
The Integrated Amendment of Sodic-Saline Soils Using Biochar and Plant Growth-Promoting Rhizobacteria Enhances Maize (Zea mays L.) Resilience to Water Salinity

 ,
, 
 , ,
, ,
Abstract
:1. Introduction
2. Results
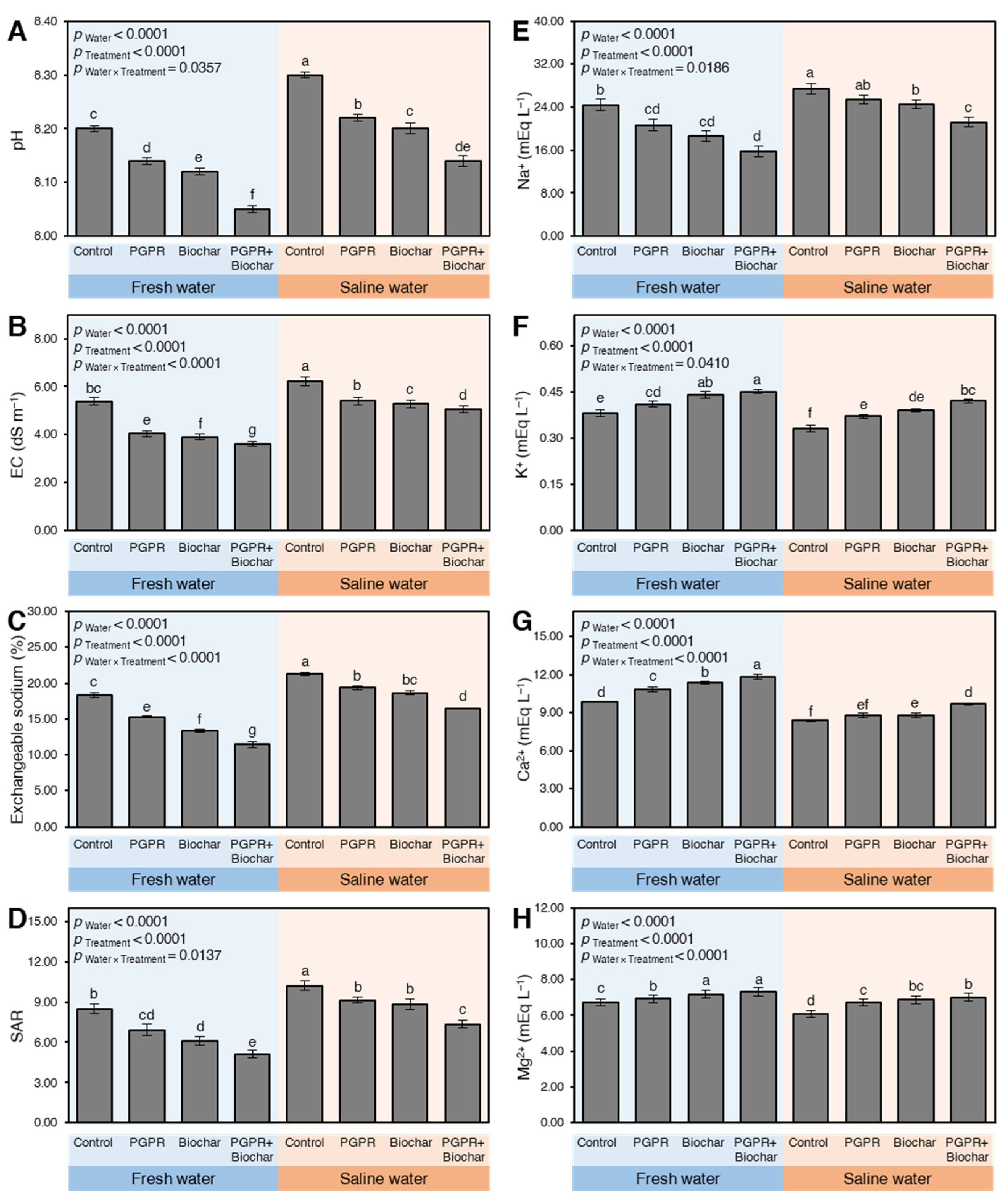
2.1. Biochar and PGBP Application Improved the Soil Ecosystem
2.1.1. Soil Physicochemical Properties
2.1.2. Activity of Soil Enzymes
2.1.3. Bacteriological Characteristics
2.2. Soil Amendment Using Biochar and PGPR Enhanced Maize Performance and Resilience to Water Salinity
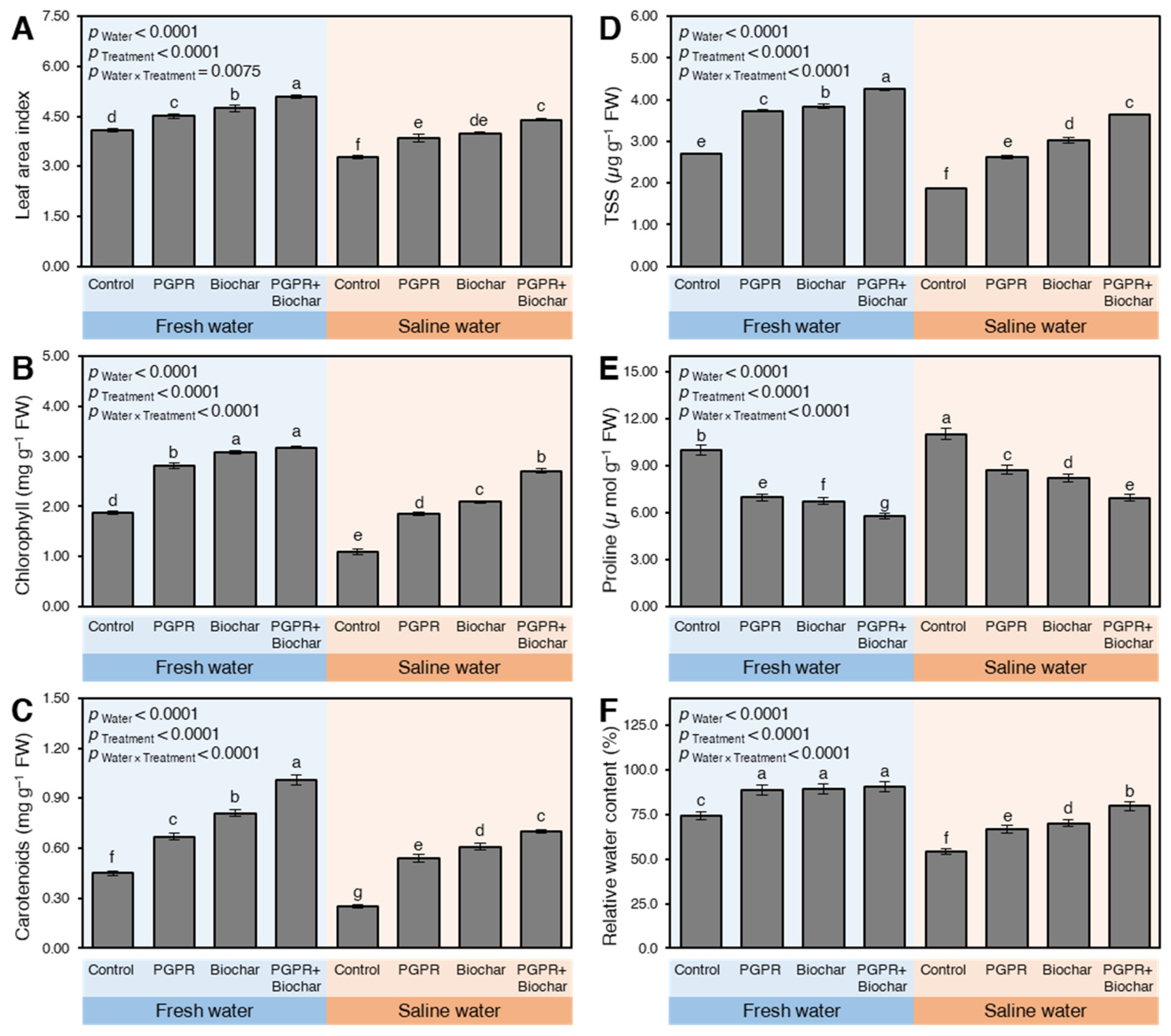
2.2.1. Leaf Area Index and Photosynthetic Pigments
2.2.2. Total Soluble Sugars (TSS), Proline, and Relative Water Content (RWC)
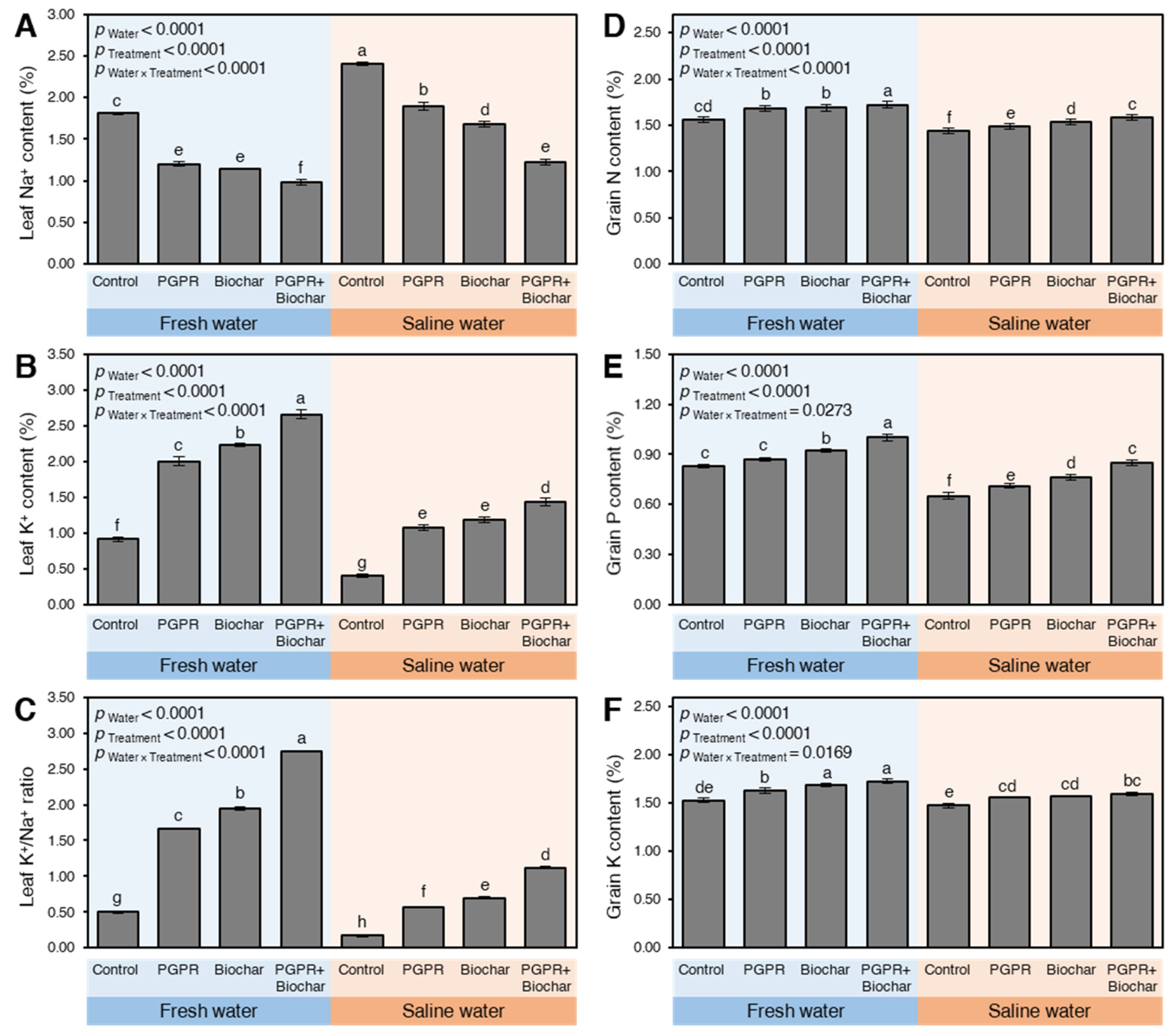
2.2.3. The Leaf Content of Na+, K+, and K+/Na+
2.2.4. The NPK Content of Maize Grains
2.2.5. Yield and Yield Components of Maize
3. Discussion
4. Materials and Methods
4.1. Source of PGPR and Growth Conditions
4.2. Biochar Characterization
4.3. Field Experiments and Growth Conditions
4.3.1. Location and Treatments
4.3.2. Soil Sampling
4.3.3. Agronomic Practices
4.4. Plant Biometrics
4.4.1. Preparation of Plant Samples
4.4.2. Photosynthetic Pigments
4.4.3. Proline Content
4.4.4. Total Soluble Sugar (TSS)
4.4.5. Relative Water Content (RWC)
4.4.6. Determination of Na+ and K+ in Maize Leaves
4.4.7. Leaf Area Index
4.5. Maize Productivity
4.5.1. Yield and Yield Components
4.5.2. NPK Content in Maize Grains
4.6. Soil Measurements
4.6.1. Soil Dehydrogenase and Urease Activity
4.6.2. Microbial Estimations
4.6.3. Soil Physicochemical Properties
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ullah, N.; Ditta, A.; Khalid, A.; Mehmood, S.; Rizwan, M.S.; Ashraf, M.; Mubeen, F.; Imtiaz, M.; Iqbal, M.M. Integrated effect of algal biochar and plant growth promoting rhizobacteria on physiology and growth of maize under deficit irrigations. J. Soil Sci. Plant Nutr. 2019, 20, 346–356. [Google Scholar] [CrossRef]
- FAO. FAOSTAT. Available online: http://www.fao.org/faostat/en/#data/QC (accessed on 20 February 2020).
- Hafez, E.M.; Omara, A.E.D.; Alhumaydhi, F.A.; El-Esawi, M.A. Minimizing hazard impacts of soil salinity and water stress on wheat plants by soil application of vermicompost and biochar. Physiol. Plant. 2020, 172, 587–602. [Google Scholar] [CrossRef]
- Hafez, E.E.-D.M.; El Hassan, W.H.A.; Gaafar, I.A.; Seleiman, M.F. Effect of gypsum application and irrigation intervals on clay saline-sodic soil characterization, rice water use efficiency, growth, and yield. J. Agric. Sci. 2015, 7, 208. [Google Scholar] [CrossRef] [Green Version]
- Ding, Z.; Kheir, A.M.; Ali, O.A.; Hafez, E.M.; El Shamey, E.A.; Zhou, Z.; Wang, B.; Lin, X.; Ge, Y.; Fahmy, A.E.; et al. A vermicompost and deep tillage system to improve saline-sodic soil quality and wheat productivity. J. Environ. Manag. 2020, 277, 111388. [Google Scholar] [CrossRef] [PubMed]
