
Antiinflammatory Medicinal Plants from the Ugandan Greater Mpigi Region Act as Potent Inhibitors in the COX-2/PGH2 Pathway

 ,
,  and
and
Abstract
:1. Introduction
2. Results
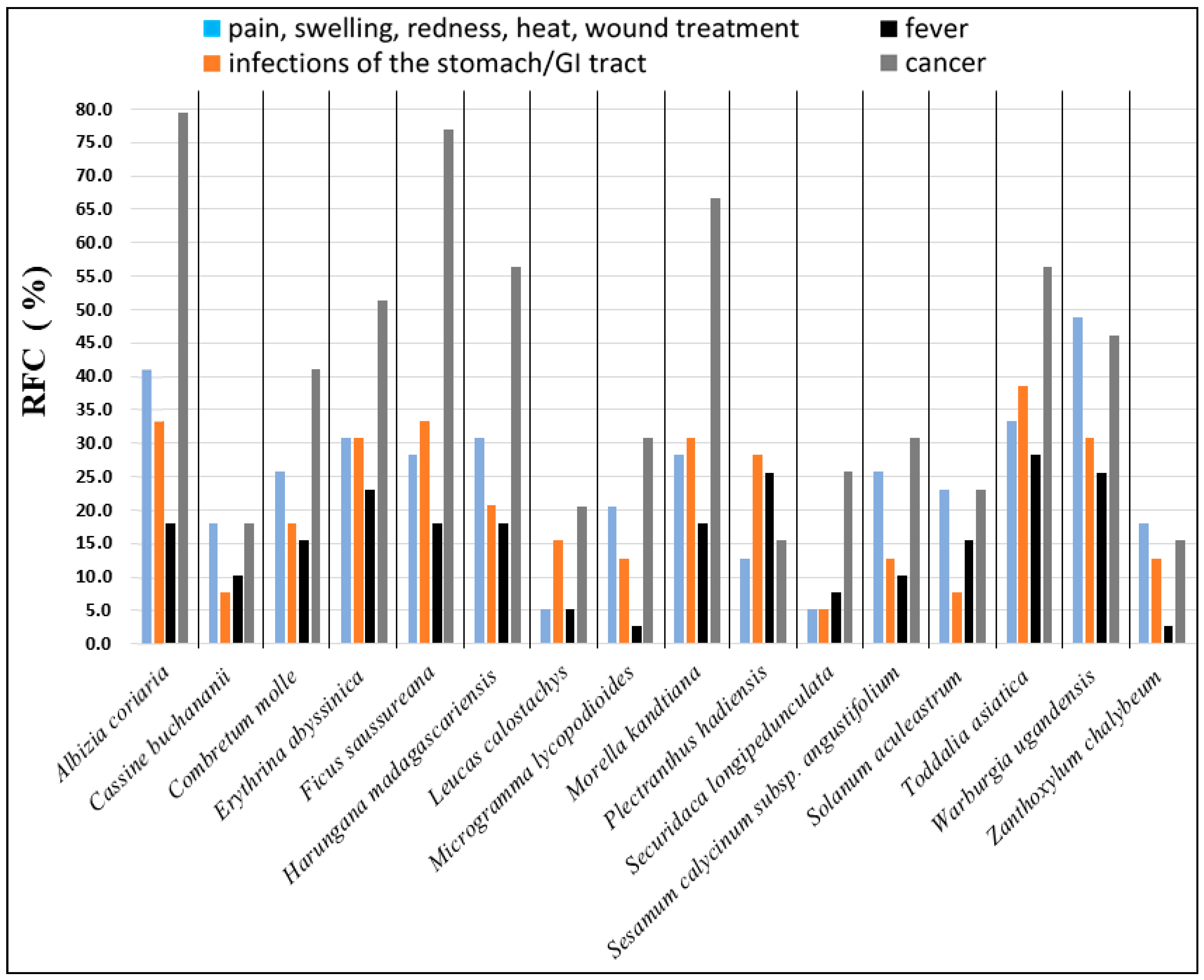
2.1. Information on Plant Species and Extractions
2.2. Selective COX-2 Inhibition Library Screen
