
Inhibitory Effects of Gyeji-Tang on MMP-9 Activity and the Expression of Adhesion Molecules in IL-4- and TNF-α-Stimulated BEAS-2B Cells


Abstract
:1. Introduction
2. Results
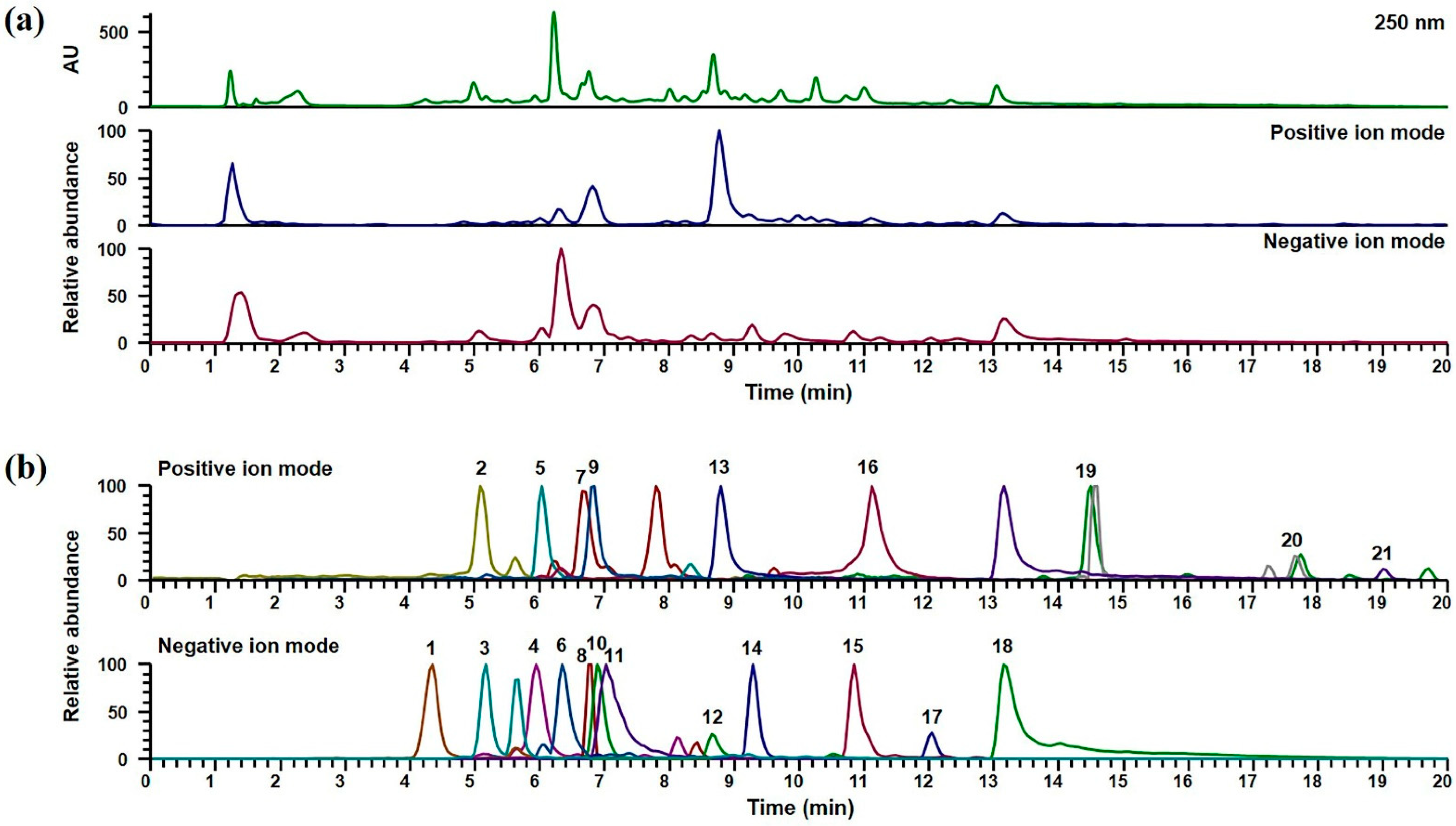
2.1. UPLC-DAD-MS/MS Analysis of GJTWE
2.2. Cytotoxicity of GJTWE in BEAS-2B Cells
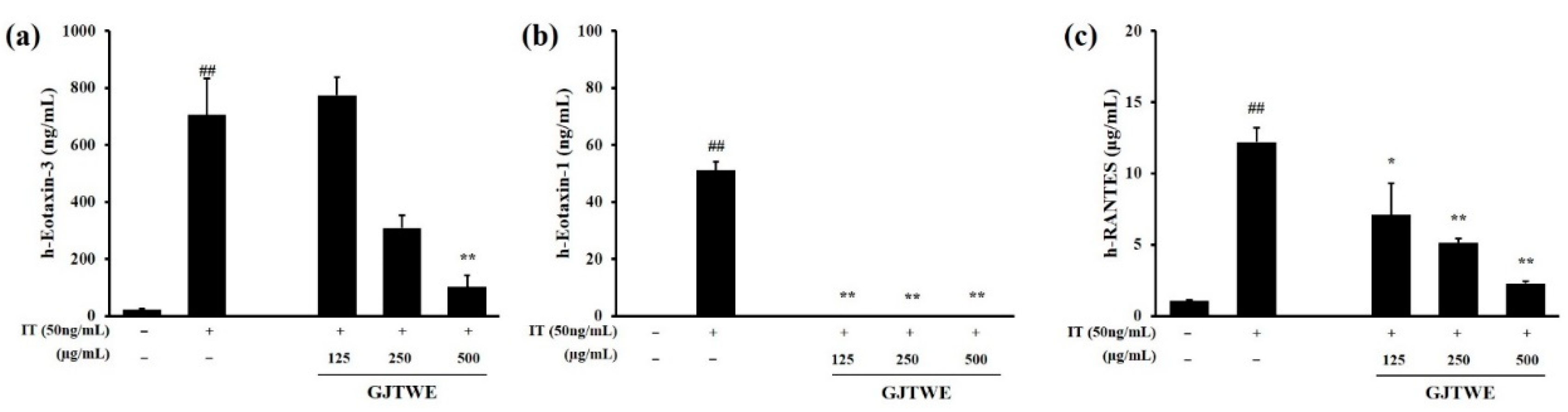
2.3. Effect of GJTWE on Chemokine Oroduction
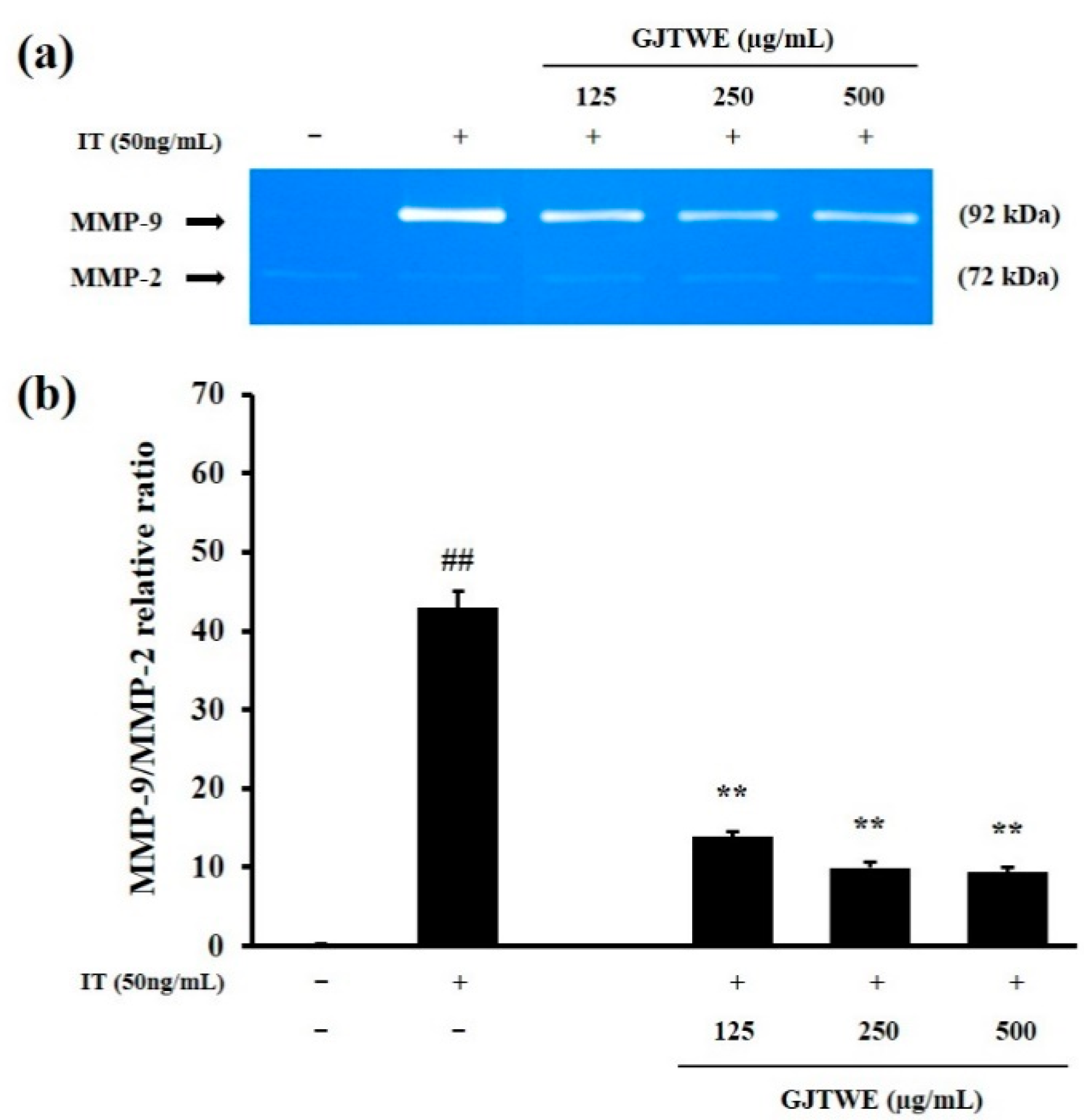
2.4. Effect of GJTWE on MMP-9 Activity
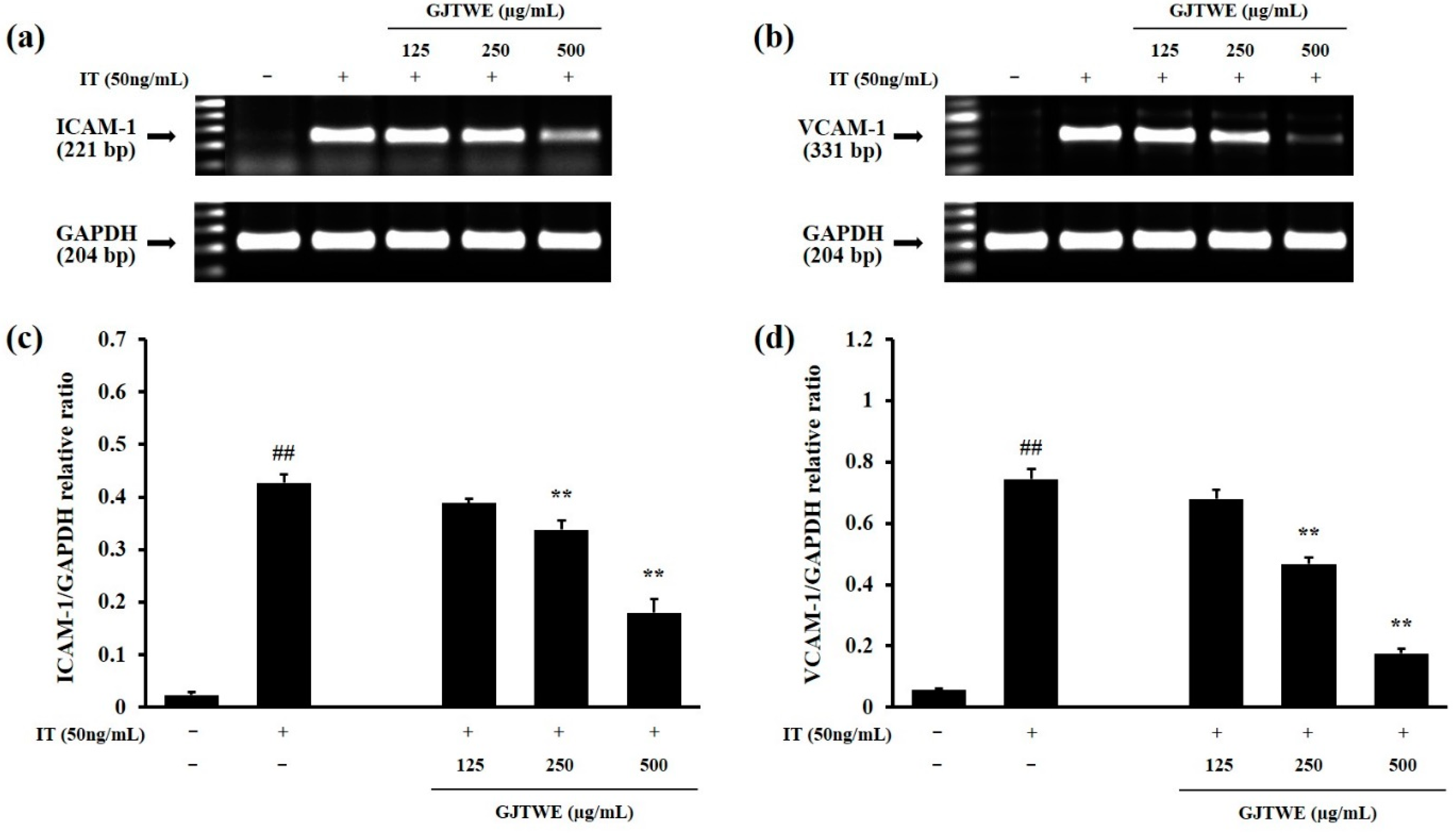
2.5. Effect of GJTWE on the Expression of Adhesion Molecules
3. Discussion
4. Materials and Methods
4.1. Materials and Reagents
4.2. Cell Culture
4.3. Cytotoxicity Assay
4.4. Cell Stimulation
4.5. Measurement of Chemokine Production
4.6. Measurement of MMP-9 Activity
4.7. Measurement of the Adhesion Molecule Expression
4.8. UPLC-DAD-MS-MS Analysis of GJTWE
4.9. Statistical Analyses
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Barnes, P.J. Immunology of asthma and chronic obstructive pulmonary disease. Nat. Rev. Immunol. 2008, 8, 183–192. [Google Scholar] [CrossRef]
- Holgate, S.T.; Lackie, P.; Wilson, S.; Roche, W.; Davies, D. Bronchial epithelium as a key regulator of airway allergen sensitization and remodeling in asthma. Am. J. Respir. Crit. Care Med. 2000, 162, S113–S117. [Google Scholar] [CrossRef]
- Verheggen, M.M.; van Hal, P.T.; Adriaansen-Soeting, P.W.; Goense, B.J.; Hoogsteden, H.C.; Brinkmann, A.O.; Versnel, M.A. Modulation of glucocorticoid receptor expression in human bronchial epithelial cell lines by IL-1 beta, TNF-alpha and LPS. Eur. Respir. J. 1996, 9, 2036–2043. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.H.; Jeon, W.Y.; Lee, M.Y.; Seo, C.S.; Lim, H.S.; Shin, H.K. Inhibitory activity of Socheongryong-tang and its constituent components against the production of RANTES, eotaxin, eotaxin-3 and MMP-9 from BEAS-2B cells. Mol. Med. Rep. 2014, 10, 3035–3046. [Google Scholar] [CrossRef]
- Takeda, N.; Sumi, Y.; Préfontaine, D.; Al Abri, J.; Al Heialy, N.; Al-Ramli, W.; Michoud, M.C.; Martin, J.G.; Hamid, Q. Epithelium-derived chemokines induce airway smooth muscle cell migration. Clin. Exp. Allergy 2009, 39, 1018–1026. [Google Scholar] [CrossRef]