- El-Ramady, H.; Alshaal, T.; Elhawat, N.; Ghazi, A.; Elsakhawy, T.; Omara, A.E.-D.; El-Nahrawy, S.; Elmahrouk, M.; Abdalla, N.; Domokos-Szabolcsy, É.; et al. Plant nutrients and their roles under saline soil conditions. In Plant Nutrients and Abiotic Stress Tolerance; Hasanuzzaman, M., Fujita, M., Oku, H., Nahar, K., Hawrylak-Nowak, B., Eds.; Springer: Singapore, 2018; pp. 297–324. [Google Scholar] [CrossRef]
- Hafez, E.M.; Kheir, A.M.S.; Badawy, S.A.; Rashwan, E.; Farig, M.; Osman, H.S. Differences in physiological and biochemical attributes of wheat in response to single and combined salicylic acid and biochar subjected to limited water irrigation in saline sodic soil. Plants 2020, 9, 1346. [Google Scholar] [CrossRef]
- Alshaal, T.; El-Ramady, H.; Al-Saeedi, A.H.; Shalaby, T.; Elsakhawy, T.; Omara, A.E.-D.; Gad, A.; Hamad, E.; El-Ghamry, A.; Mosa, A.; et al. The rhizosphere and plant nutrition under climate change. In Essential Plant Nutrients; Naeem, M., Ansari, A., Gill, S., Eds.; Springer: Cham, Switzerland, 2017; pp. 275–308. [Google Scholar] [CrossRef]
- El-Ramady, H.; Alshaal, T.; Bakr, N.; Elbana, T.; Mohamed, E.; Belal, A.A. The soils of Egypt. In World Soils Book Series, 1st ed.; El-Ramady, H., Alshaal, T., Bakr, N., Elbana, T., Mohamed, E., Belal, A.-A., Eds.; Springer International Publishing: Cham, Switzerland, 2019. [Google Scholar]
- Sofy, M.R.; Elhawat, N.; Alshaal, T. Glycine betaine counters salinity stress by maintaining high K+/Na+ ratio and antioxidant defense via limiting Na+ uptake in common bean (Phaseolus vulgaris L.). Ecotoxicol. Environ. Saf. 2020, 200, 110732. [Google Scholar] [CrossRef] [PubMed]
- Shahid, S.A.; Zaman, M.; Heng, L. Soil salinity: Historical perspectives and a world overview of the problem. In Guideline for Salinity Assessment; Springer: Cham, Switzerland, 2018; pp. 43–53. [Google Scholar] [CrossRef] [Green Version]
- Szabolcs, I. Salt-Affected Soils; CRC Press: Boca Raton, FL, USA, 1989. [Google Scholar]
- Ghassemi, F.; Jakeman, A.J.; Nix, H.A. Salinisation of Land and Water Resources: Human Causes, Extent, Management and Case Studies; CABI Publishing: Wallingford, UK, 1995; p. 526. [Google Scholar]
- Metternicht, G.; Zinck, J. Remote sensing of soil salinity: Potentials and constraints. Remote. Sens. Environ. 2003, 85, 1–20. [Google Scholar] [CrossRef]
- Guo, M.; Song, W.; Tian, J. Biochar-facilitated soil remediation: Mechanisms and efficacy variations. Front. Environ. Sci. 2020, 8, 183. [Google Scholar] [CrossRef]
- Zhang, X.; Qu, J.; Li, H.; La, S.; Tian, Y.; Gao, L. Biochar addition combined with daily fertigation improves overall soil quality and enhances water-fertilizer productivity of cucumber in alkaline soils of a semi-arid region. Geoderma 2020, 363, 114170. [Google Scholar] [CrossRef]