2.3. Dose-Response COX-2 Inhibition Experiments
2.4. COX-1 Inhibition Analysis and Selectivity Ratio Determination
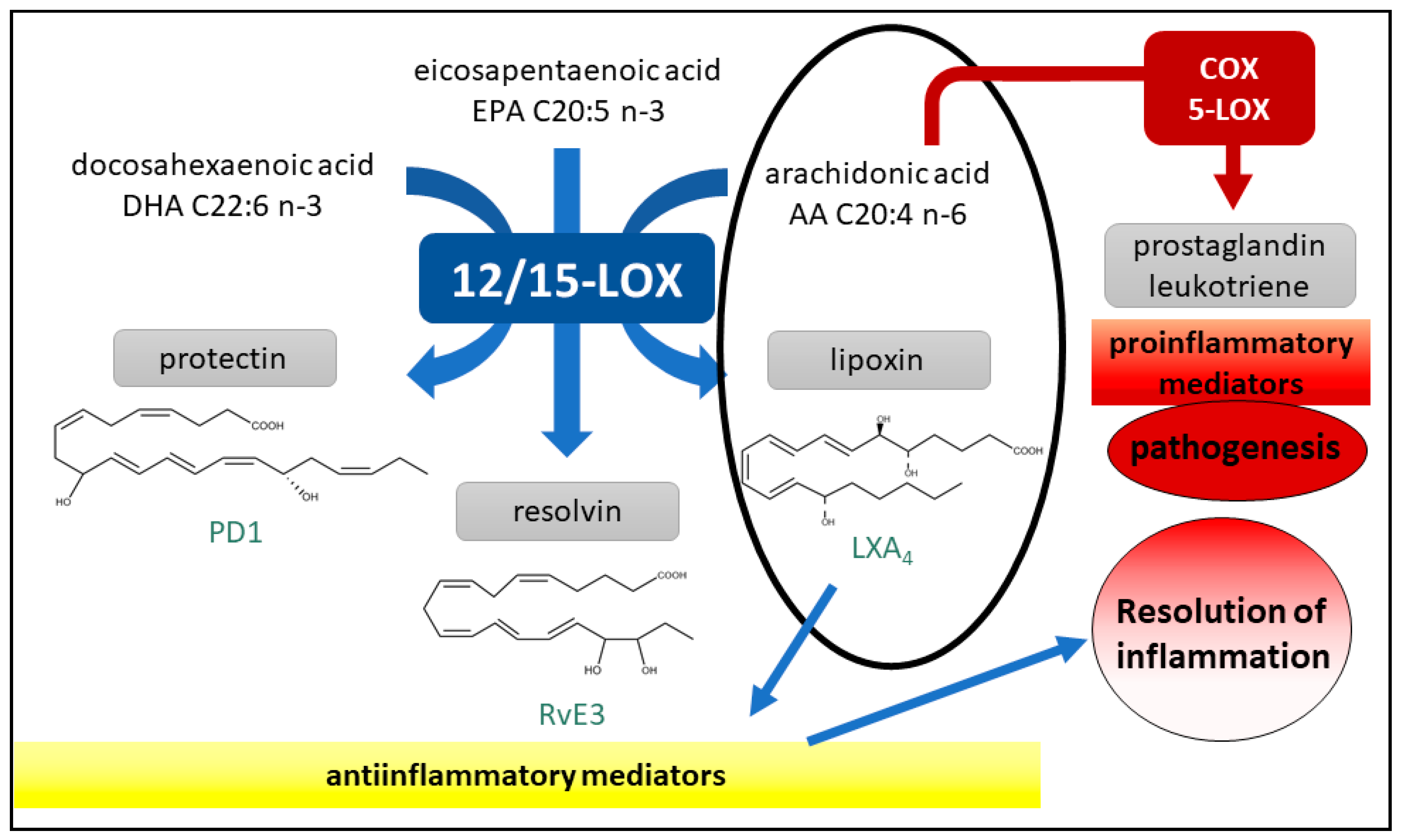
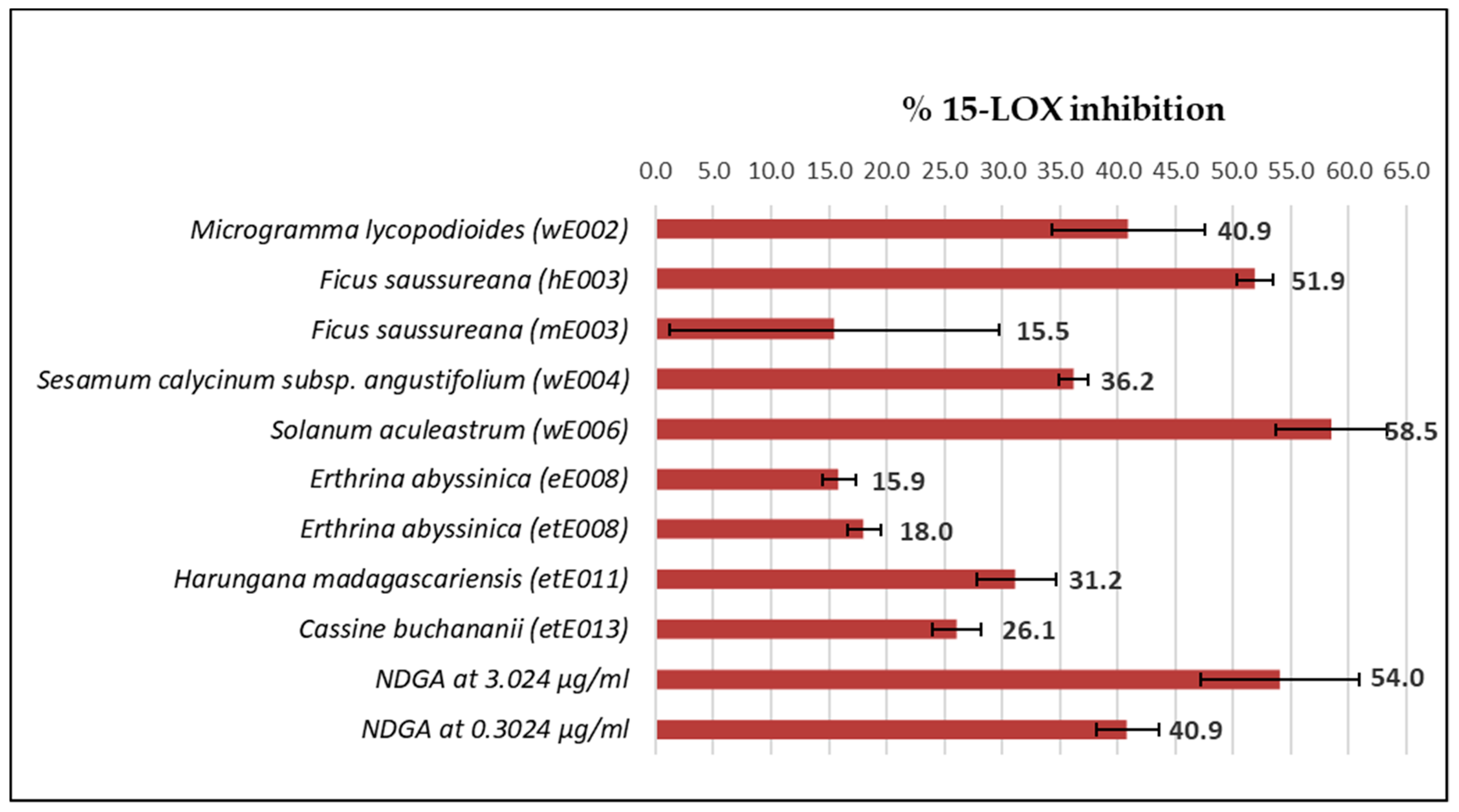
2.5. 15-LOX Inhibition Counterscreen
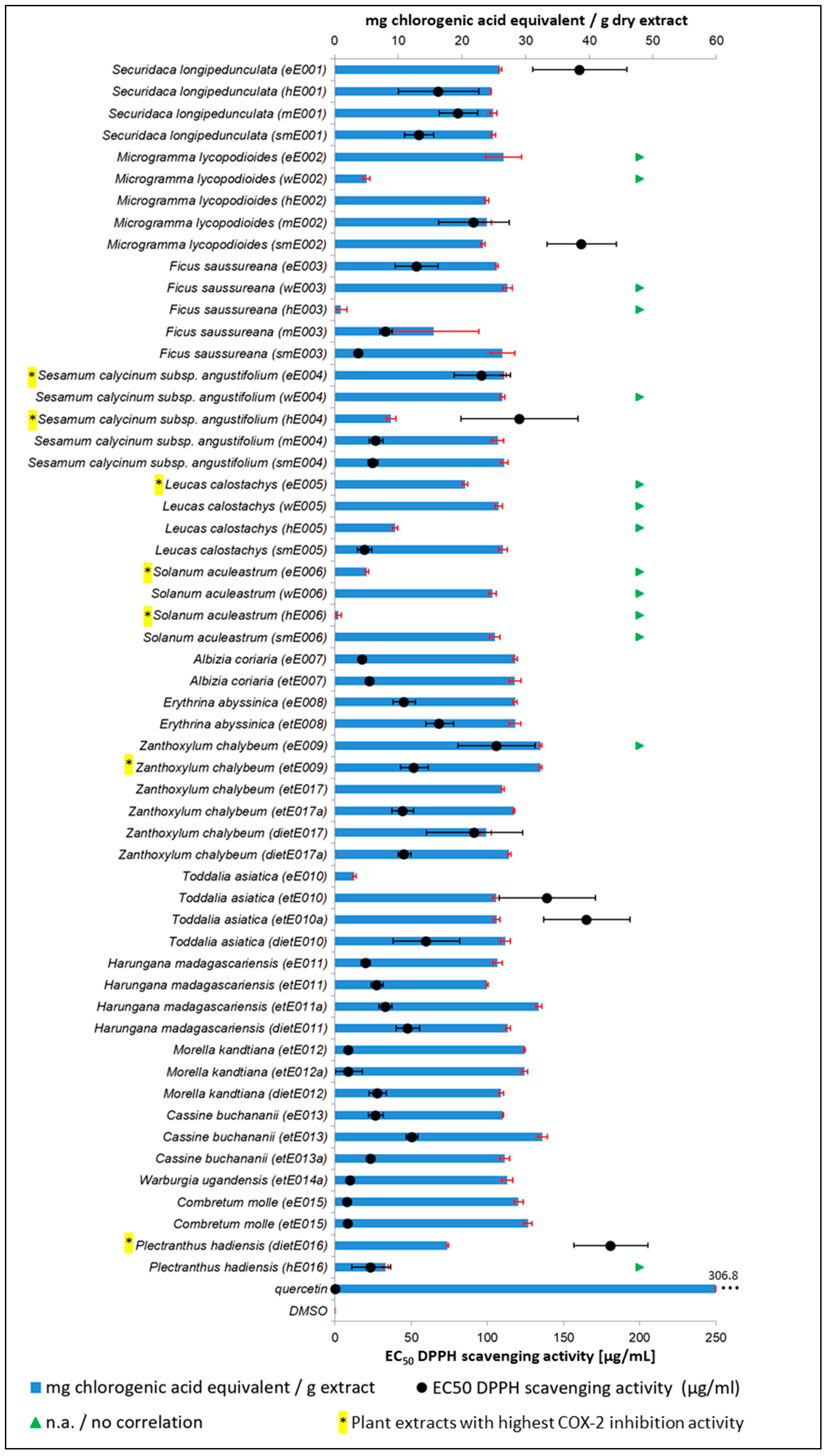
2.6. DPPH Assay for Antioxidant Activity and TPC Determination
2.7. Antibacterial Resazurin Bioassay
3. Discussion and Conclusions
- extract hE005-18 (L. calostachys, MICUAMS-1: >256 µg/mL, MIC25923: 500 µg/mL);
- extract hE006 (S. aculeastrum, MICUAMS-1: 128 µg/mL, MIC25923: 125 µg/mL);
- extract etE008 (E. abyssinica, MICUAMS-1: 64 µg/mL, MIC25923: 63 µg/mL);
- extract etE011-18 (H. madagascariensis, MICUAMS-1: 32 µg/mL, MIC25923: 31 µg/mL);
- extract hE011-18 (H. madagascariensis, MICUAMS-1: 32 µg/mL, MIC25923: 31 µg/mL); and
- extract etE013 (C. buchananii, MICUAMS-1: >256 µg/mL, MIC25923: 500 µg/mL).
