
Identification of Oil Palm’s Consistently Upregulated Genes during Early Infections of Ganoderma boninense via RNA-Seq Technology and Real-Time Quantitative PCR

 ,
,  ,
,
Abstract
:1. Introduction
2. Results
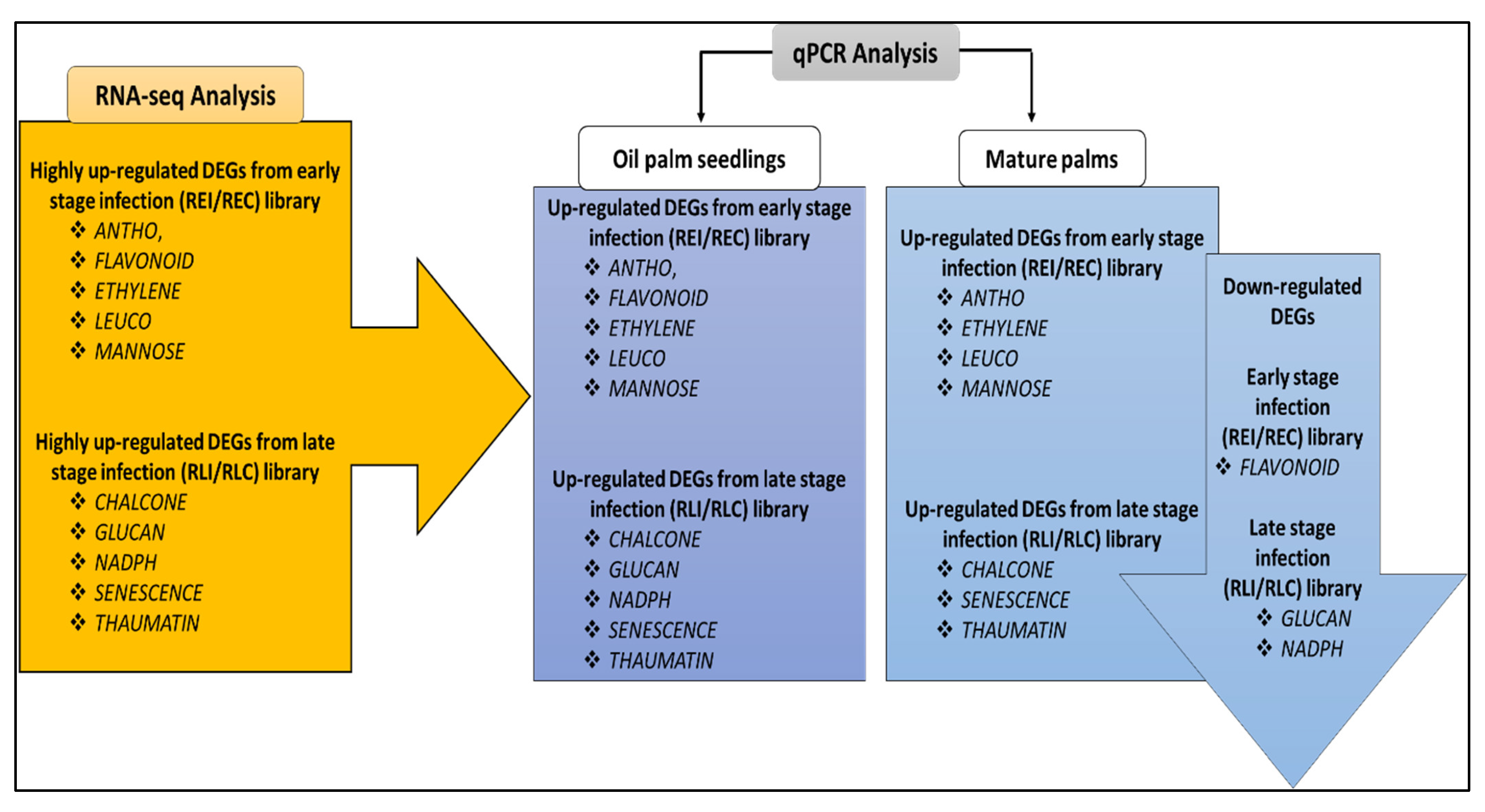
2.1. Transcriptomic Analysis via RNA-Seq
2.2. Identification of DEGs
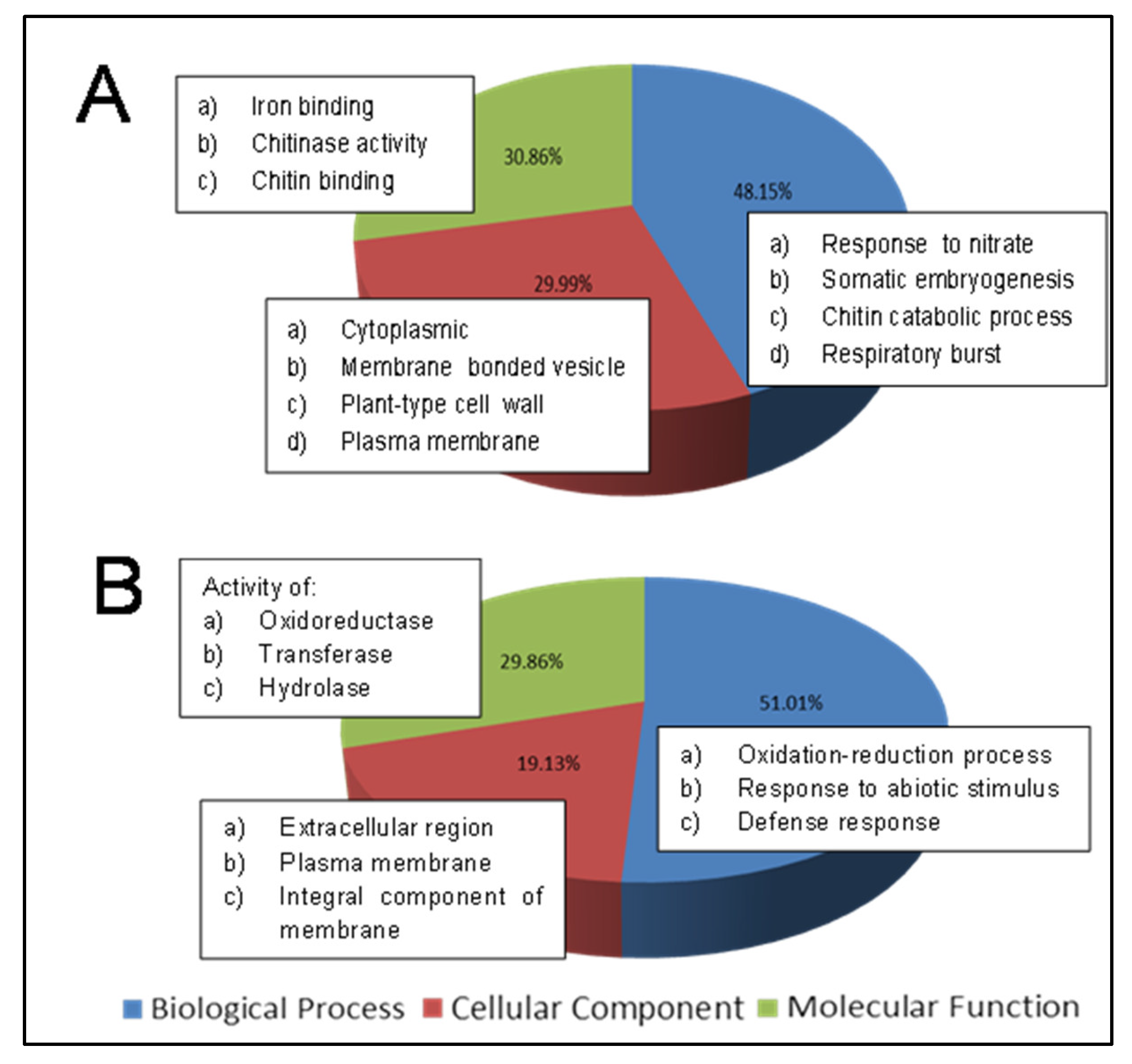
2.3. Functional Annotation
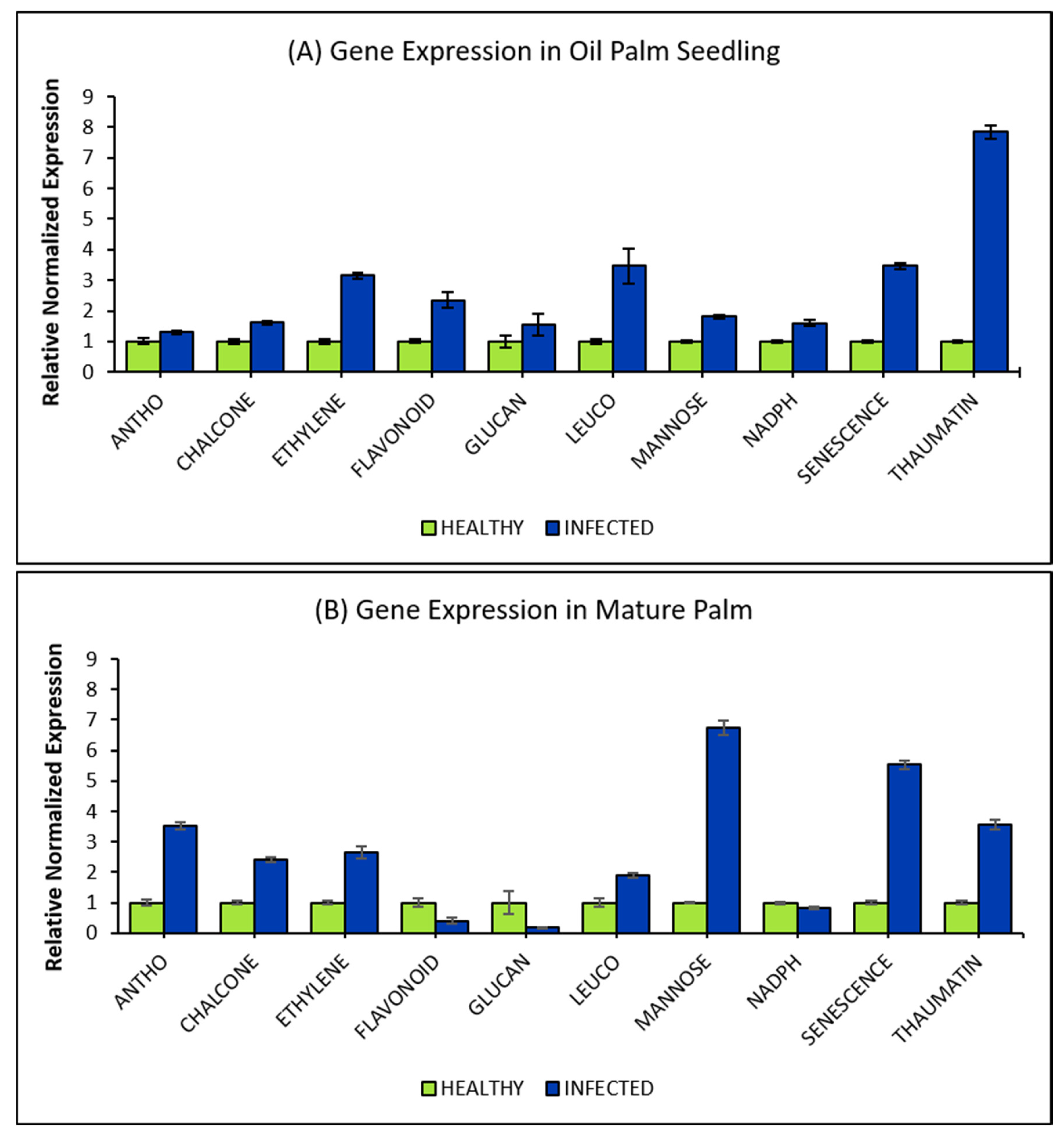
2.4. Gene Expression Profiling of Selected DEGs via qRT-PCR
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Treatments for Transcriptomic Profiling
4.2. Total RNA Extraction and cDNA Synthesis
4.3. RNA-Sequencing and Transcriptome Library Generation
4.4. Quantitative Real-Time Polymerase Chain Reaction (qRT-PCR) Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Malaysia Palm Oil Board (MPOB). Overview of the Malaysian Oil Palm Industry. 2019. Available online: http://bepi.mpob.gov.my/images/overview/Overview_of_Industry_2019.pdf (accessed on 13 June 2020).
- Nadzri, M.M.M.; Ahmad, A. Roller picker robot (ROPICOT 1.0) for loose fruit collection system. ARPN J. Eng. Appl. Sci. 2016, 11, 8983–8986. [Google Scholar]
- Abas, R.; Kamarudin, M.F.; Borhan, A.A.N.; Simeh, M.A. A study on the Malaysian oil palm biomass sector—Supply and perception of palm oil millers. Oil Palm Ind. Econ. J. 2011, 11, 28–41. [Google Scholar]
- Murphy, D.J. The future of oil palm as a major global crop: Opportunities and challenges. J. Oil Palm Res. 2014, 26, 1–24. [Google Scholar]
- Parthiban, K.; Tan, S.P.; Siti, M.A.; Idris, A.S.; Ayatollah, K.A.R.; Khairuman, H.; Hamdan, A.B.; Wahid, O. Knowledge assessment of basal stem rot disease of oil palm and its control practices among recipients of replanting assistance scheme in Malaysia. Int. J. Agric. Res. 2017, 12, 73–81. [Google Scholar] [CrossRef]
- Chung, G.F. Management of Ganoderma diseases in Oil palm plantations. Planter 2011, 87, 325–339. [Google Scholar]
- Naher, L.; Yusuf, U.K.; Ismail, A.; Tan, S.G.; Monal, M.M.A. Ecological status of Ganoderma and basal stem rot disease of oil palms (Elaeis guineensis Jacq.). Aust. J. Crop. Sci. 2013, 7, 1723–1727. [Google Scholar]
- Badalyan, M.S.; Innocenti, G.; Garibyan, G.N. Interactions between Xylotrophic mushrooms and mycoparasitic fungi in dual-culture experiments. Phytopathol. Mediterr. 2004, 43, 44–48. [Google Scholar]
- Chong, K.P.; Dayou, J.; Alexander, A. Pathogenic nature of Ganoderma boninense and basal stem rot disease. In Detection and Control of Ganoderma boninense in Oil Palm Crop; Chong, K.P., Dayou, J., Alexander, A., Eds.; Springer: Cham, Switzerland, 2017; pp. 5–12. [Google Scholar] [CrossRef]
- Durand-Gasselin, T.; Asmady, H.; Flori, A.; Jacquemard, J.C.; Hayun, Z.; Breton, F.; de Franqueville, H. Possible sources of genetic resistance in oil palm (Elaeis guineensis Jacq.) to basal stem rot caused by Ganoderma boninense-prospects for future breeding. Mycopathologia 2005, 159, 93–100. [Google Scholar] [CrossRef]