- Verspohl, E.; Podlogar, J. LPS-induced proliferation and chemokine secretion from BEAS-2B cells. Pharmacol. Pharm. 2012, 3. [Google Scholar] [CrossRef] [Green Version]
- Atsuta, J.; Plitt, J.; Bochner, B.S.; Schleimer, R.P. Inhibition of VCAM-1 expression in human bronchial epithelial cells by glucocorticoids. Am. J. Respir. Cell Mol. Biol. 1999, 20, 643–650. [Google Scholar] [CrossRef]
- Atsuta, J.; Sterbinsky, S.A.; Plitt, J.; Schwiebert, L.M.; Bochner, B.S.; Schleimer, R.P. Phenotyping and cytokine regulation of the BEAS-2B human bronchial epithelial cell: Demonstration of inducible expression of the adhesion molecules VCAM-1 and ICAM-1. Am. J. Respir. Cell Mol. Biol. 1997, 17, 571–582. [Google Scholar] [CrossRef] [PubMed]
- Zu, Y.-x.; Lu, H.-y.; Liu, W.-w.; Jiang, X.-w.; Huang, Y.; Li, X.; Zhao, Q.-c.; Xu, Z.-h. Jiang Gui Fang activated interscapular brown adipose tissue and induced epididymal white adipose tissue browning through the PPARγ/SIRT1-PGC1α pathway. J. Ethnopharmacol. 2020, 248, 112271. [Google Scholar] [CrossRef]
- Wang, S.; Chen, L.; Leng, J.; Chen, P.; Fan, X.; Cheng, Y. Fragment ion diagnostic strategies for the comprehensive identification of chemical profile of Gui-Zhi-Tang by integrating high-resolution MS, multiple-stage MS and UV information. J. Pharm. Biomed. Anal. 2014, 98, 22–35. [Google Scholar] [CrossRef]
- Kutsuwa, M.; Nakahata, N.; Kubo, M.; Hayashi, K.; Ohizumi, Y. A comparative study of Kakkon-to and Keishi-to on prostaglandin E(2) release from rabbit astrocytes. Phytomedicine 1998, 5, 275–282. [Google Scholar] [CrossRef]
- Yoo, S.R.; Kim, Y.; Lee, M.Y.; Kim, O.S.; Seo, C.S.; Shin, H.K.; Jeong, S.J. Gyeji-tang water extract exerts anti-inflammatory activity through inhibition of ERK and NF-κB pathways in lipopolysaccharide-stimulated RAW 264.7 cells. BMC Complement. Altern. Med. 2016, 16, 390. [Google Scholar] [CrossRef] [Green Version]
- Lu, X.F. Experimental study on the immunosuppressive effects of gui zhi tang. Zhong Xi Yi Jie He Za Zhi 1989, 9, 262, 283–285. [Google Scholar] [PubMed]