- extract dietE011 (H. madagascariensis, MICUAMS-1: 32 µg/mL, MIC25923: 13 µg/mL);
- extract dietE017a (Z. chalybeum, MICUAMS-1: 32 µg/mL, MIC25923: 13 µg/mL);
- extract dietE014-18 (W. ugandensis, MICUAMS-1: 64 µg/mL, MIC25923: 31 µg/mL); and
- extract hE014-18 (W. ugandensis, MICUAMS-1: 64 µg/mL, MIC25923: 31 µg/mL).
4. Materials and Methods
4.1. Ethnobotanical Data
4.2. Collection and Identification of Plant Material
4.3. Extractions
4.4. Sample Preparation
4.5. COX-1/2 Inhibition Screening Assays
4.6. 15-LOX Inhibition Assay
4.7. DPPH Assay
4.8. TPC Determination
4.9. Bacterial Strains
4.10. Resazurin Bioassay
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ghorbanpour, M.; Varma, A. Medicinal Plants and Environmental Challenges; Springer Publishing: Berlin, Germany, 2017. [Google Scholar]
- Ekor, M. The growing use of herbal medicines: Issues relating to adverse reactions and challenges in monitoring safety. Front. Pharm. 2014, 4, 177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- WHO. WHO Traditional Medicine Strategie 2014–2023; WHO: Geneva, Switzerland, 2013. [Google Scholar]
- Tugume, P.; Nyakoojo, C. Ethno-pharmacological survey of herbal remedies used in the treatment of paediatric diseases in Buhunga parish, Rukungiri District, Uganda. BMC Complement. Altern. Med. 2019, 19, 353. [Google Scholar] [CrossRef] [Green Version]
- Schultz, F.; Anywar, G.; Wack, B.; Quave, C.L.; Garbe, L.-A. Ethnobotanical study of selected medicinal plants traditionally used in the rural Greater Mpigi region of Uganda. J. Ethnopharmacol. 2020, 256, 112742. [Google Scholar] [CrossRef] [PubMed]
- Schultz, F.; Anywar, G.; Quave, C.L.; Garbe, L.-A. A Bibliographic Assessment Using the Degrees of Publication Method: Medicinal Plants from the Rural Greater Mpigi Region (Uganda). Evid. Based Complement. Alternat. Med. 2021, 6661565. [Google Scholar]
- Ricciotti, E.; FitzGerald, G.A. Prostaglandins and inflammation. Arterioscler. Thromb. Vasc. Biol. 2011, 31, 986–1000. [Google Scholar] [CrossRef]
- George, A.; Chinnappan, S.; Chintamaneni, M.; Kotak, C.V.; Choudhary, Y.; Kueper, T.; Radhakrishnan, A.K. Anti-inflammatory effects of Polygonum minus (Huds) extract (Lineminus) in in-vitro enzyme assays and carrageenan induced paw edema. BMC Complement. Altern. Med. 2014, 14, 355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tarselli, M.A.; Raehal, K.M.; Brasher, A.K.; Streicher, J.M.; Groer, C.E.; Cameron, M.D.; Bohn, L.M.; Micalizio, G.C. Synthesis of conolidine, a potent non-opioid analgesic for tonic and persistent pain. Nat. Chem. 2011, 3, 449–453. [Google Scholar] [CrossRef] [PubMed]
- Davis, K.D.; Aghaeepour, N.; Ahn, A.H.; Angst, M.S.; Borsook, D.; Brenton, A.; Burczynski, M.E.; Crean, C.; Edwards, R.; Gaudilliere, B.; et al. Discovery and validation of biomarkers to aid the development of safe and effective pain therapeutics: Challenges and opportunities. Nat. Rev. Neurol. 2020, 16, 381–400. [Google Scholar] [CrossRef] [PubMed]
