
The Metabolite Profile in Culture Supernatant of Aster yomena Callus and Its Anti-Photoaging Effect in Skin Cells Exposed to UVB

 ,
,
Abstract
:1. Introduction
2. Results
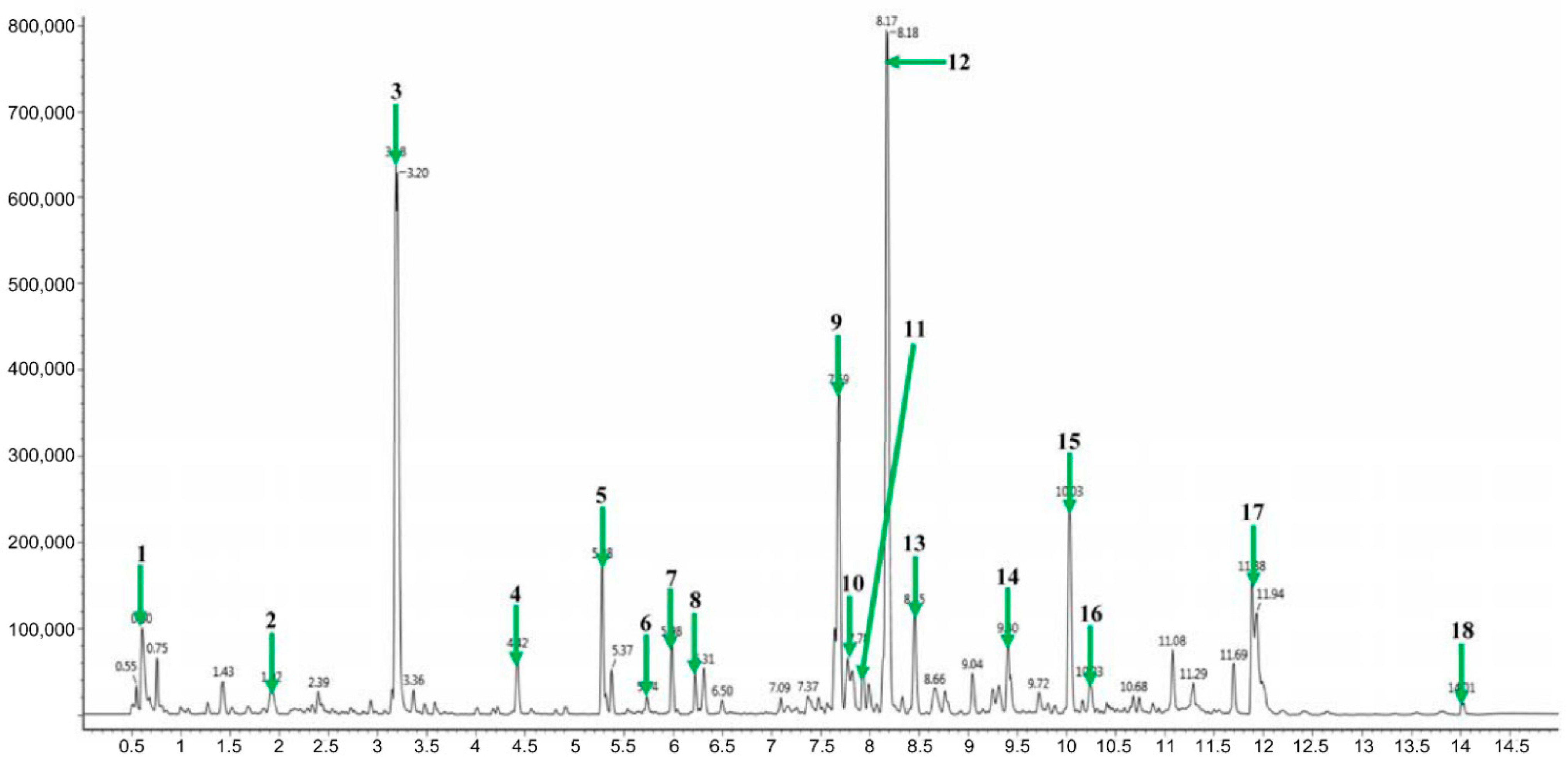
2.1. Identification of Major Metabolites from AYC-CS-E
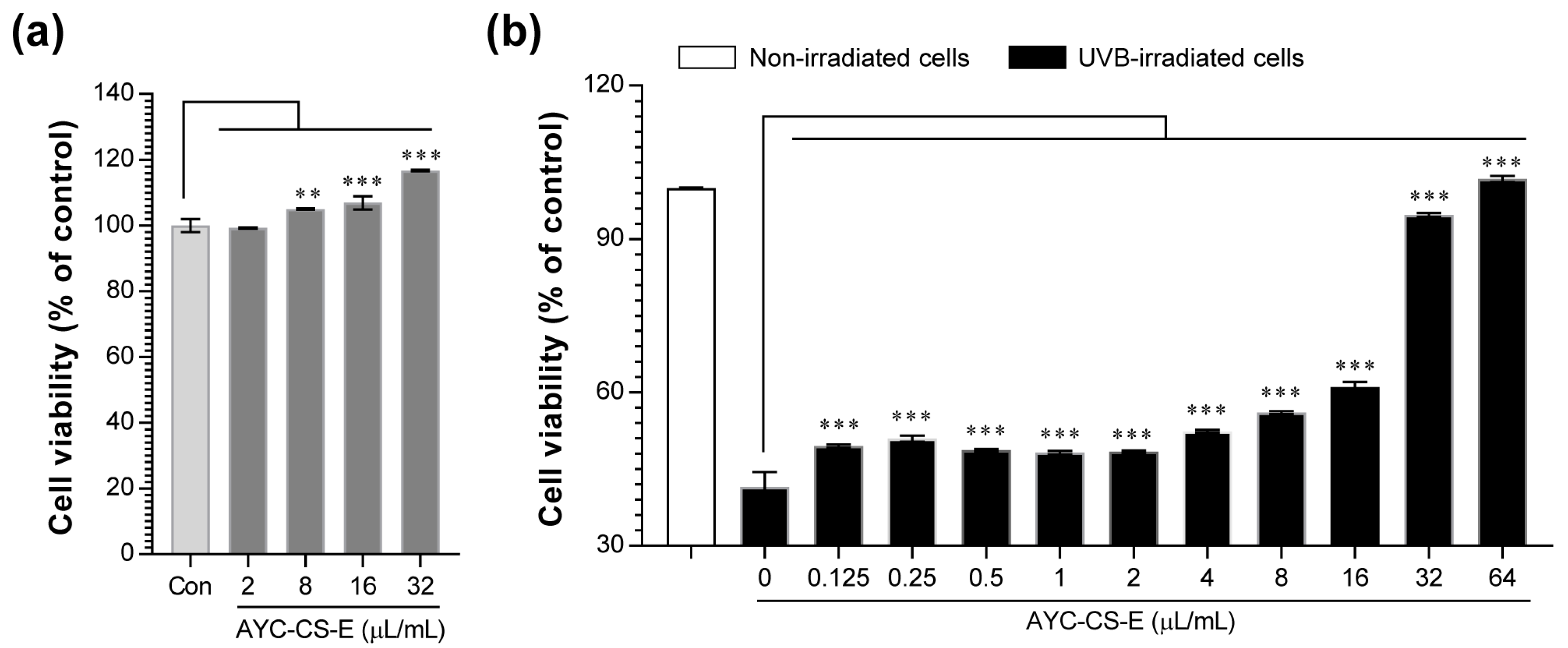
2.2. Cytoprotective Effect of AYC-CS-E on UVB-Irradiated HaCaT Cells
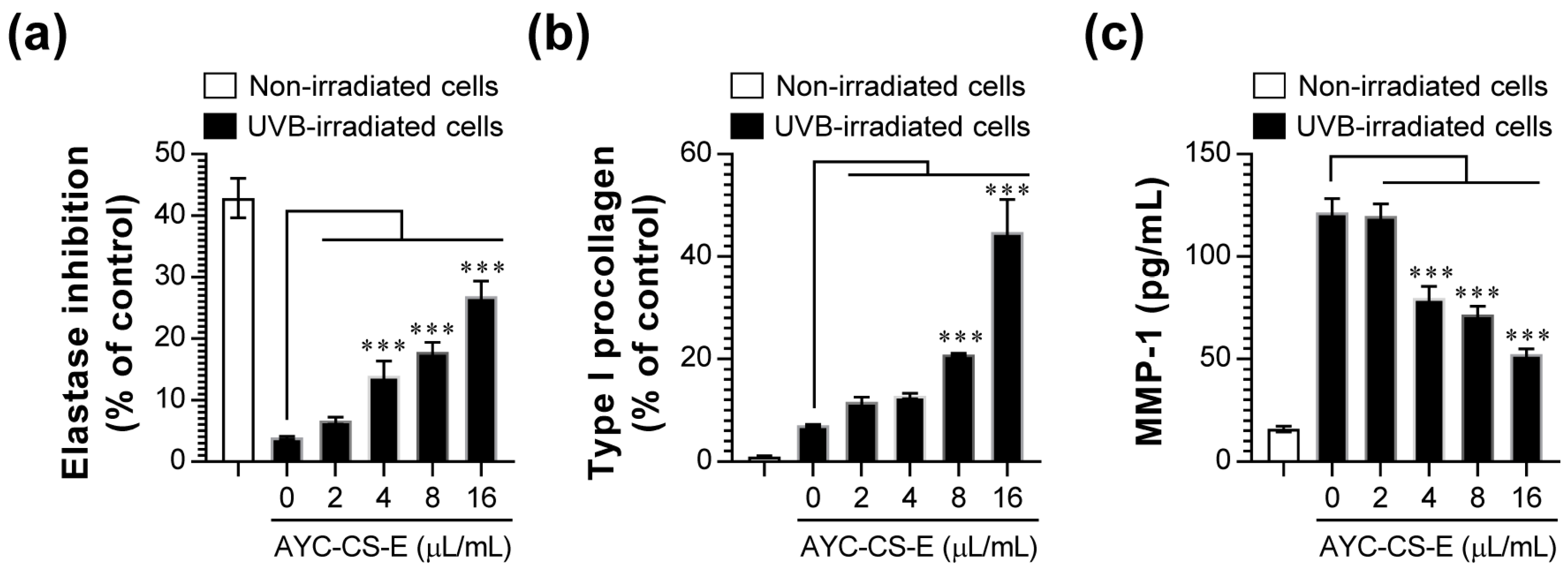
2.3. Elastase Inhibition, Type I Procollagen Synthesis Promotion, and MMP-1 Inhibition Effects of AYC-CS-E
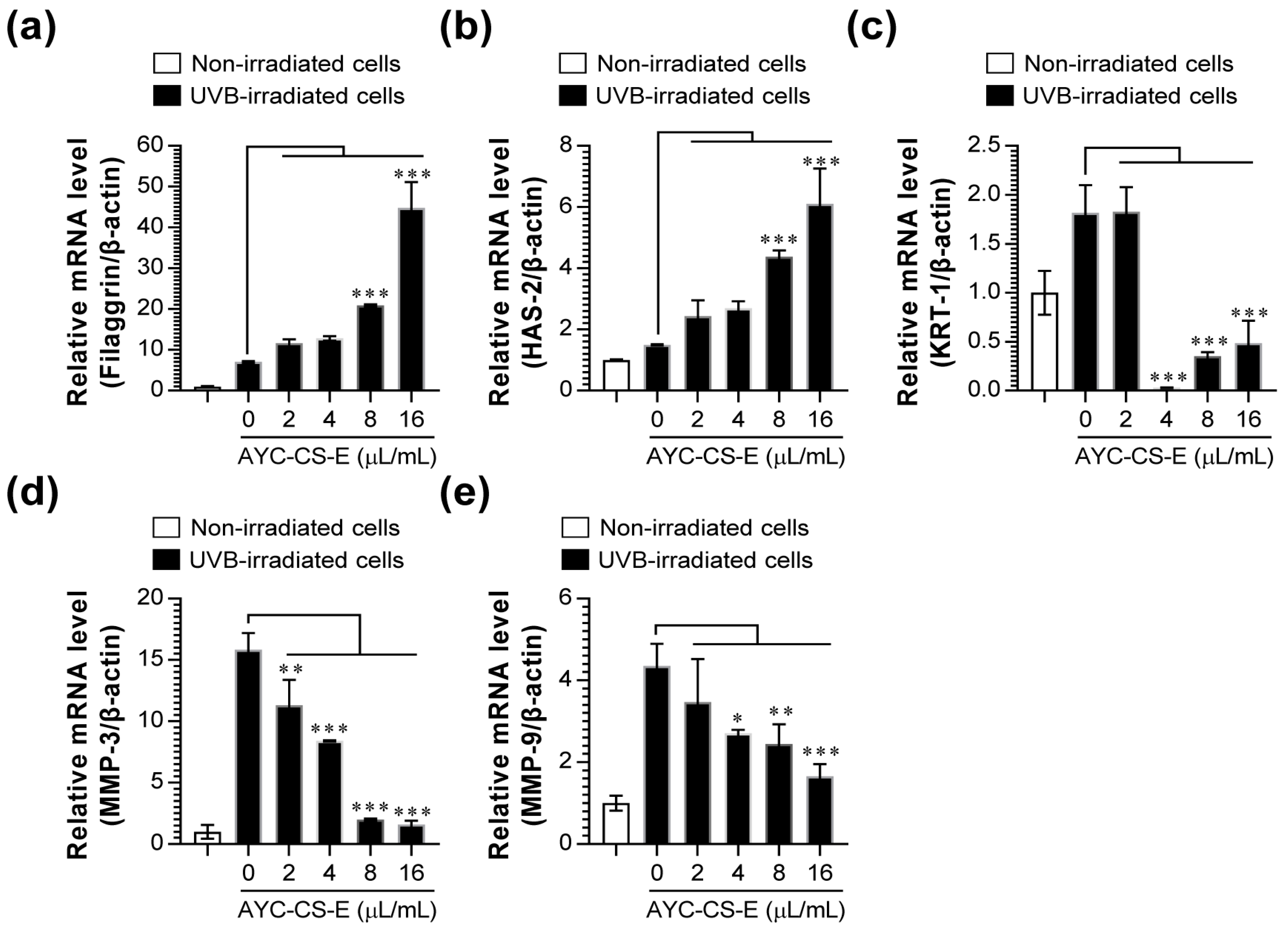
2.4. Expression of Photoaging Regulators Induced by AYC-CS-E
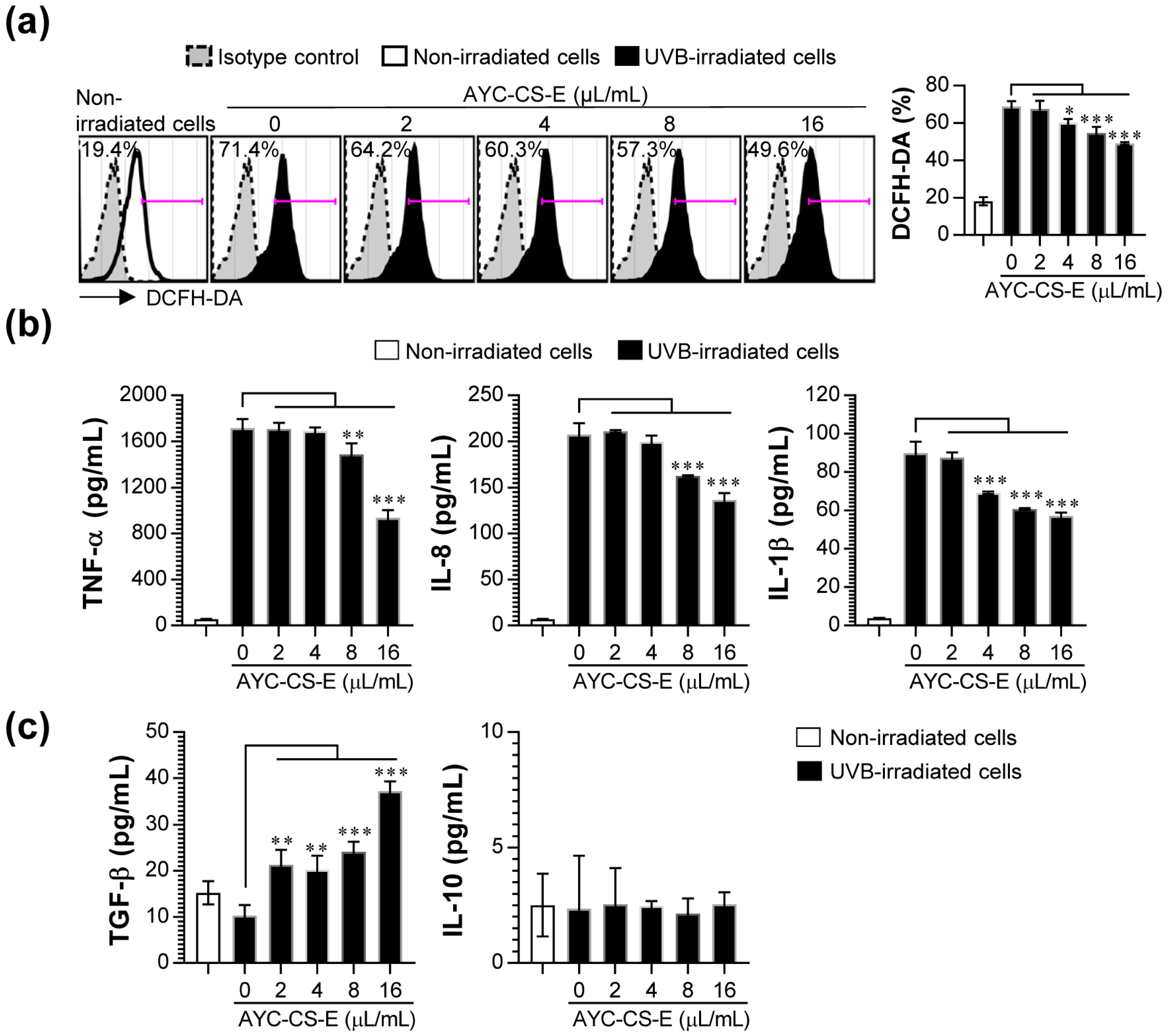
2.5. Anti-Inflammatory and Antioxidant Effect of AYC-CS-E Treatment on UVB-Irradiated HaCaT Cells
2.6. Comparison of Anti-Photoaging Efficacy between AYC-CE-E and Extract (AYC-P-E) Isolated from A. yomena Callus Pellets on UVB-Irradiated HaCaT Cells
3. Discussion
4. Materials and Methods
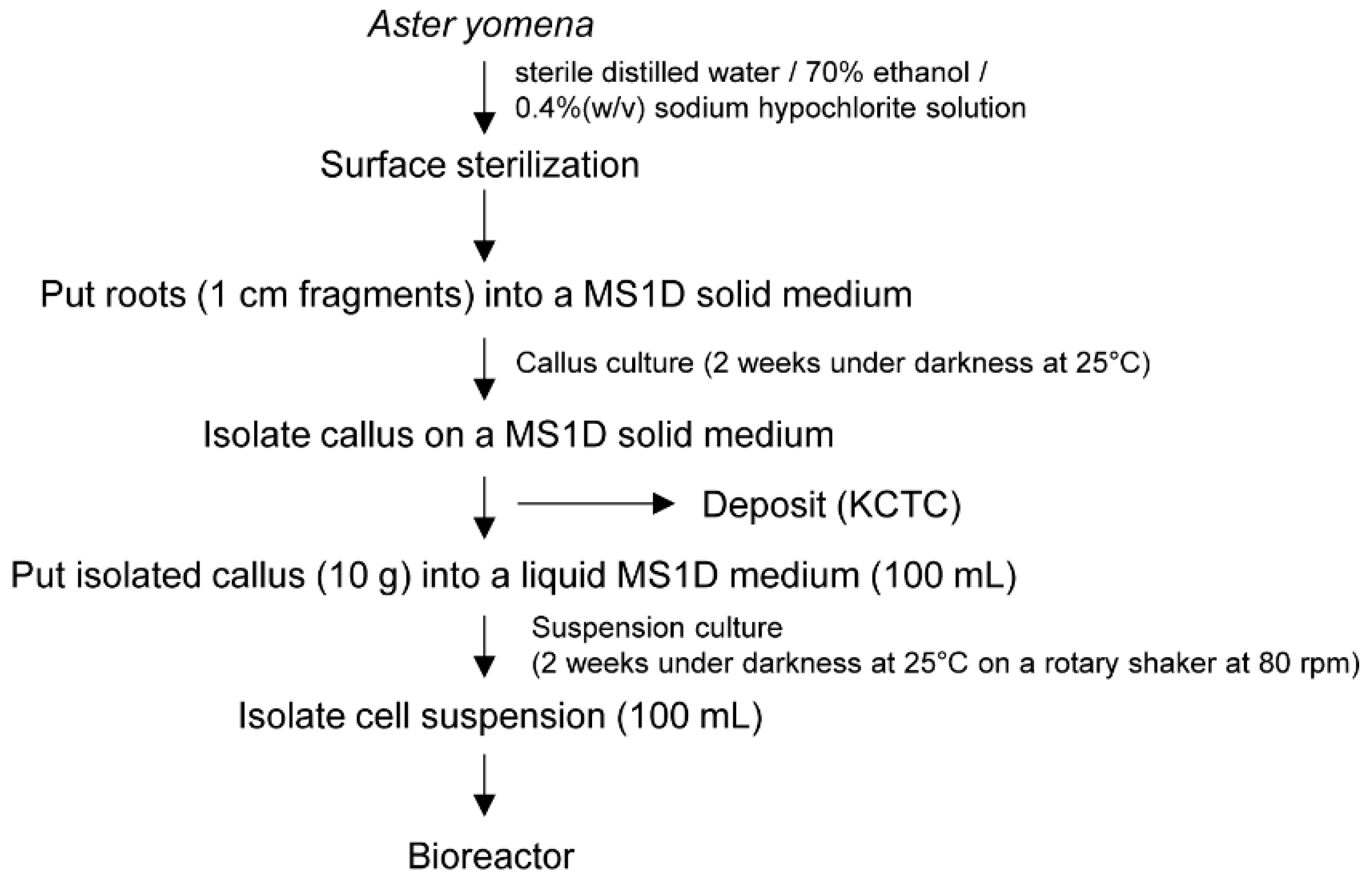
4.1. Plant Sterilization and Callus Induction
4.2. Bioreactor Experiment
4.3. Ultra-Performance Liquid Chromatography-Quadrupole Time-of-Flight/Mass Spectrometry (UPLC-QTOF/MS) Analysis
4.4. Cell Culture
4.5. Cell Viability
4.6. Measurement of Elastase Activity
4.7. Quantitative Real-Time Polymerase Chain Reaction (qRT-PCR)
4.8. Measurement of Intracellular ROS Level
4.9. Measurement of Intracellular Nrf2 and NF-κB Signals
4.10. Enzyme-Linked Immunosorbent Assay (ELISA)
4.11. DPPH Radical Scavenging Assay
4.12. Statistics
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Abbreviations
| AYC-CS-E | Extract isolated from culture supernatant of Aster yomena callus |
| AYC-P-E | Extract isolated from Aster yomena callus pellet |
| UVB | Ultraviolet B |
| qRT-PCR | Quantitative real-time polymerase chain reaction |
| UPLC | Ultra-performance liquid chromatography |
| Q-TOF-MS | Quadrupole time-of-flight mass spectrometry |
| MMP-1 | Matrix metalloproteinase-1 |
| MMP-3 | Matrix metalloproteinase-3 |
| MMP-9 | Matrix metalloproteinase-9 |
| HAS-2 | Hyaluronic acid synthase 2 |
| KRT-1 | Keratin 1 |
| LC/MS | Liquid chromatography/mass spectrometry |
| ROS | Reactive oxygen species |
| DCFH-DA | 2′,7′-dichlorofluorescein diacetate |
| ELISA | Enzyme-linked immunosorbent assay |
| TNF-α | Tumor necrosis factor-α |
| IL-8 | Interleukin-8 |
| IL-1β | Interleukin-1 beta |
| TGF-β | Transforming growth factor-beta |
| IL-10 | Interleukin-10 |
| ACN | Acetonitrile |
| DMEM | Dulbecco’s modified Eagle’s medium |
| DMSO | Dimethyl sulfoxide |