- Idris, A.S.; Kushairi, A.; Ismail, S.; Ariffin, D. Selection of partial resistance in oil palm progenies to Ganoderma basal stem rot. J. Oil Palm Res. 2004, 16, 12–18. [Google Scholar]
- Breton, F.; Hasan, Y.; Hariadi, S.; Lubis, Z.; De Franqueville, H. Characterization of parameters for the development of an early screening test for basal stem rot tolerance in oil palm progenies. J. Oil Palm Res. 2006, 18, 24–36. [Google Scholar]
- Bahari, M.N.A.; Sakeh, N.M.; Abdullah, S.N.A.; Ramli, R.R.; Kadkhodaei, S. Transciptome profiling at early infection of Elaeis guineensis by Ganoderma boninense provides novel insights on fungal transition from biotrophic to necrotrophic phase. BMC Plant Biol. 2018, 18, 377. [Google Scholar] [CrossRef]
- Parveez, G.K.A.; Hishamuddin, E.; Loh, S.K.; Ong-Abdullah, M.; Kamalrudin, M.S.; Zanal Bidin, M.N.I.; Sundram, S.; Hasan, Z.A.B.A.; Idris, Z. Oil Palm Economic Performance in Malaysia and R&D Progress in 2019. J. Oil Palm Res. 2020, 32, 159–190. [Google Scholar]
- Tee, S.S.; Tan, Y.C.; Abdullah, F.; Ong-Abdullah, M.; Ho, C.L. Transcriptome of oil palm (Elaeis guineensis Jacq.) roots treated with Ganoderma boninense. Tree Genet. Genomes 2013, 9, 377–386. [Google Scholar] [CrossRef]
- Yeoh, K.A.; Othman, A.; Meon, S.; Abdullah, F.; Ho, C.L. Sequence analysis and gene expression of putative exo- and endo-glucanases from oil palm (Elaeis guineensis) during fungal infection. J. Plant Physiol. 2012, 169, 1565–1570. [Google Scholar] [CrossRef] [PubMed]
- Yeoh, K.A.; Othman, A.; Meon, S.; Abdullah, F.; Ho, C.L. Sequence analysis and gene expression of putative oil palm chitinase and chitinase-like proteins in response to colonization of Ganoderma boninense and Trichoderma harzianum. Mol. Biol. Rep. 2013, 40, 147–158. [Google Scholar] [CrossRef] [PubMed]
- Naher, L.; Ho, C.L.; Tan, S.G.; Yusuf, U.K.; Ahmad, S.H.; Abdullah, F. Cloning of transcripts encoding chitinases from Elaeis guineensis Jacq. and their expression profiles in response to fungal infections. Physiol. Mol. Plant Pathol. 2011, 76, 96–103. [Google Scholar] [CrossRef]
- Tan, Y.C.; Yeoh, K.A.; Wong, M.Y.; Ho, C.L. Expression profiles of putative defence-related proteins in oil palm (Elaeis guineensis) colonised by Ganoderma boninense. J. Plant Physiol. 2013, 170, 1455–1460. [Google Scholar] [CrossRef]
- Ho, C.L.; Tan, Y.C.; Yeoh, K.A.; Ghazali, A.K.; Yee, W.Y.; Hoh, C.C. De novo transcriptome analyses of host fungal interactions in oil palm (Elaeis guineensis Jacq.). BMC Genom. 2016, 17, 66–84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goyer, A.; Hamlin, L.; Crosslin, J.; Buchanan, A.; Chang, J.H. RNA-Seq analysis of resistant and susceptible potato varieties during the early stages of potato virus Y infection. BMC Genom. 2015, 16, 472–484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, G.; Liu, K.; Kang, J.; Xu, K.; Zhang, Y.; Hu, L.; Zhang, J.; Li, C. Transcriptome analysis of the compatible interaction of tomato with Verticillium dahliae using RNA-sequencing. Front. Plant Sci. 2015, 6, 428–437. [Google Scholar] [CrossRef] [Green Version]
