
The In Vitro α-Glucosidase Inhibition Activity of Various Solvent Fractions of Tamarix dioica and 1H-NMR Based Metabolite Identification and Molecular Docking Analysis

 , ,
, ,  , and
, and
Abstract
:1. Introduction
2. Results
2.1. Total Phenolic and Flavonoid Contents
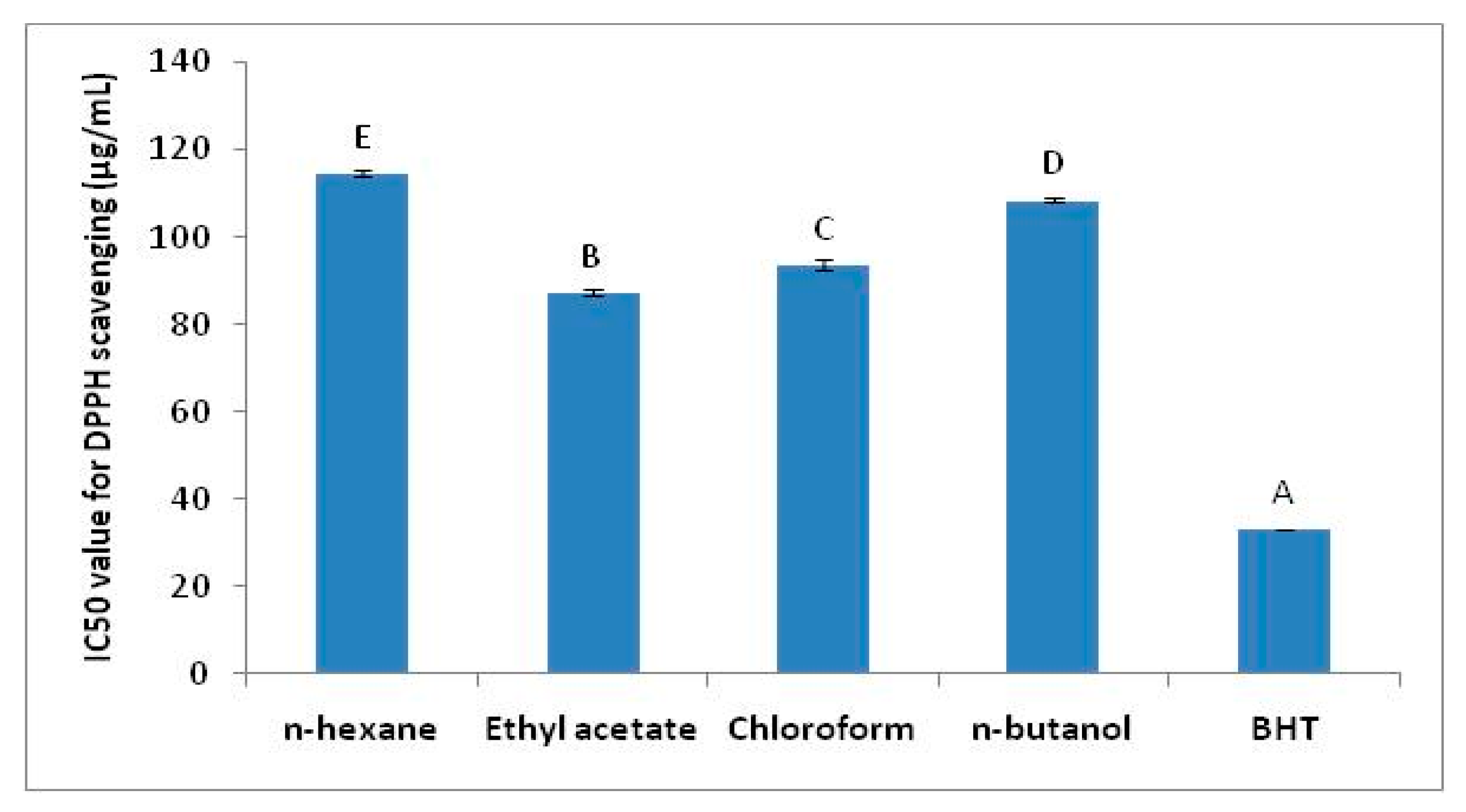
2.2. Antioxidant Activity
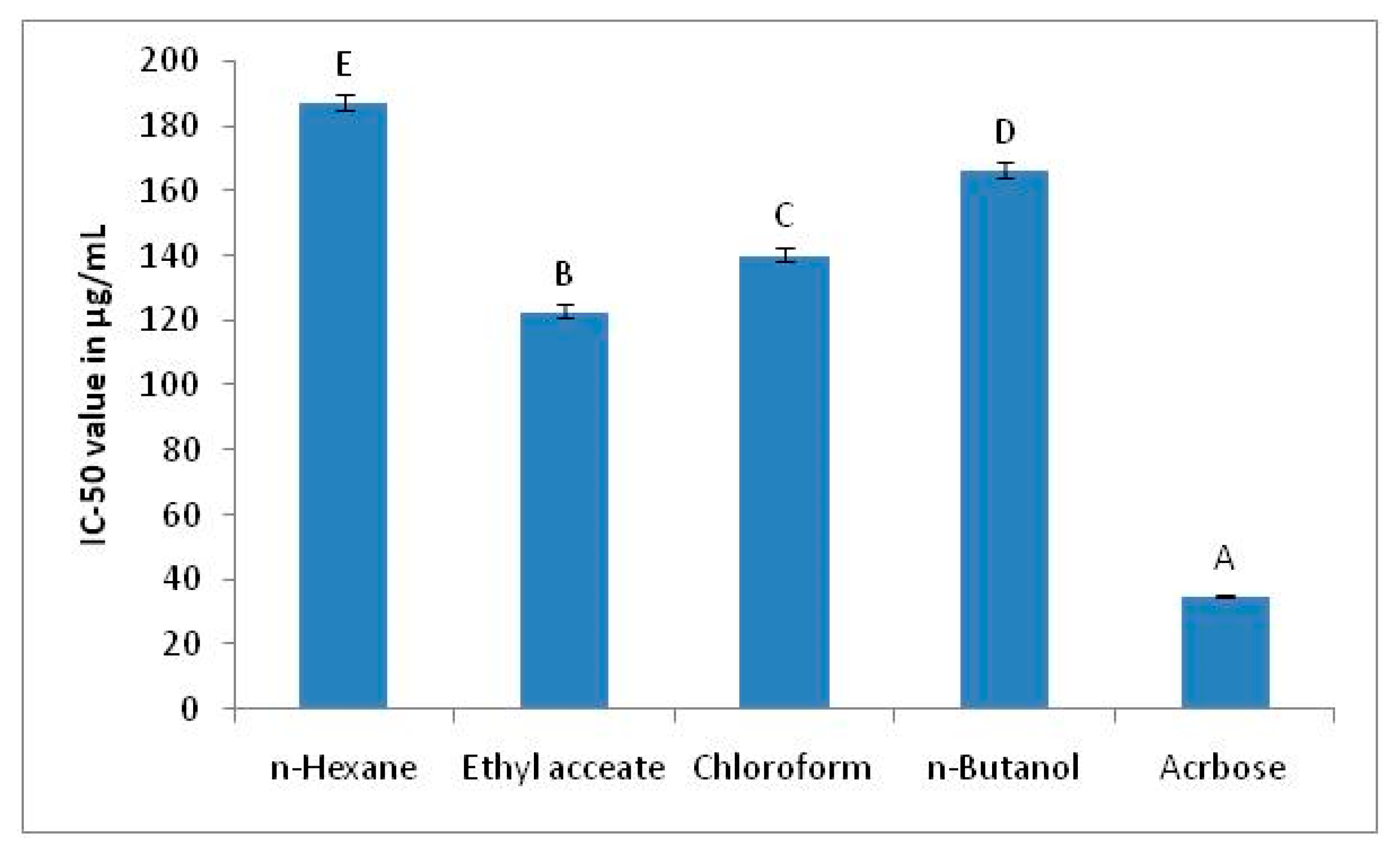
2.3. The α-Glucosidase Inhibition Assay
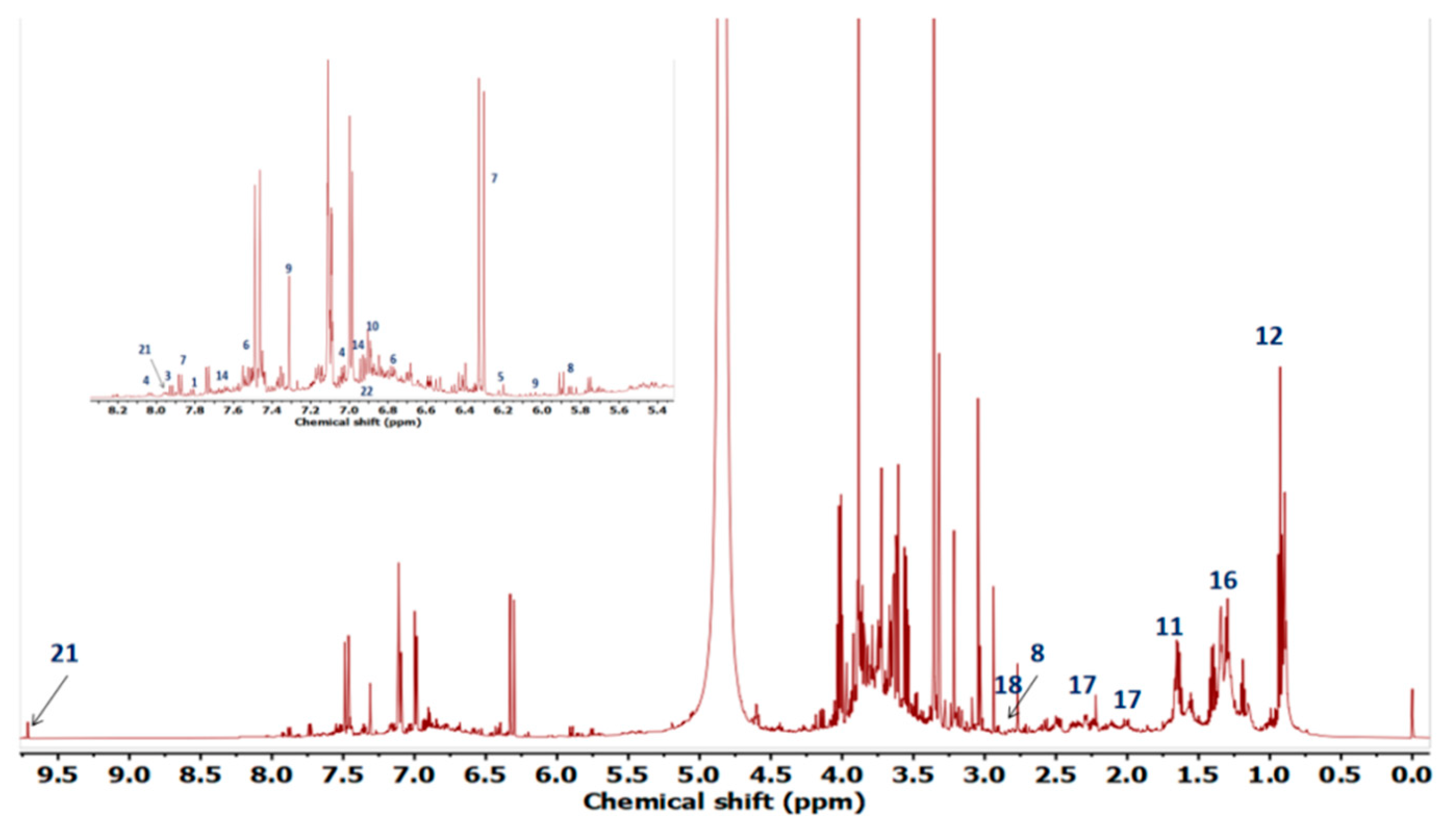
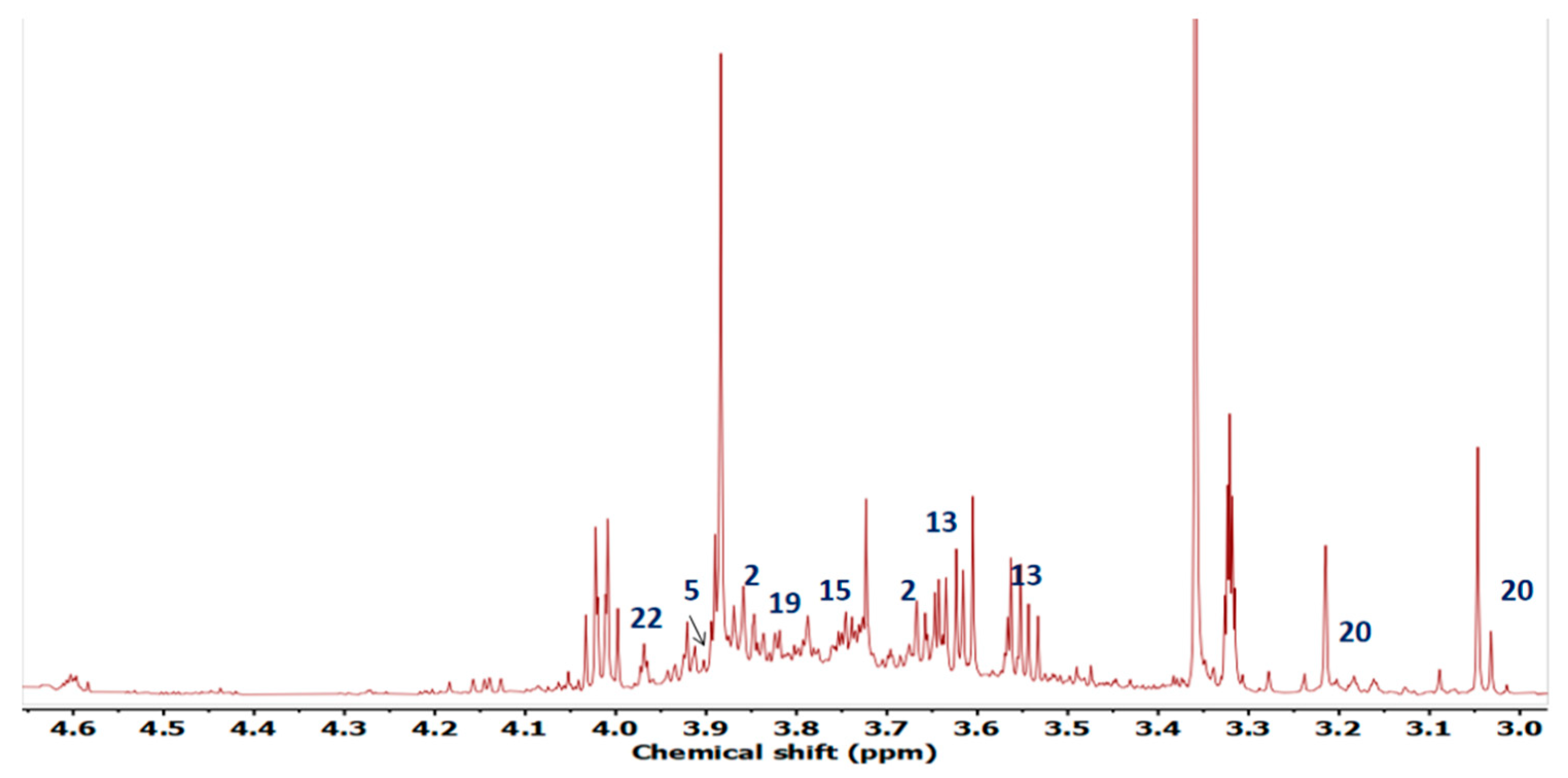
2.4. Metabolite Profiling
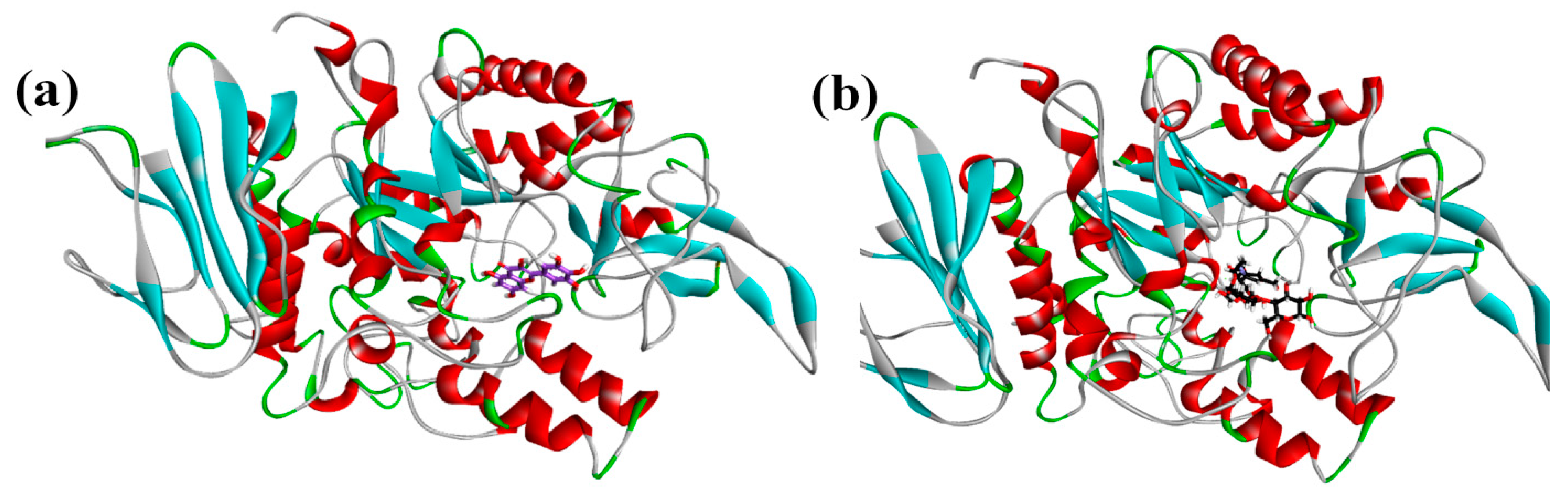
2.5. Molecular Docking Data
3. Discussion
4. Materials and Methods
4.1. Processing of Plant Material
4.2. Preparation of Extracts
4.3. DPPH Radical Based Antioxidant Activity
4.4. Total Phenolic Contents
4.5. Total Flavonoid Contents
4.6. The α-Glucosidase Inhibition Assay
4.7. Metabolite Profiling
4.8. Docking Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bahramsoltani, R.; Kalkhorani, M.; Zaidi, S.M.A.; Farzaei, M.H.; Rahimi, R. The genus Tamarix: Traditional uses, phytochemistry, and pharmacology. J. Ethnopharmacol. 2020, 246, 112245. [Google Scholar] [CrossRef] [PubMed]
- Bughio, S.H.; Samejo, M.Q.; Memon, S.; Bano, S.; Mughal, M.A.; Memon, A.A. Chemical composition of the essential oils from Tamarix dioica and determination of its antibacterial activity. Int. J. Food Prop. 2017, 20, S2660–S2667. [Google Scholar] [CrossRef] [Green Version]
- Khan, S.; Khan, G.M.; Mehsud, S.; Rahman, A.; Khan, F. Antifungal activity of Tamarix dioica—An in vitro study. Gomal J. Med. Sci. 2004, 2, 40–42. [Google Scholar]
- Samejo, M.Q.; Sumbul, A.; Shah, S.; Memon, S.B.; Chundrigar, S. Phytochemical screening of Tamarix dioica Roxb. ex Roch. J. Pharm. Res. 2013, 7, 181–183. [Google Scholar] [CrossRef]
- Imtiaz, S.M.; Aleem, A.; Saqib, F.; Ormenisan, A.N.; Elena Neculau, A.; Anastasiu, C.V. The potential involvement of an ATP-dependent potassium channel-opening mechanism in the smooth muscle relaxant properties of Tamarix dioica Roxb. Biomolecules 2019, 9, 722. [Google Scholar] [CrossRef] [Green Version]