- Fujiwara, H.; Han, Y.; Ebihara, K.; Awale, S.; Araki, R.; Yabe, T.; Matsumoto, K. Daily administration of yokukansan and keishito prevents social isolation-induced behavioral abnormalities and down-regulation of phosphorylation of neuroplasticity-related signaling molecules in mice. BMC Complement. Altern. Med. 2017, 17, 195. [Google Scholar] [CrossRef] [Green Version]
- Motoo, Y.; Su, S.B.; Xie, M.J.; Mouri, H.; Taga, H.; Sawabu, N. Effect of herbal medicine keishi-to (TJ-45) and its components on rat pancreatic acinar cell injuries in vivo and in vitro. Pancreatology 2001, 1, 102–109. [Google Scholar] [CrossRef]
- Wang, J.; Su, B.; Jiang, H.; Cui, N.; Yu, Z.; Yang, Y.; Sun, Y. Traditional uses, phytochemistry and pharmacological activities of the genus Cinnamomum (Lauraceae): A review. Fitoterapia 2020, 146, 104675. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.X.; Braun, M.S.; Wetterauer, P.; Wetterauer, B.; Wink, M. Antioxidant, Cytotoxic, and Antimicrobial Activities of Glycyrrhiza glabra L., Paeonia lactiflora Pall., and Eriobotrya japonica (Thunb.) Lindl. Extracts. Medicines 2019, 6, 43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Z.; Jiang, M.; Wei, X.; Shi, J.; Geng, Z.; Yang, S.; Fu, C.; Guo, L. Rapid discovery of chemical constituents and absorbed components in rat serum after oral administration of Fuzi-Lizhong pill based on high-throughput HPLC-Q-TOF/MS analysis. Chin. Med. 2019, 14, 6. [Google Scholar] [CrossRef] [Green Version]
- Lee, M.-K.; Kim, H.-W.; Kim, Y.; Lee, S.-H.; Jang, H.-H.; Jung, H.-A.; Kim, S.-B.; Lee, S.-H.; Choe, J.-S.; Kim, J.-B. Profiling of flavonoid glycosides in fruits and leaves of jujube (Zizyphus jujuba var. inermis (Bunge) Rehder) using UPLC-DAD-QTOF/MS. Korean J. Food Preserv. 2016, 23, 1004–1011. [Google Scholar] [CrossRef]
- Park, E.J.; Choi, J.; Park, Y.K.; Park, K. Oxidative stress induced by cerium oxide nanoparticles in cultured BEAS-2B cells. Toxicology 2008, 245, 90–100. [Google Scholar] [CrossRef]