- Cen, R.; Feng, W.; Yang, F.; Wu, W.; Liao, H.; Qu, Z. Effect mechanism of biochar application on soil structure and organic matter in semi-arid areas. J. Environ. Manag. 2021, 286, 112198. [Google Scholar] [CrossRef] [PubMed]
- Kong, C.; Camps-Arbestain, M.; Clothier, B.; Bishop, P.; Vázquez, F.M. Use of either pumice or willow-based biochar amendments to decrease soil salinity under arid conditions. Environ. Technol. Innov. 2021, 24, 101849. [Google Scholar] [CrossRef]
- Guo, M.; He, Z.; Uchimiya, S.M. Introduction to biochar as an agricultural and environmental amendment. In Agricultural and Environmental Applications of Biochar: Advances and Barriers; Guo, M., He, Z., Uchimiya, S.M., Eds.; Soil Science Society of America, Inc.: Madison, WI, USA, 2016; Volume 63, pp. 1–14. [Google Scholar]
- Hafez, E.; Omara, A.E.D.; Ahmed, A. The coupling effects of plant growth promoting rhizobacteria and salicylic acid on physiological modifications, yield traits, and productivity of wheat under water deficient conditions. Agronomy 2019, 9, 524. [Google Scholar] [CrossRef] [Green Version]
- IBI. Standardized Product Definition and Product Testing Guidelines for Biochar that is Used in Soil; International Biochar Initiative (IBI): Canandaigua, NY, USA, 2014. [Google Scholar]
- Sarfraz, R.; Hussain, A.; Sabir, A.; Ben Fekih, I.; Ditta, A.; Xing, S. Role of biochar and plant growth promoting rhizobacteria to enhance soil carbon sequestration—A review. Environ. Monit. Assess. 2019, 191, 251. [Google Scholar] [CrossRef]
- Ullah, S.; Bano, A. Isolation of plant-growth-promoting rhizobacteria from rhizospheric soil of halophytes and their impact on maize (Zea mays L.) under induced soil salinity. Can. J. Microbiol. 2015, 61, 307–313. [Google Scholar] [CrossRef] [PubMed]
- Niranjana, S.R.; Hariprasad, P. Understanding the mechanism involved in PGPR-mediated growth promotion and suppression of biotic and abiotic stress in plants. In Future Challenges in Crop Protection Against Fungal Pathogens; Goyal, A., Manoharachary, C., Eds.; Springer: New York, NY, USA, 2014; pp. 59–108. [Google Scholar] [CrossRef]
- Gaudin, V.; Vrain, T.; Jouanin, L. Bacterial genes modifying hormonal balances in plants. Plant Physiol. Biochem. 1994, 32, 11–29. [Google Scholar]
- Asghar, H.; Zahir, Z.; Arshad, M.; Khaliq, A. Relationship between in vitro production of auxins by rhizobacteria and their growth-promoting activities in Brassica juncea L. Biol. Fertil. Soils 2002, 35, 231–237. [Google Scholar] [CrossRef]
- Vessey, J.K. Plant growth promoting rhizobacteria as biofertilizers. Plant Soil 2003, 255, 571–586. [Google Scholar] [CrossRef]
- Glick, B.R.; Penrose, D.M.; Li, J. A Model for the lowering of plant ethylene concentrations by plant growth-promoting bacteria. J. Theor. Biol. 1998, 190, 63–68. [Google Scholar] [CrossRef]
