienti umidi della bassa Valsugana (Trentino). Doc. Phytosoc. 1995, 15, 417–449. [Google Scholar]
- Tasinazzo, S.; Fiorentin, R. I relitti boschetti ad Alnus glutinosa delle risorgive vicentine (Pianura Veneta). Ann. Mus. Civ. Rovereto 2003, 17, 125–135. [Google Scholar]
- Pedrotti, F. Ricerche geobotaniche al Laghestel di Pin‚ (1967–2001). Braun-Blanquetia 2004, 35, 1–55. [Google Scholar]
- Lonati, M.; Lonati, S. Le comunità a Carex elata All. della torbiera di Vanzone (Piemonte, Vercelli). Fitosociologia 2005, 42, 15–21. [Google Scholar]
- Tisi, A.; Minuzzo, C.; Siniscalco, C.; Caramiello, R. La vegetazione acquatica e palustre della zona dei “Cinque Laghi” di Ivrea. Riv. Piem. St. Nat. 2007, 28, 87–126. [Google Scholar]
- Sburlino, G.; Poldini, L.; Venanzoni, R.; Ghirelli, L. Italian black alder swamps: Their syntaxonomic relationships and originality within the European context. Plant Biosyst. 2011, 145, 148–171. [Google Scholar] [CrossRef]
- Gennai, M.; Gabellini, A.; Viciani, D.; Venanzoni, R.; Dell’Olmo, L.; Giunti, M.; Lucchesi, F.; Monacci, F.; Mugnai, M.; Foggi, B. The floodplain woods of Tuscany: Towards a phytosociological synthesis. Plant Sociol. 2021, 58, 1–28. [Google Scholar] [CrossRef]
- Sciandrello, S.; Cambria, S.; Giusso del Galdo, G.; Guarino, R.; Minissale, P.; Pasta, S.; Tavilla, G.; Cristaudo, A. Floristic and Vegetation Changes on a Small Mediterranean Island over the Last Century. Plants 2021, 10, 680. [Google Scholar] [CrossRef]
- Macchia, F. Vegetazione e flora dei Laghi Alimini. Atti e Relazioni Acc. Pugliese Scienze ns. Cl. Sc. Fis. Med. Nat. 1967, 25, 221–267. [Google Scholar]
- Annese, B.; Beccarisi, L. Thelypteris palustris Schott. In Notule Pteridologiche Italiche, 3 (64–84); Marchetti, D., Ed.; Ann. Mus. Civ. Rovereto Sez. Arch. St. Sc. Nat.: Sardinia, Italy, 2003; Volume 18, pp. 65–81. Available online: http://www.museocivico.rovereto.tn.it/UploadDocs/202_C__WINDOWS_Desktop_annali_pdf_18_art04_marchetti.pdf (accessed on 1 June 2021).
- Crisafulli, A.; Cannavò, S.; Maiorca, G.; Musarella, C.M.; Signorino, G.; Spampinato, G. Aggiornamenti floristici per la Calabria. Inform. Bot. Ital. 2010, 42, 431–442. [Google Scholar]
- Bilz, M.; Kell, S.P.; Maxted, N.; Lansdown, R.V. European Red List of Vascular Plants; Publications Office of the European Union: Luxembourg, 2011; p. 130. [Google Scholar]
- Orsenigo, S.; Fenu, G.; Gargano, D.; Montagnani, C.; Abeli, T.; Alessandrini, A.; Bacchetta, G.; Bartolucci, F.; Carta, A.; Castello, M.; et al. Red list of threatened vascular plants in Italy. Plant Biosyst. 2021, 155, 310–335. [Google Scholar] [CrossRef]
- IUCN. Guidelines for Using the IUCN Red List Categories and Criteria, 14th ed.; IUCN Standards and Petitions Committee: Gland, Switzerland, 2019; p. 113. Available online: http://cmsdocs.s3.amazonaws.com/RedListGuidelines.pdf (accessed on 1 January 2021).
