
The Steroid Saponin Protodioscin Modulates Arabidopsis thaliana Root Morphology Altering Auxin Homeostasis, Transport and Distribution

 ,
, 
Abstract
:1. Introduction
2. Results
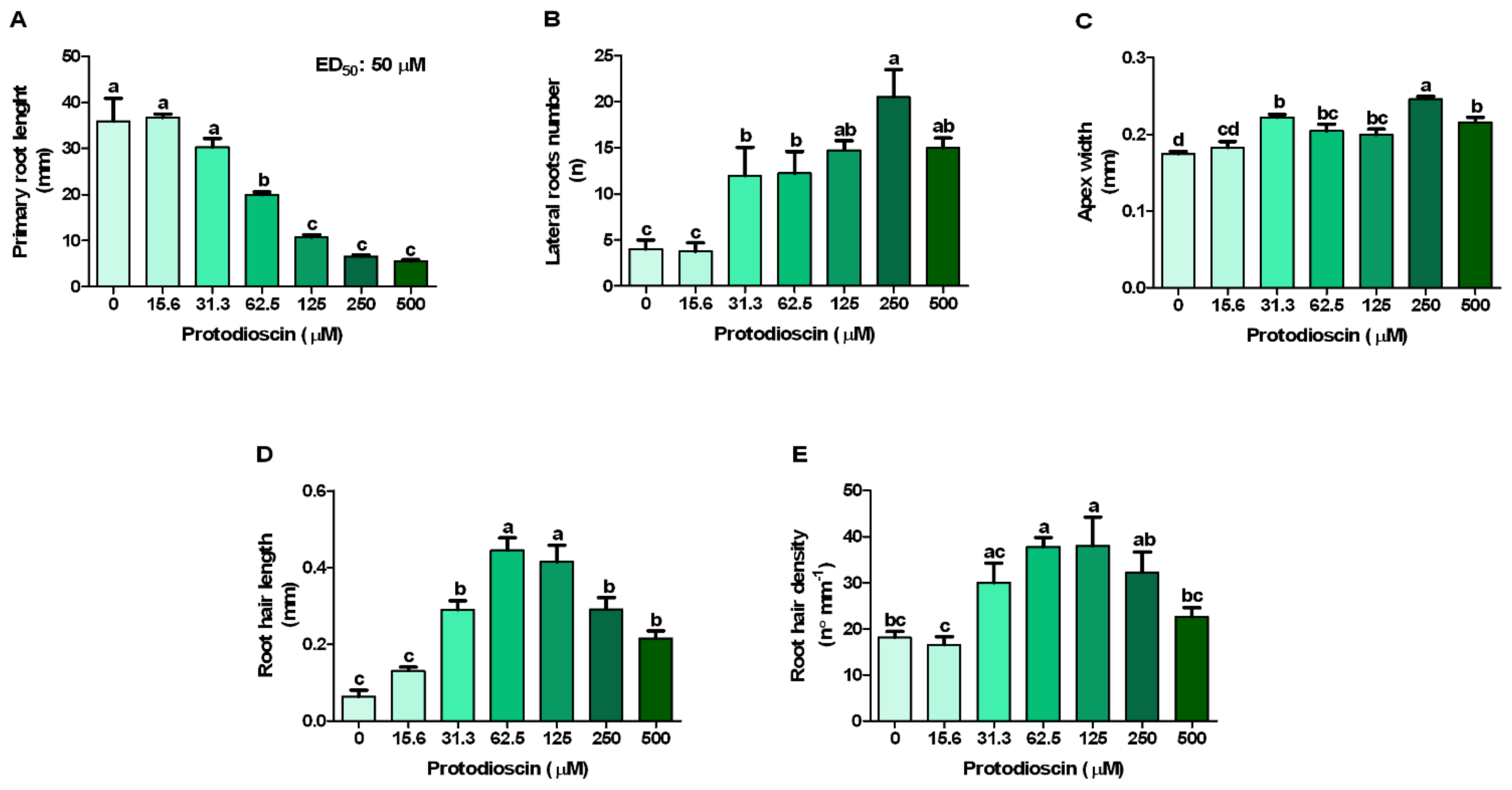
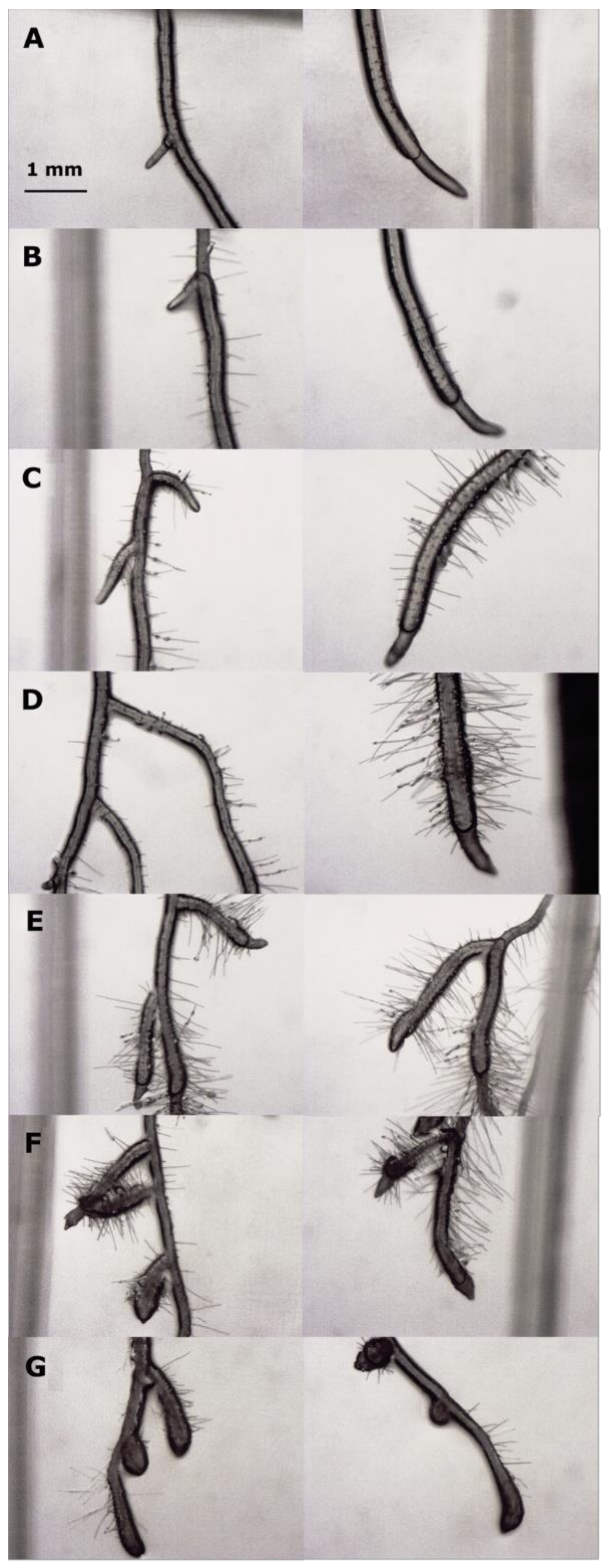
2.1. Dose-Response Curves
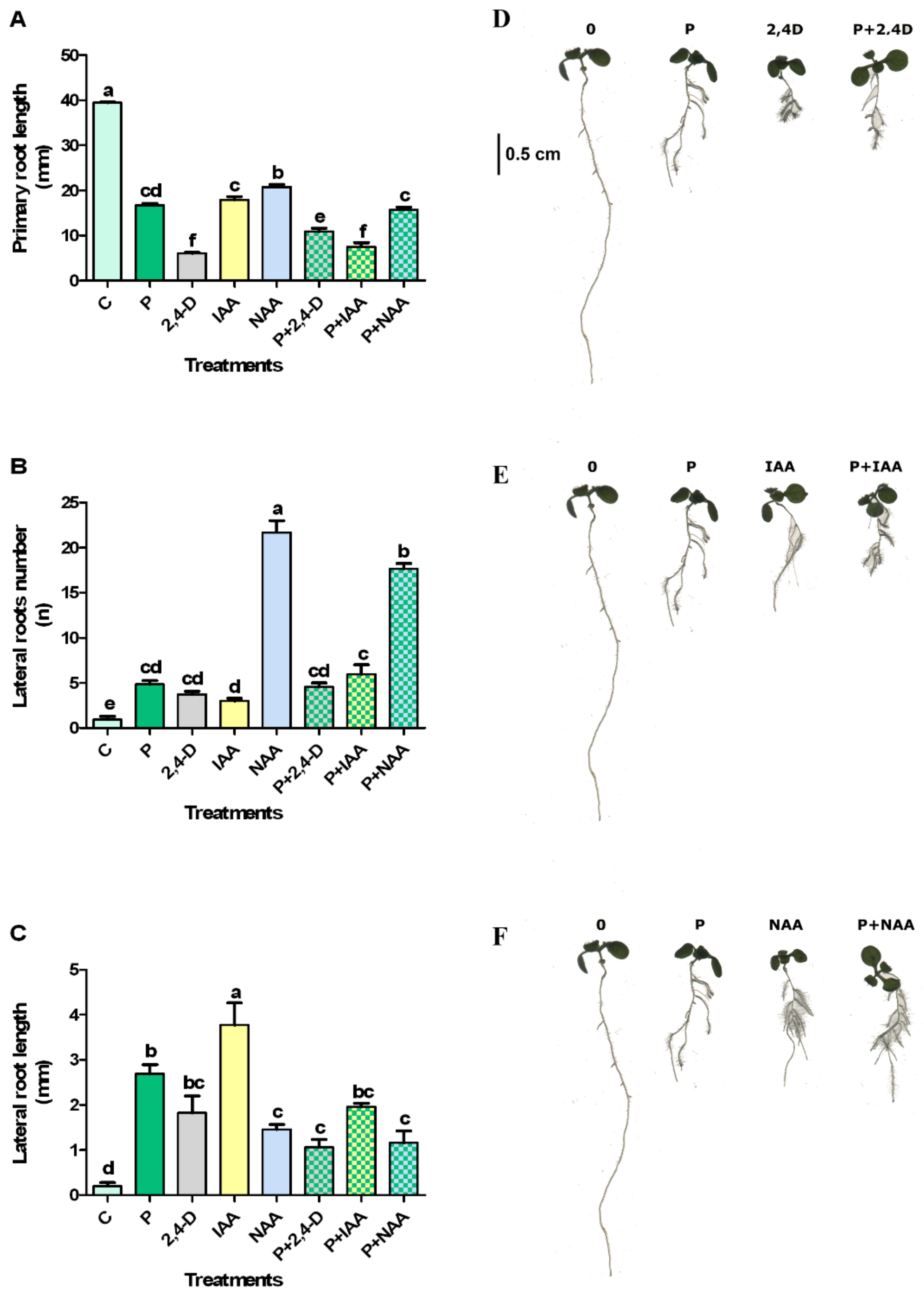
2.2. Protodioscin and Natural/Synthetic Auxin Interactions
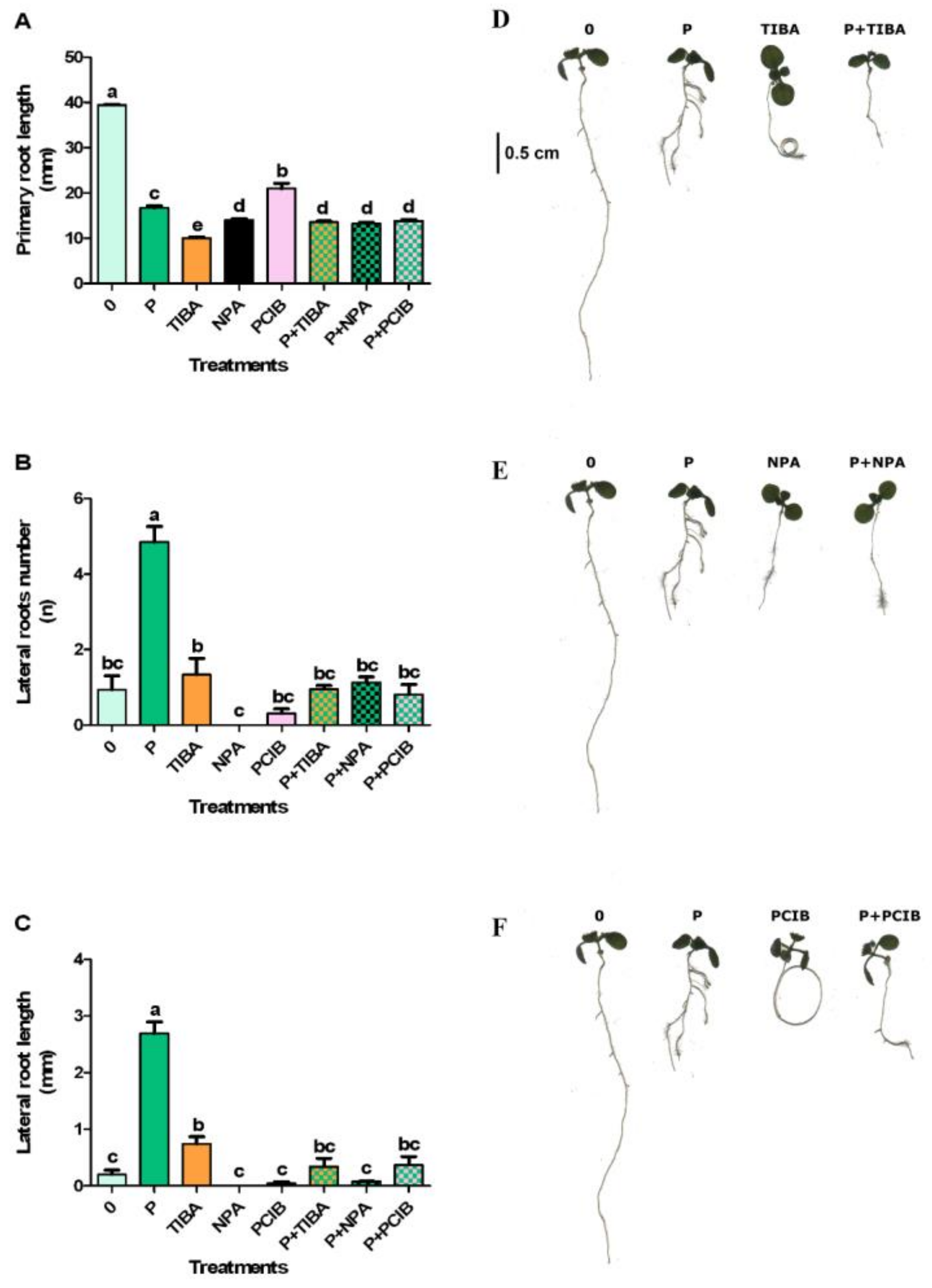
2.3. Effects of Protodioscin and Auxin Inhibitors
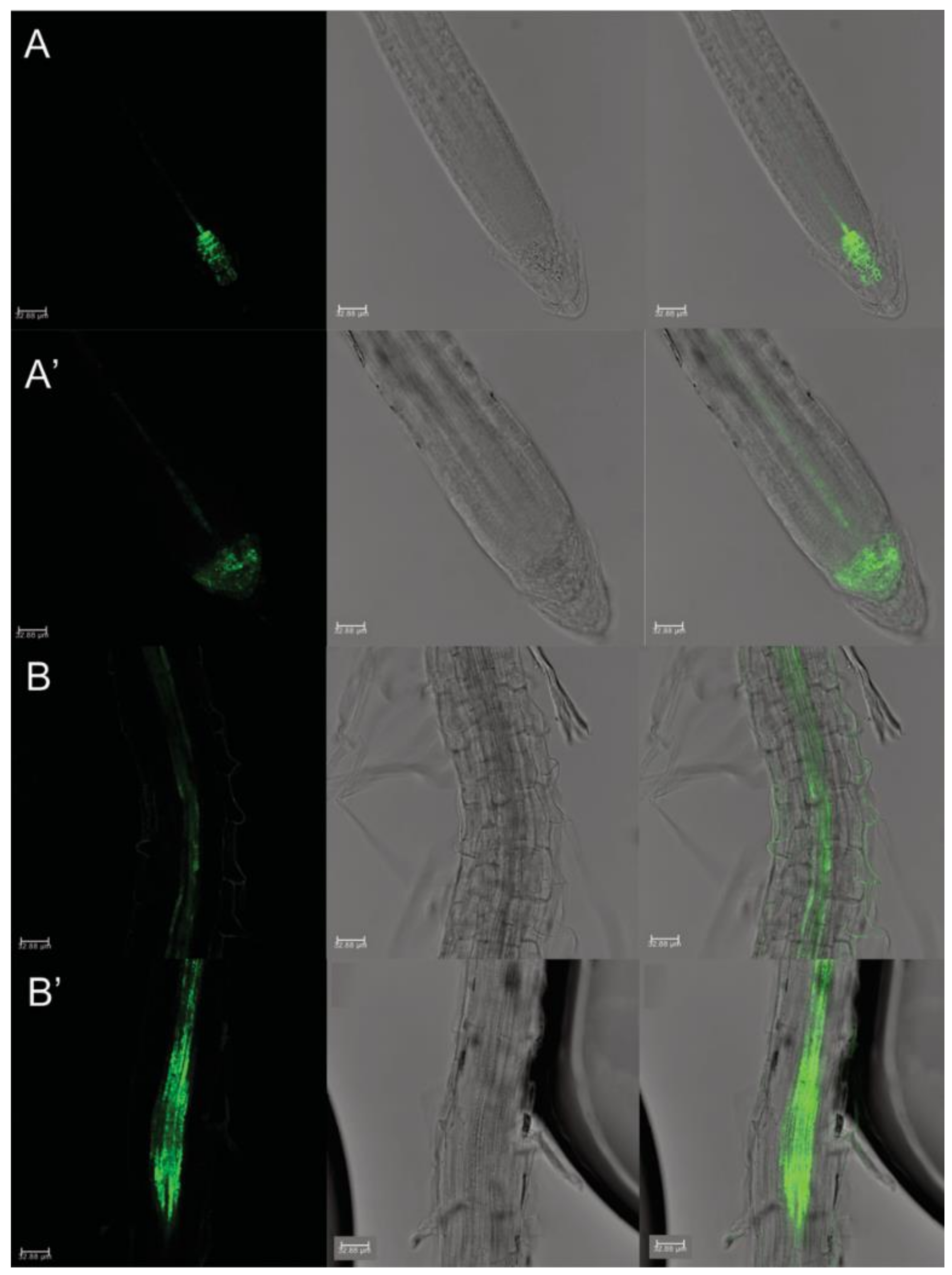
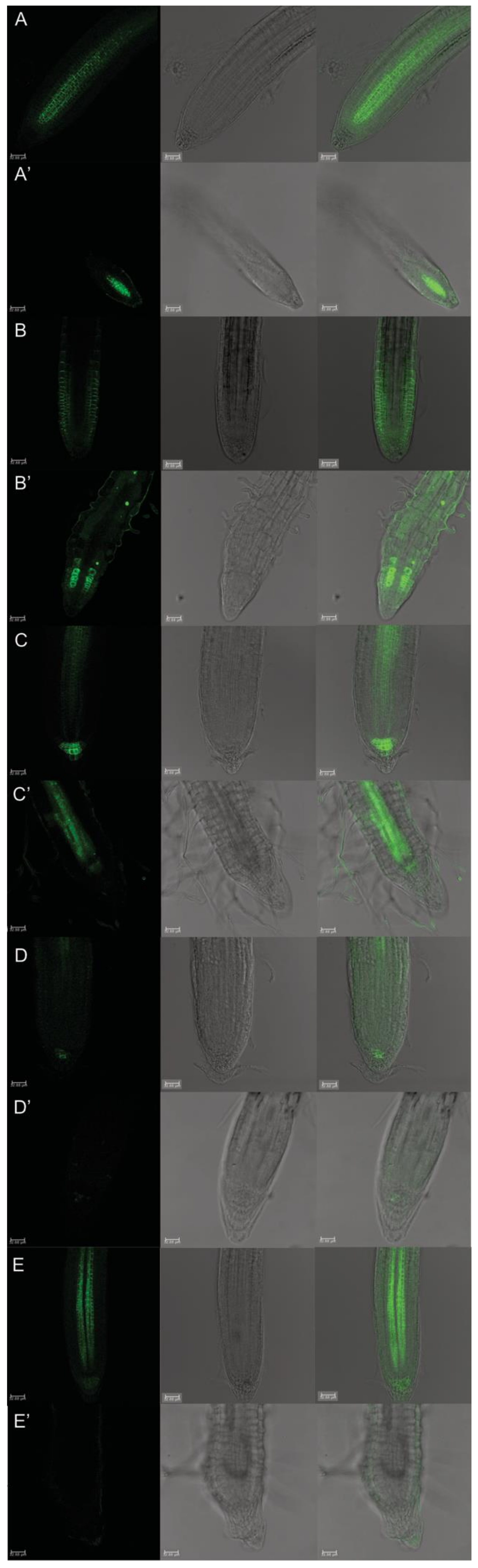
2.4. Auxin Distribution through Auxin-Responsive Reporter pDR5::GFP and Its Relative Quantification
2.5. Protodioscin Affected Auxin Gradient and Polar Transport
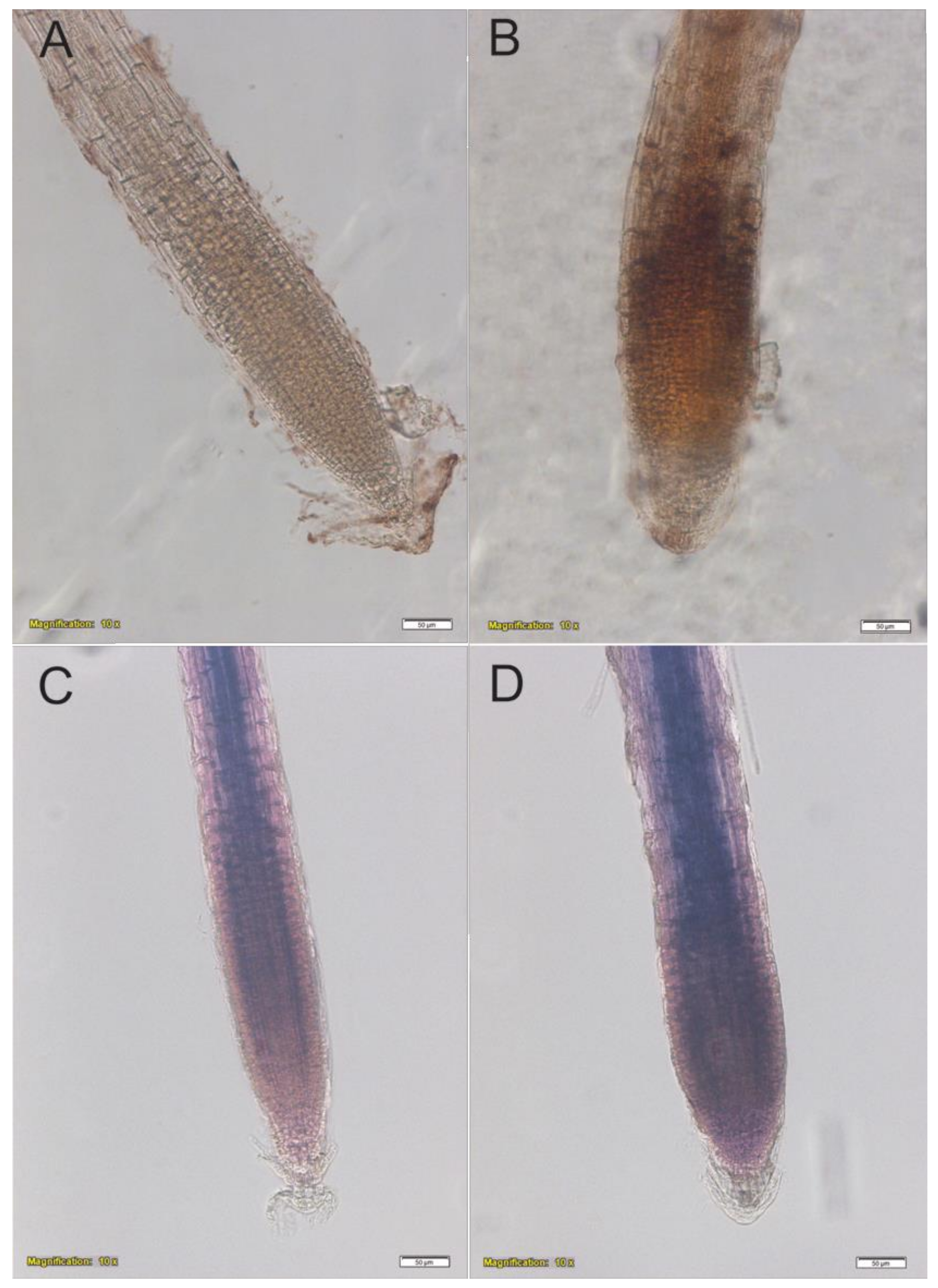
2.6. In Situ Semi-Quantitative Determination of H2O2 and O2−
3. Discussion
4. Materials and Methods
4.1. Plant Material and Experimental Design
4.2. Protodioscin and Auxins Interaction in Roots of A. thaliana
4.3. Arabidopsis Transgenic Reporter Lines Bioassay
4.4. In Situ Semi-Quantitative Determination of H2O2 and O2−
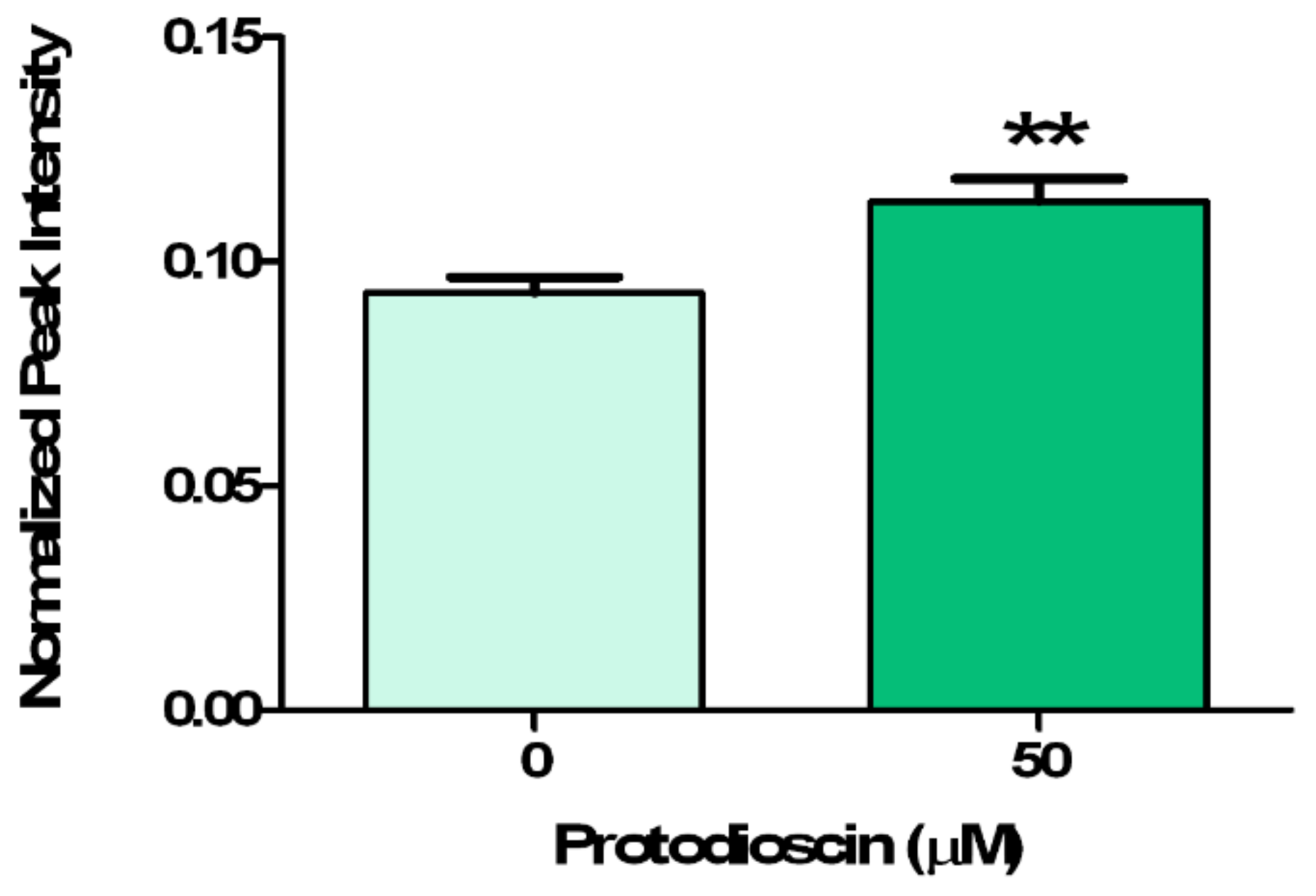
4.5. IAA Relative Quantification through GC-MS Analysis
4.6. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Chauhan, B.S. Grand challenges in weed management. Front. Agron. 2020, 1, 3. [Google Scholar] [CrossRef]
- Monaco, T.J.; Weller, S.C.; Ashton, F.M. Weed Science: Principles and Practices; John Wiley & Sons: Hoboken, NJ, USA, 2002. [Google Scholar]
- Clapp, J. Explaining growing glyphosate use: The political economy of herbicide-dependent agriculture. Glob. Environ. Chang. 2021, 67, 102239. [Google Scholar] [CrossRef]
- Rosculete, C.A.; Bonciu, E.; Rosculete, E.; Olaru, L.A. Determination of the environmental pollution potential of some herbicides by the assessment of cytotoxic and genotoxic effects on Allium cepa. Int. J. Environ. Res. Public Health 2019, 16, 75. [Google Scholar] [CrossRef] [Green Version]
- Heap, I. International Herbicide-Resistant Weed Database. 2019. Available online: www.weedscience.org (accessed on 26 February 2020).
- Al-Samarai, G.F.; Mahdi, W.M.; Al-Hilali, B.M. Reducing environmental pollution by chemical herbicides using natural plant derivatives—Allelopathy effect. Ann. Agric. Environ. Med. 2018, 25, 449–452. [Google Scholar] [CrossRef] [PubMed]
- Verdeguer, M.; Sánchez-Moreiras, A.M.; Araniti, F. Phytotoxic effects and mechanism of action of essential oils and terpenoids. Plants 2020, 9, 1571. [Google Scholar] [CrossRef] [PubMed]
- Dayan, F.E. Is there a natural route to the next generation of herbicides? Outlooks Pest Manag. 2018, 29, 54–57. [Google Scholar] [CrossRef]
