
Vegetative Reproduction Is More Advantageous Than Sexual Reproduction in a Canopy-Forming Clonal Macroalga under Ocean Warming Accompanied by Oligotrophication and Intensive Herbivory

 ,
,
Abstract
:1. Introduction
2. Results
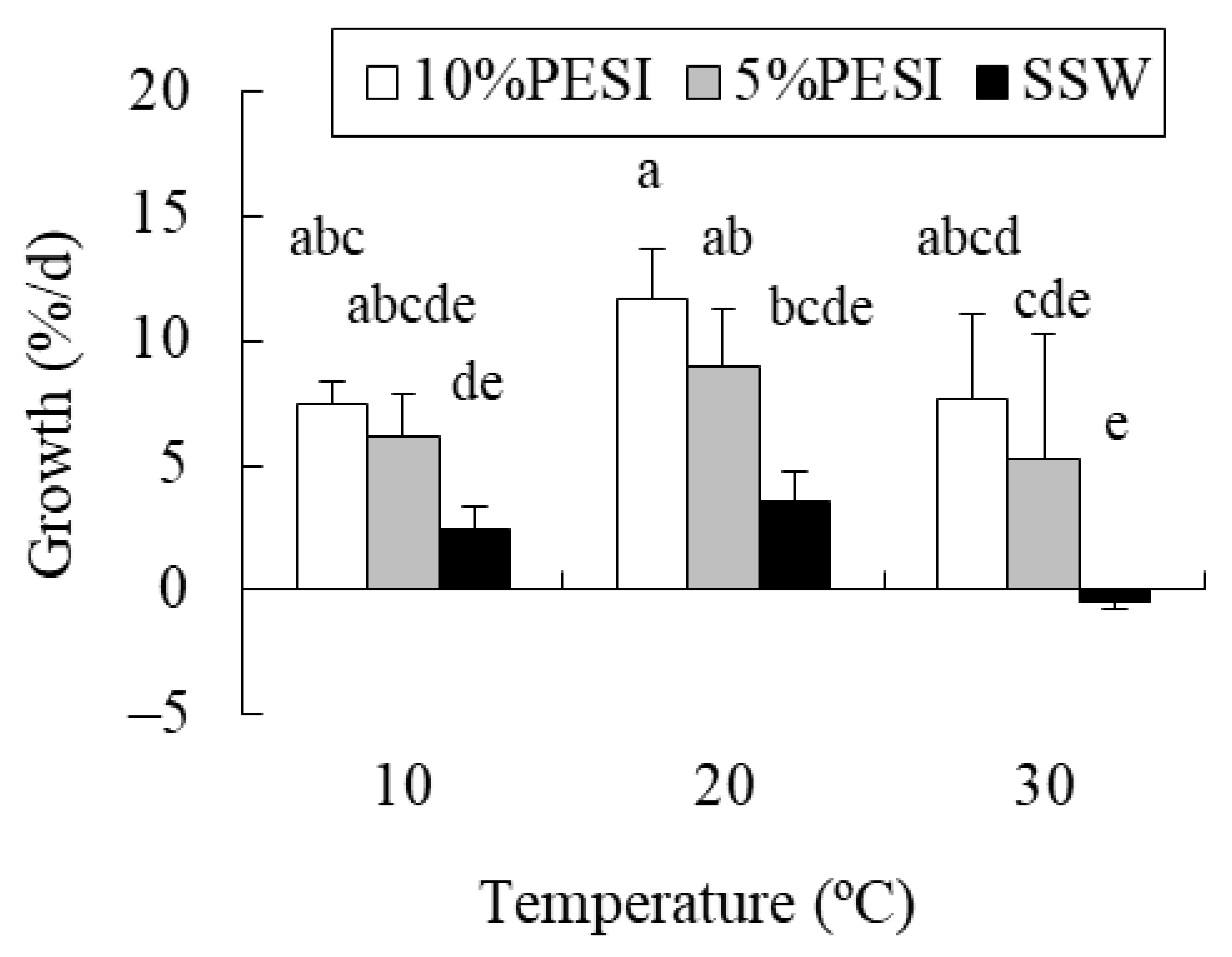
2.1. Experiment 1: Combined Effects of Temperature and Nutrients on Propagules
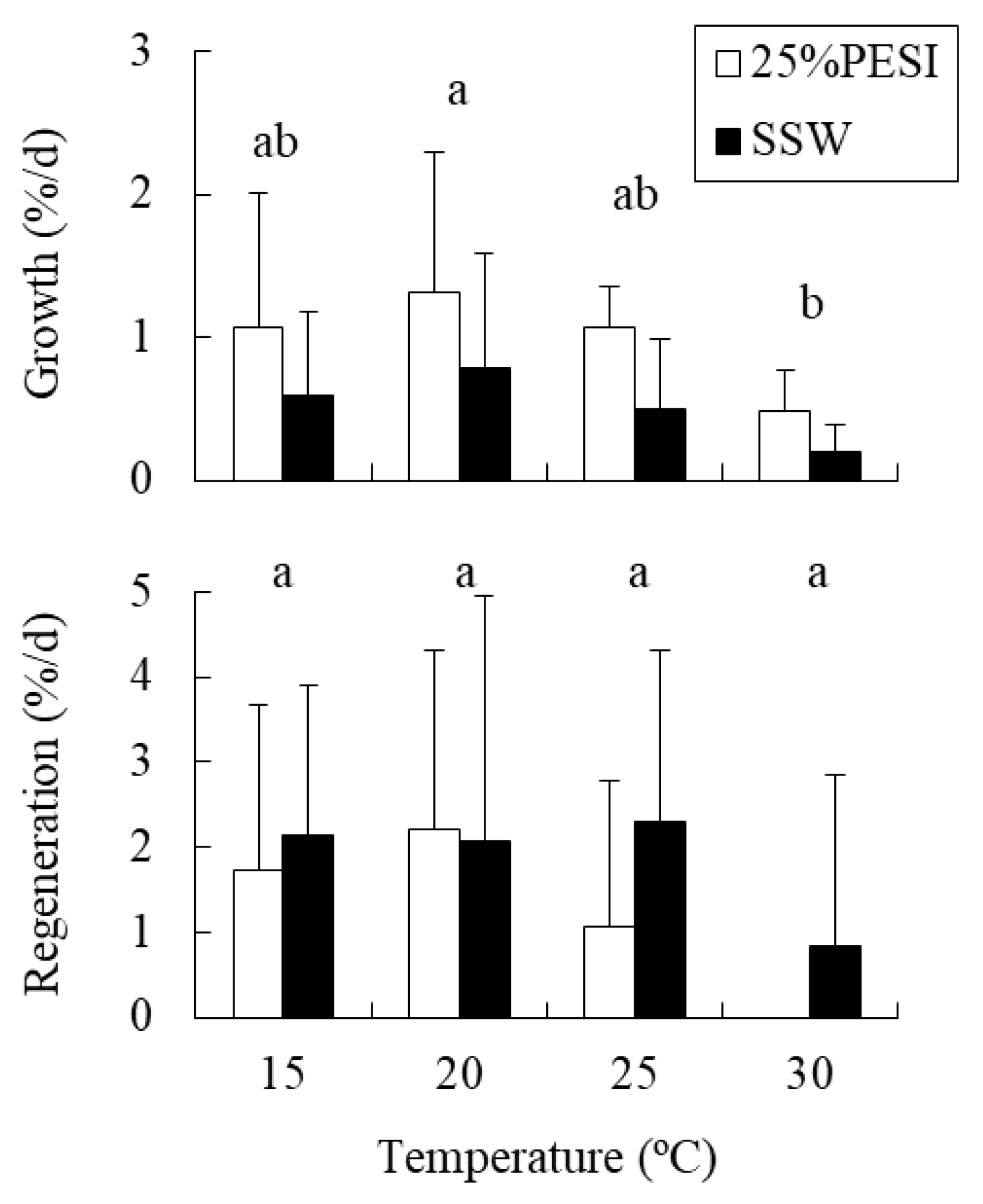
2.2. Experiment 2: Combined Effects of Temperature and Nutrients on Holdfasts
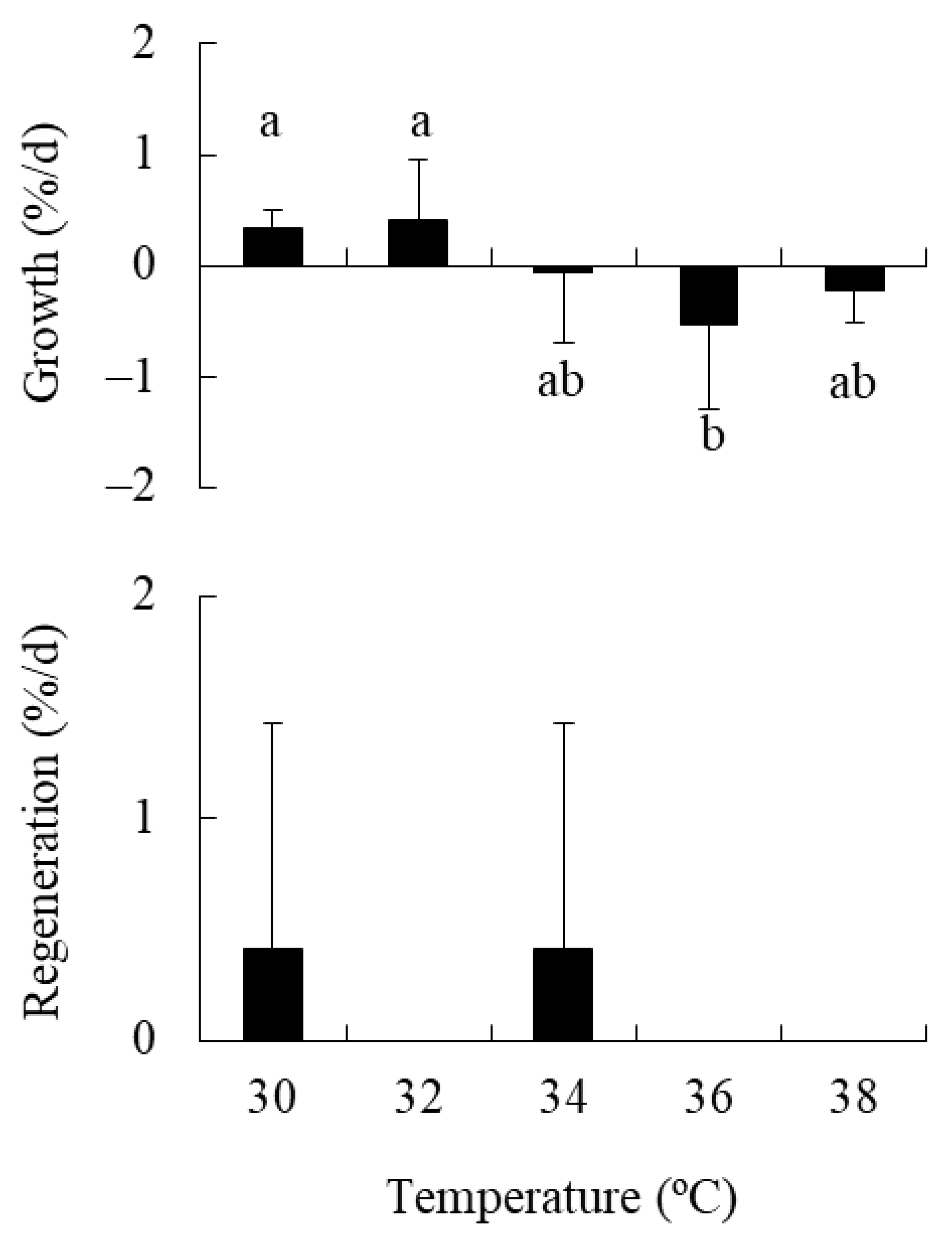
2.3. Experiment 3: Effect of High Temperature on Holdfasts
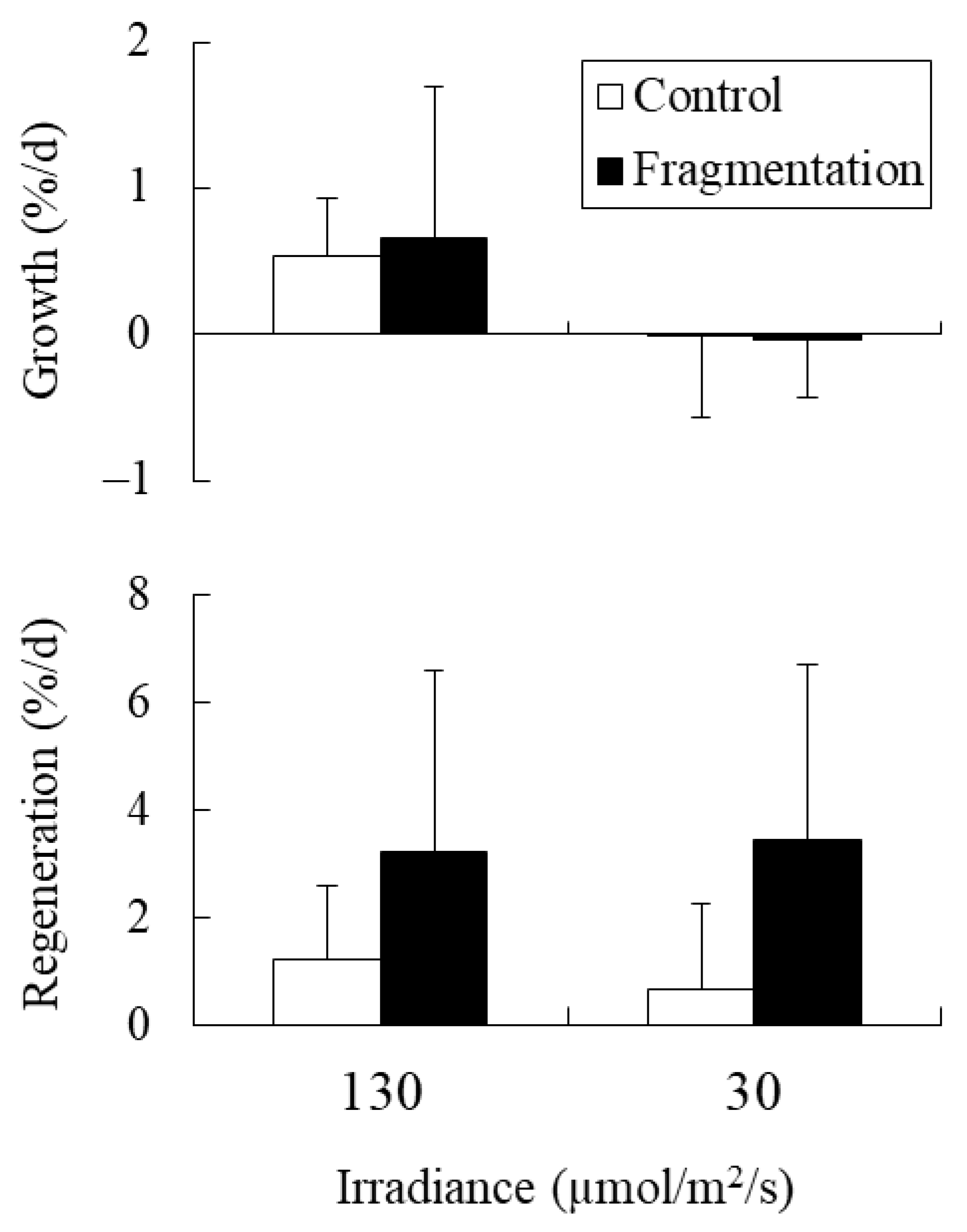
2.4. Experiment 4: Combined Effects of Fragmentation and Irradiance on Holdfasts
3. Discussion
4. Materials and Methods
4.1. Experiment 1: Combined Effects of Temperature and Nutrients on Propagules
4.2. Experiment 2: Combined Effects of Temperature and Nutrients on Holdfasts
4.3. Experiment 3: Effect of High Temperature on Holdfasts
4.4. Experiment 4: Combined Effects of Fragmentation and Irradiance on Holdfasts
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Schmidt, A.; Schmid, M.W.; Grossniklaus, U. Plant germline formation: Common concepts and developmental flexibility in sexual and asexual reproduction. Development 2015, 142, 229–241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roiloa, S.R.; Retuerto, R. Effects of fragmentation and seawater submergence on photochemical efficiency and growth in the clonal invader Carpobrotus edulis. Flora 2016, 225, 45–51. [Google Scholar] [CrossRef] [Green Version]
- Zhou, J.; Li, H.L.; Alpert, P.; Zhang, M.X.; Yu, F.H. Fragmentation of the invasive, clonal plant Alternanthera philoxeroides decreases its growth but not its competitive effect. Flora 2017, 228, 17–23. [Google Scholar] [CrossRef]