| N-STANA | N-succinyl-tri-alanyl-p-nitroanilide |
| RNA | Ribonucleic acid |
| cDNA | Complementary DNA |
| DPPH | 2,2-diphenyl-1-picrylhydrazyl |
References
- Mukherjee, P.K.; Maity, N.; Nema, N.K.; Sarkar, B.K. Bioactive compounds from natural resources against skin aging. Phytomed. Int. J. Phytother. Phytopharm. 2011, 19, 64–73. [Google Scholar] [CrossRef] [PubMed]
- Gopinath, H.; Karthikeyan, K. Turmeric: A condiment, cosmetic and cure. Indian J. Dermatol. Venereol. Leprol. 2018, 84, 16–21. [Google Scholar] [CrossRef]
- Amberg, N.; Fogarassy, C. Green consumer behavior in the cosmetics market. Resources 2019, 8, 137. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, B.M. Responsible use of medicinal plants for cosmetics. HortScience 2012, 47, 985–991. [Google Scholar] [CrossRef]
- Karim, A.A.; Azlan, A.; Ismail, A.; Hashim, P.; Abd Gani, S.S.; Zainudin, B.H.; Abdullah, N.A. Phenolic composition, antioxidant, anti-wrinkles and tyrosinase inhibitory activities of cocoa pod extract. BMC Complement. Altern. Med. 2014, 14, 381. [Google Scholar]
- Brunt, E.G.; Burgess, J.G. The promise of marine molecules as cosmetic active ingredients. Int. J. Cosmet. Sci. 2018, 40, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Kim, C.R.; Kim, Y.M.; Lee, M.K.; Kim, I.H.; Choi, Y.H.; Nam, T.J. Pyropia yezoensis peptide promotes collagen synthesis by activating the tgf-beta/smad signaling pathway in the human dermal fibroblast cell line hs27. Int. J. Mol. Med. 2017, 39, 31–38. [Google Scholar] [CrossRef] [Green Version]
- Martin, J.L.; Maris, V.; Simberloff, D.S. The need to respect nature and its limits challenges society and conservation science. Proc. Natl. Acad. Sci. USA 2016, 113, 6105–6112. [Google Scholar] [CrossRef] [Green Version]
- Rojas-Downing, M.M.; Nejadhashemi, A.P.; Harrigan, T.; Woznicki, S.A. Climate change and livestock: Impacts, adaptation, and mitigation. Clim. Risk Manag. 2017, 16, 145–163. [Google Scholar] [CrossRef]
- Gheorghe, I.F.; Ion, B. The effects of air pollutants on vegetation and the role of vegetation in reducing atmospheric pollution. In The Impact of Air Pollution on Health, Economy, Environment and Agricultural Sources; Books on Demand: Norderstedt, Germany, 2011; pp. 241–280. [Google Scholar]
- Hameed, B.B.; Ismail, Z.Z. Decolorization, biodegradation and detoxification of reactive blue azo dye using immobilized mixed cells. J. Eng. 2019, 25, 53–66. [Google Scholar] [CrossRef] [Green Version]
