
Strain-Specific Biostimulant Effects of Chlorella and Chlamydomonas Green Microalgae on Medicago truncatula


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Characterization of the Selected Microalgae Strains
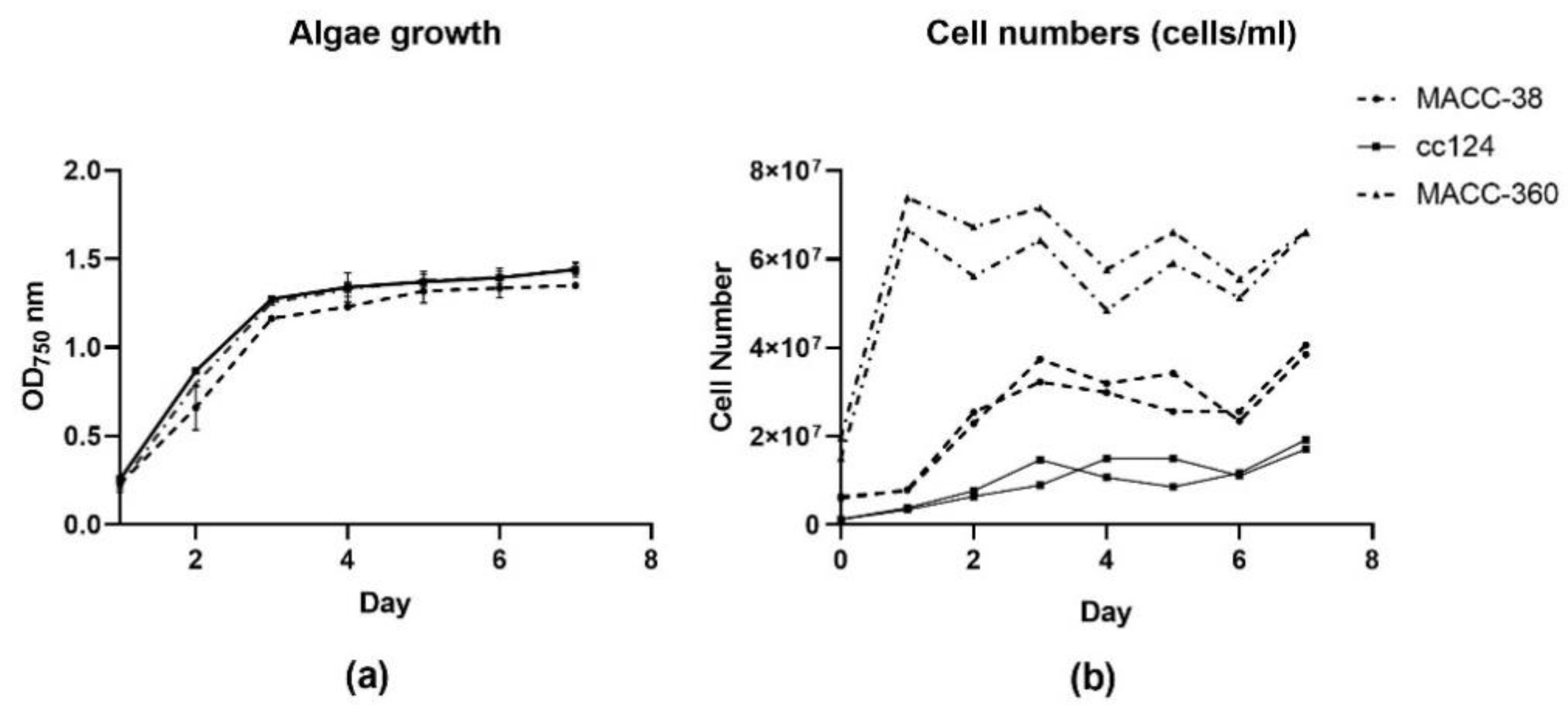
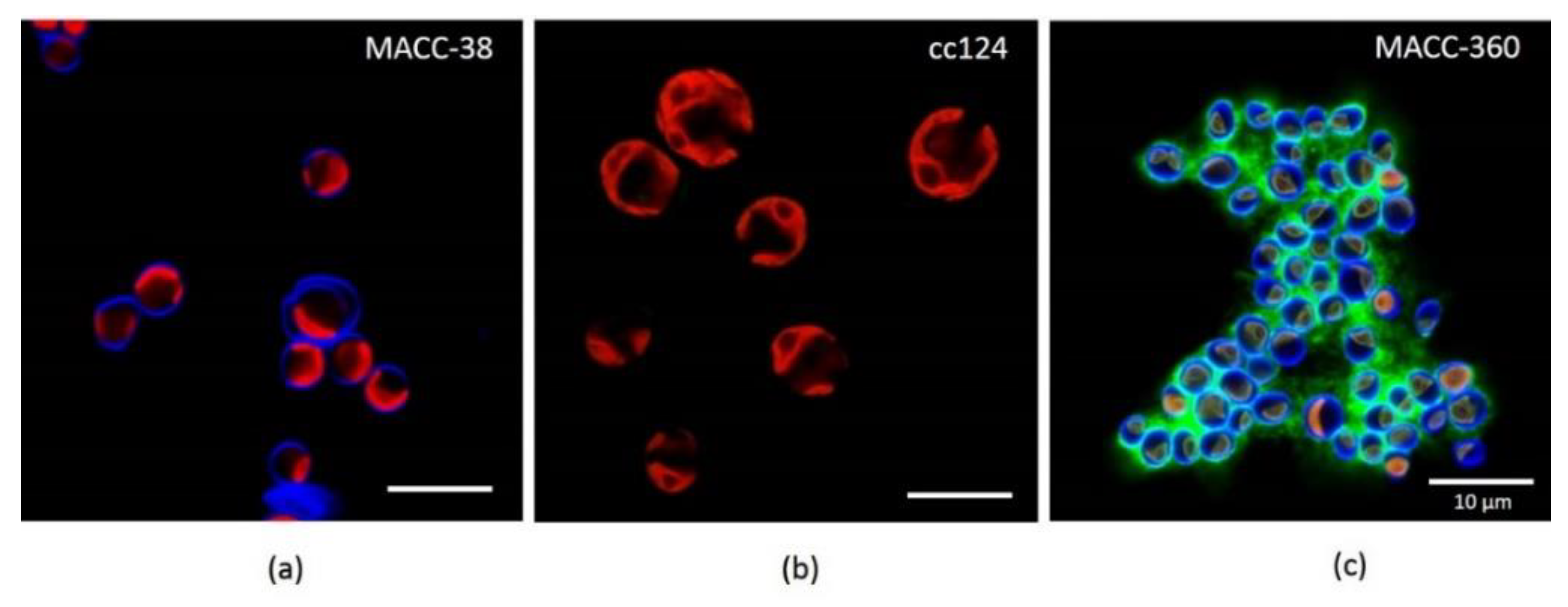
2.1.1. Algae Growth and Morphology
2.1.2. Extracellular Polysaccharide Production of the Selected Microalgae
2.2. Effect of Microalgae Application on Plant Architecture and Canopy Cover
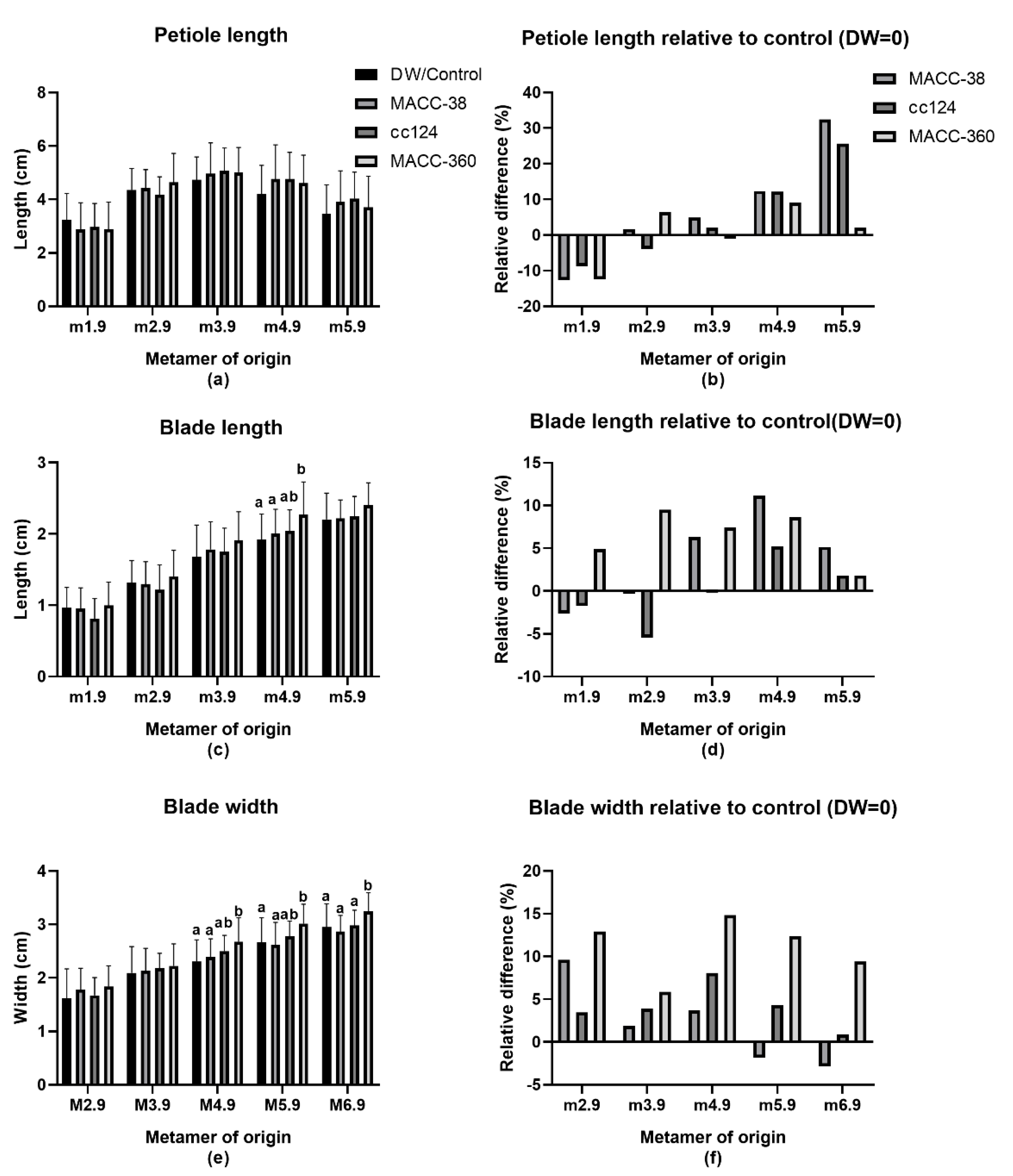
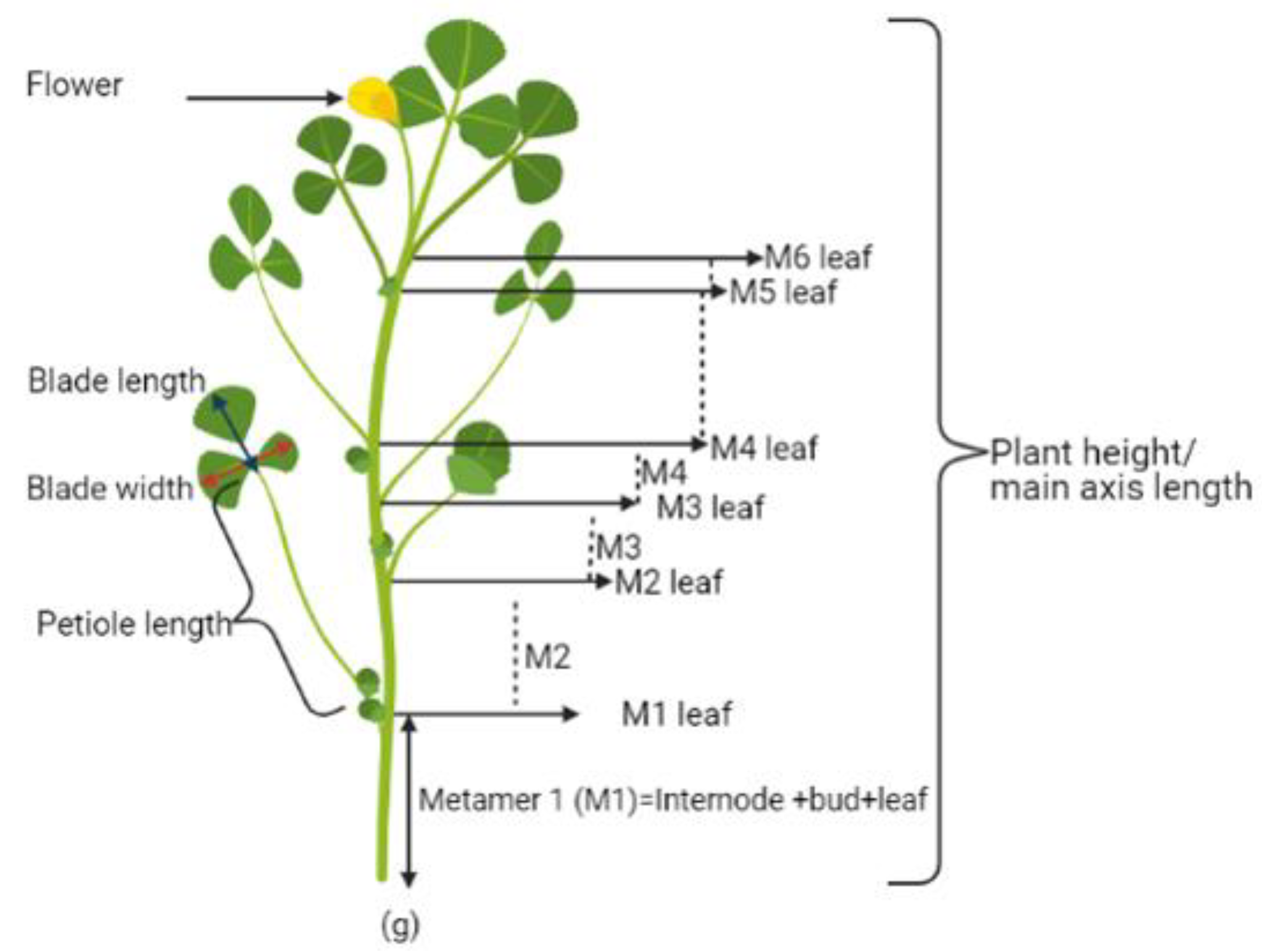
2.3. Effect of Microalgae Application on Leaf Parameters
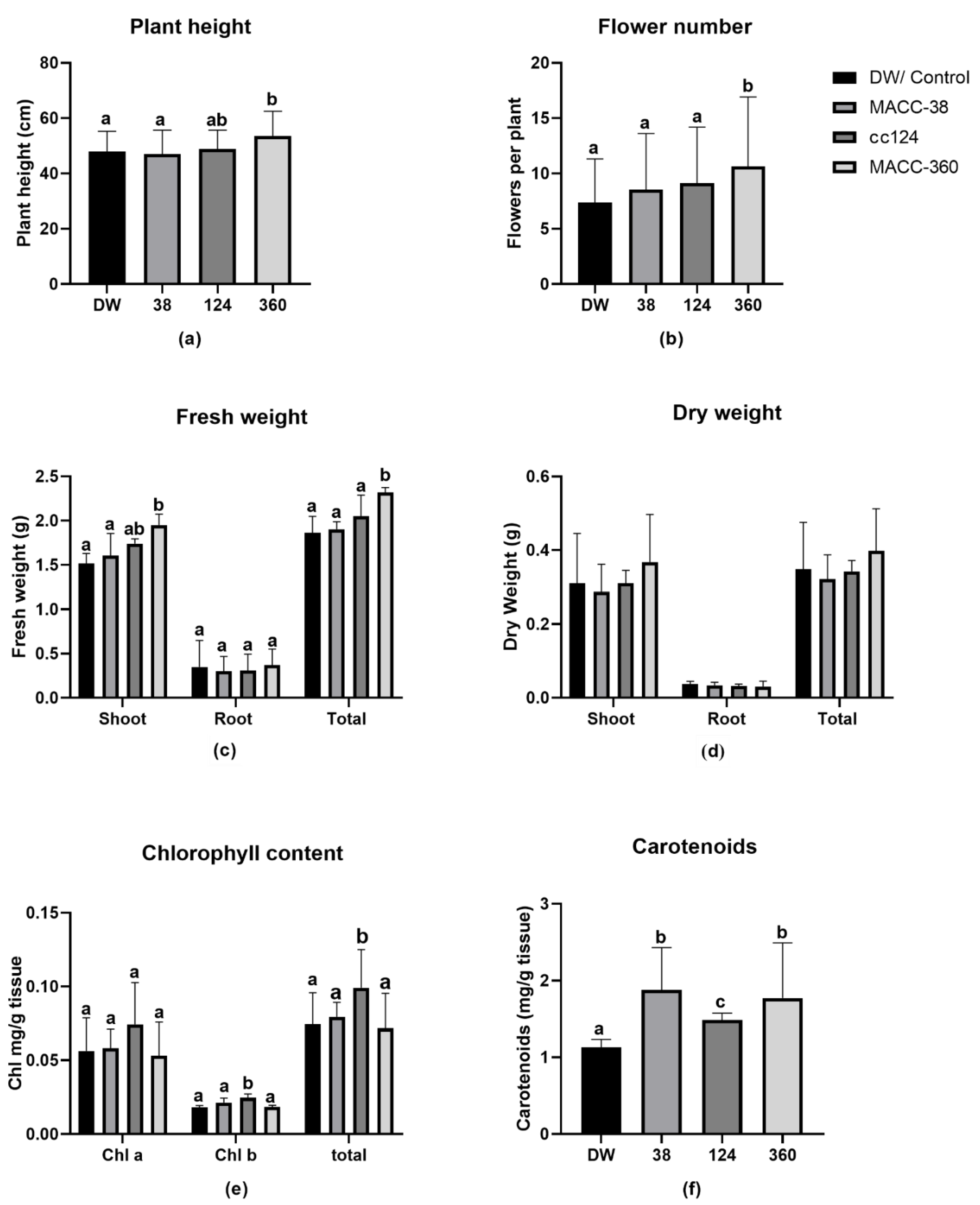
2.4. Effect of Microalgae Application on Plant Height, Flowers, Fresh Weight, Chlorophylls and Carotenoids
3. Discussion
4. Materials and Methods
4.1. Algae Strains
4.1.1. Determination of Algal Growth
4.1.2. Determination of Cell Numbers
4.2. Microscopy
4.2.1. SEM—Scanning Electron Microscopy
4.2.2. CLSM—Confocal Laser Scanning Microscopy
4.3. Preparation of the Algae for Plant Treatment
4.4. Greenhouse-Based Bioassays
4.5. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Singh, R.P. Improving Seed Systems Resiliency at Local Level through Participatory Approach for Adaptation to Climate Change. Adv. Plants Agric. Res. 2016, 6, 00-200. [Google Scholar] [CrossRef] [Green Version]
- Brain, K.R.; Chalopin, M.C.; Turner, T.D.; Blunden, G.; Wildgoose, P.B. Cytokinin activity of commercial aqueous seaweed extract. Plant Sci. Lett. 1973, 1, 241–245. [Google Scholar] [CrossRef]
- Melo, P.C.; Collela, C.F.; Sousa, T.; Pacheco, D.; Cotas, J.; Gonçalves, A.M.; Bahcevandziev, K.; Pereira, L. Seaweed-Based Products and Mushroom β-Glucan as Tomato Plant Immunological Inducers. Vaccines 2020, 8, 524. [Google Scholar] [CrossRef] [PubMed]
- Digruber, T.; Sassa, L.; Cseria, A.; Paula, K.; Nagya, A.V.; Remenyikc, J.; Molnárb, I.; Vassa, I.; Toldid, O.; Gyuriczae, C.; et al. Stimulation of energy willow biomass with triacontanol and seaweed extract. Ind. Crop. Prod. 2018, 120, 104–112. [Google Scholar] [CrossRef]
- Fayzi, L.; Dayan, M.; Cherifi, O.; Boufous, E.H.; Cherifi, K. Biostimulant effect of four moroccan seaweed extracts applied as seed treatment and foliar spray on maize. Asian J. Plant Sci. 2020, 19. [Google Scholar] [CrossRef]
- Kavipriya, R.; Dhanalakshmi, P.K.; Jayashree, S.; Thangaraju, N. Seaweed extract as a biostimulant for legume crop, green gram. J. Ecobiotechnol. 2011, 3, 16–19. [Google Scholar]
- Kocira, S.; Kocira, A.; Kornas, R.; Koszel, M.; Szmigielski, M.; Krajewska, M.; Szparaga, A.; Krzysiak, Z. Effects of seaweed extract on yield and protein content of two common bean (Phaseolus vulgaris L.) cultivars. Legume Res. 2018, 41, 589–593. [Google Scholar] [CrossRef]
- Michalak, I.; Tuhy, Ł.; Chojnacka, K. Seaweed extract by microwave assisted extraction as plant growth biostimulant. Open Chem. 2015, 13, 1183–1195. [Google Scholar] [CrossRef]
- Pomin, V.H. Seaweeds: Agricultural Uses, Biological and Antioxidant Agents; Nova Science Publishers, Inc.: Hauppauge, NY, USA, 2014. [Google Scholar]