- Chen, T.; Lv, Y.; Zhao, T.; Li, N.; Yang, Y.; Yu, W.; He, X.; Liu, T.; Zhang, B. Comparative transcriptome profiling of a resistant vs. susceptible tomato (Solanum lycopersicum) cultivar in response to infection by tomato Yellow Leaf Curl Virus. PLoS ONE 2013, 8, e80816. [Google Scholar] [CrossRef] [PubMed]
- Kamber, T.; Buchmann, J.P.; Pothier, J.F.; Smits, T.H.M.; Wicker, T.; Duffy, B. Fire blight disease reactome: RNAseq transcriptional profile of apple host plant defense responses to Erwinia amylovora pathogen infection. Sci. Rep. 2016, 6, 21600. [Google Scholar] [CrossRef] [Green Version]
- Chakraborty, S.; Britton, M.; Martinez-Garcia, M.P.J.; Dandekar, A.M. Deep RNA-Seq profile reveals biodiversity, plant–microbe interactions and a large family of NBS-LRR resistance genes in walnut (Juglans regia) tissues. AMB Express 2016, 6, 12–24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ho, C.L.; Tan, Y.C.; Yeoh, K.A.; Lee, W.K.; Ghazali, A.K.; Yee, W.Y.; Hoh, C.C. Transcriptional response of oil palm (Elaeis guineensis Jacq.) inoculated simultaneously with both Ganoderma boninense and Trichoderma harzianum. Plant Gene 2018, 13, 56–63. [Google Scholar] [CrossRef]
- Kendziorski, C.; Irizarry, R.A.; Chen, K.S.; Haag, J.D.; Gould, M.N. On the utility of pooling biological samples in microarray experiments. Proc. Natl. Acad. Sci. USA 2005, 102, 4252–4257. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Biswas, S.; Agrawal, Y.N.; Mucyn, T.S.; Dangl, J.L.; Jones, C.D. Biological averaging in RNA-seq. arXiv 2013, arXiv:1309.0670. [Google Scholar]
- Trapnell, C.; Williams, B.A.; Pertea, G.; Mortazavi, A.; Kwan, G.; van Baren, M.J.; Salzberg, S.L.; Wold, B.J.; Pachter, L. Transcript assembly and quantification by RNA-seq reveals unannotated transcripts and isoform switching during cell differentiation. Nat. Biotechnol. 2010, 28, 511–515. [Google Scholar] [CrossRef] [Green Version]
- Bustin, S.A.; Benes, V.; Garson, J.A.; Hellemans, J.; Huggett, J.; Kubista, M.; Mueller, R.; Nolan, T.; Pfaffl, M.W.; Shipley, G.L.; et al. The MIQE guidelines: Minimum information for publication of quantitative real-time PCR experiments. Clin. Chem. 2009, 55, 611–622. [Google Scholar] [CrossRef] [Green Version]
- Vandesompele, J.; De Preter, K.; Pattyn, F.; Poppe, B.; Van Roy, N.; De Paepe, A.; Speleman, F. Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes. Genome Biol. 2002, 3, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Song, W.; Jiang, K.; Zhang, F.; Lin, Y.; Ma, L. RNA-sequencing of the sturgeon Acipenser baeri provides insights into expression dynamics of morphogenic differentiation and developmental regulatory genes in early versus late developmental stages. BMC Genom. 2016, 17, 564–576. [Google Scholar] [CrossRef] [Green Version]