- Huang, H.; Wei, Y.; Zhai, Y.; Ouyang, K.; Chen, X.; Bai, L. High frequency regeneration of plants via callus-mediated organogenesis from cotyledon and hypocotyl cultures in a multipurpose tropical tree (Neolamarkia cadamba). Sci. Rep. 2020, 10, 4558. [Google Scholar] [CrossRef] [Green Version]
- Bidabadi, S.S.; Jain, S.M. Cellular, molecular, and physiological aspects of in vitro plant regeneration. Plants 2020, 9, 702. [Google Scholar] [CrossRef]
- Chung, H.H.; Ouyang, H.Y. Use of thidiazuron for high-frequency callus induction and organogenesis of wild strawberry (Fragaria vesca). Plants 2020, 10, 67. [Google Scholar] [CrossRef]
- Georgiev, V.; Slavov, A.; Vasileva, I.; Pavlov, A. Plant cell culture as emerging technology for production of active cosmetic ingredients. Eng. Life Sci. 2018, 18, 779–798. [Google Scholar] [CrossRef] [PubMed]
- Trehan, S.; Michniak-Kohn, B.; Beri, K. Plant stem cells in cosmetics: Current trends and future directions. Future Sci. OA 2017, 3, FSO226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ikeuchi, M.; Sugimoto, K.; Iwase, A. Plant callus: Mechanisms of induction and repression. Plant Cell 2013, 25, 3159–3173. [Google Scholar] [CrossRef] [Green Version]
- Barbulova, A.; Apone, F.; Colucci, G. Plant cell cultures as source of cosmetic active ingredients. Cosmetics 2014, 1, 94–104. [Google Scholar] [CrossRef]
- Malinski, M.P.; Kikowska, M.A.; Soluch, A.; Kowalczyk, M.; Stochmal, A.; Thiem, B. Phytochemical screening, phenolic compounds and antioxidant activity of biomass from Lychnis flos-cuculi L. In vitro cultures and intact plants. Plants 2021, 10, 206. [Google Scholar] [CrossRef]
- Zhong, J.J. Plant cell culture for production of paclitaxel and other taxanes. J. Biosci. Bioeng. 2002, 94, 591–599. [Google Scholar] [CrossRef]
- Gallego-Jara, J.; Lozano-Terol, G.; Sola-Martinez, R.A.; Canovas-Diaz, M.; de Diego Puente, T. A compressive review about taxol®: History and future challenges. Molecules 2020, 25, 5986. [Google Scholar] [CrossRef] [PubMed]
- Hwang, K.A.; Hwang, Y.J.; Song, J. Aster yomena extract ameliorates pro-inflammatory immune response by suppressing nf-kappab activation in raw 264.7 cells. J. Chin. Med. Assoc. 2018, 81, 102–110. [Google Scholar] [CrossRef] [PubMed]
- Hwang, K.-A.; Hwang, Y.-J.; Song, J. Anti-allergic effect of aster yomena on ovalbumin-sensitized mouse and rhl-2h3 cells via th1/th2 cytokine balance. J. Funct. Foods 2018, 44, 1–8. [Google Scholar] [CrossRef]
- Sim, J.H.; Lee, H.S.; Lee, S.; Park, D.E.; Oh, K.; Hwang, K.A.; Kang, H.R.; Ye, S.K.; Kim, H.R. Anti-asthmatic activities of an ethanol extract of aster yomena in an ovalbumin-induced murine asthma model. J. Med. Food 2014, 17, 606–611. [Google Scholar] [CrossRef] [Green Version]