- Wang, J.; Xu, T.; Wang, Y.; Li, G.; Abdullah, I.; Zhong, Z.; Liu, J.; Zhu, W.; Wang, L.; Wang, D.; et al. A meta-analysis of effects of physiological integration in clonal plants under homogeneous vs. heterogeneous environments. Funct. Ecol. 2021, 35, 578–589. [Google Scholar] [CrossRef]
- Dayton, P.K. Toward understanding of community resilience and the potential effects of enrichment to the benthos at McMurdo Sound, Antarctica. In Proceedings of the Colloquium on Conservation Problems in Antarctica; Parker, B.C., Ed.; Allen Press: Lawrence, KS, USA, 1972; pp. 81–95. [Google Scholar]
- Coleman, M.A.; Wernberg, T. Forgotten underwater forests: The key role of fucoids on Australian temperate reefs. Ecol. Evol. 2017, 7, 8406–8418. [Google Scholar] [CrossRef] [Green Version]
- Eger, A.M.; Marzinelli, E.; Gribben, P.; Johnson, C.R.; Layton, C.; Steinberg, P.D.; Wood, G.; Silliman, B.R.; Vergés, A. Playing to the positives: Using synergies to enhance kelp forest restoration. Front. Mar. Sci. 2020, 7, 544. [Google Scholar] [CrossRef]
- Filbee-Dexter, K.; Wernberg, T. Substantial blue carbon in overlooked Australian kelp forests. Sci. Rep. 2020, 10, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, K.; Yoshida, G.; Hori, M.; Umezawa, Y.; Moki, H.; Kuwae, T. Macroalgal metabolism and lateral carbon flows can create significant carbon sinks. Biogeosciences 2020, 17, 2425–2440. [Google Scholar] [CrossRef]
- Smale, D.A. Impacts of ocean warming on kelp forest ecosystems. New Phytol. 2020, 225, 1447–1454. [Google Scholar] [CrossRef] [Green Version]
- Gerard, V.A. The role of nitrogen nutrition in high-temperature tolerance of the kelp, Laminaria saccharina (Chromophyta). J. Phycol. 1997, 33, 800–810. [Google Scholar] [CrossRef]