- Guglielmetto Mugion, L.; Montacchini, F. La vegetazione del Lago di Viverone. Allionia 1994, 32, 1–26. [Google Scholar]
- Poldini, L.; Vidali, M.; Castello, M.; Sburlino, G. A novel insight into the remnants of hygrophilous forests and scrubs of the Po Plain biogeographical transition area (Northern Italy). Plant Sociol. 2020, 57, 17–69. [Google Scholar] [CrossRef]
- Gellini, R.; Pedrotti, F.; Venanzoni, R. Le associazioni forestali ripariali e palustri della Selva di San Rossore (Pisa). Doc. Phytosoc. n.s. 1986, 10, 27–41. [Google Scholar]
- Brullo, S.; Minissale, P.; Spampinato, G. Studio fitosociologico della vegetazione lacustre dei Monti Nebrodi (Sicilia settentrionale). Fitosociologia 1994, 27, 5–50. [Google Scholar]
- Biondi, E.; Blasi, C.; Allegrezza, M.; Anzellotti, I.; Azzella, M.M.; Carli, E.; Casavecchia, S.; Copiz, R.; Del Vico, E.; Facioni, L.; et al. Plant communities of Italy: The Vegetation Prodrome. Plant Biosyst. 2014, 148, 728–814. [Google Scholar] [CrossRef] [Green Version]
- Brullo, S.; Grillo, M. Ricerche fitosociologiche sui pascoli dei monti Nebrodi (Sicilia settentrionale). Not. Fitosoc. 1978, 13, 23–61. [Google Scholar]
- Brullo, S.; Scelsi, F.; Spampinato, G. La vegetazione dell’Aspromonte. In Studio Fitosociologico; Laruffa: Reggio Calabria, Italy, 2001; p. 370. [Google Scholar]
- Landucci, F.; Gigante, D.; Venanzoni, R.; Chytrý, M. Wetland vegetation of the class Phragmito-Magno-Caricetea in central Italy. Phytocoenologia 2013, 43, 67–100. [Google Scholar] [CrossRef] [Green Version]
- Ciaschetti, G.; Pirone, G.; Venanzoni, R. Sedge vegetation of the ‘Major Highlands of Abruzzo’ (Central Italy): Updated knowledge after new discoveries. Plant Biosyst. 2021, 155, 647–662. [Google Scholar] [CrossRef]
- Bazan, G.; Marino, P.; Guarino, R.; Domina, G.; Schicchi, R. Bioclimatology and vegetation series in Sicily: A geostatistical approach. Ann. Bot. Fenn. 2015, 52, 1–18. [Google Scholar] [CrossRef]
- Brullo, C.; Brullo, S.; Giusso del Galdo, G.; Guarino, R.; Siracusa, G.; Sciandrello, S. The class Querco-Fagetea sylvaticae in Sicily: An example of boreal-temperate vegetation in the central mediterranean area. Ann. Bot. 2012, 2, 19–38. [Google Scholar]
- Henriquet, M.; Dominguez, S.; Barreca, G.; Malavieille, J.; Monaco, C. Structural and tectono-stratigraphic review of the Sicilian orogen and new insights from analogue modeling. Earth-Sci. Rev. 2020, 208, 103257. [Google Scholar] [CrossRef]
- Lentini, F.; Carbone, S. Geologia della Sicilia—Geology of Sicily. Mem. Descr. Carta Geol. D’italia 2014, 95, 7–414. [Google Scholar]
- Rivas-Martínez, S.; Penas, A.; Diaz, T.E. Biogeographic Map of Europe; Cartographic Service, University of Léon: León, Spain, 2004. [Google Scholar]
- Bisculm, M.; Colombaroli, D.; Vescovi, E.; van Leeuwen, J.F.N.; Henne, P.D.; Rothen, J.; Procacci, G.; Pasta, S.; La Mantia, T.; Tinner, W. Holocene vegetation and fire dynamics in the supra-mediterranean belt of the Nebrodi Mountains (Sicily, Italy). J. Quat. Sci. 2012, 27, 687–698. [Google Scholar] [CrossRef]