- Francesca, S.; Carmen, A.; Mele, B.H.; Schettini, C.; Ambrosinom, P.; Barone, A.; Rigano, M.M. The use of a plant-based biostimulant improves plant performances and fruit quality in tomato plants grown at elevated temperatures. Agronomy 2020, 10, 363. [Google Scholar] [CrossRef] [Green Version]
- Godlewska, A.; Ciepiela, G.A. Assessment of the effect of various biostimulants on medicago X varia T. Martyn yielding and content of selected organic components. Appl. Ecol. Environ. Res. 2018, 16, 5571–5581. [Google Scholar] [CrossRef]
- Gramss, G.; Voigt, K.D.; Bergmann, H. Irrigation with plant extracts in ecofarming increases biomass production and mineral and organic nitrogen content of plants. J. Plant Nutr. Soil Sci. 2003, 166, 612–620. [Google Scholar] [CrossRef]
- Coban, G.A.; Dasgan, H.Y.; Akhoundnejad, Y.; Cimen, B.A. Use of microalgae (Chlorella vulgaris) to save mineral nutrients in soilless grown tomato. Acta Hortic. 2020, 1273. [Google Scholar] [CrossRef]
- Dineshkumar, R.; Kumaravel, R.; Gopalsamy, J.; Sikder, M.N.A.; Sampathkumar, P. Microalgae as Bio-fertilizers for Rice Growth and Seed Yield Productivity. Waste Biomass Valorization 2018, 9, 793–800. [Google Scholar] [CrossRef]
- Dineshkumar, R.; Subramanian, J.; Gopalsamy, J.; Jayasingam, P.; Arumugam, A. The Impact of Using Microalgae as Biofertilizer in Maize (Zea mays L.). Waste Biomass Valorization 2019, 10, 1101–1110. [Google Scholar] [CrossRef]
- Iriti, M.; Scarafoni, A.; Pierce, S.; Castorina, G.; Vitalini, S. Soil application of effective microorganisms (EM) maintains leaf photosynthetic efficiency, increases seed yield and quality traits of bean (Phaseolus vulgaris L.) plants grown on different substrates. Int. J. Mol. Sci. 2019, 20, 2327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kublanovskaya, A.A.; Khapchaeva, S.A.; Zotov, V.S.; Zaytsev, P.A.; Lobakova, E.S.; Solovchenko, A.E. The Effect of the Microalga Chlorella vulgaris Ippas C-1 Biomass Application on Yield, Biological Activity, and the Microbiome of the Soil during Bean Growing. Mosc. Univ. Biol. Sci. Bull. 2019, 74. [Google Scholar] [CrossRef]
- Viaene, T.; Langendries, S.; Beirinckx, S.; Maes, M.; Goormachtig, S. Streptomyces as a plant’s best friend? FEMS Microbiol. Ecol. 2016, 92, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Ahmad, M.T.; Shariff, M.; Yusoff, F.; Goh, Y.M.; Banerjee, S. Applications of microalga Chlorella vulgaris in aquaculture. Rev. Aquac. 2020, 12. [Google Scholar] [CrossRef]
- Hakim, M.; Thaher, M.; Aljabri, H.; Alghasal, G.; Das, P. Microalgae biomass production in municipal wastewater and use of the produced biomass as sustainable biofertilizer. QScience Proc. 2016, 4. [Google Scholar] [CrossRef]
- Sassi, K.K.B.; da Silva, J.A.; Calixto, C.D.; Sassi, R.; Sassi, C.F.d.C. Metabolites of interest for food technology produced by microalgae from the Northeast Brazil. Rev. Cienc. Agron. 2019, 50, 54–65. [Google Scholar] [CrossRef]
- Skjånes, K.; Rebours, C.; Lindblad, P. Potential for green microalgae to produce hydrogen, pharmaceuticals and other high value products in a combined process. Crit. Rev. Biotechnol. 2013, 33, 172–215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suganya, T.; Varman, M.; Masjuki, H.H.; Renganathan, S. Macroalgae and microalgae as a potential source for commercial applications along with biofuels production: A biorefinery approach. Renew. Sustain. Energy Rev. 2016, 55, 909–941. [Google Scholar] [CrossRef]