- Gao, X.; Endo, H.; Taniguchi, K.; Agatsuma, Y. Combined effects of seawater temperature and nutrient condition on growth and survival of juvenile sporophytes of the kelp Undaria pinnatifida (Laminariales; Phaeophyta) cultivated in northern Honshu, Japan. J. Appl. Phycol. 2013, 25, 269–275. [Google Scholar] [CrossRef]
- Gao, X.; Endo, H.; Nagaki, M.; Agatsuma, Y. Growth and survival of juvenile sporophytes of the kelp Ecklonia cava in response to different nitrogen and temperature regimes. Fish. Sci. 2016, 82, 623–629. [Google Scholar] [CrossRef]
- Gao, X.; Endo, H.; Nagaki, M.; Agatsuma, Y. Interactive effects of nutrient availability and temperature on growth and survival of different size classes of Saccharina japonica (Laminariales, Phaeophyceae). Phycologia 2017, 56, 253–260. [Google Scholar] [CrossRef]
- Vergés, A.; Steinberg, P.D.; Hay, M.E.; Poore, A.G.; Campbell, A.H.; Ballesteros, E.; Heck, K.L.; Booth, D.J.; Coleman, M.A.; Feary, D.A.; et al. The tropicalization of temperate marine ecosystems: Climate-mediated changes in herbivory and community phase shifts. Proc. R. Soc. B Biol. Sci. 2014, 281, 20140846. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, Y.W.; Ishida, H.; Nakano, T.; Nagai, N. Spatiotemporal decreases of nutrients and chlorophyll-a in the surface mixed layer of the western North Pacific from 1971 to 2000. J. Oceanogr. 2005, 61, 1011–1016. [Google Scholar] [CrossRef]
- Kumagai, N.H.; Molinos, J.G.; Yamano, H.; Takao, S.; Fujii, M.; Yamanaka, Y. Ocean currents and herbivory drive macroalgae-to-coral community shift under climate warming. Proc. Natl. Acad. Sci. USA 2018, 115, 8990–8995. [Google Scholar] [CrossRef] [Green Version]