- Matsumura, Y.; Ananthaswamy, H.N. Toxic effects of ultraviolet radiation on the skin. Toxicol. Appl. Pharmacol. 2004, 195, 298–308. [Google Scholar] [CrossRef] [PubMed]
- de Macedo, L.M.; Santos, E.M.D.; Militao, L.; Tundisi, L.L.; Ataide, J.A.; Souto, E.B.; Mazzola, P.G. Rosemary (Rosmarinus officinalis L. Syn salvia Rosmarinus spenn.) and its topical applications: A review. Plants 2020, 9, 651. [Google Scholar] [CrossRef]
- Wang, A.S.; Dreesen, O. Biomarkers of cellular senescence and skin aging. Front. Genet. 2018, 9, 247. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Zhang, K.; Mu, X.; Tian, Q.; Liu, W.; Gao, T.; Ma, X.; Zhang, J. Astragalin attenuates uvb radiation-induced actinic keratosis formation. Anti-Cancer Agents Med. Chem. 2018, 18, 1001–1008. [Google Scholar] [CrossRef] [PubMed]
- Petruk, G.; Del Giudice, R.; Rigano, M.M.; Monti, D.M. Antioxidants from plants protect against skin photoaging. Oxidative Med. Cell. Longev. 2018, 2018, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Song, K.; Sivanesan, I.; Ak, G.; Zengin, G.; Cziaky, Z.; Jeko, J.; Rengasamy, K.R.; Lee, O.N.; Kim, D.H. Screening of bioactive metabolites and biological activities of calli, shoots, and seedlings of Mertensia maritima (L.) gray. Plants 2020, 9, 1551. [Google Scholar] [CrossRef]
- Altemimi, A.; Lakhssassi, N.; Baharlouei, A.; Watson, D.G.; Lightfoot, D.A. Phytochemicals: Extraction, isolation, and identification of bioactive compounds from plant extracts. Plants 2017, 6, 42. [Google Scholar] [CrossRef]
- Xu, G.; Ye, X.; Chen, J.; Liu, D. Effect of heat treatment on the phenolic compounds and antioxidant capacity of citrus peel extract. J. Agric. Food Chem. 2007, 55, 330–335. [Google Scholar] [CrossRef] [PubMed]
- Katiyar, S.K.; Mantena, S.K.; Meeran, S.M. Silymarin protects epidermal keratinocytes from ultraviolet radiation-induced apoptosis and DNA damage by nucleotide excision repair mechanism. PLoS ONE 2011, 6, e21410. [Google Scholar] [CrossRef] [PubMed]
- Moon, H.J.; Lee, S.H.; Ku, M.J.; Yu, B.C.; Jeon, M.J.; Jeong, S.H.; Stonik, V.A.; Zvyagintseva, T.N.; Ermakova, S.P.; Lee, Y.H. Fucoidan inhibits uvb-induced mmp-1 promoter expression and down regulation of type i procollagen synthesis in human skin fibroblasts. Eur. J. Dermatol. 2009, 19, 129–134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Angel, P.; Szabowski, A.; Schorpp-Kistner, M. Function and regulation of ap-1 subunits in skin physiology and pathology. Oncogene 2001, 20, 2413–2423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rhim, D.-B.; Kim, C.; Hwang, J.-K. Inhibitory effect of mojaban (Sargassum fulvellum) extract on ultraviolet b-induced photoaging in hairless mouse skin. Korean J. Food Sci. Technol. 2020, 52, 467–475. [Google Scholar]