- Dayan, F.E. Current status and future prospects in herbicide discovery. Plants 2019, 8, 341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Araniti, F.; Mancuso, R.; Lupini, A.; Giofrè, S.V.; Sunseri, F.; Gabriele, B.; Abenavoli, M.R. Phytotoxic potential and biological activity of three synthetic coumarin derivatives as new natural-like herbicides. Molecules 2015, 20, 17883–17902. [Google Scholar] [CrossRef] [Green Version]
- Araniti, F.; Mancuso, R.; Lupini, A.; Sunseri, F.; Abenavoli, M.R.; Gabriele, B. Benzofuran-2-acetic esters as a new class of natural-like herbicides. Pest Manag. Sci. 2020, 76, 395–404. [Google Scholar] [CrossRef]
- Dayan, F.E.; Duke, S.O. Natural compounds as next-generation herbicides. Plant Physiol. 2014, 166, 1090–1105. [Google Scholar] [CrossRef] [Green Version]
- Aşkin Çelik, T.; Aslantürk, Ö.S. Evaluation of cytotoxicity and genotoxicity of Inula viscosa leaf extracts with Allium test. J. Biomed. Biotechnol. 2010, 2010, 189252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lupini, A.; Araniti, F.; Mauceri, A.; Princi, M.; Sorgonà, A.; Sunseri, F.; Varanini, Z.; Abenavoli, M. Coumarin enhances nitrate uptake in maize roots through modulation of plasma membrane H+-ATPase activity. Plant Biol. 2018, 20, 390–398. [Google Scholar] [CrossRef]
- Scavo, A.; Abbate, C.; Mauromicale, G. Plant allelochemicals: Agronomic, nutritional and ecological relevance in the soil system. Plant Soil 2019, 442, 23–48. [Google Scholar] [CrossRef]
- Araniti, F.; Landi, M.; Lupini, A.; Sunseri, F.; Guidi, L.; Abenavoli, M. Origanum vulgare essential oils inhibit glutamate and aspartate metabolism altering the photorespiratory pathway in Arabidopsis thaliana seedlings. J. Plant Physiol. 2018, 231, 297–309. [Google Scholar] [CrossRef] [PubMed]
- Araniti, F.; Miras-Moreno, B.; Lucini, L.; Landi, M.; Abenavoli, M.R. Metabolomic, proteomic and physiological insights into the potential mode of action of thymol, a phytotoxic natural monoterpenoid phenol. Plant Physiol. Biochem. 2020, 153, 141–153. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Moreiras, A.M.; Graña, E.; Reigosa, M.J.; Araniti, F. Imaging of chlorophyll a fluorescence in natural compound-induced stress detection. Front. Plant Sci. 2020, 11, 1991. [Google Scholar] [CrossRef] [PubMed]
- Ishii-Iwamoto, E.; Abrahim, D.; Sert, M.; Bonato, C.; Kelmer-Bracht, A.; Bracht, A. Mitochondria as a site of allelochemical action. In Allelopathy; Springer: Berlin/Heidelberg, Germany, 2006; pp. 267–284. [Google Scholar]
- Landi, M.; Misra, B.B.; Muto, A.; Bruno, L.; Araniti, F. Phytotoxicity, morphological, and metabolic effects of the sesquiterpenoid nerolidol on Arabidopsis thaliana seedling roots. Plants 2020, 9, 1347. [Google Scholar] [CrossRef]
- Na, X.; Hu, Y.; Yue, K.; Lu, H.; Jia, P.; Wang, H.; Wang, X.; Bi, Y. Narciclasine modulates polar auxin transport in Arabidopsis roots. J. Plant Physiol. 2011, 168, 1149–1156. [Google Scholar] [CrossRef] [PubMed]