- ReMukesh, R.; Namita, P. Medicinal plants with antidiabetic potential-A review. Am-Eurasian. J. Agric. Environ. Sci. 2017, 13, 81–94. [Google Scholar]
- Yamazaki, D.; Hitomi, H.; Nishiyama, A. Hypertension with diabetes mellitus complications. Hypertens. Res. 2018, 41, 147–156. [Google Scholar] [CrossRef] [PubMed]
- Deshmukh, C.D.; Jain, A. Diabetes mellitus: A review. Int. J. Pure App. Biosci. 2015, 3, 224–230. [Google Scholar]
- Saltiel, A.R.; Olefsky, J.M. Thiazolidinediones in the treatment of insulin resistance and type II diabetes. Diabetes 1996, 45, 1661–1669. [Google Scholar] [CrossRef]
- Folli, F.; Corradi, D.; Fanti, P.; Davalli, A.; Paez, A.; Giaccari, A.; Perego, C.; Muscogiuri, G. The role of oxidative stress in the pathogenesis of type 2 diabetes mellitus micro-and macrovascular complications: Avenues for a mechanistic-based therapeutic approach. Curr. Diabetes Rev. 2011, 7, 313–324. [Google Scholar] [CrossRef]
- Babizhayev, M.A.; Strokov, I.A.; Nosikov, V.V.; Savel’yeva, E.L.; Sitnikov, V.F.; Yegorov, Y.E.; Lankin, V.Z. The role of oxidative stress in diabetic neuropathy: Generation of free radical species in the glycation reaction and gene polymorphisms encoding antioxidant enzymes to genetic susceptibility to diabetic neuropathy in population of type I diabetic patients. Cell Biochem. Biophys. 2015, 71, 1425–1443. [Google Scholar] [CrossRef]
- Van de Laar, F.A.; Lucassen, P.L.; Akkermans, R.P.; Van de Lisdonk, E.H.; De Grauw, W.J. Alpha-glucosidase inhibitors for people with impaired glucose tolerance or impaired fasting blood glucose. Cochrane Database Syst. Rev. 2006, 18, CD005061. [Google Scholar] [CrossRef] [Green Version]
- William, J.; John, P.; Mumtaz, M.W.; Ch, A.R.; Adnan, A.; Mukhtar, H.; Sharif, S.; Raza, S.A.; Akhtar, M.T. Antioxidant activity, α-glucosidase inhibition and phytochemical profiling of Hyophorbelagenicaulis leaf extracts. PeerJ 2019, 7, e7022. [Google Scholar] [CrossRef] [Green Version]
- Arshad, M.; Chaudhary, A.R.; Mumtaz, M.W.; Raza, S.A.; Ahmad, M.; Mukhtar, H.; Bashir, R. Polyphenol fingerprinting and hypoglycemic attributes of optimized Cycas circinalis leaf extracts. J. Sci. Food Agric. 2021, 101, 1530–1537. [Google Scholar] [CrossRef] [PubMed]