- Khan, M.S.; Zaidi, A.; Wani, P.A. Role of phosphate-solubilizing microorganisms in sustainable agriculture—A review. Agron. Sustain. Dev. 2007, 27, 29–43. [Google Scholar] [CrossRef]
- Cohen, A.; Bottini, R.; Piccoli, P.N. Azospirillum brasilense sp. 245 produces ABA in chemically-defined culture medium and increases ABA content in arabidopsis plants. Plant Growth Regul. 2007, 54, 97–103. [Google Scholar] [CrossRef]
- Boiero, L.; Perrig, D.; Masciarelli, O.; Penna, C.; Cassán, F.; Luna, V. Phytohormone production by three strains of Bradyrhizobium japonicum and possible physiological and technological implications. Appl. Microbiol. Biotechnol. 2007, 74, 874–880. [Google Scholar] [CrossRef]
- Cohen, A.; Travaglia, C.N.; Bottini, R.; Piccoli, P.N. Participation of abscisic acid and gibberellins produced by endophytic Azospirillum in the alleviation of drought effects in maize. Botany 2009, 87, 455–462. [Google Scholar] [CrossRef]
- Devi, B.; Chitdeshwari, T. Physiological and morphological traits of Maize hybrids under saline water irrigation. Indian, J. Exp. Biol. 2019, 57, 188–194. [Google Scholar]
- Turan, M.A.; Elkarim, A.H.A.; Taban, N.; Taban, S. Effect of salt stress on growth, stomatal resistance, proline and chlorophyll concentrations on maize plant. Afr. J. Agric. Res. 2009, 4, 893–897. [Google Scholar]
- Alsaeedi, A.H.; El-Ramady, H.; Alshaal, T.; El-Garawani, M.; Elhawat, N.; Almohsen, M. Engineered silica nanoparticles alleviate the detrimental effects of Na+ stress on germination and growth of common bean (Phaseolus vulgaris). Environ. Sci. Pollut. Res. 2017, 24, 21917–21928. [Google Scholar] [CrossRef] [PubMed]
- Alsaeedi, A.; El-Ramady, H.; Alshaal, T.; El-Garawani, M.; Elhawat, N.; Al-Otaibi, A. Exogenous nanosilica improves germination and growth of cucumber by maintaining K+/Na+ ratio under elevated Na+ stress. Plant Physiol. Biochem. 2018, 125, 164–171. [Google Scholar] [CrossRef]
- Osman, H.; Gowayed, S.; Elbagory, M.; Omara, A.; El-Monem, A.; El-Razek, U.A.; Hafez, E. Interactive impacts of beneficial microbes and Si-Zn nanocomposite on growth and productivity of soybean subjected to water deficit under salt-affected soil conditions. Plants 2021, 10, 1396. [Google Scholar] [CrossRef]
- Flexas, J.; Diaz-Espejo, A.; Galmés, J.; Kaldenhoff, R.; Medrano, H.; Ribas-Carbo, M. Rapid variations of mesophyll conductance in response to changes in CO2 concentration around leaves. Plant, Cell Environ. 2007, 30, 1284–1298. [Google Scholar] [CrossRef]
- Di Martino, C.; Delfine, S.; Alvino, A.; Loreto, F. Photorespiration rate in spinach leaves under moderate NaCl stress. Photosynthetica 1999, 36, 233. [Google Scholar] [CrossRef]
- Marcelis, L.; Van Hooijdonk, J. Effect of salinity on growth, water use and nutrient use in radish (Raphanus sativus L.). Plant Soil 1999, 215, 57–64. [Google Scholar] [CrossRef]