- Céspedes, C.L.; Marín, J.C.; Domínguez, M.; Avila, J.G.; Serrato, B. Plant growth inhibitory activities by secondary metabolites isolated from Latin American flora. Adv. Phytomed. 2006, 2, 373–410. [Google Scholar]
- Lotina-Hennsen, B.; King-Diaz, B.; Aguilar, M.; Terrones, M.H. Plant secondary metabolites. Targets and mechanisms of allelopathy. In Allelopathy; Springer: Berlin/Heidelberg, Germany, 2006; pp. 229–265. [Google Scholar]
- Zhang, W.; Lu, L.-Y.; Hu, L.-Y.; Cao, W.; Sun, K.; Sun, Q.-B.; Siddikee, A.; Shi, R.-H.; Dai, C.-C. Evidence for the involvement of auxin, ethylene and ROS signaling during primary root inhibition of arabidopsis by the allelochemical benzoic acid. Plant Cell Physiol. 2018, 59, 1889–1904. [Google Scholar] [CrossRef] [PubMed]
- López-González, D.; Costas-Gil, A.; Reigosa, M.J.; Araniti, F.; Sánchez-Moreiras, A.M. A natural indole alkaloid, norharmane, affects PIN expression patterns and compromises root growth in Arabidopsis thaliana. Plant Physiol. Biochem. 2020, 151, 378–390. [Google Scholar] [CrossRef]
- Araniti, F.; Grana, E.; Krasuska, U.; Bogatek, R.; Reigosa, M.J.; Abenavoli, M.R.; Sanchez-Moreiras, A.M. Loss of gravitropism in farnesene-treated arabidopsis is due to microtubule malformations related to hormonal and ROS unbalance. PLoS ONE 2016, 11, e0160202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lupini, A.; Araniti, F.; Sunseri, F.; Abenavoli, M.R. Coumarin interacts with auxin polar transport to modify root system architecture in Arabidopsis thaliana. Plant Growth Regul. 2014, 74, 23–31. [Google Scholar] [CrossRef]
- Graña, E.; Sotelo, T.; Díaz-Tielas, C.; Araniti, F.; Krasuska, U.; Bogatek, R.; Reigosa, M.J.; Sánchez-Moreiras, A.M. Citral induces auxin and ethylene-mediated malformations and arrests cell division in Arabidopsis thaliana roots. J. Chem. Ecol. 2013, 39, 271–282. [Google Scholar] [CrossRef] [Green Version]
- Wu, J.; Long, J.; Lin, X.; Chang, Z.; Baerson, S.R.; Ding, C.; Wu, X.; Pan, Z.; Song, Y.; Zeng, R. Momilactone B inhibits Arabidopsis growth and development via disruption of ABA and auxin signaling. bioRxiv 2020. [Google Scholar] [CrossRef]
- Teale, W.D.; Paponov, I.A.; Palme, K. Auxin in action: Signalling, transport and the control of plant growth and development. Nat. Rev. Mol. Cell Biol. 2006, 7, 847–859. [Google Scholar] [CrossRef]
- Kleine-Vehn, J.; Ding, Z.; Jones, A.R.; Tasaka, M.; Morita, M.T.; Friml, J. Gravity-induced PIN transcytosis for polarization of auxin fluxes in gravity-sensing root cells. Proc. Natl. Acad. Sci. USA 2010, 107, 22344–22349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Im Kim, J.; Sharkhuu, A.; Jin, J.B.; Li, P.; Jeong, J.C.; Baek, D.; Lee, S.Y.; Blakeslee, J.J.; Murphy, A.S.; Bohnert, H.J. yucca6, a dominant mutation in Arabidopsis, affects auxin accumulation and auxin-related phenotypes. Plant Physiol. 2007, 145, 722–735. [Google Scholar] [CrossRef] [Green Version]