- Gong, J.H.; Shin, D.; Han, S.Y.; Kim, J.L.; Kang, Y.H. Kaempferol suppresses eosionphil infiltration and airway inflammation in airway epithelial cells and in mice with allergic asthma. J. Nutr. 2012, 142, 47–56. [Google Scholar] [CrossRef]
- Matsukura, S.; Osakabe, Y.; Sekiguchi, A.; Inoue, D.; Kakiuchi, Y.; Funaki, T.; Yamazaki, Y.; Takayasu, H.; Tateno, H.; Kato, E.; et al. Overexpression of microRNA-155 suppresses chemokine expression induced by Interleukin-13 in BEAS-2B human bronchial epithelial cells. Allergol. Int. 2016, 65, S17–S23. [Google Scholar] [CrossRef] [Green Version]
- Jose, P.J.; Griffiths-Johnson, D.A.; Collins, P.D.; Walsh, D.T.; Moqbel, R.; Totty, N.F.; Truong, O.; Hsuan, J.J.; Williams, T.J. Eotaxin: A potent eosinophil chemoattractant cytokine detected in a guinea pig model of allergic airways inflammation. J. Exp. Med. 1994, 179, 881–887. [Google Scholar] [CrossRef] [Green Version]
- Forssmann, U.; Uguccioni, M.; Loetscher, P.; Dahinden, C.A.; Langen, H.; Thelen, M.; Baggiolini, M. Eotaxin-2, a novel CC chemokine that is selective for the chemokine receptor CCR3, and acts like eotaxin on human eosinophil and basophil leukocytes. J. Exp. Med. 1997, 185, 2171–2176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kitaura, M.; Suzuki, N.; Imai, T.; Takagi, S.; Suzuki, R.; Nakajima, T.; Hirai, K.; Nomiyama, H.; Yoshie, O. Molecular cloning of a novel human CC chemokine (Eotaxin-3) that is a functional ligand of CC chemokine receptor 3. J. Biol. Chem. 1999, 274, 27975–27980. [Google Scholar] [CrossRef] [Green Version]
- Provost, V.; Larose, M.C.; Langlois, A.; Rola-Pleszczynski, M.; Flamand, N.; Laviolette, M. CCL26/eotaxin-3 is more effective to induce the migration of eosinophils of asthmatics than CCL11/eotaxin-1 and CCL24/eotaxin-2. J. Leukoc. Biol. 2013, 94, 213–222. [Google Scholar] [CrossRef] [PubMed]
- Alam, R.; Stafford, S.; Forsythe, P.; Harrison, R.; Faubion, D.; Lett-Brown, M.A.; Grant, J.A. RANTES is a chemotactic and activating factor for human eosinophils. J. Immunol. 1993, 150, 3442–3448. [Google Scholar]
- Brand, K.H.; Ahout, I.M.; de Groot, R.; Warris, A.; Ferwerda, G.; Hermans, P.W. Use of MMP-8 and MMP-9 to assess disease severity in children with viral lower respiratory tract infections. J. Med. Virol. 2012, 84, 1471–1480. [Google Scholar] [CrossRef] [PubMed]