- Elhafiz, A.A.; Elhafiz, A.A.; Gaur, S.S.; Hamdany, N.; Osman, M.; Lakshmi, T.V.R. Chlorella vulgaris and Chlorella pyrenoidosa live cells appear to be promising sustainable biofertilizer to grow rice, lettuce, cucumber and eggplant in the UAE soils. Recent Res. Sci. Technol. 2015, 7, 14–21. [Google Scholar] [CrossRef]
- Marks, E.A.N.; Montero, O.; Rad, C. The biostimulating effects of viable microalgal cells applied to a calcareous soil: Increases in bacterial biomass, phosphorus scavenging, and precipitation of carbonates. Sci. Total Environ. 2019, 692. [Google Scholar] [CrossRef]
- Kumar, R.; Sharma, A.K.; Ahluwalia, S.S. Advances in Environmental Biotechnology; Springer: Singapore, 2017; pp. 1–288. [Google Scholar] [CrossRef]
- Ramanan, R.; Kim, B.H.; Cho, D.H.; Oh, H.M.; Kim, H.S. Algae-bacteria interactions: Evolution, ecology and emerging applications. Biotechnol. Adv. 2016, 34, 4–29. [Google Scholar] [CrossRef] [Green Version]
- Toyama, T.; Kasuya, M.; Hanaoka, T.; Kobayashi, N.; Tanaka, Y.; Inoue, D.; Sei, K.; Morikawa, M.; Mori, K. Biotechnology for Biofuels Growth promotion of three microalgae, Chlamydomonas reinhadtii, Chlorella vulgaris, and Euglena gracilis, by in situ indigenous bacteria in wastewater effluent. Biotechnol. Biofuels 2018, 11, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Ciepiela, G.A.; Godlewska, A.; Jankowska, J. The effect of biostimulant on yields of mixed grass/red clover stands and chlorophyll content in crop plant leaves under different nitrogen fertilisation regimes. Fresenius Environ. Bull. 2013, 22, 3700–3708. [Google Scholar]
- Paulert, R.; Talamini, V.; Cassolato, J.E.; Duarte, M.E.; Noseda, M.D.; Smania, A.; Stadnik, M.J. Effects of sulfated polysaccharide and alcoholic extracts from green seaweed Ulva fasciata on anthracnose severity and growth of common bean (Phaseolus vulgaris L.). J. Plant Dis. Prot. 2009, 116, 263–270. [Google Scholar] [CrossRef]
- Rengasamy, K.R.; Kulkarni, M.G.; Stirk, W.A.; Van Staden, J. Eckol-a new plant growth stimulant from the brown seaweed Ecklonia maxima. J. Appl. Phycol. 2015, 27, 581–587. [Google Scholar] [CrossRef]
- Navarro-López, E.; Ruíz-Nieto, A.; Ferreira, A.; Acién, F.G.; Gouveia, L. Biostimulant Potential of Scenedesmus obliquus Grown in Brewery Wastewater. Molecules 2020, 25, 664. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rathore, S.S.; Chaudhary, D.R.; Boricha, G.N.; Ghosh, A.; Bhatt, B.P.; Zodape, S.T.; Patolia, J.S. Effect of seaweed extract on the growth, yield and nutrient uptake of soybean (Glycine max) under rainfed conditions. S. Afr. J. Bot. 2009, 75, 351–355. [Google Scholar] [CrossRef] [Green Version]
- Gatamaneni Loganathan, B.; Orsat, V.; Lefsrud, M. Utilizing the microalgal biomass of Chlorella variabilis and Scenedesmus obliquus produced from the treatment of synthetic dairy wastewater as a biofertilizer. J. Plant Nutr. 2020, 1–12. [Google Scholar] [CrossRef]
- El-Sharkawy, M.; El-Beshsbeshy, T.; Al-Shal, R.; Missaoui, A. Effect of Plant Growth Stimulants on Alfalfa Response to Salt Stress. Agric. Sci. 2017, 8, 267–291. [Google Scholar] [CrossRef] [Green Version]