- Raza, S.A.; Chaudhary, A.R.; Mumtaz, M.W.; Ghaffar, A.; Adnan, A.; Waheed, A. Antihyperglycemic effect of Conocarpus erectus leaf extract in alloxan-induced diabetic mice. Pak. J. Pharm. Sci. 2018, 31, 637–642. [Google Scholar] [PubMed]
- William, J.; John, P.; Mumtaz, M.W.; Ch, A.R.; Adnan, A.; Mukhtar, H.; Sharif, S.; Raza, S.A. Antioxidant activity, Hypoglycemic potential and metabolite profiling of Hyophorbe indica leaf extract. Pak. J. Pharm. Sci. 2018, 31, 2737–2742. [Google Scholar] [PubMed]
- Farooq, M.U.; Mumtaz, M.W.; Mukhtar, H.; Rashid, U.; Akhtar, M.T.; Raza, S.A.; Nadeem, M. UHPLC-QTOF-MS/MS based phytochemical characterization and anti-hyperglycemic prospective of hydro-ethanolic leaf extract of Butea monosperma. Sci. Rep. 2020, 10, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdul Rahman, H.; Saari, N.; Abas, F.; Ismail, A.; Mumtaz, M.W.; Abdul Hamid, A. Anti-obesity and antioxidant activities of selected medicinal plants and phytochemical profiling of bioactive compounds. Int. J. Food Prop. 2017, 20, 2616–2629. [Google Scholar] [CrossRef]
- Oh, J.; Yoon, D.H.; Han, J.G.; Choi, H.K.; Sung, G.H. 1H NMR based metabolite profiling for optimizing the ethanol extraction of Wolfiporiacocos. Saudi J. Biol. Sci. 2018, 25, 1128–1134. [Google Scholar] [CrossRef]
- Aabideen, Z.U.; Mumtaz, M.W.; Akhtar, M.T.; Mukhtar, H.; Raza, S.A.; Touqeer, T.; Saari, N. Anti-obesity attributes; UHPLC-QTOF-MS/MS-based metabolite profiling and molecular docking insights of Taraxacum officinale. Molecules 2020, 25, 4935. [Google Scholar] [CrossRef]
- Umesh, H.; Ramesh, K.; Devaraju, K. Molecular docking studies of phytochemicals against trehalose–6–phosphate phosphatases of pathogenic microbes. Beni-Suef Univ. J. Basic Appl. Sci. 2020, 9, 5. [Google Scholar] [CrossRef] [Green Version]
- Nadeem, M.; Mumtaz, M.W.; Danish, M.; Rashid, U.; Mukhtar, H.; Irfan, A. Antidiabetic functionality of Vitex negundo L. leaves based on UHPLC-QTOF-MS/MS based bioactives profiling and molecular docking insights. Ind. Crops Prod. 2020, 152, 112445. [Google Scholar] [CrossRef]