- Malla, N.; Sjøli, S.; Winberg, J.O.; Hadler-Olsen, E.; Uhlin-Hansen, L. Biological and pathobiological functions of gelatinase dimers and complexes. Connect. Tissue Res. 2008, 49, 180–184. [Google Scholar] [CrossRef]
- Hozumi, A.; Nishimura, Y.; Nishiuma, T.; Kotani, Y.; Yokoyama, M. Induction of MMP-9 in normal human bronchial epithelial cells by TNF-alpha via NF-kappa B-mediated pathway. Am. J. Physiol. Lung Cell Mol. Physiol. 2001, 281, L1444–L1452. [Google Scholar] [CrossRef]
- Phillips, P.G.; Birnby, L.M. Nitric oxide modulates caveolin-1 and matrix metalloproteinase-9 expression and distribution at the endothelial cell/tumor cell interface. Am. J. Physiol. Lung Cell Mol. Physiol. 2004, 286, L1055–L1065. [Google Scholar] [CrossRef] [PubMed]
- Proud, D.; Leigh, R. Epithelial cells and airway diseases. Immunol. Rev. 2011, 242, 186–204. [Google Scholar] [CrossRef]
- Nakajima, H.; Sano, H.; Nishimura, T.; Yoshida, S.; Iwamoto, I. Role of vascular cell adhesion molecule 1/very late activation antigen 4 and intercellular adhesion molecule 1/lymphocyte function-associated antigen 1 interactions in antigen-induced eosinophil and T cell recruitment into the tissue. J. Exp. Med. 1994, 179, 1145–1154. [Google Scholar] [CrossRef]
- Yeh, C.F.; Chang, J.S.; Wang, K.C.; Shieh, D.E.; Chiang, L.C. Water extract of Cinnamomum cassia Blume inhibited human respiratory syncytial virus by preventing viral attachment, internalization, and syncytium formation. J. Ethnopharmacol. 2013, 147, 321–326. [Google Scholar] [CrossRef]
- Shou, Q.; Lang, J.; Jin, L.; Fang, M.; Cao, B.; Cai, Y.; Ni, Z.; Qiu, F.; Li, C.; Cao, G.; et al. Total glucosides of peony improve ovalbumin-induced allergic asthma by inhibiting mast cell degranulation. J. Ethnopharmacol. 2019, 244, 112136. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Chang, B.; Wang, B. The effect of herbal medicine including astragalus membranaceus (fisch) bge, codonpsis pilosula and glycyrrhiza uralensis fisch on airway responsiveness. Zhonghua Jie He He Hu Xi Za Zhi 1998, 21, 287–288. [Google Scholar] [PubMed]