- Endo, H.; Nishigaki, T.; Yamamoto, K.; Takeno, K. Age-and size-based morphological comparison between the brown alga Sargassum macrocarpum (Heterokonta; Fucales) from different depths at an exposed coast in northern Kyoto, Japan. J. Appl. Phycol. 2013, 25, 1815–1822. [Google Scholar] [CrossRef]
- Endo, H.; Nishigaki, T.; Yamamoto, K.; Takeno, K. Subtidal macroalgal succession and competition between the annual, Sargassum horneri, and the perennials, Sargassum patens and Sargassum piluliferum, on an artificial reef in Wakasa Bay, Japan. Fish. Sci. 2019, 85, 61–69. [Google Scholar] [CrossRef]
- Yoshida, G.; Shimabukuro, H. Seasonal population dynamics of Sargassum fusiforme (Fucales, Phaeophyta), Suo-Oshima Is., Seto Inland Sea, Japan—development processes of a stand characterized by high density and productivity. J. Appl. Phycol. 2017, 29, 639–648. [Google Scholar] [CrossRef] [Green Version]
- Low, J.K.; Fong, J.; Todd, P.A.; Chou, L.M.; Bauman, A.G. Seasonal variation of Sargassum ilicifolium (Phaeophyceae) growth on equatorial coral reefs. J. Phycol. 2019, 55, 289–296. [Google Scholar] [CrossRef]
- Ito, R.; Terawaki, T.; Satuito, C.G.; Kitamura, H. Storage, cutting and culture conditions of filamentous roots of Hiziki Sargassum fusiforme. Aquacult. Sci. 2009, 57, 579–585, (In Japanese with English Abstract). [Google Scholar]
- Yatsuya, K.; Kiyomoto, S.; Yoshida, G.; Yoshimura, T. Regeneration of the holdfast in 13 species of the genus Sargassum grown along the western coast of Kyushu, south-western Japan. Jpn. J. Phycol. 2012, 60, 41–45, (In Japanese with English Abstract). [Google Scholar]
- Loffler, Z.; Graba-Landry, A.; Kidgell, J.T.; McClure, E.C.; Pratchett, M.S.; Hoey, A.S. Holdfasts of Sargassum swartzii are resistant to herbivory and resilient to damage. Coral Reefs 2018, 37, 1075–1084. [Google Scholar] [CrossRef]