- Signorello, G.; Prato, C.; Marzo, A.; Ientile, R.; Cucuzza, G.; Sciandrello, S.; Martínez-López, J.; Balbi, S.; Villa, F. Are protected areas covering important biodiversity sites? An assessment of the nature protection network in Sicily (Italy). Land Use Policy 2018, 78, 593–602. [Google Scholar] [CrossRef]
- Lellinger, D.B. A Modern Multilingual Glossary for Taxonomic Pteridology. Pteridologia 2002, 3, 1–263. [Google Scholar]
- Tryon, A.F.; Lugardon, B. Spores of Pteridophyta: Surface, Wall Structure and Diversity Based on Electron Microscope Studies; Springer: New York, NY, USA, 1991; p. 648. [Google Scholar]
- Westhoff, V.; van der Maarel, E. The Braun-Blanquet approach. In Classification of Plant Communities, 2nd ed.; Whittaker, R.H., Ed.; Junk: The Hague, The Netherlands, 1978; pp. 287–297. [Google Scholar]
- Gargano, D. Verso la redazione di nuove Liste Rosse della flora d’Italia: Una griglia standard per la misura dell’Area of Occupancy (AOO). Inform. Bot. Ital. 2011, 43, 35–38. [Google Scholar]
- Mucina, L.; Bültmann, H.; Dierßen, K.; Theurillat, J.-P.; Raus, T.; Čarni, A.; Šumberová, K.; Willner, W.; Dengler, J.; García, R.G.; et al. Vegetation of Europe: Hierarchical floristic classification system of vascular plant, bryophyte, lichen, and algal communities. Appl. Veg. Sci. 2016, 19, 3–264. [Google Scholar] [CrossRef]
- Pignatti, S. Volume 1: Flora d’Italia & Flora Digitale. In Flora d’Italia: In 4 Volumi, 2nd ed.; Edagricole-Edizioni Agricole di New Business Media srl: Milano, Italy, 2017. [Google Scholar]
- Pignatti, S. Volume 2: Flora d’Italia & Flora Digitale. In Flora d’Italia: In 4 Volumi, 2nd ed.; Edagricole-Edizioni Agricole di New Business Media srl: Milano, Italy, 2017. [Google Scholar]
- Pignatti, S. Volume 3: Flora d’Italia & Flora Digitale. In Flora d’Italia: In 4 Volumi, 2nd ed.; Edagricole-Edizioni Agricole di New Business Media srl: Milano, Italy, 2018. [Google Scholar]
- Pignatti, S.; Guarino, R.; La Rosa, M. Volume 4: Flora d’Italia & Flora Digitale. In Flora d’Italia: In 4 Volumi, 2nd ed.; Edagricole-Edizioni Agricole di New Business Media srl: Milano, Italy, 2019. [Google Scholar]
- Rull, V. Microrefugia. J. Biogeogr. 2009, 36, 481–484. [Google Scholar] [CrossRef]
- Cannarozzo, M.; Noto, L.V.; Viola, F. Spatial distribution of rainfall trends in Sicily (1921–2000). Phys. Chem. Earth 2006, 31, 1201–1211. [Google Scholar] [CrossRef]
- Arnone, E.; Pumo, D.; Viola, F.; Noto, L.V.; La Loggia, G. Rainfall statistics changes in Sicily. Hydrol. Earth Syst. Sci. 2013, 17, 2449–2458. [Google Scholar] [CrossRef] [Green Version]
- Cuena-Lombraña, A.; Fois, M.; Cogoni, A.; Bacchetta, G. Where we Come from and where to Go: Six Decades of Botanical Studies in the Mediterranean Wetlands, with Sardinia (Italy) as a Case Study. Wetlands 2021, 41, 69. [Google Scholar] [CrossRef]
- Kavak, S. Thelypteris palustris. The IUCN Red List of Threatened Species 2014: e.T164136A42331187. Available online: https://dx.doi.org/10.2305/IUCN.UK.2014-1.RLTS.T164136A42331187.en (accessed on 1 January 2021).