- Hajinezhad, M.R.; Jamshidian, A.; Shahri, S.H. Effect of hydroalcoholic extract of Tamarix dioica on liver histopathological changes in rats fed with high fat diet. Qom Univ. Med. Sci. J. 2019, 13, 1–8. [Google Scholar] [CrossRef]
- Babbar, N.; Oberoi, H.S.; Sandhu, S.K.; Bhargav, V.K. Influence of different solvents in extraction of phenolic compounds from vegetable residues and their evaluation as natural sources of antioxidants. J. Food Sci. Technol. 2014, 51, 2568–2575. [Google Scholar] [CrossRef] [Green Version]
- Pandey, K.B.; Rizvi, S.I. Plant polyphenols as dietary antioxidants in human health and disease. Oxidative Med. Cell. Longev. 2009, 2, 270–278. [Google Scholar] [CrossRef] [Green Version]
- Tungmunnithum, D.; Thongboonyou, A.; Pholboon, A.; Yangsabai, A. Flavonoids and other phenolic compounds from medicinal plants for pharmaceutical and medical aspects: An overview. Medicines 2018, 5, 93. [Google Scholar] [CrossRef] [PubMed]
- Villaño, D.; Fernández-Pachón, M.; Moyá, M.L.; Troncoso, A.; García-Parrilla, M. Radical scavenging ability of polyphenolic compounds towards DPPH free radical. Talanta 2007, 71, 230–235. [Google Scholar] [CrossRef]
- Raza, S.A.; Rashid, A.; William, J.; Arshed, S.F.; Arshad, M. Comparison of antioxidant activity of some medicinally important plants from Pakistan. Acta Sci. Pol. Technol. Aliment. 2013, 12, 403–410. [Google Scholar]
- Raza, S.A.; Chaudhary, A.R.; Mumtaz, M.W.; Adnan, A.; Mukhtar, H.; Akhtar, M.T. Metabolite profiling and antidiabetic attributes of ultrasonicated leaf extracts of Conocarpu slancifolius. Asian Pac. J. Trop. Biomed. 2020, 10, 353. [Google Scholar]
- Elya, B.; Basah, K.; Mun’im, A.; Yuliastuti, W.; Bangun, A.; Septiana, E.K. Screening of α-glucosidase inhibitory activity from some plants of Apocynaceae, Clusiaceae, Euphorbiaceae, and Rubiaceae. J. Biomed. Biotechnol. 2012, 2012, 281078. [Google Scholar] [CrossRef] [Green Version]
- Goboza, M.; Meyer, M.; Aboua, Y.G.; Oguntibeju, O.O. In vitro antidiabetic and antioxidant effects of different extracts of Catharanthus roseus and its indole alkaloid, vindoline. Molecules 2020, 25, 5546. [Google Scholar] [CrossRef]