- Yamaguchi, A.; Furumitsu, K.; Yagishita, N.; Kume, G. Biology of herbivorous fish in the coastal areas of western Japan. In Coastal Environmental and Ecosystem Issues of the East China Sea; Ishimatsu, A., Lie, H.-J., Eds.; TERRAPUB and Nagasaki University: Nagasaki, Japan, 2010; pp. 181–2190. [Google Scholar]
- Shimabukuro, H.; Terada, R.; Sotobayashi, J.; Nishihara, G.N.; Noro, T. Phenology of Sargassum duplicatum (Fucales, Phaeophyceae) from the southern coast of Satsuma Peninsula, Kagoshima, Japan. Nippon Suisan Gakkaishi 2007, 73, 454–460, (In Japanese with English Abstract). [Google Scholar] [CrossRef]
- Tsuchiya, Y.; Sakaguchi, Y.; Terada, R. Phenology and enrivonmental characteristics of four Sargassum species (Fucales): S. piluliferum, S. patens, S. crispifolium, and S. alternato-pinnatum from Sakurajima, Kagoshima Bay, southern Japan. Jpn. J. Phycol. 2011, 59, 1–8, (In Japanese with English Abstract). [Google Scholar]
- Nakashima, H.; Tanaka, T.; Yoshimitsu, S.; Terada, R. Phenology of three species of Sargassum (Fucales) and the long-term change of seaweed community structure from Kasasa, Kagoshima Prefecture, Japan. Jpn. J. Phycol. 2013, 61, 97–105, (In Japanese with English Abstract). [Google Scholar]
- Baba, M. Effects of temperature and irradiance on germling growth in eight Sargassaceous species. Rep. Mar. Ecol. Res. Inst. 2007, 10, 9–20. [Google Scholar]
- Graba-Landry, A.C.; Loffler, Z.; McClure, E.C.; Pratchett, M.S.; Hoey, A.S. Impaired growth and survival of tropical macroalgae (Sargassum spp.) at elevated temperatures. Coral Reefs 2020, 39, 475–486. [Google Scholar] [CrossRef]
- Hay, K.B.; Millers, K.A.; Poore, A.G.; Lovelock, C.E. The use of near infrared reflectance spectrometry for characterization of brown algal tissue. J. Phycol. 2010, 46, 937–946. [Google Scholar] [CrossRef]