- Schultz, J.C.; Appel, H.M.; Ferrieri, A.P.; Arnold, T.M. Flexible resource allocation during plant defense responses. Front. Plant Sci. 2013, 4, 324. [Google Scholar] [CrossRef] [Green Version]
- Gorinova, N.; Nedkovska, M.; Bakalova, E.; Atanassov, A.; Ohkawa, H. Biotransformation of xenobiotics and endogenous substrates by plant cytochrome P450s. Biotechnol. Biotec. Eq. 1999, 13, 13–19. [Google Scholar] [CrossRef]
- Cheng, Z.; Yu, X.; Li, S.; Wu, Q. Genome-wide transcriptome analysis and identification of benzothiadiazole-induced genes and pathways potentially associated with defense response in banana. BMC Genom. 2018, 19, 454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Banday, Z.Z.; Nandi, A.K. Interconnection between flowering time control and activation of systemic acquired resistance. Front. Plant Sci. 2015, 6, 174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Conrath, U. Systemic acquired resistance. Plant Signal Behav. 2006, 1, 79–84. [Google Scholar] [CrossRef]
- Hwang, I.S.; Hwang, B.K. The pepper mannose-binding lectin gene CaMBL1 is required to regulate cell death and defense responses to microbial pathogens. Plant Physiol. 2011, 155, 447–463. [Google Scholar] [CrossRef] [Green Version]
- Butt, A.; Mousley, C.; Morris, K.; Beynon, J.; Can, C.; Holub, E.; Greenberg, J.T.; Buchanan-Wollaston, V. Differential expression of a senescence-enhanced metallothionein gene in Arabidopsis in response to isolates of Peronospora parasitica and Pseudomonas syringae. Plant J. 1998, 16, 209–221. [Google Scholar] [CrossRef] [PubMed]
- Pontier, D.; Gan, S.; Amasino, R.M.; Roby, D.; Lam, E. Markers for hypersensitive response and senescence show distinct patterns of expression. Plant Mol. Biol. 1999, 39, 1243–1255. [Google Scholar] [CrossRef]
- Liu, J.J.; Sturrock, R.; Ekramoddoullah, A.K. The superfamily of thaumatin-like proteins: Its origin, evolution, and expression towards biological function. Plant Cell Rep. 2010, 29, 419–436. [Google Scholar] [CrossRef] [PubMed]
- Shatters, R.G., Jr.; Boykin, L.M.; Lapointe, S.L.; Hunter, W.B.; Weathersbee, A.A., III. Phylogenetic and structural relationships of the PR5 gene family reveal an ancient multigene family conserved in plants and select animal taxa. J. Mol. Evol. 2006, 63, 12–29. [Google Scholar] [CrossRef]
- Dafoe, N.J.; Gowen, B.E.; Constabel, C.P. Thaumatin-like proteins are differentially expressed and localized in phloem tissues of hybrid poplar. BMC Plant Biol. 2010, 10, 191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, M.; Zhang, D.; Xing, Y.; Guo, S. Cloning and expression of three thaumatin-like protein genes from Polyporus umbellatus. Acta Pharm. Sin. B 2017, 7, 373–380. [Google Scholar] [CrossRef]
- Zuk, M.; Dzialo, M.; Richter, D.; Dyminska, L.; Matula, J.; Kotecki, A.; Hanuza, J.; Szopa, J. Chalcone synthase (CHS) gene suppression in flax leads to changes in wall synthesis and sensing genes, cell wall chemistry and stem morphology parameters. Front. Plant Sci. 2016, 7, 894. [Google Scholar] [CrossRef] [Green Version]