- Habib, S.H.; Kausar, H.; Saud, H.M. Plant growth-promoting rhizobacteria enhance salinity stress tolerance in okra through ROS-scavenging enzymes. BioMed Res. Int. 2016, 2016, 6284547. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Machado, R.M.A.; Serralheiro, R.P. Soil salinity: Effect on vegetable crop growth. Management practices to prevent and mitigate soil salinization. Horticulturae 2017, 3, 30. [Google Scholar] [CrossRef]
- Chaganti, V.N.; Crohn, D.M. Evaluating the relative contribution of physiochemical and biological factors in ameliorating a saline–sodic soil amended with composts and biochar and leached with reclaimed water. Geoderma 2015, 259–260, 45–55. [Google Scholar] [CrossRef]
- Huang, M.; Zhang, Z.; Zhu, C.; Zhai, Y.; Lu, P. Effect of biochar on sweet corn and soil salinity under conjunctive irrigation with brackish water in coastal saline soil. Sci. Hortic. 2019, 250, 405–413. [Google Scholar] [CrossRef]
- Fukami, J.; De La Osa, C.; Ollero, F.J.; Megías, M.; Hungria, M. Co-inoculation of maize with Azospirillum brasilense and Rhizobium tropici as a strategy to mitigate salinity stress. Funct. Plant Biol. 2018, 45, 328–339. [Google Scholar] [CrossRef]
- Pandit, N.R.; Schmidt, H.-P.; Mulder, J.; Hale, S.E.; Husson, O.; Cornelissen, G. Nutrient effect of various composting methods with and without biochar on soil fertility and maize growth. Arch. Agron. Soil Sci. 2019, 66, 250–265. [Google Scholar] [CrossRef] [Green Version]
- Akhtar, S.S.; Andersen, M.N.; Naveed, M.; Zahir, Z.A.; Liu, F. Interactive effect of biochar and plant growth-promoting bacterial endophytes on ameliorating salinity stress in maize. Funct. Plant Biol. 2015, 42, 770. [Google Scholar] [CrossRef] [PubMed]
- Naeem, M.A.; Khalid, M.; Aon, M.; Abbas, G.; Tahir, M.; Amjad, M.; Murtaza, B.; Yang, A.; Akhtar, S.S. Effect of wheat and rice straw biochar produced at different temperatures on maize growth and nutrient dynamics of a calcareous soil. Arch. Agron. Soil Sci. 2017, 63, 2048–2061. [Google Scholar] [CrossRef]
- Kamaluldeen, J.; Yunusa, I.; Zerihun, A.; Bruhl, J.J.; Kristiansen, P. Uptake and distribution of ions reveal contrasting tolerance mechanisms for soil and water salinity in okra (Abelmoschus esculentus) and tomato (Solanum esculentum). Agric. Water Manag. 2014, 146, 95–104. [Google Scholar] [CrossRef]
- Elzobair, K.A.; Stromberger, M.E.; Ippolito, J.A.; Lentz, R.D. Contrasting effects of biochar versus manure on soil microbial communities and enzyme activities in an Aridisol. Chemosphere 2016, 142, 145–152. [Google Scholar] [CrossRef]
- Ahmad, M.T.; Asghar, H.N.; Saleem, M.; Khan, M.Y.; Zahir, Z.A. Synergistic effect of rhizobia and biochar on growth and physiology of maize. Agron. J. 2015, 107, 2327–2334. [Google Scholar] [CrossRef]