- Endo, H.; Suehiro, K.; Kinoshita, J.; Gao, X.; Agatsuma, Y. Combined effects of temperature and nutrient availability on growth and phlorotannin concentration of the brown alga Sargassum patens (Fucales; Phaeophyceae). Am. J. Plant Sci. 2013, 4, 14–20. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Liu, Y.; Liu, Y.; Wang, Q.; Gao, X.; Gong, Q. Effects of temperature and salinity on the growth and biochemical composition of the brown alga Sargassum fusiforme (Fucales, Phaeophyceae). J. Appl. Phycol. 2019, 31, 3061–3068. [Google Scholar] [CrossRef]
- Loffler, Z.; Hoey, A.S. Microtopographic refuges enhance recruitment and survival but inhibit growth of propagules of the tropical macroalga Sargassum swartzii. Mar. Ecol. Prog. Ser. 2019, 627, 61–70. [Google Scholar] [CrossRef]
- Yoshida, T. Marine Algae of Japan; Uchida Rokakuho: Tokyo, Japan, 1998. (In Japanese) [Google Scholar]
- Hu, Z.M.; Li, J.J.; Sun, Z.M.; Gao, X.; Yao, J.T.; Choi, H.G.; Endo, H.; Duan, D.L. Hidden diversity and phylogeographic history provide conservation insights for the edible seaweed Sargassum fusiforme in the Northwest Pacific. Evol. Appl. 2017, 10, 366–378. [Google Scholar] [CrossRef]
- Liu, L.; Lin, L. Effect of heat stress on Sargassum fusiforme leaf metabolome. J. Plant Biol. 2020, 63, 229–241. [Google Scholar] [CrossRef] [Green Version]
- Gao, X.; Endo, H.; Taniguchi, K.; Agatsuma, Y. Genetic differentiation of high-temperature tolerance in the kelp Undaria pinnatifida sporophytes from geographically separated populations along the Pacific coast of Japan. J. Appl. Phycol. 2013, 25, 567–574. [Google Scholar] [CrossRef]