- Chuck, G.; Muszynski, M.; Kellogg, E.; Hake, S.; Schmidt, R.J. The control of spikelet meristem identity by the branched silkless1 gene in maize. Science 2002, 298, 1238–1241. [Google Scholar] [CrossRef]
- Ecker, J.R.; Davis, R.W. Plant defense genes are regulated by ethylene. Proc. Natl. Acad. Sci. USA 1987, 84, 5202–5206. [Google Scholar] [CrossRef] [Green Version]
- Sun, W.; Meng, X.; Liang, L.; Jiang, W.; Huang, Y.; He, J.; Hu, H.; Almqvist, J.; Gao, X.; Wang, L. Molecular and biochemical analysis of chalcone synthase from Freesia hybrid in flavonoid biosynthetic pathway. PLoS ONE 2015, 10, e0119054. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shimada, S.; Inoue, Y.T.; Sakuta, M. Anthocyanidin synthase in non-anthocyanin-producing Caryophyllales species. Plant J. 2005, 44, 950–959. [Google Scholar] [CrossRef] [PubMed]
- Bogs, J.; Downey, M.O.; Harvey, J.S.; Ashton, A.R.; Tanner, G.J.; Robinson, S.P. Proanthocyanidin synthesis and expression of genes encoding leucoanthocyanidin reductase and anthocyanidin reductase in developing grape berries and grapevine leaves. Plant Physiol. 2005, 139, 652–663. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liao, L.; Vimolmangkang, S.; Wei, G.; Zhou, H.; Korban, S.S.; Han, Y. Molecular characterization of genes encoding leucoanthocyanidin reductase involved in proanthocyanidin biosynthesis in apple. Front. Plant Sci. 2015, 6, 243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paolocci, F.; Robbins, M.P.; Madeo, L.; Arcioni, S.; Martens, S.; Damiani, F. Ectopic expression of a basic helix-loop-helix gene transactivates parallel pathways of proanthocyanidin biosyn thesis: Structure, expression analysis, and genetic control of leucoanthocyanidin 4-reductase and anthocyanidin reductase genes in Lotus corniculatus. Plant Physiol. 2007, 143, 504–516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.S.; Xu, Y.J.; Gao, L.P.; Yu, O.; Wang, X.Z.; He, X.J.; Jiang, X.L.; Liu, Y.J.; Xia, T. Functional analysis of flavonoid 3′,5′-hydroxylase from tea plant (Camellia sinensis): Critical role in the accumulation of catechins. BMC Plant Biol. 2014, 14, 347. [Google Scholar] [CrossRef] [Green Version]
- Winkel-Shirley, B. Biosynthesis of flavonoids and effects of stress. Curr. Opin. Plant Biol. 2002, 5, 218–223. [Google Scholar] [CrossRef]
- Sekher-Pannala, A.; Chan, T.S.; O’Brien, P.J.; Rice-Evans, C.A. Flavonoid B-ring chemistry and antioxidant activity: Fast reaction kinetics. Biochem. Biophys. Res. Commun. 2001, 282, 1161–1168. [Google Scholar] [CrossRef]
- Sharma, M.; Chai, C.; Morohashi, K.; Grotewold, E.; Snook, M.E.; Chopra, S. Expression of flavonoid 3′-hydroxylase is controlled by P1, the regulator of 3-deoxyflavonoid biosynthesis in maize. BMC Plant Biol. 2012, 12, 196. [Google Scholar] [CrossRef] [Green Version]