- Kang, W.; Choi, D.; Park, S.; Park, T. Carvone decreases melanin content by inhibiting melanoma cell proliferation via the cyclic adenosine monophosphate (camp) pathway. Molecules 2020, 25, 5191. [Google Scholar] [CrossRef]
- Lee, S.J.; Lee, K.B.; Son, Y.H.; Shin, J.; Lee, J.H.; Kim, H.J.; Hong, A.Y.; Bae, H.W.; Kwon, M.A.; Lee, W.J.; et al. Transglutaminase 2 mediates uv-induced skin inflammation by enhancing inflammatory cytokine production. Cell Death Dis. 2017, 8, e3148. [Google Scholar] [CrossRef]
- Wang, P.W.; Cheng, Y.C.; Hung, Y.C.; Lee, C.H.; Fang, J.Y.; Li, W.T.; Wu, Y.R.; Pan, T.L. Red raspberry extract protects the skin against uvb-induced damage with antioxidative and anti-inflammatory properties. Oxidative Med. Cell. Longev. 2019, 2019, 9529676. [Google Scholar] [CrossRef] [Green Version]
- Yaar, M.; Gilchrest, B.A. Photoageing: Mechanism, prevention and therapy. Br. J. Dermatol. 2007, 157, 874–887. [Google Scholar] [CrossRef]
- Saha, S.; Buttari, B.; Panieri, E.; Profumo, E.; Saso, L. An overview of nrf2 signaling pathway and its role in inflammation. Molecules 2020, 25, 5474. [Google Scholar] [CrossRef]
- Ahmed, S.M.U.; Luo, L.; Namani, A.; Wang, X.J.; Tang, X. Nrf2 signaling pathway: Pivotal roles in inflammation. Biochim. Biophys. Acta Mol. Basis Dis. 2017, 1863, 585–597. [Google Scholar] [CrossRef]
- Jariashvili, K.; Madhan, B.; Brodsky, B.; Kuchava, A.; Namicheishvili, L.; Metreveli, N. Uv damage of collagen: Insights from model collagen peptides. Biopolymers 2012, 97, 189–198. [Google Scholar] [CrossRef] [Green Version]
- Solano, F. Photoprotection and skin pigmentation: Melanin-related molecules and some other new agents obtained from natural sources. Molecules 2020, 25, 1537. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baron, A.E.; Asdigian, N.L.; Gonzalez, V.; Aalborg, J.; Terzian, T.; Stiegmann, R.A.; Torchia, E.C.; Berwick, M.; Dellavalle, R.P.; Morelli, J.G.; et al. Interactions between ultraviolet light and mc1r and oca2 variants are determinants of childhood nevus and freckle phenotypes. Cancer Epidemiol. Biomark. Prev. 2014, 23, 2829–2839. [Google Scholar] [CrossRef] [Green Version]