- Daoud-Bouattour, A.; Muller, S.D.; Ferchichi-Ben Jamaa, H.; Ghrabi-Gammar, Z.; Rhazi, L.; Mokhtar Gammar, A.; Raouf Karray, M.; Soulié-Märsche, I.; Zouaïdia, A.; de Bélair, G.; et al. Recent discovery of the small pillwort (Pilularia minuta Durieu, Marsileaceae) in Tunisia: Hope for an endangered emblematic species of Mediterranean temporary pools? Comptes Rendus Biol. 2009, 332, 886–897. [Google Scholar] [CrossRef]
- Gianguzzi, L.; D’Amico, A.; Troia, A. Notes on the distribution, ecology and conservation status of two very rare sedges (Carex, Cyperaceae) rediscovered in Sicily (Italy). Bot. Lett. 2017, 164, 339–349. [Google Scholar] [CrossRef]
- Minissale, P.; Sciandrello, S. Ecological features affect patterns of plant communities in Mediterranean temporary rock pools. Plant Biosyst. 2016, 150, 171–179. [Google Scholar] [CrossRef]
- Minissale, P.; Santo, A.; Sciandrello, S. Analisi geobotanica del SIC: Capo Murro di Porco, Penisola della Maddalena e Grotta Pellegrino (Siracusa, Sicilia). Fitosociologia 2011, 48, 77–98. [Google Scholar]
- Minissale, P.; Molina, J.A.; Sciandrello, S. Pilularia minuta Durieu (Marsileaceae) discovered in south-eastern-Sicily: New insights on its ecology, distribution and conservation status. Bot. Lett. 2017, 164, 197–208. [Google Scholar] [CrossRef]
- Sciandrello, S.; Puglisi, M.; Privitera, M.; Minissale, P. Diversity and spatial patterns of plant communities in volcanic temporary ponds of Sicily (Italy). Biologia 2016, 71, 793–803. [Google Scholar] [CrossRef]
- Gazaix, A.; Klesczewski, M.; Bouchet, M.A.; Cartereau, M.; Molina, J.; Michaud, H.; Muller, S.D.; Pirsoul, L.; Gauthier, P.; Grillas, P.; et al. A history of discoveries and disappearances of the rare annual plant Lythrum thesioides M. Bieb.: New insights into its ecology and biology. Bot. Lett. 2020, 167, 201–211. [Google Scholar] [CrossRef]
- Taylor, N.G.; Grillas, P.; Al Hreisha, H.; Balkız, Ö.; Borie, M.; Boutron, O.; Catita, A.; Champagnon, J.; Cherif, S.; Çiçek, K.; et al. The future for Mediterranean wetlands: 50 key issues and 50 important conservation research questions. Reg. Environ. Chang. 2021, 21, 1–17. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pools | 18 | 18 | 18 | 18 | 18 | 18 | 18 | 18 | 18 | 18 | 18 | 18 | 18 | 31 | |||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Relevé number | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | |||
| Altitude (m a.s.l.) | 1000 | 1000 | 1000 | 1000 | 1000 | 1030 | 1030 | 1028 | 1028 | 1028 | 1000 | 1000 | 1028 | 1057 | |||
| Surface (mq) | 50 | 50 | 50 | 50 | 50 | 50 | 50 | 50 | 50 | 50 | 50 | 50 | 50 | 50 | |||
| Coverage(%) | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | |||
| Slope (°) | - | - | - | - | - | - | - | 5 | 5 | 5 | 5 | 5 | 5 | - | |||
| Aspect | - | - | - | - | - | - | - | W | W | W | W | W | W | - | |||
| Vegetation height (m) | 1.5 | 1.3 | 1.3 | 14 | 1.5 | 1.5 | 1.3 | 1.3 | 1.4 | 1.4 | 1.4 | 1.3 | 1.2 | 1.5 | |||
| No. species | 18 | 19 | 20 | 19 | 20 | 25 | 27 | 23 | 24 | 27 | 28 | 24 | 25 | 23 | |||
| Simpson_1-D | 0.9 | 0.9 | 0.9 | 0.9 | 0.9 | 0.95 | 0.95 | 0.95 | 0.95 | 0.95 | 0.96 | 0.95 | 0.95 | 0.95 | |||