- Endo, H.; Inomata, E.; Gao, X.; Kinoshita, J.; Sato, Y.; Agatsuma, Y. Heat stress promotes nitrogen accumulation in meristems via apical blade erosion in a brown macroalga with intercalary growth. Front. Mar. Sci. 2020, 7, 575721. [Google Scholar] [CrossRef]
- IPCC. Climate change 2013: The physical science basis. In Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change (IPCC); Cambridge University Press: Cambridge, MA, USA, 2013. [Google Scholar]
- Pang, S.J.; Chen, L.T.; Zhuang, D.G.; Fei, X.G.; Sun, J.Z. Cultivation of the brown alga Hizikia fusiformis (Harvey) Okamura: Enhanced seedling production in tumbled culture. Aquaculture 2005, 245, 321–329. [Google Scholar] [CrossRef]
- Pang, S.J.; Zhang, Z.H.; Zhao, H.J.; Sun, J.Z. Cultivation of the brown alga Hizikia fusiformis (Harvey) Okamura: Stress resistance of artificially raised young seedlings revealed by chlorophyll fluorescence measurement. J. Appl. Phycol. 2007, 19, 557–565. [Google Scholar] [CrossRef]
- Pang, S.J.; Shan, T.F.; Zhang, Z.H.; Sun, J.Z. Cultivation of the intertidal brown alga Hizikia fusiformis (Harvey) Okamura: Mass production of zygote-derived seedlings under commercial cultivation conditions, a case study experience. Aquacult. Res. 2008, 39, 1408–1415. [Google Scholar] [CrossRef]
- Tatewaki, M. Formation of a crustaceous sporophyte with unilocular sporangia in Scytosiphon lomentaria. Phycologia 1966, 6, 62–66. [Google Scholar] [CrossRef]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Source | df | MS | F | p | |
|---|---|---|---|---|---|
| Temperature (T) | 2 | 1.385 | 32.408 | <0.001 | * |
| Nutrient (N) | 2 | 2.420 | 56.647 | <0.001 | * |
| T × N | 4 | 0.351 | 8.219 | <0.001 | * |
| Source | df | MS | F | p | |
|---|---|---|---|---|---|
| Growth rate | |||||