- Sauer, M.; Balla, J.; Luschnig, C.; Wiśniewska, J.; Reinöhl, V.; Friml, J.; Benková, E. Canalization of auxin flow by Aux/IAA-ARF-dependent feedback regulation of PIN polarity. Genes Dev. 2006, 20, 2902–2911. [Google Scholar] [CrossRef] [Green Version]
- Steenackers, W.; Cesarino, I.; Klíma, P.; Quareshy, M.; Vanholme, R.; Corneillie, S.; Kumpf, R.P.; Van de Wouwer, D.; Ljung, K.; Goeminne, G. The allelochemical MDCA inhibits lignification and affects auxin homeostasis. Plant Physiol. 2016, 172, 874–888. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Gruber, M.Y.; Hegedus, D.D.; Lydiate, D.J.; Gao, M.-J. Effects of a coumarin derivative, 4-methylumbelliferone, on seed germination and seedling establishment in Arabidopsis. J. Chem. Ecol. 2011, 37, 880. [Google Scholar] [CrossRef] [PubMed]
- Graña, E.; Costas-Gil, A.; Longueira, S.; Celeiro, M.; Teijeira, M.; Reigosa, M.J.; Sánchez-Moreiras, A.M. Auxin-like effects of the natural coumarin scopoletin on Arabidopsis cell structure and morphology. J. Plant Physiol. 2017, 218, 45–55. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Na, X.; Li, J.; Yang, L.; You, J.; Liang, X.; Wang, J.; Peng, L.; Bi, Y. Narciclasine, a potential allelochemical, affects subcellular trafficking of auxin transporter proteins and actin cytoskeleton dynamics in Arabidopsis roots. Planta 2015, 242, 1349–1360. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Yang, L.; Na, X.; You, J.; Hu, W.; Liang, X.; Liu, J.; Mao, L.; Wang, X.; Wang, H. Narciclasine inhibits the responses of Arabidopsis roots to auxin. Planta 2012, 236, 597–612. [Google Scholar] [CrossRef]
- Jones, D.; Blancaflor, E.; Kochian, L.; Gilroy, S. Spatial coordination of aluminium uptake, production of reactive oxygen species, callose production and wall rigidification in maize roots. Plant Cell Environ. 2006, 29, 1309–1318. [Google Scholar] [CrossRef]
- Zhang, P.; Luo, Q.; Wang, R.; Xu, J. Hydrogen sulfide toxicity inhibits primary root growth through the ROS-NO pathway. Sci. Rep. 2017, 7, 868. [Google Scholar] [CrossRef]
- Dunand, C.; Crèvecoeur, M.; Penel, C. Distribution of superoxide and hydrogen peroxide in Arabidopsis root and their influence on root development: Possible interaction with peroxidases. New Phytol. 2007, 174, 332–341. [Google Scholar] [CrossRef]
- Romero-Puertas, M.; Rodríguez-Serrano, M.; Corpas, F.; Gomez, M.D.; Del Rio, L.; Sandalio, L. Cadmium-induced subcellular accumulation of O2·− and H2O2 in pea leaves. Plant Cell Environ. 2004, 27, 1122–1134. [Google Scholar] [CrossRef]