- Stuhlmüller, B.; Ungethüm, U.; Scholze, S.; Martinez, L.; Backhaus, M.; Kraetsch, H.-G.; Kinne, R.W.; Burmester, G.-R. Identification of known and novel genes in activated monocytes from patients with rheumatoid arthritis. Arthritis Rheum. 2000, 43, 775–790. [Google Scholar] [CrossRef]
- Michaëlsson, E.; Holmdahl, M.; Engström, A.; Burkhardt, H.; Scheynius, A.; Holmdahl, R. Macrophages, but not dendritic cells, present collagen to T cells. Eur. J. Immunol. 1995, 25, 2234–2241. [Google Scholar] [CrossRef] [PubMed]
- Williams, C.S.; Mann, M.; DuBois, R.N. The role of cyclooxygenases in inflammation, cancer, and development. Oncogene 1999, 18, 7908–7916. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Limongelli, V.; Bonomi, M.; Marinelli, L.; Gervasio, F.L.; Cavalli, A.; Novellino, E.; Parrinello, M. Molecular basis of cyclooxygenase enzymes (COXs) selective inhibition. Proc. Natl. Acad. Sci. USA 2010, 107, 5411–5416. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, H.; Yu, R.; Choi, Y.; Ma, Z.Z.; Zhang, H.; Xiang, W.; Lee, D.Y.; Berman, B.M.; Moudgil, K.D.; Fong, H.H.; et al. Discovery of cyclooxygenase inhibitors from medicinal plants used to treat inflammation. Pharmacol. Res. 2010, 61, 519–524. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gorwitz, R.J. A Review of Community-Associated Methicillin-Resistant Staphylococcus aureus Skin and Soft Tissue Infections. Pediatric Infect. Dis. J. 2008, 27, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Shaikh, R.U.; Pund, M.M.; Gacche, R.N. Evaluation of anti-inflammatory activity of selected medicinal plants used in Indian traditional medication system in vitro as well as in vivo. J. Tradit. Complementary Med. 2016, 6, 355–361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kurumbail, R.G.; Kiefer, J.R.; Marnett, L.J. Cyclooxygenase enzymes: Catalysis and inhibition. Curr. Opin. Struct. Biol. 2001, 11, 752–760. [Google Scholar] [CrossRef]
- Zarghi, A.; Arfaei, S. Selective COX-2 Inhibitors: A Review of Their Structure-Activity Relationships. Iran J. Pharm. Res. 2011, 10, 655–683. [Google Scholar] [PubMed]
- Botting, R.M. Inhibitors of cyclooxygenases: Mechanisms, selectivity and uses. J. Physiol. Pharmacol. 2006, 57 (Suppl. S5), 113–124. [Google Scholar]
- Konturek, P.C.; Kania, J.; Burnat, G.; Hahn, E.G.; Konturek, S.J. Prostaglandins as mediators of COX-2 derived carcinogenesis in gastrointestinal tract. J. Physiol. Pharmacol. 2005, 56 (Suppl. S5), 57–73. [Google Scholar] [PubMed]
- Hashemi Goradel, N.; Najafi, M.; Salehi, E.; Farhood, B.; Mortezaee, K. Cyclooxygenase-2 in cancer: A review. J. Cell. Physiol. 2019, 234, 5683–5699. [Google Scholar] [CrossRef] [PubMed]
- Young, J.L.; Jazaeri, A.A.; Darus, C.J.; Modesitt, S.C. Cyclooxygenase-2 in cervical neoplasia: A review. Gynecol. Oncol. 2008, 109, 140–145. [Google Scholar] [CrossRef]
- Menter, D.G.; Schilsky, R.L.; DuBois, R.N. Cyclooxygenase-2 and Cancer Treatment: Understanding the Risk Should Be Worth the Reward. Clin. Cancer Res. 2010, 16, 1384–1390. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sheng, J.; Sun, H.; Yu, F.-B.; Li, B.; Zhang, Y.; Zhu, Y.-T. The Role of Cyclooxygenase-2 in Colorectal Cancer. Int. J. Med. Sci. 2020, 17, 1095–1101. [Google Scholar] [CrossRef] [PubMed]