- Mahfoudhi, A.; Grosso, C.; Gonçalves, R.F.; Khelifi, E.; Hammami, S.; Achour, S.; Trabelsi-Ayadi, M.; Valentão, P.; Andrade, P.B.; Mighri, Z. Evaluation of antioxidant, anticholinesterase, and antidiabetic potential of dry leaves and stems in Tamarix aphylla growing wild in Tunisia. Chem. Biodivers. 2016, 13, 1747–1755. [Google Scholar] [CrossRef] [PubMed]
- Nakanishi, H.; Onose, S.; Kitahara, E.; Chumchuen, S.; Takasaki, M.; Konishi, H.; Kanekatsu, R. Effect of environmental conditions on the α-glucosidase inhibitory activity of mulberry leaves. Biosci. Biotechnol. Biochem. 2011, 75, 2293–2296. [Google Scholar] [CrossRef] [PubMed]
- Raza, S.; Chaudhary, A.R.; Mumtaz, M.W.; Bashir, S. Antioxidant, iron chelating, lipase inhibition activities and metabolite’s prediction of hyrdoethanolic leaf extract of Conocarpus erectus. Food Res. 2019, 4, 482–487. [Google Scholar] [CrossRef]
- AL-Zuaidy, M.H.; Hamid, A.A.; Ismail, A.; Mohamed, S.; Abdul Razis, A.F.; Mumtaz, M.W.; Salleh, S.Z. Potent antidiabetic activity and metabolite profiling of Melicopelunu-ankenda leaves. J. Food Sci. 2016, 81, C1080–C1090. [Google Scholar] [CrossRef] [PubMed]
- Mediani, A.; Abas, F.; Khatib, A.; Tan, C.P.; Ismail, I.S.; Shaari, K.; Ismail, A.; Lajis, N. Phytochemical and biological features of Phyllanthus niruri and Phyllanthus urinaria harvested at different growth stages revealed by 1H NMR-based metabolomics. Ind. Crop Prod. 2015, 77, 602–613. [Google Scholar] [CrossRef]
- Sultanova, N.; Makhmoor, T.; Abilov, Z.; Parween, Z.; Omurkamzinova, V.; Choudhary, M.I. Antioxidant and antimicrobial activities of Tamarix ramosissima. J. Ethnopharmacol. 2001, 78, 201–205. [Google Scholar] [CrossRef]
- Arshad, M.; Mumtaz, M.W.; Chaudhary, A.R.; Rashid, U.; Ali, M.; Mukhtar, H.; Adnan, A.; Raza, S.A. Metabolite profiling of Cycas revoluta leaf extract and docking studies on alpha-glucosidase inhibitory molecular targets by phytochemicals. Pak. J. Pharm. Sci. 2019, 32, 871–874. [Google Scholar]