- Liu, G.; Gao, S.; Tian, H.; Wu, W.; Robert, H.S.; Ding, Z. Local transcriptional control of YUCCA regulates auxin promoted root-growth inhibition in response to aluminium stress in Arabidopsis. PLoS Genet. 2016, 12, e1006360. [Google Scholar] [CrossRef] [PubMed]
- Gauthaman, K.; Ganesan, A.P.; Prasad, R. Sexual effects of puncturevine (Tribulus terrestris) extract (protodioscin): An evaluation using a rat model. J. Altern. Complementary Med. 2003, 9, 257–265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leal, E.S.; Ítavo, L.C.V.; do Valle, C.B.; Ítavo, C.C.B.F.; Dias, A.M.; dos Santos Difante, G.; Barbosa-Ferreira, M.; Nonato, L.M.; de Melo, G.K.A.; Gurgel, A.L.C. Influence of protodioscin content on digestibility and in vitro degradation kinetics in Urochloa brizantha cultivars. Crop Pasture Sci. 2020, 71, 278–284. [Google Scholar] [CrossRef]
- Sao Miguel, A.; Pacheco, L.; Souza, E.; Silva, C.; Carvalho, I. Cover crops in the weed management in soybean culture. Planta Daninha 2018, 36. [Google Scholar] [CrossRef]
- Oliveira, R., Jr.; Rios, F.; Constantin, J.; Ishii-Iwamoto, E.; Gemelli, A.; Martini, P. Grass straw mulching to suppress emergence and early growth of weeds. Planta Daninha 2014, 32, 11–17. [Google Scholar] [CrossRef] [Green Version]
- Silva, A. Isolamento e identificacao de aleloquımicos de diversas especies de plantas com potencial herbicida. Dissertation Univ. Mar. Braz. 2012, 115. [Google Scholar]
- Nepomuceno, M.; Chinchilla, N.; Varela, R.M.; Molinillo, J.M.; Lacret, R.; Alves, P.L.; Macias, F.A. Chemical evidence for the effect of Urochloa ruziziensis on glyphosate-resistant soybeans. Pest Manag. Sci. 2017, 73, 2071–2078. [Google Scholar] [CrossRef]
- Mito, M.; Silva, A.; Kagami, F.; Almeida, J.; Mantovanelli, G.; Barbosa, M.; Kern-Cardoso, K.; Ishii-Iwamoto, E. Responses of the weed Bidens pilosa L. to exogenous application of the steroidal saponin protodioscin and plant growth regulators 24-epibrassinolide, indol-3-acetic acid and abscisic acid. Plant Biol. 2019, 21, 326–335. [Google Scholar]
- Menezes, P.V.M.C.; Silva, A.A.; Mito, M.S.; Mantovanelli, G.C.; Stulp, G.F.; Wagner, A.L.; Constantin, R.P.; Baldoqui, D.C.; Gonçales Silva, R.; Oliveira do Carmo, A.A.; et al. Morphogenic responses and biochemical alterations induced by the cover crop Urochloa ruziziensis and its component protodioscin in weed species. Plant Physiol. Biochem. 2021, 166, 857–873. [Google Scholar] [CrossRef]
- Pennacchio, M.; Jefferson, L.V.; Havens, K. Arabidopsis thaliana: A new test species for phytotoxic bioassays. J. Chem. Ecol. 2005, 31, 1877–1885. [Google Scholar] [CrossRef]
- Reigosa, M.; Pazos-Malvido, E. Phytotoxic effects of 21 plant secondary metabolites on Arabidopsis thaliana germination and root growth. J. Chem. Ecol. 2007, 33, 1456–1466. [Google Scholar] [CrossRef]