- Ahmed, F.; Arthur, E.; Plauborg, F.; Razzaghi, F.; Kørup, K.; Andersen, M.N. Biochar amendment of fluvio-glacial temperate sandy subsoil: Effects on maize water uptake, growth and physiology. J. Agron. Crop. Sci. 2017, 204, 123–136. [Google Scholar] [CrossRef]
- Kheir, A.M.S.; Abouelsoud, H.M.; Hafez, E.M.; Ali, O.A.M. Integrated effect of nano-Zn, nano-Si, and drainage using crop straw–filled ditches on saline sodic soil properties and rice productivity. Arab. J. Geosci. 2019, 12, 471. [Google Scholar] [CrossRef]
- Hafez, E.M.; Alsohim, A.S.; Farig, M.; Omara, A.E.-D.; Rashwan, E.; Kamara, M.M. Synergistic effect of biochar and plant growth promoting rhizobacteria on alleviation of water deficit in rice plants under salt-affected soil. Agronomy 2019, 9, 847. [Google Scholar] [CrossRef] [Green Version]
- Hafez, E.E.-D.M. Influence of salicylic acid on ion distribution, enzymatic activity and some agromorphological characteristics of wheat under salt-affected soil. Egypt. J. Agron. 2016, 38, 455–469. [Google Scholar] [CrossRef] [Green Version]
- Leogrande, R.; Vitti, C.; Lopedota, O.; Ventrella, D.; Montemurro, F.; Rita, L.; Carolina, V.; Ornella, L.; Domenico, V.; Francesco, M. Saline water and MSW compost: Effects on yield of maize crop and soil responses. J. Plant Nutr. 2016, 39, 1863–1873. [Google Scholar] [CrossRef]
- Hafez, E.M.; Gharib, H.S. Effect of exogenous application of ascorbic acid on physiological and biochemical characteristics of wheat under water stress. Int. J. Plant Prod. 2016, 10, 579–596. [Google Scholar]
- Calamai, A.; Chiaramonti, D.; Casini, D.; Masoni, A.; Palchetti, E. Short-term effects of organic amendments on soil properties and maize (Zea maize L.) growth. Agriculture 2020, 10, 158. [Google Scholar] [CrossRef]
- Li, H.Q.; Jiang, X.W. Inoculation with plant growth-promoting bacteria (PGPB) improves salt tolerance of maize seedling. Russ. J. Plant Physiol. 2017, 64, 235–241. [Google Scholar] [CrossRef]
- Hafez, E.; Osman, H.; Gowayed, S.; Okasha, S.; Omara, A.; Sami, R.; El-Monem, A.A.; El-Razek, U.A. Minimizing the adversely impacts of water deficit and soil salinity on maize growth and productivity in response to the application of plant growth-promoting rhizobacteria and silica nanoparticles. Agronomy 2021, 11, 676. [Google Scholar] [CrossRef]
- Ahmed, B.O.; Inoue, M.; Moritani, S. Effect of saline water irrigation and manure application on the available water content, soil salinity, and growth of wheat. Agric. Water Manag. 2010, 97, 165–170. [Google Scholar] [CrossRef]
- Jensen, H.L. Notes on the biology of azotobacter. Proc. Soc. Appl. Bacteriol. 1951, 14, 89–94. [Google Scholar] [CrossRef]
- King, O.E.; Ward, M.K.; Raney, E.D. Two simple media for the demonstration of pyocyanin and fluorescin. J. Lab. Clin. Med. 1954, 44, 301–307. [Google Scholar] [PubMed]
- Gharib, H.S.; El-Henawy, A.; Meleha, M. Influence of irrigation regimes at different growth stages on development and yield in maize (Zea mays, L.). Egypt. J. Agron. 2016, 38, 509–530. [Google Scholar] [CrossRef]