- Khan, A.M.; Shahzad, M.; Raza Asim, M.B.; Imran, M.; Shabbir, A. Zingiber officinale ameliorates allergic asthma via suppression of Th2-mediated immune response. Pharm. Biol. 2015, 53, 359–367. [Google Scholar] [CrossRef]
- Ninave, P.B.; Patil, S.D. Antiasthmatic potential of Zizyphus jujuba Mill and Jujuboside B.—Possible role in the treatment of asthma. Respir. Physiol. Neurobiol. 2019, 260, 28–36. [Google Scholar] [CrossRef]
- Zhou, H.; Wu, Q.; Wei, L.; Peng, S. Paeoniflorin inhibits PDGF-BB-induced human airway smooth muscle cell growth and migration. Mol. Med. Rep. 2018, 17, 2660–2664. [Google Scholar] [CrossRef] [Green Version]
- Zhang, T.; Yang, Z.; Yang, S.; Du, J.; Wang, S. Immunoregulatory Effects of Paeoniflorin Exerts Anti-asthmatic Effects via Modulation of the Th1/Th2 Equilibrium. Inflammation 2015, 38, 2017–2025. [Google Scholar] [CrossRef]
- Ram, A.; Mabalirajan, U.; Das, M.; Bhattacharya, I.; Dinda, A.K.; Gangal, S.V.; Ghosh, B. Glycyrrhizin alleviates experimental allergic asthma in mice. Int. Immunopharmacol. 2006, 6, 1468–1477. [Google Scholar] [CrossRef]
- Ma, C.; Ma, Z.; Liao, X.L.; Liu, J.; Fu, Q.; Ma, S. Immunoregulatory effects of glycyrrhizic acid exerts anti-asthmatic effects via modulation of Th1/Th2 cytokines and enhancement of CD4(+)CD25(+)Foxp3+ regulatory T cells in ovalbumin-sensitized mice. J. Ethnopharmacol. 2013, 148, 755–762. [Google Scholar] [CrossRef] [PubMed]
- Hocaoglu, A.B.; Karaman, O.; Erge, D.O.; Erbil, G.; Yilmaz, O.; Bagriyanik, A.; Uzuner, N. Glycyrrhizin and long-term histopathologic changes in a murine model of asthma. Curr. Ther. Res. Clin. Exp. 2011, 72, 250–261. [Google Scholar] [CrossRef] [Green Version]
- Yocum, G.T.; Hwang, J.J.; Mikami, M.; Danielsson, J.; Kuforiji, A.S.; Emala, C.W. Ginger and its bioactive component 6-shogaol mitigate lung inflammation in a murine asthma model. Am. J. Physiol. Lung Cell Mol. Physiol. 2020, 318, L296–Ll303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Z.; Liu, Z.; Uddandrao, V.V.S.; Ponnusamy, P.; Balakrishnan, S.; Brahmanaidu, P.; Vadivukkarasi, S.; Ganapathy, S. Asthma-Alleviating Potential of 6-Gingerol: Effect on Cytokines, Related mRNA and c-Myc, and NFAT1 Expression in Ovalbumin-Sensitized Asthma in Rats. J. Environ. Pathol. Toxicol. Oncol. 2019, 38, 41–50. [Google Scholar] [CrossRef] [PubMed]