- Overvoorde, P.; Fukaki, H.; Beeckman, T. Auxin control of root development. Cold Spring Harb. Perspect. Biol. 2010, 2, a001537. [Google Scholar] [CrossRef] [Green Version]
- Abenavoli, M.R.; Sorgonà, A.; Albano, S.; Cacco, G. Coumarin differentially affects the morphology of different root types of maize seedlings. J. Chem. Ecol. 2004, 30, 1871–1883. [Google Scholar] [CrossRef] [PubMed]
- Lupini, A.; Sorgonà, A.; Miller, A.J.; Abenavoli, M.R. Short-term effects of coumarin along the maize primary root axis. Plant Signal. Behav. 2010, 5, 1395–1400. [Google Scholar] [CrossRef]
- Baskin, T.I.; Beemster, G.T.; Judy-March, J.E.; Marga, F. Disorganization of cortical microtubules stimulates tangential expansion and reduces the uniformity of cellulose microfibril alignment among cells in the root of Arabidopsis. Plant Physiol. 2004, 135, 2279–2290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chaimovitsh, D.; Abu-Abied, M.; Belausov, E.; Rubin, B.; Dudai, N.; Sadot, E. Microtubules are an intracellular target of the plant terpene citral. Plant J. 2010, 61, 399–408. [Google Scholar] [CrossRef]
- Häntzschel, K.; Weber, G. Blockage of mitosis in maize root tips using colchicine-alternatives. Protoplasma 2010, 241, 99–104. [Google Scholar] [CrossRef] [PubMed]
- Bruno, L.; Talarico, E.; Cabeiras-Freijanes, L.; Madeo, M.L.; Muto, A.; Minervino, M.; Araniti, F. Coumarin interferes with polar auxin transport altering microtubule cortical array organization in Arabidopsis thaliana (L.) Heynh. root apical meristem. Int. J. Mol. Sci. 2021, 22, 7305. [Google Scholar] [CrossRef]
- Li, J.; Xu, H.-H.; Liu, W.-C.; Zhang, X.-W.; Lu, Y.-T. Ethylene inhibits root elongation during alkaline stress through AUXIN1 and associated changes in auxin accumulation. Plant Physiol. 2015, 168, 1777–1791. [Google Scholar] [CrossRef] [Green Version]
- Soltys, D.; Rudzińska-Langwald, A.; Gniazdowska, A.; Wiśniewska, A.; Bogatek, R. Inhibition of tomato (Solanum lycopersicum L.) root growth by cyanamide is due to altered cell division, phytohormone balance and expansin gene expression. Planta 2012, 236, 1629–1638. [Google Scholar] [CrossRef] [Green Version]
- Steenackers, W.; Klíma, P.; Quareshy, M.; Cesarino, I.; Kumpf, R.P.; Corneillie, S.; Araújo, P.; Viaene, T.; Goeminne, G.; Nowack, M.K. cis-Cinnamic acid is a novel, natural auxin efflux inhibitor that promotes lateral root formation. Plant Physiol. 2017, 173, 552–565. [Google Scholar] [CrossRef] [Green Version]