- Liu, S.; Ju, J.; Xia, G. Identification of the flavonoid 3′-hydroxylase and flavonoid 3′,5′-hydroxylase genes from Antarctic moss and their regulation during abiotic stress. Gene 2014, 543, 145–152. [Google Scholar] [CrossRef]
- Forkmann, G. Flavonoids as flower pigments: The formation of the natural spectrum and its extension by genetic engineering. Plant Breed. 1991, 106, 1–26. [Google Scholar] [CrossRef]
- Roubalová, L.; Dinkova-Kostovac, A.T.; Biedermann, D.; Křend, V.; Ulrichová, J.; Vrba, J. Flavonolignan 2,3-dehydrosilydianin activates Nrf2 and upregulates NAD(P) H:quinone oxidoreductase 1 in Hepa1c1c7 cells. Fitoterapia 2017, 119, 115–120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shetty, N.P.; Jensen, J.D.; Knudsen, A.; Finnie, C.; Geshi, N.; Blennow, A.; Collinge, D.B.; Jørgensen, H.J.L. Effects of β-1,3-glucan from Septoria tritici on structural defence responses in wheat. J. Exp. Bot. 2009, 60, 4287–4300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jongsma, M.A.; Bolter, C. The adaptation of insect to plant protease inhibitors. J. Insect. Physiol. 1997, 43, 885–895. [Google Scholar] [CrossRef]
- Paterson, R.R.M. Ganoderma disease of oil palm—A white rot perspective necessary for integrated control. J. Crop. Prot. 2007, 26, 1369–1376. [Google Scholar] [CrossRef] [Green Version]
- Sundram, S.; Meon, S.; Seman, I.A.; Othman, R. Symbiotic interaction of endophytic bacteria with arbuscular mycorrhizal fungi and its antagonistic effect on Ganoderma boninense. J. Microbiol. 2011, 49, 551. [Google Scholar] [CrossRef]
- Idris, A.S.; Kushairi, D.; Ariffin, D.; Basri, M.W. Technique for inoculation of oil palm germinated seeds with Ganoderma. MPOB Inf. Ser. 2006, 314, 1–4. [Google Scholar]
- Abdullah, F.; Ilias, G.N.M.; Nelson, M.; Izzati, N.A.M.Z.; Yusuf, K.U. Disease assessment and the efficacy of Trichoderma as a biocontrol agent of basal stem rot of oil palms. Sci. Putra 2003, 12, 31–33. [Google Scholar]
- Sundram, S.; Abdullah, F.; Ahmad, Z.A.M.; Yusuf, U.K. Efficacy of single and mixed treatments of Trichoderma harzianum as biocontrol agents of Ganoderma basal stem rot in oil palm. J. Oil Palm Res. 2008, 20, 470–483. [Google Scholar]
- Zeng, Y.; Yang, T. RNA isolation from highly viscous samples rich in polyphenols and polysaccharides. Plant Mol. Biol. Rep. 2002, 20, 417. [Google Scholar] [CrossRef]
- Zuhar, L.M.; Madihah, A.Z.; Ahmad, S.A.; Zainal, Z.; Idris, A.S.; Shaharuddin, N.A. Determination of reference genes for normalization of gene expression study of Ganoderma-infected oil palms. J. Oil Palm Res. 2019, 31, 550–556. [Google Scholar] [CrossRef]
- Langmead, B.; Trapnell, C.; Pop, M.; Salzberg, S. Ultrafast and memory-efficient alignment of short DNA sequences to the human genome. Genome Biol. 2009, 10, R25. [Google Scholar] [CrossRef] [Green Version]