- Sosnowski, J.; Jankowski, K.; Malinowska, E.; Truba, M. The effect of Ecklonia maxima extract on Medicago x varia T. Martyn biomass. J. Soil Sci. Plant Nutr. 2017, 17, 770–780. [Google Scholar] [CrossRef] [Green Version]
- Brahmbhatt, N.H.; Kalasariya, H.S. Effect of algae on seedling growth of “Queen of Forages”. Int. J. Eng. Res. Gen. Sci. 2015, 3, 827–833. [Google Scholar]
- Satish, L.; Ramesh, M. Algae-based extracts as a natural biostimulant for plant growth and development: Current and future prospects. In Photobioreactors: Advancements, Applications and Research; Nova Science Publishers, Inc.: Hauppauge, NY, USA, 2017; pp. 1–13. [Google Scholar]
- Puglisi, I.; Barone, V.; Fragalà, F.; Stevanato, P.; Baglieri, A.; Vitale, A. Effect of microalgal extracts from Chlorella vulgaris and Scenedesmus quadricauda on germination of beta vulgaris seeds. Plants 2020, 9, 675. [Google Scholar] [CrossRef] [PubMed]
- Dineshkumar, R.; Subramanian, J.; Arumugam, A.; Rasheeq, A.A.; Sampathkumar, P. Exploring the microalgae biofertilizer effect on onion cultivation by field experiment. Waste Biomass Valorization 2020, 11, 77–87. [Google Scholar] [CrossRef]
- Faheed, F.A.; Abd-el Fattah, Z. Effect of Chlorella vulgaris as Bio-fertilizer on Growth Parameters and Metabolic Aspects of Lettuce Plant. J. Agric. Soc. Sci. 2008, 4, 165–169. [Google Scholar]
- Agwa, O.K.; Ogugbue, C.J.; Williams, E.E. Field Evidence of Chlorella vulgaris Potentials as a Biofertilizer for Hibiscus esculentus. Int. J. Agric. Res. 2017, 12, 181–189. [Google Scholar] [CrossRef] [Green Version]
- Suleiman, A.K.; Lourenço, K.S.; Clark, C.; Luz, R.L.; da Silva, G.H.; Vet, L.E.; Cantarella, H.; Fernandes, T.V.; Kuramae, E.E. Fertilization with microalgal biomass from wastewater impacts the soil and rhizosphere active microbiomes, greenhouse gas emissions and plant growth. Resour. Conserv. Recycl. 2020, 161. [Google Scholar] [CrossRef]
- De Siqueira Castro, J.; Calijuri, M.L.; Mattiello, E.M.; Ribeiro, V.J.; Assemany, P.P. Algal biomass from wastewater: Soil phosphorus bioavailability and plants productivity. Sci. Total Environ. 2020, 711. [Google Scholar] [CrossRef]
- Rasheed, R.; Bounnit, T.; Al Jabri, H.; Saadaoui, I. Algae-based biofertilizer for date palm cultivation in Qatar. In Proceedings of the Qatar University Annual Research Forum and Exhibition (QUARFE 2020), Qatar, Doha, 28 October 2020. [Google Scholar] [CrossRef]
- Jo, S.W.; Do, J.M.; Na, H.; Hong, J.W.; Kim, I.S.; Yoon, H.S. Assessment of biomass potentials of microalgal communities in open pond raceways using mass cultivation. PeerJ 2020, 8. [Google Scholar] [CrossRef] [PubMed]
- Kholssi, R.; Marks, E.A.; Miñón, J.; Montero, O.; Debdoubi, A.; Rad, C. Biofertilizing effect of Chlorella sorokiniana suspensions on wheat growth. J. Plant Growth Regul. 2019, 38, 644–649. [Google Scholar] [CrossRef]
- Bumandalai, O.; Tserennadmid, R. Effect of Chlorella vulgaris as a biofertilizer on germination of tomato and cucumber seeds. Int. J. Aquat. Biol. 2019, 7, 95–99. [Google Scholar] [CrossRef]
- Özdemİr, S.; Sukatar, A.; Öztekin, G.B. Production of Chlorella vulgaris and its effects on plant growth, yield and fruit quality of organic tomato grown in greenhouse as biofertilizer. Tarim Bilim. Derg. 2016, 22, 596–605. [Google Scholar] [CrossRef]