- Nadeem, M.; Mumtaz, M.W.; Danish, M.; Rashid, U.; Mukhtar, H.; Anwar, F.; Raza, S.A. Calotropis procera: UHPLC-QTOF-MS/MS based profiling of bioactives, antioxidant and anti-diabetic potential of leaf extracts and an insight into molecular docking. J. Food Meas. Charact. 2019, 13, 3206–3220. [Google Scholar] [CrossRef]
- Pereira, D.F.; Cazarolli, L.H.; Lavado, C.; Mengatto, V.; Figueiredo, M.S.R.B.; Guedes, A.; Pizzolatti, M.G.; Silva, F.R.M.B. Effects of flavonoids on α-glucosidase activity: Potential targets for glucose homeostasis. Nutrition 2011, 27, 1161–1167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rahman, N.; Muhammad, I.; Khan, H.; Aschner, M.; Filosa, R.; Daglia, M. Molecular docking of isolated alkaloids for possible α-glucosidase inhibition. Biomolecules 2019, 9, 544. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ivanović, M.; Makoter, K.; IslamčevićRazboršek, M. Comparative study of chemical composition and antioxidant activity of essential oils and crude extracts of four characteristic Zingiberaceae Herbs. Plants 2021, 10, 501. [Google Scholar] [CrossRef]
- Chew, K.; Khoo, M.; Ng, S.; Thoo, Y.; Aida, W.W.; Ho, C. Effect of ethanol concentration, extraction time and extraction temperature on the recovery of phenolic compounds and antioxidant capacity of Orthosiphon stamineus extracts. Int. Food Res. J. 2011, 18, 1427. [Google Scholar]
- Makkar, H.P.; Blümmel, M.; Borowy, N.K.; Becker, K. Gravimetric determination of tannins and their correlations with chemical and protein precipitation methods. J. Sci. Food Agric. 1993, 61, 161–165. [Google Scholar] [CrossRef]
- Phuyal, N.; Jha, P.K.; Raturi, P.P.; Rajbhandary, S. Total phenolic, flavonoid contents, and antioxidant activities of fruit, seed, and bark extracts of Zanthoxylum armatum DC. Sci. World J. 2020, 2020, 8780704. [Google Scholar] [CrossRef] [Green Version]
- Mediani, A.; Abas, F.; Khatib, A.; Maulidiani, H.; Shaari, K.; Choi, Y.H.; Lajis, N. 1H-NMR-based metabolomics approach to understanding the drying effects on the phytochemicals in Cosmos caudatus. Food Res. Int. 2012, 49, 763–770. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Solvent Fraction | TPC (mg GAE/g DE) | TFC (mg QE/g DE) |