- Hwang, Y.H.; Ma, J.Y. Preventive Effects of an UPLC-DAD-MS/MS Fingerprinted Hydroalcoholic Extract of Citrus aurantium in a Mouse Model of Ulcerative Colitis. Planta Med. 2018, 84, 1101–1109. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Rt 1 (min) | Calculated (m/z) | Measured (m/z) | Adduct | Error (ppm) | Formula | MS/MS (m/z) | Identifications |
|---|---|---|---|---|---|---|---|---|
| 1 | 4.32 | 153.0193 | 153.0187 | [M−H]− | −3.9699 | C7H6O4 | 153.0176, 109.0276 | Protocatechuic acid [16] |
| 2 | 5.09 | 139.0390 | 139.0389 | [M + H]+ | −0.5419 | C7H6O3 | 139.0388, 121.0285, 111.0443, 93.0340 | Protocatechualdehyde [16] |
| 3 | 5.16 | 289.0718 | 289.0710 | [M−H]− | 3.5430 | C15H14O6 | 289.0709, 245.0809, 203.0701, 179.0491, 165.0176, 137.0228 | Catechin [17] |
| 4 | 5.93 | 563.1406 | 563.1392 | [M−H]− | −2.5017 | C26H28O14 | 563.1381, 473.1062, 443.0975, 383.0745, 353.0655 | Schaftoside [17] |
| 5 | 6.04 | 481.1704 | 481.1698 | [M + H]+ | −1.3305 | C23H28O11 | 179.0700, 151.0752, 133.0648 | Albiflorin [17] |
| 6 | 6.28 | 525.1614 | 525.1634 | [M + HCO2]− | 3.8676 | C23H28O11 | 327.1074, 165.0541, 121.0277 | Paeoniflorin [17] |
| 7 | 6.67 | 433.1129 | 433.1126 | [M + H]+ | −0.7161 | C21H20O10 | 433.1061, 415.1018, 397.0916, 379.0807, 351.0856, 313.0702 | Isovitexin [19] |
| 8 | 6.73 | 549.1614 | 549.1637 | [M−H]− | 4.2543 | C26H30O13 | 549.1605, 417.1173, 255.0652, 135.0069, 119.0487 | Liquiritin apioside [17] |
| 9 | 6.85 | 183.0652 | 183.0652 | [M + H]+ | −0.1077 | C9H10O4 | 183.0652, 123.0442 | Syringaldehyde [16] |
| 10 | 6.87 | 417.1191 | 417.1182 | [M−H]− | −2.2678 | C21H22O9 | 417.1183, 255.0653, 135.0070, 119.0483 | Liquiritin [17] |
| 11 | 7.01 | 939.1109 | 939.1087 | [M−H]− | −2.3388 | C41H32O26 | 769.0862, 617.0756, 447.0557, 295.0450, 169.0126 | 1,2,3,4,6-O-Pentagalloylglucose [17] |