- Shoulders, M.D.; Raines, R.T. Collagen structure and stability. Annu. Rev. Biochem. 2009, 78, 929–958. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rittie, L.; Fisher, G.J. Natural and sun-induced aging of human skin. Cold Spring Harbor Perspect. Med. 2015, 5, a015370. [Google Scholar] [CrossRef]
- Pandel, R.; Poljsak, B.; Godic, A.; Dahmane, R. Skin photoaging and the role of antioxidants in its prevention. ISRN Dermatol. 2013, 2013, 930164. [Google Scholar] [CrossRef]
- Zhang, Z.; Zhu, H.; Zheng, Y.; Zhang, L.; Wang, X.; Luo, Z.; Tang, J.; Lin, L.; Du, Z.; Dong, C. The effects and mechanism of collagen peptide and elastin peptide on skin aging induced by d-galactose combined with ultraviolet radiation. J. Photochem. Photobiol. B Biol. 2020, 210, 111964. [Google Scholar] [CrossRef]
- Tigges, J.; Krutmann, J.; Fritsche, E.; Haendeler, J.; Schaal, H.; Fischer, J.W.; Kalfalah, F.; Reinke, H.; Reifenberger, G.; Stuhler, K.; et al. The hallmarks of fibroblast ageing. Mech. Ageing Dev. 2014, 138, 26–44. [Google Scholar] [CrossRef]
- Chung, J.H.; Seo, J.Y.; Choi, H.R.; Lee, M.K.; Youn, C.S.; Rhie, G.; Cho, K.H.; Kim, K.H.; Park, K.C.; Eun, H.C. Modulation of skin collagen metabolism in aged and photoaged human skin in vivo. J. Investig. Dermatol. 2001, 117, 1218–1224. [Google Scholar] [CrossRef] [Green Version]
- Im, A.R.; Lee, B.; Kang, D.J.; Chae, S. Protective effects of tyndallized lactobacillus acidophilus idcc 3302 against uvbinduced photodamage to epidermal keratinocytes cells. Int. J. Mol. Med. 2019, 43, 2499–2506. [Google Scholar]
- Yoshizumi, M.; Nakamura, T.; Kato, M.; Ishioka, T.; Kozawa, K.; Wakamatsu, K.; Kimura, H. Release of cytokines/chemokines and cell death in uvb-irradiated human keratinocytes, hacat. Cell Biol. Int. 2008, 32, 1405–1411. [Google Scholar] [CrossRef]
- Pavicic, T.; Gauglitz, G.G.; Lersch, P.; Schwach-Abdellaoui, K.; Malle, B.; Korting, H.C.; Farwick, M. Efficacy of cream-based novel formulations of hyaluronic acid of different molecular weights in anti-wrinkle treatment. J. Drugs Dermatol. 2011, 10, 990–1000. [Google Scholar] [PubMed]
- McAleer, M.A.; Irvine, A.D. The multifunctional role of filaggrin in allergic skin disease. J. Allergy Clin. Immunol. 2013, 131, 280–291. [Google Scholar] [CrossRef]
- Weidenthaler, B.; Hausser, I.; Anton-Lamprecht, I. Is filaggrin really a filament-aggregating protein in vivo? Arch. Dermatol. Res. 1993, 285, 111–120. [Google Scholar] [CrossRef] [PubMed]
- Moravcova, M.; Libra, A.; Dvorakova, J.; Viskova, A.; Muthny, T.; Velebny, V.; Kubala, L. Modulation of keratin 1, 10 and involucrin expression as part of the complex response of the human keratinocyte cell line hacat to ultraviolet radiation. Interdiscip. Toxicol. 2013, 6, 203–208. [Google Scholar] [CrossRef] [Green Version]