| Shannon_H | 2.8 | 2.8 | 2.9 | 2.9 | 2.9 | 3.1 | 3.2 | 3.1 | 3.1 | 3.2 | 3.2 | 3.1 | 3.1 | 3.1 | |||
| Evenness_e^H/S | 0.9 | 0.9 | 0.9 | 0.9 | 0.9 | 0.9 | 0.9 | 0.9 | 0.9 | 0.89 | 0.9 | 0.9 | 0.9 | 0.9 | |||
| Equitability_J | 0.96 | 0.96 | 0.97 | 0.97 | 0.97 | 0.97 | 0.97 | 0.97 | 0.97 | 0.96 | 0.97 | 0.96 | 0.97 | 0.97 | presence | ||
| LF | Corology | Characteristic species | |||||||||||||||
| G | Subcosmop | Thelypteris palustris Schott subsp. palustris | 2 | 1 | 2 | 1 | 2 | + | + | 2 | 2 | 3 | 3 | 2 | 1 | - | 13 |
| G | Circumbor. | Epipactis palustris (L.) Crantz | + | + | - | + | - | - | -. | + | - | + | + | - | - | - | |
| Char. Magnocaricion elatae and Magnocaricetalia | |||||||||||||||||
| H | Europ.-Caucas. | Carex paniculata L. | 4 | 3 | 2 | 3 | 3 | 4 | 3 | 3 | 4 | 4 | 4 | 3 | 1 | 4 | 14 |
| H | Euri-Medit. | Galium palustre L. subsp. elongatum (C. Presl) Arcang. | 1 | 1 | 2 | 1 | 1 | 1 | 2 | 2 | 1 | 2 | 1 | 2 | 1 | 1 | 14 |
| G | Paleotemp. | Cyperus longus L. | 1 | 2 | 1 | 1 | + | - | - | 1 | 1 | 1 | + | 1 | + | + | 12 |
| G | Europ. | Carex flacca Schreb. subsp. flacca | - | - | + | + | - | - | + | + | - | + | + | - | - | - | 6 |
| H | Circumbor. | Carex pallescens L. | . | . | + | . | . | + | . | . | . | + | . | . | . | - | 3 |
| H | Eurasiat. | Rumex conglomeratus Murray | . | . | . | . | . | . | . | . | . | . | . | . | . | + | 1 |
| Char. Phragmito-Magnocaricetea | |||||||||||||||||
| H | Paleotemp | Mentha aquatica L. | 1 | + | 2 | 1 | 1 | 1 | 2 | 1 | 1 | + | 1 | 1 | 1 | 1 | 14 |
| H | Orof. NE-Medit. | Cirsium creticum (Lam.) d’Urv.subsp. triumfettii (Lacaita) K. Werner | + | 1 | + | 1 | 1 | 2 | 1 | + | 1 | + | + | 1 | + | + | 14 |
| G | Europ.-Caucas. | Juncus subnodulosus Schrank | 2 | 3 | 3 | 2 | 2 | 1 | 2 | 2 | 1 | 1 | 1 | 3 | 1 | . | 13 |
| H | Eurosiber. | Angelica sylvestris L. | + | + | + | + | 1 | 1 | + | + | 1 | + | 1 | 1 | + | . | 13 |
| G | Circumbor. | Equisetum palustre L. | 1 | 2 | + | 1 | + | 2 | 1 | 1 | 1 | 1 | + | + | 3 | . | 13 |
| H | Euri-Medit. | Carex distans L. | 2 | 1 | 1 | 1 | + | 2 | 1 | 1 | + | 1 | 1 | 1 | . | 2 | 13 |
| H | Paleotemp. | Hypericum tetrapterum Fr. | + | + | 1 | 1 | 1 | + | + | . | . | 1 | 1 | + | . | 1 | 11 |
| G | Subcosmop. | Phragmites australis (Cav.) Trin. ex Steud. | . | + | + | . | + | + | + | + | + | + | + | . | . | . | 9 |
| H | Paleotemp. | Epilobium parviflorum Schreb. | + | 1 | + | + | + | . | . | . | . | + | + | + | . | . | 8 |
| H | Paleotemp. | Lolium arundinaceum (Schreb.) Darbysh. | . | . | . | . | . | + | 1 | + | + | + | . | + | . | + | 7 |
| H | Paleotemp. | Eupatorium cannabinum L. | 1 | 1 | 1 | + | 1 | . | . | . | . | . | + | + | . | . | 7 |
| Ch | Medit. | Lotus rectus L. | . | . | + | . | . | . | . | . | + | + | 1 | + | . | . | 5 |
| G | Subcosmop. | Eleocharis palustris (L.) Roem. & Schult. | . | . | . | . | . | . | . | + | + | + | + | . | . | 1 | 5 |
| H | Euri-Medit. | Helosciadium nodiflorum (L.) W.D.J. Koch | . | . | . | . | . | + | + | . | . | . | . | . | 1 | 1 | 4 |
| G | Subcosmop | Glyceria spicata Guss. | . | . | . | . | . | + | + | . | . | . | . | . | + | 1 | 4 |