- Mousa, N.A.; Siaguru, P.; Wiryowidagdo, S.; Wagih, M.E. Evaluation and selection of elite clonal genotypes of the sweet crop licorice (Glycyrrhiza glabra) in a new environment. Sugar Tech 2007, 9, 83–94. [Google Scholar] [CrossRef]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid determination of free proline for water-stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Ibragimova, M.V.; Rumyantseva, M.L.; Onishchuk, O.P.; Belova, V.S.; Kurchak, O.N.; Andronov, E.; Dzyubenko, N.; Simarov, B.V. Symbiosis between the root-nodule bacterium Sinorhizobium meliloti and alfalfa (Medicago sativa) under salinization conditions. Microbiology 2006, 75, 77–81. [Google Scholar] [CrossRef]
- Barrs, H.; Weatherley, P. A re-examination of the relative turgidity technique for estimating water deficits in leaves. Aust. J. Biol. Sci. 1962, 15, 413–428. [Google Scholar] [CrossRef] [Green Version]
- Temminghoff, E.E.J.M.; Houba, V.J.G. Plant Analysis Procedures; Springer: Berlin/Heidelberg, Germany, 2004; p. 179. [Google Scholar]
- AOAC. Official Methods of Analysis of the Association of Official Analytical Chemists; AOAC: Rockville, MD, USA, 1975; p. 1018. [Google Scholar]
- Sparks, D.L.; Page, A.L.; Helmke, P.A.; Loeppert, R.H. Methods of Soil Analysis Part 3—Chemical Methods; SSSA Book Series; Soil Science Society of America, American Society of Agronomy: Madison, WI, USA, 1996. [Google Scholar]
- Filep, G.Y. Soil Analysis: Lecture Notes; University of Agricultural Sciences: Debrecen, Hungary, 1995. (In Hungarian) [Google Scholar]
- Mersi, V.W. Enzymes involved in intracellular metabolism: Dehydrogenase activity with the substrate INT. In Methods in Soil Biology; Schinner, F., Öhlinger, R., Kandeler, E., Margesin, R., Eds.; Springer: Berlin/Heidelberg, Germany, 1996; pp. 243–245. [Google Scholar]
- Abd-El-Malek, Y.; Ishac, Y.Z. Evaluation of methods used in counting azotobacters. J. Appl. Bacteriol. 1968, 31, 267–275. [Google Scholar] [CrossRef]
- Cochran, W.G. Estimation of bacterial densities by means of the “most probable number”. Biometrics 1950, 6, 105. [Google Scholar] [CrossRef]
- Casida, L.E.; Klein, D.A.; Santoro, T. Soil dehydrogenase activity. Soil Sci. 1964, 98, 371–376. [Google Scholar] [CrossRef]
- Seilsepour, M.; Rashidi, M.; Khabbaz, B.G. Prediction of soil exchangeable sodium percentage based on soil sodium adsorption ratio. Am. J. Agric. Environ. Sci. 2009, 5, 1–4. [Google Scholar]
- Richards, L.A. Diagnosis and Improvement of Saline and Alkali Soils; Richards, L.A., Ed.; United States Department of Agriculture: Washington, DC, USA, 1954.
- SAS. JMP® Data Analysis Software—Version 15; SAS Institute Inc.: Cary, NC, USA, 1989–2021; JMP Homepage; Available online: http://www.jmp.com/ (accessed on 15 February 2021).