- Trapnell, C.; Roberts, A.; Goff, L.; Pertea, G.; Kim, D.; Kelley, D.R.; Pimentel, H.; Salzberg, S.L.; Rinn, J.L.; Pachter, L. Differential gene and transcript expression analysis of RNA-seq experiments with TopHat and Cufflinks. Nat. Protoc. 2012, 7, 562–578. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ashburner, M.; Ball, C.A.; Blake, J.A.; Botstein, D.; Butler, H.; Cherry, J.M.; Davis, A.P.; Dolinski, K.; Dwight, S.S.; Eppig, J.T.; et al. Gene ontology: Tool for the unification of biology. The Gene Ontology Consortium. Nat. Genet. 2000, 25, 25–29. [Google Scholar] [CrossRef] [Green Version]
- Blast2GO. Available online: www.blast2go.com (accessed on 23 July 2016).
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT–PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef]
- Enke, R. qPCR Primer Efficiency Standard Curve Analysis. CSHL DNALC RNA-Seq for the Next Generation Working Group. 2016. Available online: http://www.rnaseqforthenextgeneration.org/profiles/raymond-enke.html#teaching (accessed on 30 June 2020).
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using RealTime Quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Samples | Total Reads (bp) | Low-Quality Reads (%) | Total High-Quality Reads (bp) | PhiX Contamination (%) | Total Sample Reads (bp) |
|---|---|---|---|---|---|
| REI | 16,536,956 | 8.51 | 15,129,373 | 0.32 | 15,080,896 |
| RLI | 21,075,290 | 9.49 | 19,075,010 | 0.31 | 19,014,726 |
| REC | 17,741,550 | 8.71 | 16,195,459 | 0.33 | 16,141,333 |
| RLC | 27,247,240 | 10.79 | 24,305,602 | 0.32 | 24,227,629 |
| Oil Palm Root Sample Pairs | Total Significant DEGs | Up-Regulated Transcripts | Down-Regulated Transcripts |
|---|---|---|---|
| REI/REC | 21 | 12 | 9 |
| RLI/RLC | 103 | 69 | 34 |
| Gene ID | Annotation Genes | Labels | FPKM | Log 2 Fold-Change | |
|---|---|---|---|---|---|
| Control | Treated | ||||
| Highly Upregulated Root Genes at Early Stage of Infection (REI/REC) | |||||
| XLOC_022484 | anthocyanidin synthase | ANTHO | 5.95 | 189.31 | 31.79 |
| XLOC_009804 | leucoanthocyanidin reductase-like | LEUCO | 16.66 | 198.57 | 11.91 |
| XLOC_019742 | ethylene-responsive transcription factor 1b-like | ETHYLENE | 4.09 | 109.77 | 26.85 |
| XLOC_005112 | flavonoid 3–hydroxylase | FLAVONOID | 0 | 19.56 | Positive Infinity |
| XLOC_013737 | mannose-specific lectin-like | MANNOSE | 85.08 | 1114.43 | 13.09 |
| Highly Upregulated Root Genes at Late Stage of Infection (RLI/RLC) | |||||
| XLOC_016957 | chalcone synthase | CHALCONE | 0 | 674.91 | Positive Infinity |
| XLOC_009482 | glucan endo-beta-glucosidase-like | GLUCAN | 555.34 | 9543.37 | 17.18 |
| XLOC_009990 | nadph:quinone oxidoreductase-like | NADPH | 5.69 | 107.89 | 18.96 |
| XLOC_017303 | senescence-associated partial | SENESCENCE | 91.86 | 1540.91 | 16.77 |
| XLOC_001542 | thaumatin-like protein | THAUMATIN | 13.10 | 187.77 | 14.33 |
| Gene | Amplicon Size (bp) | BLAST Results | Score (Bits) | E-Value | Percentage of Identity (%) | Accession No. |
|---|---|---|---|---|---|---|
| ANTHO | 138 | Anthocyanidin synthase (Delphinium grandiflorum) | 51.6 | 9 × 10−7 | 92 | BAO04186.1 |