| Trasgr. Holoschoenetalia vulgaris | |||||||||||||||||
| H | Europ.-Caucas. Subatl. | Lysimachia nemorum L. | 2 | + | 1 | 1 | 1 | 1 | 1 | 1 | + | 1 | 1 | 2 | 2 | . | 13 |
| H | Circumbor. | Holcus lanatus L. | 1 | + | 1 | + | + | + | + | + | + | + | + | + | + | . | 13 |
| G | Medit. | Dactylorhiza maculata subsp. saccifera (Brongn.) Diklić | + | + | . | . | . | + | 1 | + | + | + | 1 | + | + | 1 | 11 |
| G | Cosmop. | Juncus effusus L. | . | . | . | + | + | 1 | 2 | + | + | . | + | 1 | 1 | 1 | 10 |
| G | Eurosiber. | Juncus conglomeratus L. | + | . | + | + | + | + | + | . | + | + | + | . | . | . | 9 |
| H | NE-Medit. | Rhynchocorys elephas (L.) Griseb. | . | . | . | . | . | + | + | 1 | 1 | + | 1 | + | 1 | . | 8 |
| H | Medit. | Lythrum junceum Banks & Sol. | . | . | . | . | . | + | + | + | + | + | + | + | + | 1 | 9 |
| Other species | |||||||||||||||||
| Calliergonella cuspidata (Hedw.) Loeske | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 14 | ||
| H | Paleotemp. | Lathyrus pratensis L. | . | . | . | . | . | . | + | 1 | 2 | + | + | + | . | + | 7 |
| H/T | Euri-Medit.-Sett. | Myosotis sicula Guss. | . | . | . | . | . | + | + | . | . | + | + | + | . | 1 | 6 |
| H | Circumbor. | Prunella vulgaris L. | . | . | . | . | . | . | + | + | + | . | + | . | + | . | 5 |
| G | Circumbor. | Juncus articulatus L. | . | . | . | + | + | . | . | . | . | . | . | . | 2 | 2 | 4 |
| H | Subcosmop. | Samolus valerandi L. | . | . | . | . | . | + | + | . | . | . | . | . | 1 | + | 4 |
| H | Subcosmop. | Isolepis cernua (Vahl) Roem. & Schult. | . | . | . | . | . | + | + | . | . | . | . | . | 1 | + | 4 |
| H | Europ.-Caucas. | Carex remota L. | . | . | . | . | . | . | . | . | . | . | . | . | 3 | . | 1 |
| H | Paleotemp. | Trifolium repens L. | . | + | + | . | . | . | . | . | . | . | . | . | . | . | 2 |
| T | Euri-Medit. | Ranunculus ophioglossifolius Vill. | . | . | . | . | . | . | . | . | . | . | . | . | + | + | 2 |
| G | NE-Medit | Geranium versicolor L. | . | . | . | . | . | . | . | . | . | . | . | . | + | . | 1 |
| H | Eurasiat. | Ajuga reptans L. | . | . | . | . | . | . | . | . | . | . | . | . | . | + | 1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sciandrello, S.; Cambria, S.; del Galdo, G.G.; Tavilla, G.; Minissale, P. Unexpected Discovery of Thelypteris palustris (Thelypteridaceae) in Sicily (Italy): Morphological, Ecological Analysis and Habitat Characterization. Plants 2021, 10, 2448. https://doi.org/10.3390/plants10112448
Sciandrello S, Cambria S, del Galdo GG, Tavilla G, Minissale P. Unexpected Discovery of Thelypteris palustris (Thelypteridaceae) in Sicily (Italy): Morphological, Ecological Analysis and Habitat Characterization. Plants. 2021; 10(11):2448. https://doi.org/10.3390/plants10112448
Chicago/Turabian StyleSciandrello, Saverio, Salvatore Cambria, Gianpietro Giusso del Galdo, Gianmarco Tavilla, and Pietro Minissale. 2021. "Unexpected Discovery of Thelypteris palustris (Thelypteridaceae) in Sicily (Italy): Morphological, Ecological Analysis and Habitat Characterization" Plants 10, no. 11: 2448. https://doi.org/10.3390/plants10112448