- Tu, Y.; Quan, T. Oxidative stress and human skin connective tissue aging. Cosmetics 2016, 3, 28. [Google Scholar] [CrossRef]
- Krstic, J.; Trivanovic, D.; Mojsilovic, S.; Santibanez, J.F. Transforming growth factor-beta and oxidative stress interplay: Implications in tumorigenesis and cancer progression. Oxid. Med. Cell. Longev. 2015, 2015, 654594. [Google Scholar] [CrossRef] [PubMed]
- Kucich, U.; Rosenbloom, J.C.; Abrams, W.R.; Rosenbloom, J. Transforming growth factor-beta stabilizes elastin mrna by a pathway requiring active smads, protein kinase c-delta, and p38. Am. J. Respir. Cell Mol. Biol. 2002, 26, 183–188. [Google Scholar] [CrossRef]
- Chu, C.C.; Di Meglio, P.; Nestle, F.O. Harnessing dendritic cells in inflammatory skin diseases. Semin. Immunol. 2011, 23, 28–41. [Google Scholar] [CrossRef]
- Sugaya, M. The role of th17-related cytokines in atopic dermatitis. Int. J. Mol. Sci. 2020, 21, 1314. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Helou, D.G.; Martin, S.F.; Pallardy, M.; Chollet-Martin, S.; Kerdine-Romer, S. Nrf2 involvement in chemical-induced skin innate immunity. Front. Immunol. 2019, 10, 1004. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, K.; Hasegawa, J.; Asamitsu, K.; Okamoto, T. Prevention of the ultraviolet b-mediated skin photoaging by a nuclear factor kappab inhibitor, parthenolide. J. Pharmacol. Exp. Ther. 2005, 315, 624–630. [Google Scholar] [CrossRef] [Green Version]
- Tungmunnithum, D.; Thongboonyou, A.; Pholboon, A.; Yangsabai, A. Flavonoids and other phenolic compounds from medicinal plants for pharmaceutical and medical aspects: An overview. Mediences 2018, 5, 93. [Google Scholar] [CrossRef]
- Moise, A.R.; Bobis, O. Baccharis dracunculifolia and Dalbergia ecastophyllum, main plant sources for bioactive properties in green and red Brazilian propolis. Plants 2020, 9, 1619. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.Y.; Go, K.C.; Song, Y.S.; Jeong, Y.S.; Kim, E.J.; Kim, B.J. Extract of the mycelium of t. Matsutake inhibits elastase activity and tpa-induced mmp-1 expression in human fibroblasts. Int. J. Mol. Med. 2014, 34, 1613–1621. [Google Scholar] [CrossRef] [PubMed]
- De Palma, M.; Ambrosone, A.; Leone, A.; Del Gaudio, P.; Ruocco, M.; Turiak, L.; Bokka, R.; Fiume, I.; Tucci, M.; Pocsfalvi, G. Plant roots release small extra cellular vesicles with antifungal activity. Plants 2020, 9, 1777. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peak No. | Retention Time (min) | Identification | Exact Mass (m/z) | Fragment Ions (m/z) |