| 12 | 8.65 | 417.1191 | 417.1183 | [M−H]− | 4.0974 | C21H22O9 | 417.1176, 255.0652, 135.0070, 119.0481 | Isoliquiritin [17] |
| 13 | 8.80 | 147.0441 | 147.0439 | [M + H]+ | −0.9875 | C9H6O2 | 147.0439, 103.0547, 91.0548, 77.0394, 65.0394 | Coumarin [16] |
| 14 | 9.28 | 255.0663 | 255.0656 | [M−H]− | −2.7953 | C15H12O4 | 255.0651, 135.0069, 119.0483 | Liquiritigenin [17] |
| 15 | 10.84 | 629.1876 | 629.1862 | [M + HCO2]− | −2.1643 | C30H32O12 | 431.1359, 165.0540, 121.0276 | Benzoylpaeoniflorin [17] |
| 16 | 11.14 | 133.0648 | 133.0648 | [M + H]+ | −0.0089 | C9H8O | 133.0647, 115.0544, 105.0702, 103.0546, 91.0548, 79.0550, 77.0394, 55.0188 | Cinnamaldehyde [16] |
| 17 | 12.02 | 255.0663 | 255.0671 | [M−H]− | 3.1272 | C15H12O4 | 255.0652, 153.0180, 135.0069, 119.0483 | Isoliquiritigenin [17] |
| 18 | 13.18 | 821.3965 | 821.3999 | [M−H]− | 4.1300 | C42H62O16 | 821.3943, 351.0556, 193.0343 | Glycyrrhizin [17] |
| 19 | 14.52 | 317.1723 | 317.1720 | [M + Na] + | −0.9535 | C17H26O4 | 317.1690, 299.1064 | 6-Gingerol [18] |
| 20 | 17.68 | 277.1798 | 277.1809 | [M + H]+ | 4.0361 | C17H24O3 | 277.2159, 259.2050, 137.0596 | 6-Shogaol [18] |
| 21 | 19.02 | 471.3469 | 471.3463 | [M + H]+ | −1.2879 | C30H46O4 | 471.3464, 407.3315, 317.2107, 189.1635 | Glycyrrhetinic acid [17] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, Y.J.; Jeon, W.-Y.; Hwang, Y.-H.; Lee, M.-Y. Inhibitory Effects of Gyeji-Tang on MMP-9 Activity and the Expression of Adhesion Molecules in IL-4- and TNF-α-Stimulated BEAS-2B Cells. Plants 2021, 10, 951. https://doi.org/10.3390/plants10050951
Kim YJ, Jeon W-Y, Hwang Y-H, Lee M-Y. Inhibitory Effects of Gyeji-Tang on MMP-9 Activity and the Expression of Adhesion Molecules in IL-4- and TNF-α-Stimulated BEAS-2B Cells. Plants. 2021; 10(5):951. https://doi.org/10.3390/plants10050951
Chicago/Turabian StyleKim, Yu Jin, Woo-Young Jeon, Youn-Hwan Hwang, and Mee-Young Lee. 2021. "Inhibitory Effects of Gyeji-Tang on MMP-9 Activity and the Expression of Adhesion Molecules in IL-4- and TNF-α-Stimulated BEAS-2B Cells" Plants 10, no. 5: 951. https://doi.org/10.3390/plants10050951