| Temperature (T) | 3 | 0.915 | 3.887 | 0.016 | * |
| Nutrient (N) | 1 | 1.129 | 4.797 | 0.034 | * |
| T × N | 3 | 0.052 | 0.222 | 0.881 | |
| Regeneration rate | |||||
| Temperature (T) | 3 | 0.254 | 2.478 | 0.075 | |
| Nutrient (N) | 1 | 0.100 | 0.973 | 0.330 | |
| T × N | 3 | 0.045 | 0.441 | 0.725 |
| Source | df | MS | F | p | |
|---|---|---|---|---|---|
| Growth rate | |||||
| Fragmentation (F) | 1 | 0.013 | 0.029 | 0.866 | |
| Irradiance (I) | 1 | 2.260 | 5.228 | 0.033 | * |
| F × I | 1 | 0.038 | 0.089 | 0.769 | |
| Regeneration rate | |||||
| Fragmentation (F) | 1 | 33.982 | 5.156 | 0.034 | * |
| Irradiance (I) | 1 | 0.187 | 0.028 | 0.868 | |
| F × I | 1 | 0.995 | 0.151 | 0.702 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Endo, H.; Sugie, T.; Yonemori, Y.; Nishikido, Y.; Moriyama, H.; Ito, R.; Okunishi, S. Vegetative Reproduction Is More Advantageous Than Sexual Reproduction in a Canopy-Forming Clonal Macroalga under Ocean Warming Accompanied by Oligotrophication and Intensive Herbivory. Plants 2021, 10, 1522. https://doi.org/10.3390/plants10081522
Endo H, Sugie T, Yonemori Y, Nishikido Y, Moriyama H, Ito R, Okunishi S. Vegetative Reproduction Is More Advantageous Than Sexual Reproduction in a Canopy-Forming Clonal Macroalga under Ocean Warming Accompanied by Oligotrophication and Intensive Herbivory. Plants. 2021; 10(8):1522. https://doi.org/10.3390/plants10081522
Chicago/Turabian StyleEndo, Hikaru, Toru Sugie, Yukiko Yonemori, Yuki Nishikido, Hikari Moriyama, Ryusei Ito, and Suguru Okunishi. 2021. "Vegetative Reproduction Is More Advantageous Than Sexual Reproduction in a Canopy-Forming Clonal Macroalga under Ocean Warming Accompanied by Oligotrophication and Intensive Herbivory" Plants 10, no. 8: 1522. https://doi.org/10.3390/plants10081522