- Oono, Y.; Ooura, C.; Rahman, A.; Aspuria, E.T.; Hayashi, K.-i.; Tanaka, A.; Uchimiya, H. p-Chlorophenoxyisobutyric acid impairs auxin response in Arabidopsis root. Plant Physiol. 2003, 133, 1135–1147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Rybel, B.; Audenaert, D.; Xuan, W.; Overvoorde, P.; Strader, L.C.; Kepinski, S.; Hoye, R.; Brisbois, R.; Parizot, B.; Vanneste, S. A role for the root cap in root branching revealed by the non-auxin probe naxillin. Nat. Chem. Biol. 2012, 8, 798–805. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blilou, I.; Xu, J.; Wildwater, M.; Willemsen, V.; Paponov, I.; Friml, J.; Heidstra, R.; Aida, M.; Palme, K.; Scheres, B. The PIN auxin efflux facilitator network controls growth and patterning in Arabidopsis roots. Nature 2005, 433, 39–44. [Google Scholar] [CrossRef]
- Swarup, R.; Kramer, E.M.; Perry, P.; Knox, K.; Leyser, H.O.; Haseloff, J.; Beemster, G.T.; Bhalerao, R.; Bennett, M.J. Root gravitropism requires lateral root cap and epidermal cells for transport and response to a mobile auxin signal. Nat. Cell Biol. 2005, 7, 1057–1065. [Google Scholar] [CrossRef]
- Křeček, P.; Skůpa, P.; Libus, J.; Naramoto, S.; Tejos, R.; Friml, J.; Zažímalová, E. The PIN-FORMED (PIN) protein family of auxin transporters. Genome Biol. 2009, 10, 249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muday, G.K.; Murphy, A.S. An emerging model of auxin transport regulation. Plant Cell 2002, 14, 293–299. [Google Scholar] [CrossRef] [PubMed]
- Araniti, F.; Bruno, L.; Sunseri, F.; Pacenza, M.; Forgione, I.; Bitonti, M.B.; Abenavoli, M.R. The allelochemical farnesene affects Arabidopsis thaliana root meristem altering auxin distribution. Plant Physiol. Biochem. 2017, 121, 14–20. [Google Scholar] [CrossRef]
- Sun, P.; Tian, Q.-Y.; Chen, J.; Zhang, W.-H. Aluminium-induced inhibition of root elongation in Arabidopsis is mediated by ethylene and auxin. J. Exp. Bot. 2010, 61, 347–356. [Google Scholar] [CrossRef] [Green Version]
- Michniewicz, M.; Zago, M.K.; Abas, L.; Weijers, D.; Schweighofer, A.; Meskiene, I.; Heisler, M.G.; Ohno, C.; Zhang, J.; Huang, F. Antagonistic regulation of PIN phosphorylation by PP2A and PINOID directs auxin flux. Cell 2007, 130, 1044–1056. [Google Scholar] [CrossRef] [Green Version]
- Dubrovsky, J.G.; Sauer, M.; Napsucialy-Mendivil, S.; Ivanchenko, M.G.; Friml, J.; Shishkova, S.; Celenza, J.; Benková, E. Auxin acts as a local morphogenetic trigger to specify lateral root founder cells. Proc. Natl. Acad. Sci. USA 2008, 105, 8790–8794. [Google Scholar] [CrossRef] [Green Version]
- Alarcón, M.; Salguero, J.; Lloret, P.G. Auxin modulated initiation of lateral roots is linked to pericycle cell length in maize. Front. Plant Sci. 2019, 10, 11. [Google Scholar] [CrossRef] [Green Version]
- Casimiro, I.; Beeckman, T.; Graham, N.; Bhalerao, R.; Zhang, H.; Casero, P.; Sandberg, G.; Bennett, M.J. Dissecting Arabidopsis lateral root development. Trends Plant Sci. 2003, 8, 165–171. [Google Scholar] [CrossRef]