|---|---|---|
| n-Hexane | 164.44 ± 4.76 d | 78.51 ± 0.61 d |
| Ethyl accetate | 205.45 ± 1.36 a | 156.85 ± 1.33 a |
| Chloroform | 188.35 ± 1.63 b | 111.15 ± 2.29 b |
| n-Butanol | 173.88 ± 2.58 c | 83.19 ± 0.96 c |
| Sr. No. | Metabolites | Chemical Shift (δH ppm) with J Resolved Values |
|---|---|---|
| 1 | Tamarixetin | 7.8 (dd, J = 8.9 Hz, 1.4 Hz) |
| 2 | D-mannitol | 3.68 (dd, J = 8.28 Hz, 5.64 Hz); 3.84 (dd, J = 11.34 Hz, 1.26 Hz) |
| 3 | Gardenin B | 7.90 (d, J = 8.9 Hz) |
| 4 | Nevadensin A | 7.14 (d, J = 2.76 Hz); 8.03 (J = 8.9 Hz) |
| 5 | Tamaridone | 3.90 (s); 6.20 (s) |
| 6 | Tamadone | 6.77 (s); 7.47 (d, J = 1.92 Hz) |
| 7 | Apigenin | 6.30 (s); 7.73 (d, 8.8 Hz) |
| 8 | Catechin | 2.82 (dd, J = 23.04 Hz); 5.86 (d, J = 11.6 Hz) |
| 9 | Myricetin | 6.18 (d, J = 2.04 Hz); 7.36 (s) |
| 10 | Kaempferol | 6.90 (m) |
| 11 | Nonanal | 1.65 (m) |
| 12 | Tetradecane | 0.90 (m) |
| 13 | Rutin | 3.55 (dd, J = 11.6 Hz, 6.3 Hz); 3.64 (dd, J = 13.56 Hz, 2.52 Hz) |
| 14 | Quercetin | 6.89 (d, J = 1.86 Hz); 7.66 (d, J = 2.5 Hz) |
| 15 | Leucine | 3.73 (m) |
| 16 | Lysine | 1.39 (m) |
| 17 | Glutamic acid | 2.39 (m); 2.01 (m); 2.10 (m) |
| 18 | Aspartic acid | 2.80 (dd, J = 14.88 Hz, 7.02 Hz) |
| 19 | Serine | 3.83 (dd, J = 15.66 Hz, 7.8 Hz) |
| 20 | Tyrosine | 3.01 (dd, J = 19.56 Hz, 7.08 Hz) 3.17 (dd, J = 12.54 Hz, 5.46 Hz) |
| 21 | Isorhamnetin | 7.71 (d, J = 31.98 Hz) 9.73 (s) |
| 22 | Vanillic acid | 3.90 (s), 6.94 (d, J = 8.6 Hz) |
| Sr. No. | Metabolites | Binding Energy (kJ/mol) | Hydrogen Bonding | Other Interactions |
|---|---|---|---|---|
| 1 | Tamarixetin | −14.9304 | HIS279, HIS239, ARG312 | |
| 2 | D-mannitol | −11.4684 | SER156, GLY160, LYS155, ASP408 | PHE158, ASN412 |
| 3 | Gardenin B | −11.8613 | ARG312, SER 156 | HIS279, PHE311, TYR31 |
| 4 | Nevadensin A | −14.0441 | GLY159, PHE311, LYS155, PHE157, PHE300, ALA278, TYR313, ARG312 | |
| 5 | Tamaridone | −14.8680 | THR307 | PRO240, HIS239, HIS279, PHE157, PRO309, VAL305 |
| 6 | Tamadone | −12.9285 | PHE157, HIS279, LYS155, HIS239, PRO240, ARG312 | |
| 7 | Apigenin | −12.6378 | ASN241 | HIS279, ARG312 |