- Schwarz, D.; Gross, W. Algae affecting lettuce growth in hydroponic systems. J. Hortic. Sci. Biotechnol. 2004, 79, 554–559. [Google Scholar] [CrossRef]
- Kopta, T.; Pavlikova, M.; Sękara, A.; Pokluda, R.; Maršálek, B. Effect of bacterial-algal biostimulant on the yield and internal quality of lettuce (Lactuca sativa L.) produced for spring and summer crop. Not. Bot. Horti Agrobot. Cluj-Napoca 2018, 46, 615–621. [Google Scholar] [CrossRef] [Green Version]
- Grzesik, M.; Romanowska-Duda, Z. Ability of Cyanobacteria and Green Algae to Improve Metabolic Activity and Development of Willow Plants. Pol. J. Environ. Stud. 2015, 24, 1003–1012. [Google Scholar] [CrossRef]
- Zayadan, B.K.; Matorin, D.N.; Baimakhanova, G.B.; Bolathan, K.; Oraz, G.D.; Sadanov, A.K. Promising microbial consortia for producing biofertilizers for rice fields. Microbiology 2014, 83, 391–397. [Google Scholar] [CrossRef]
- Renuka, N.; Guldhe, A.; Prasanna, R.; Singh, P.; Bux, F. Microalgae as multi-functional options in modern agriculture: Current trends, prospects and challenges. Biotechnol. Adv. 2018, 36, 1255–1273. [Google Scholar] [CrossRef]
- Odgerel, B.; Tserendulam, D. Effect of Chlorella as a biofertilizer on germination of wheat and barley grains. Proc. Mong. Acad. Sci. 2016, 26–31. [Google Scholar] [CrossRef] [Green Version]
- Dineshkumar, R.; Subramanian, J.; Sampathkumar, P. Prospective of Chlorella vulgaris to Augment Growth and Yield Parameters Along with Superior Seed Qualities in Black Gram, Vigna mungo (L.). Waste Biomass Valorization 2020, 11. [Google Scholar] [CrossRef]
- Ergun, O.; Dasgan, H.Y.; Isik, O. Effects of microalgae Chlorella vulgaris on hydroponically grown lettuce. Acta Hortic. 2020, 1273. [Google Scholar] [CrossRef]
- Kim, M.J.; Shim, C.K.; Kim, Y.K.; Ko, B.G.; Park, J.H.; Hwang, S.G.; Kim, B.H. Effect of biostimulator Chlorella fusca on improving growth and qualities of chinese chives and spinach in organic farm. Plant Pathol. J. 2018, 34, 567–574. [Google Scholar] [CrossRef] [PubMed]
- De Lima, J.F. Utilization of Chlorella sp. as biostimulant in the germination of melon seeds (Cucumis melo L.). J. Agric. Stud. 2020, 8, 750–773. [Google Scholar] [CrossRef]
- Yanni, Y.G.; Elashmouny, A.A.; Elsadany, A.Y. Differential Response of Cotton Growth, Yield and Fiber Quality to Foliar Application of Spirulina platensis and Urea Fertilizer. Asian J. Adv. Agric. Res. 2020, 29–40. [Google Scholar] [CrossRef] [Green Version]
- Uddin, A.F.M.J.; Rakibuzzaman, M.; naher Wasin, E.W.; Husna, M.A.; Mahato, A.K. Foliar application of Spirulina and Oscillatoria on growth and yield of okra as bio-fertilizer. J. Biosci. Agric. Res. 2019, 22, 1840–1844. [Google Scholar] [CrossRef]
- Wuang, S.C.; Khin, M.C.; Chua, P.Q.D.; Luo, Y.D. Use of Spirulina biomass produced from treatment of aquaculture wastewater as agricultural fertilizers. Algal Res. 2016, 15, 59–64. [Google Scholar] [CrossRef]
- Carillo, P.; Ciarmiello, L.F.; Woodrow, P.; Corrado, G.; Chiaiese, P.; Rouphael, Y. Enhancing sustainability by improving plant salt tolerance through macro-and micro-algal biostimulants. Biology 2020, 9, 253. [Google Scholar] [CrossRef]