{kind=link}
{kind=link}
{kind=link}
| Treatment | Urease (mg NH4+ g−1 Dry Soil d−1) | Dehydrogenase (mg TPF g−1 Dry Soil d−1) | Bacteria (Log cfu g−1 Soil) | Azotobacter (Log cfu g−1 Soil) | Bacillus spp. (Log cfu g−1 Soil) | |
|---|---|---|---|---|---|---|
| Fresh water | Control | 128.86 ± 3.60 d | 64.01 ± 1.52 e | 3.44 ± 0.05 e | 0.93 ± 0.01 g | 1.74 ± 0.02 e |
| PGPB † | 165.26 ± 2.65 c | 95.85 ± 1.60 c | 4.23 ± 0.06 d | 1.11 ± 0.01 f | 2.14 ± 0.10 d | |
| Biochar ‡ | 187.38 ± 2.16 b | 115.97 ± 1.79 b | 5.82 ± 0.07 b | 1.63 ± 0.01 c | 3.13 ± 0.03 b | |
| PGPR + biochar ¥ | 229.32 ± 3.51 a | 156.14 ± 3.34 b | 6.02 ± 0.07 a | 2.13 ± 0.01 a | 3.75 ± 0.04 a | |
| Saline water | Control | 99.29 ± 2.10 e | 49.92 ± 2.41 f | 2.35 ± 0.04 f | 0.63 ± 0.02 h | 1.07 ± 0.02 f |
| PGPB | 121.39 ± 3.57 d | 80.65 ± 2.65 d | 3.45 ± 0.05 e | 1.23 ± 0.02 e | 1.75 ± 0.02 e | |
| Biochar | 127.66 ± 2.66 d | 94.22 ± 1.64 c | 3.54 ± 0.05 e | 1.39 ± 0.01 d | 1.85 ± 0.03 e | |
| PGPR + biochar | 165.51 ± 3.05 c | 121.44 ± 1.00 a | 5.25 ± 0.04 c | 1.89 ± 0.01 b | 2.95 ± 0.03 c | |
| F-test | ||||||
| pWater | <0.0001 | <0.0001 | <0.0001 | <0.0001 | <0.0001 | |
| pTreatment | <0.0001 | <0.0001 | <0.0001 | <0.0001 | <0.0001 | |
| pWater × Treatment | <0.0001 | <0.0001 | <0.0001 | <0.0001 | <0.0001 | |
| Treatment | Number of Grains Ear−1 | 100-Grain Weight (g) | Grain Yield (ton/ha) | Stover Yield (ton/ha) | Harvest Index (%) | |
|---|---|---|---|---|---|---|
| Fresh water | Control | 425.27 ± 1.6 d | 30.45 ± 0.49 c | 4.55 ± 0.04 e | 8.86 ± 0.06 e | 33.95 ± 0.15 d |
| PGPB † | 436.69 ± 0.9 b | 32.30 ± 0.32 b | 5.22 ± 0.03 c | 9.76 ± 0.07 c | 34.87 ± 0.26 cd | |
| Biochar ‡ | 437.94 ± 0.9 b | 33.55 ± 0.47 b | 5.43 ± 0.03 b | 9.92 ± 0.04 b | 35.36 ± 0.66 bc | |
| PGPR + biochar ¥ | 446.63 ± 0.9 a | 35.77 ± 0.96 a | 5.95 ± 0.02 a | 10.29 ± 0.04 a | 36.66 ± 0.14 a | |
| Saline water | Control | 408.83 ± 0.62 f | 23.38 ± 0.67 e | 4.3 ± 0.05 f | 8.43 ± 0.07 f | 33.79 ± 0.29 d |
| PGPB | 421.45 ± 1.25 e | 27.13 ± 0.78 d | 4.63 ± 0.06 e | 8.87 ± 0.04 e | 34.32 ± 0.37 cd | |
| Biochar | 424.84 ± 0.82 d | 28.32 ± 0.67 d | 4.76 ± 0.04 d | 8.95 ± 0.03 e | 34.70 ± 0.21 cd | |
| PGPR + biochar | 429.65 ± 0.71 c | 32.72 ± 0.31 b | 5.26 ± 0.02 c | 9.36 ± 0.08 d | 35.97 ± 0.63 ab | |
| F-test | ||||||
| pWater | <0.0001 | <0.0001 | <0.0001 | <0.0001 | <0.0001 | |
| pTreatment | <0.0001 | <0.0001 | <0.0001 | <0.0001 | <0.0001 | |
| pWater × Treatment | =0.0196 | =0.0004 | <0.0001 | <0.0001 | =0.0310 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nehela, Y.; Mazrou, Y.S.A.; Alshaal, T.; Rady, A.M.S.; El-Sherif, A.M.A.; Omara, A.E.-D.; Abd El-Monem, A.M.; Hafez, E.M. The Integrated Amendment of Sodic-Saline Soils Using Biochar and Plant Growth-Promoting Rhizobacteria Enhances Maize (Zea mays L.) Resilience to Water Salinity. Plants 2021, 10, 1960. https://doi.org/10.3390/plants10091960
Nehela Y, Mazrou YSA, Alshaal T, Rady AMS, El-Sherif AMA, Omara AE-D, Abd El-Monem AM, Hafez EM. The Integrated Amendment of Sodic-Saline Soils Using Biochar and Plant Growth-Promoting Rhizobacteria Enhances Maize (Zea mays L.) Resilience to Water Salinity. Plants. 2021; 10(9):1960. https://doi.org/10.3390/plants10091960
Chicago/Turabian StyleNehela, Yasser, Yasser S. A. Mazrou, Tarek Alshaal, Asmaa M. S. Rady, Ahmed M. A. El-Sherif, Alaa El-Dein Omara, Ahmed M. Abd El-Monem, and Emad M. Hafez. 2021. "The Integrated Amendment of Sodic-Saline Soils Using Biochar and Plant Growth-Promoting Rhizobacteria Enhances Maize (Zea mays L.) Resilience to Water Salinity" Plants 10, no. 9: 1960. https://doi.org/10.3390/plants10091960