| 8 | Catechin | −14.3305 | PHE310, PRO309, GLU304 | HIS279, PHE157, ARG312 |
| 9 | Myricetin | −15.3993 | SER156, HIS239, GLN350 | ARG312, PHE157 |
| 10 | Kaempferol | −14.6258 | ARG439 | PHE158, HIS279, PHE157 |
| 11 | Nonanal | −8.0170 | ARG312 | PHE311, LYS155 |
| 12 | Tetradecane | −8.7492 | HIS279, PHE157, HIS239, LYS155, PHE311 | |
| 13 | Rutin | −15.3011 | HIS279, THR307 | PRO309, TRP242, HIS245 |
| 14 | Quercetin | −14.6382 | GLN350, ASN241 | HIS279, HIS239, PHE157, ARG312 |
| 15 | Leucine | −9.2032 | LYS155, PHE157, HIS239, PRO240 | |
| 16 | Lysine | −10.3665 | PHE157 | PHE157 |
| 17 | Glutamic acid | −11.2994 | ASP408, PHE157 | |
| 18 | Aspartic acid | −9.8152 | GLY160, SER235, PHE310 | GLY159 |
| 19 | Serine | −9.8837 | SER156, PHE310, GLY159 | |
| 20 | Tyrosine | −12.0217 | ASN241, PHE157 | HIS239, PHE158 |
| 21 | Isorhamnetin | −13.8628 | PRO309, HIS239, ARG312 | HIS279, PHE157 |
| 22 | Vanillic acid | −10.5649 | ASP408 | ARG312, HIS239 |
| 23 | Acarbose | −16.4212 | ASP68, ASP349, GLU304 | HIS348, PHE300, ARG312 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Niaz, A.; Adnan, A.; Bashir, R.; Mumtaz, M.W.; Raza, S.A.; Rashid, U.; Tan, C.P.; Tan, T.B. The In Vitro α-Glucosidase Inhibition Activity of Various Solvent Fractions of Tamarix dioica and 1H-NMR Based Metabolite Identification and Molecular Docking Analysis. Plants 2021, 10, 1128. https://doi.org/10.3390/plants10061128
Niaz A, Adnan A, Bashir R, Mumtaz MW, Raza SA, Rashid U, Tan CP, Tan TB. The In Vitro α-Glucosidase Inhibition Activity of Various Solvent Fractions of Tamarix dioica and 1H-NMR Based Metabolite Identification and Molecular Docking Analysis. Plants. 2021; 10(6):1128. https://doi.org/10.3390/plants10061128
Chicago/Turabian StyleNiaz, Aamir, Ahmad Adnan, Rashida Bashir, Muhammad Waseem Mumtaz, Syed Ali Raza, Umer Rashid, Chin Ping Tan, and Tai Boon Tan. 2021. "The In Vitro α-Glucosidase Inhibition Activity of Various Solvent Fractions of Tamarix dioica and 1H-NMR Based Metabolite Identification and Molecular Docking Analysis" Plants 10, no. 6: 1128. https://doi.org/10.3390/plants10061128