- Singh, R.; Parihar, P.; Singh, M.; Bajguz, A.; Kumar, J.; Singh, S.; Singh, V.P.; Prasad, S.M.; Singh, R.; Parihar, P. Uncovering potential applications of cyanobacteria and algal metabolites in biology, agriculture and medicine: Current status and future prospects. Front. Microbiol. 2017, 8, 515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stirk, W.A.; Ördög, V.; Van Staden, J.; Jäger, K. Cytokinin- and auxin-like activity in Cyanophyta and microalgae. J. Appl. Phycol. 2002, 14, 215–221. [Google Scholar] [CrossRef]
- Stirk, W.A.; Ördög, V.; Novák, O.; Rolčík, J.; Strnad, M.; Bálint, P.; van Staden, J. Auxin and cytokinin relationships in 24 microalgal strains1. J. Phycol. 2013, 49, 459–467. [Google Scholar] [CrossRef]
- Kuhn, D.M.; Chandra, J.; Mukherjee, P.K.; Ghannoum, M.A. Comparison of biofilms formed by Candida albicans and Candida parapsilosis on bioprosthetic surfaces. Infect. Immun. 2002, 70, 878–888. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, M.Y.; Lee, D.J.; Tay, J.H. Distribution of extracellular polymeric substances in aerobic granules. Appl. Microbiol. Biotechnol. 2007, 73, 1463–1469. [Google Scholar] [CrossRef] [PubMed]
- Bucciarelli, B.; Hanan, J.; Palmquist, D.; Vance, C.P. A standardized method for analysis of Medicago truncatula phenotypic development. Plant Physiol. 2006, 142, 207–219. [Google Scholar] [CrossRef] [Green Version]
- Chinnaswamy, A.; Coba de la Peña, T.; Stoll, A.; de la Peña Rojo, D.; Bravo, J.; Rincón, A.; Lucas, M.M.; Pueyo, J.J. A nodule endophytic Bacillus megaterium strain isolated from Medicago polymorpha enhances growth, promotes nodulation by Ensifer medicae and alleviates salt stress in alfalfa plants. Ann. Appl. Biol. 2018, 172, 295–308. [Google Scholar] [CrossRef]
- Defez, R.; Andreozzi, A.; Romano, S.; Pocsfalvi, G.; Fiume, I.; Esposito, R.; Angelini, C.; Bianco, C. Bacterial IAA-delivery into medicago root nodules triggers a balanced stimulation of C and N metabolism leading to a biomass increase. Microorganisms 2019, 7, 403. [Google Scholar] [CrossRef] [Green Version]
- Kępczyńska, E.; Karczyński, P. Medicago truncatula root developmental changes by growth-promoting microbes isolated from Fabaceae, growing on organic farms, involve cell cycle changes and WOX5 gene expression. Planta 2020, 251, 1–19. [Google Scholar] [CrossRef]
- Wang, H.; Xu, Y.; Hong, L.; Zhang, X.; Wang, X.; Zhang, J.; Ding, Z.; Meng, Z.; Wang, Z.Y.; Long, R.; et al. HEADLESS Regulates Auxin Response and Compound Leaf Morphogenesis in Medicago truncatula. Front. Plant Sci. 2019, 10, 1024. [Google Scholar] [CrossRef] [Green Version]
- Ge, L.; Peng, J.; Berbel, A.; Madueño, F.; Chen, R. Regulation of compound leaf development by PHANTASTICA in Medicago truncatula. Plant Physiol. 2014, 164, 216–228. [Google Scholar] [CrossRef] [Green Version]
- Ge, L.; Chen, R. PHANTASTICA regulates leaf polarity and petiole identity in Medicago truncatula. Plant Signal. Behav. 2014, 9, e28121. [Google Scholar] [CrossRef] [Green Version]