- Laskowski, M.J.; Williams, M.E.; Nusbaum, H.C.; Sussex, I.M. Formation of lateral root meristems is a two-stage process. Development 1995, 121, 3303–3310. [Google Scholar] [CrossRef] [PubMed]
- Potters, G.; Pasternak, T.P.; Guisez, Y.; Jansen, M.A. Different stresses, similar morphogenic responses: Integrating a plethora of pathways. Plant Cell Environ. 2009, 32, 158–169. [Google Scholar] [CrossRef] [PubMed]
- Gniazdowska, A.; Krasuska, U.; Andrzejczak, O.; Soltys, D. Allelopathic compounds as oxidative stress agents: YES or NO. In Reactive Oxygen and Nitrogen Species Signaling and Communication in Plants; Springer: Berlin/Heidelberg, Germany, 2015; pp. 155–176. [Google Scholar]
- Rudrappa, T.; Bonsall, J.; Gallagher, J.L.; Seliskar, D.M.; Bais, H.P. Root-secreted allelochemical in the noxious weed Phragmites australis deploys a reactive oxygen species response and microtubule assembly disruption to execute rhizotoxicity. J. Chem. Ecol. 2007, 33, 1898–1918. [Google Scholar] [CrossRef] [PubMed]
- Bais, H.P.; Vepachedu, R.; Gilroy, S.; Callaway, R.M.; Vivanco, J.M. Allelopathy and exotic plant invasion: From molecules and genes to species interactions. Science 2003, 301, 1377–1380. [Google Scholar] [CrossRef] [PubMed]
- Rawlinson, C.; Kamphuis, L.G.; Gummer, J.P.; Singh, K.B.; Trengove, R.D. A rapid method for profiling of volatile and semi-volatile phytohormones using methyl chloroformate derivatisation and GC–MS. Metabolomics 2015, 11, 1922–1933. [Google Scholar] [CrossRef] [PubMed] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Molecules | Concentrations μM |
|---|---|
| P | 50 |
| TIBA | 15 |
| NPA | 5 |
| PCIB | 15 |
| 2,4-D | 0.1 |
| IAA | 0.1 |
| NAA | 0.1 |
| P + TIBA | 50 + 15 |
| P + NPA | 50 + 5 |
| P + PCIB | 50 + 15 |
| P + 2,4-D | 50 + 0.1 |
| P + IAA | 50 + 0.1 |
| P + NAA | 50 + 0.1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Santos Wagner, A.L.; Araniti, F.; Bruno, L.; Ishii-Iwamoto, E.L.; Abenavoli, M.R. The Steroid Saponin Protodioscin Modulates Arabidopsis thaliana Root Morphology Altering Auxin Homeostasis, Transport and Distribution. Plants 2021, 10, 1600. https://doi.org/10.3390/plants10081600
Santos Wagner AL, Araniti F, Bruno L, Ishii-Iwamoto EL, Abenavoli MR. The Steroid Saponin Protodioscin Modulates Arabidopsis thaliana Root Morphology Altering Auxin Homeostasis, Transport and Distribution. Plants. 2021; 10(8):1600. https://doi.org/10.3390/plants10081600
Chicago/Turabian StyleSantos Wagner, Ana Luiza, Fabrizio Araniti, Leonardo Bruno, Emy Luiza Ishii-Iwamoto, and Maria Rosa Abenavoli. 2021. "The Steroid Saponin Protodioscin Modulates Arabidopsis thaliana Root Morphology Altering Auxin Homeostasis, Transport and Distribution" Plants 10, no. 8: 1600. https://doi.org/10.3390/plants10081600