|---|---|---|---|---|
| 1 | 0.60 | Robustic acid | 381.07 | 349, 251, 233, 175 |
| 2 | 1.92 | 3,4-Dicaffeoyl-1,5-quinolactone | 499.12 | 319, 163 |
| 3 | 3.18 | Acetylpterosin C | 277.10 | 235, 217, 131 |
| 4 | 4.42 | Pterosin N | 235.09 | 217, 175, 147, 91 |
| 5 | 5.28 | L-Thyronine | 274.27 | 256, 230 |
| 6 | 5.74 | 3,5-Di-O-methyl-8-prenylafzelechin-4beta-ol | 387.18 | 147, 105 |
| 7 | 5.98 | Dehydrophytosphingosine | 316.28 | 298, 280 |
| 8 | 6.28 | Phytosphingosine | 318.30 | 300, 270, 155 |
| 9 | 7.69 | LysoPC(18:2) | 520.33 | 502, 337, 184, 104 |
| 10 | 7.78 | α-Linolenic acid | 279.23 | 261, 243, 109, 95, 81 |
| 11 | 7.93 | LysoPC(16:0) | 496.33 | 478, 313, 184 |
| 12 | 8.17 | 478, 313, 184, 104 | ||
| 13 | 8.45 | LysoPC(18:1) | 522.35 | 504, 184, 104 |
| 14 | 9.40 | LysoPC(18:0) | 524.36 | 506, 341, 184, 104 |
| 15 | 10.03 | Palmitic amide | 256.26 | 186 |
| 16 | 10.23 | Oleamide | 282.27 | 265, 247 |
| 17 | 11.88 | 13Z-Docosenamide | 338.34 | 321, 303 |
| 18 | 14.01 | PC(18:2/16:0) | 758.57 | 575, 337, 184 |
| Primers | Forward Primer Sequence (5‘ to 3’) | Reverse Primer Sequence (5‘ to 3’) |
|---|---|---|
| Filaggrin | ATCTTCTCGGGAGCAGTCAA | ACCCGGATTCACCATAATCA |
| HAS-2 | CCTGGGCTATGCAACAAAAT | TAAGGCAGCTGGCAAAAGAT |
| KRT-1 | AAGCTGAATGACCTGGAGGA | ACCTCCACTGATGGTGGTGT |
| MMP-3 | CTTTCCTFFCATCCCGAAGT | GCATAGGCATGGGCCAAAAC |
| MMP-9 | GTGCTCCTGGTGCTGGGCTG | GGTGCCACTTGAGGTCGCCC |
| β-actin | GCACCACACCTTCTACAATG | TGCTTGCTGATCCACATCTG |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, W.S.; Seo, J.H.; Lee, J.-I.; Ko, E.-S.; Cho, S.-M.; Kang, J.-R.; Jeong, J.-H.; Jeong, Y.J.; Kim, C.Y.; Cha, J.-D.; et al. The Metabolite Profile in Culture Supernatant of Aster yomena Callus and Its Anti-Photoaging Effect in Skin Cells Exposed to UVB. Plants 2021, 10, 659. https://doi.org/10.3390/plants10040659
Kim WS, Seo JH, Lee J-I, Ko E-S, Cho S-M, Kang J-R, Jeong J-H, Jeong YJ, Kim CY, Cha J-D, et al. The Metabolite Profile in Culture Supernatant of Aster yomena Callus and Its Anti-Photoaging Effect in Skin Cells Exposed to UVB. Plants. 2021; 10(4):659. https://doi.org/10.3390/plants10040659
Chicago/Turabian StyleKim, Woo Sik, Jeong Hun Seo, Jae-In Lee, Eun-Sil Ko, Sang-Min Cho, Jea-Ran Kang, Jong-Hoon Jeong, Yu Jeong Jeong, Cha Young Kim, Jeong-Dan Cha, and et al. 2021. "The Metabolite Profile in Culture Supernatant of Aster yomena Callus and Its Anti-Photoaging Effect in Skin Cells Exposed to UVB" Plants 10, no. 4: 659. https://doi.org/10.3390/plants10040659