| LEUCO | 170 | PREDICTED: leucoanthocyanidin reductase-like(Elaeis guineensis) | 72.4 | 4 × 10−14 | 100 | XP 010916146.1 |
| ETHYLENE | 142 | PREDICTED: ethylene-responsive transcription factor 1B-like (Elaeis guineensis) | 70.5 | 2 × 10−13 | 100 | XP 010904582.1 |
| FLAVONOID | 155 | flavonoid 3′–hydroxylase (Sorghum bicolor) | 56.2 | 3 × 10−8 | 83 | ABG54321.1 |
| MANNOSE | 105 | PREDICTED: mannose-specific lectin-like(Elaeis guineensis) | 43.9 | 3 × 10−4 | 100 | XP 010910930.1 |
| CHALCONE | 135 | PREDICTED: chalcone synthase-like (Elaeis guineensis) | 48.1 | 9 × 10−5 | 85 | XP 010910945.1 |
| GLUCAN | 110 | PREDICTED: glucan endo- 1,3-beta-glucosidase-like (Elaeis guineensis) | 36.2 | 0.67 | 79 | XP 010921890.1 |
| NADPH | 116 | PREDICTED: NADPH:quinone oxidoreductase-like (Elaeis guineensis) | 40.8 | 0.010 | 100 | XP 010922202.1 |
| SENESCENCE | 169 | putative senescence-associated protein (Cupressus sempervirens) | 90.5 | 1 × 10−21 | 100 | ACA30301.1 |
| THAUMATIN | 110 | PREDICTED: Elaeis guineensis thaumatin-like protein (LOC105033331), mRNA | 75.0 | 2 × 10−10 | 94 | XM 010908090.1 |
| Primer Name | Primer Sequence (5′–3′) | Product Length (bp) | |
|---|---|---|---|
| Forward Primer | Reverse Primer | ||
| ANTHO | ACAACATGGTCCCCGGTCT | GGTGGAGGATGCTCTTGTAGGT | 138 |
| LEUCO | TCCGTTTTGGGCGGTTCT | CGGCGGACTTTCCTCTTTTC | 170 |
| ETHYLENE | AAGAGCAAGGCAGGGAATGG | CTTCTGCGCTGTCAAAGGTTC | 142 |
| FLAVONOID | GTTTGTGGTGGGAGACTTCGTG | CCTCATTCTGCTCGGTTGGAC | 155 |
| MANNOSE | TCGGATGGGAACCTTGTGG | CCGATCTCGTTGGAGGATACAG | 105 |
| CHALCONE | GAGCAGATCCAATGCAAGGTGT | GGTTGAGGAGGTGGAAGGTGA | 135 |
| GLUCAN | AGCAAGCTACTGGGTCCAAAC | GCACATACTGGGCTTTATCTCC | 110 |
| NADPH | CGAGATTGATGGCAAGTGTCC | TCAGAGGAGCTGGGATGGAGT | 116 |
| SENESCENCE | GGCACGGCCATCAGTAGAGTA | AGCCAAGCGTTCATAGCGAC | 169 |
| THAUMATIN | ACGAGGGAGATGTCGATGAA | GACTGCGGTGGTAAACTTGC | 110 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zuhar, L.M.; Madihah, A.Z.; Ahmad, S.A.; Zainal, Z.; Idris, A.S.; Shaharuddin, N.A. Identification of Oil Palm’s Consistently Upregulated Genes during Early Infections of Ganoderma boninense via RNA-Seq Technology and Real-Time Quantitative PCR. Plants 2021, 10, 2026. https://doi.org/10.3390/plants10102026
Zuhar LM, Madihah AZ, Ahmad SA, Zainal Z, Idris AS, Shaharuddin NA. Identification of Oil Palm’s Consistently Upregulated Genes during Early Infections of Ganoderma boninense via RNA-Seq Technology and Real-Time Quantitative PCR. Plants. 2021; 10(10):2026. https://doi.org/10.3390/plants10102026
Chicago/Turabian StyleZuhar, Liyana Mohd, Ahmad Zairun Madihah, Siti Aqlima Ahmad, Zamri Zainal, Abu Seman Idris, and Noor Azmi Shaharuddin. 2021. "Identification of Oil Palm’s Consistently Upregulated Genes during Early Infections of Ganoderma boninense via RNA-Seq Technology and Real-Time Quantitative PCR" Plants 10, no. 10: 2026. https://doi.org/10.3390/plants10102026