- Peng, J.; Chen, R. Auxin efflux transporter MtPIN10 regulates compound leaf and flower development in Medicago truncatula. Plant Signal. Behav. 2011, 6, 1537–1544. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meng, Y.; Liu, H.; Wang, H.; Liu, Y.; Zhu, B.; Wang, Z.; Hou, Y.; Zhang, P.; Wen, J.; Yang, H.; et al. HEADLESS, a WUSCHEL homolog, uncovers novel aspects of shoot meristem regulation and leaf blade development in Medicago truncatula. J. Exp. Bot. 2019, 70, 149–163. [Google Scholar] [CrossRef] [Green Version]
- Tadege, M.; Lin, H.; Bedair, M.; Berbel, A.; Wen, J.; Rojas, C.M.; Niu, L.; Tang, Y.; Sumner, L.; Ratet, P.; et al. STENOFOLIA regulates blade outgrowth and leaf vascular patterning in Medicago truncatula and Nicotiana sylvestris. Plant Cell 2011, 23, 2125–2142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, F.; Wang, Y.; Li, G.; Tang, Y.; Kramer, E.M.; Tadege, M. STENOFOLIA recruits TOPLESS to repress ASYMMETRIC LEAVES2 at the leaf margin and promote leaf blade outgrowth in Medicago truncatula. Plant Cell. 2014, 26, 650–664. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, F.; Wang, H.; Tadege, M. Mechanistic insights into STENOFOLIA-mediated leaf blade outgrowth in Medicago truncatula. Model Legume Med. Truncatula 2019, 173–182. [Google Scholar] [CrossRef]
- Niu, L.; Lin, H.; Zhang, F.; Watira, T.W.; Li, G.; Tang, Y.; Wen, J.; Ratet, P.; Mysore, K.S.; Tadege, M.L. LOOSE FLOWER, a WUSCHEL-like Homeobox gene, is required for lateral fusion of floral organs in Medicago truncatula. Plant J. 2015, 81, 480–492. [Google Scholar] [CrossRef] [PubMed]
- Dineshkumar, R.; Duraimurugan, M.; Sharmiladevi, N.; Lakshmi, L.P.; Rasheeq, A.A.; Arumugam, A.; Sampathkumar, P. Microalgal liquid biofertilizer and biostimulant effect on green gram (Vigna radiata L) an experimental cultivation. Biomass Convers. Biorefinery 2020, 1–21. [Google Scholar] [CrossRef]
- Puglisi, I.; La Bella, E.; Rovetto, E.I.; Lo Piero, A.R.; Baglieri, A. Biostimulant effect and biochemical response in lettuce seedlings treated with a Scenedesmus quadricauda extract. Plants 2020, 9, 123. [Google Scholar] [CrossRef] [Green Version]
- Umamaheswari, J.; Shanthakumar, S. Paddy-soaked rice mill wastewater treatment by phycoremediation and feasibility study on use of algal biomass as biofertilizer. J. Chem. Technol. Biotechnol. 2021, 96, 394–403. [Google Scholar] [CrossRef]
- Bianco, C.; Defez, R. Improvement of phosphate solubilization and Medicago plant yield by an indole-3-acetic acid-overproducing strain of Sinorhizobium meliloti. Appl. Environ. Microbiol. 2010, 76, 4626–4632. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maymon, M.; Martínez-Hidalgo, P.; Tran, S.S.; Ice, T.; Craemer, K.; Anbarchian, T.; Sung, T.; Hwang, L.H.; Chou, M.; Fujishige, N.A.; et al. Mining the phytomicrobiome to understand how bacterial coinoculations enhance plant growth. Front. Plant Sci. 2015, 6, 784. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rey, T.; Dumas, B. Plenty Is No Plague: Streptomyces Symbiosis with Crops. Trends Plant Sci. 2017, 22, 30–37. [Google Scholar] [CrossRef]
- Varma, A.; Tripathi, S.; Prasad, R. (Eds.) Plant Microbe Interface; Springer: Berlin/Heidelberg, Germany, 2019; pp. 219–253. [Google Scholar]
- Ördög, V.; Stirk, W.A.; Van Staden, J.; Novák, O.; Strnad, M. Endogenous cytokinins in three genera of microalgae from the chlorophyte. J. Phycol. 2004, 40, 88–95. [Google Scholar] [CrossRef]
- Stirk, W.A.; Bálint, P.; Tarkowská, D.; Novák, O.; Maróti, G.; Ljung, K.; Turečková, V.; Strnad, M.; Ördög, V.; Van Staden, J. Effect of light on growth and endogenous hormones in Chlorella minutissima (Trebouxiophyceae). Plant Physiol. Biochem. 2014, 79, 66–76. [Google Scholar] [CrossRef] [PubMed]
- Ortiz-Moreno, M.L.; Sandoval-Parra, K.X.; Solarte-Murillo, L.V. Chlorella, a potential biofertilizer? Orinoquia 2019, 23, 71–78. [Google Scholar] [CrossRef]
- Yuan, Q.; Li, H.; Wei, Z.; Lv, K.; Gao, C.; Liu, Y.; Zhao, L. Isolation, structures and biological activities of polysaccharides from Chlorella: A review. Int. J. Biol. Macromol. 2020, 163, 2199–2209. [Google Scholar] [CrossRef] [PubMed]
- El-Naggar, N.E.; Hussein, M.H.; Shaaban-Dessuuki, S.A.; Dalal, S.R. Production, extraction and characterization of Chlorella vulgaris soluble polysaccharides and their applications in AgNPs biosynthesis and biostimulation of plant growth. Sci. Rep. 2020, 10, 1–19. [Google Scholar] [CrossRef] [Green Version]
- Colica, G.; Li, H.; Rossi, F.; Li, D.; Liu, Y.; De Philippis, R. Microbial secreted exopolysaccharides affect the hydrological behavior of induced biological soil crusts in desert sandy soils. Soil Biol. Biochem. 2014, 68, 62–70. [Google Scholar] [CrossRef]
- Zakeel, M.C.M.; Safeena, M.I.S. Biofilmed Biofertilizer for Sustainable Agriculture. In Plant Health under Biotic Stress; Springer: Singapore, 2019; pp. 65–82. [Google Scholar]
- Lan, S.; Zhang, Q.; Wu, L.; Liu, Y.; Zhang, D.; Hu, C. Artificially accelerating the reversal of desertification: Cyanobacterial inoculation facilitates the succession of vegetation communities. Environ. Sci. Technol. 2014, 48, 307–315. [Google Scholar] [CrossRef]
- Chamizo, S.; Rodríguez-Caballero, E.; Román, J.R.; Cantón, Y. Effects of biocrust on soil erosion and organic carbon losses under natural rainfall. Catena 2017, 148, 117–125. [Google Scholar] [CrossRef]
- Manolopoulou, E.; Varzakas, T.; Petsalaki, A. Chlorophyll determination in green pepper using two different extraction methods. Curr. Res. Nutr. Food Sci. J. 2016, 4, 52–60. [Google Scholar] [CrossRef]
- Lichtenthaler, H.; Wellburn, A.R. Determinations of total carotenoids and chlorophylls a and b of leaf extracts in different solvents. Adv. Photosynth. Res. 1984, 11, 591–592. [Google Scholar] [CrossRef] [Green Version]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gitau, M.M.; Farkas, A.; Balla, B.; Ördög, V.; Futó, Z.; Maróti, G. Strain-Specific Biostimulant Effects of Chlorella and Chlamydomonas Green Microalgae on Medicago truncatula. Plants 2021, 10, 1060. https://doi.org/10.3390/plants10061060
Gitau MM, Farkas A, Balla B, Ördög V, Futó Z, Maróti G. Strain-Specific Biostimulant Effects of Chlorella and Chlamydomonas Green Microalgae on Medicago truncatula. Plants. 2021; 10(6):1060. https://doi.org/10.3390/plants10061060
Chicago/Turabian StyleGitau, Margaret Mukami, Attila Farkas, Benedikta Balla, Vince Ördög, Zoltán Futó, and Gergely Maróti. 2021. "Strain-Specific Biostimulant Effects of Chlorella and Chlamydomonas Green Microalgae on Medicago truncatula" Plants 10, no. 6: 1060. https://doi.org/10.3390/plants10061060