- Saba, N.F.; Choi, M.; Muller, S.; Shin, H.J.C.; Tighiouart, M.; Papadimitrakopoulou, V.A.; El-Naggar, A.K.; Khuri, F.R.; Chen, Z.G.; Shin, D.M. Role of cyclooxygenase-2 in tumor progression and survival of head and neck squamous cell carcinoma. Cancer Prev. Res. 2009, 2, 823–829. [Google Scholar] [CrossRef] [Green Version]
- Kuźbicki, Ł.; Brożyna, A.A. Expression of Cyclooxygenase-2 in Human Epithelial Skin Lesions: A Systematic Review of Immunohistochemical Studies. Appl. Immunohistochem. Mol. Morphol. 2020. [Google Scholar] [CrossRef]
- AlFadly, E.D.; Elzahhar, P.A.; Tramarin, A.; Elkazaz, S.; Shaltout, H.; Abu-Serie, M.M.; Janockova, J.; Soukup, O.; Ghareeb, D.A.; El-Yazbi, A.F.; et al. Tackling neuroinflammation and cholinergic deficit in Alzheimer’s disease: Multi-target inhibitors of cholinesterases, cyclooxygenase-2 and 15-lipoxygenase. Eur. J. Med. Chem. 2019, 167, 161–186. [Google Scholar] [CrossRef] [PubMed]
- Bitto, A.; Giuliani, D.; Pallio, G.; Irrera, N.; Vandini, E.; Canalini, F.; Zaffe, D.; Ottani, A.; Minutoli, L.; Rinaldi, M.; et al. Effects of COX1-2/5-LOX blockade in Alzheimer transgenic 3xTg-AD mice. Inflamm. Res. 2017, 66, 389–398. [Google Scholar] [CrossRef]
- Mattmiller, S.A.; Carlson, B.A.; Sordillo, L.M. Regulation of inflammation by selenium and selenoproteins: Impact on eicosanoid biosynthesis. J. Nutr. Sci. 2013, 2, e28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amtul, Z.; Attaur, R. Chapter 11—Nutraceuticals Neuroprotect Naturally: Alzheimer’s disease, Parkinson’s disease, Stroke and Major Depressive Disorder. In Studies in Natural Products Chemistry; Attaur, R., Ed.; Elsevier: Amsterdam, The Netherlands, 2016; Volume 50, pp. 373–397. [Google Scholar]
- Schneider, C.; Pozzi, A. Cyclooxygenases and lipoxygenases in cancer. Cancer Metastasis Rev. 2011, 30, 277–294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Green, G.A. Understanding NSAIDs: From aspirin to COX-2. Clin. Cornerstone 2001, 3, 50–59. [Google Scholar] [CrossRef]
- Crofford, L.J. Use of NSAIDs in treating patients with arthritis. Arthritis Res. Ther. 2013, 15, S2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pirlamarla, P.; Bond, R.M. FDA labeling of NSAIDs: Review of nonsteroidal anti-inflammatory drugs in cardiovascular disease. Trends Cardiovasc. Med. 2016, 26, 675–680. [Google Scholar] [CrossRef] [PubMed]
- Varrassi, G.; Pergolizzi, J.V.; Dowling, P.; Paladini, A. Ibuprofen Safety at the Golden Anniversary: Are all NSAIDs the Same? A Narrative Review. Adv. Ther. 2020, 37, 61–82. [Google Scholar] [CrossRef] [Green Version]
- Wongrakpanich, S.; Wongrakpanich, A.; Melhado, K.; Rangaswami, J. A Comprehensive Review of Non-Steroidal Anti-Inflammatory Drug Use in The Elderly. Aging Dis. 2018, 9, 143–150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ho, K.Y.; Gwee, K.A.; Cheng, Y.K.; Yoon, K.H.; Hee, H.T.; Omar, A.R. Nonsteroidal anti-inflammatory drugs in chronic pain: Implications of new data for clinical practice. J. Pain Res. 2018, 11, 1937–1948. [Google Scholar] [CrossRef] [Green Version]
- Mitchell, J.A.; Warner, T.D. Cyclo-oxygenase-2: Pharmacology, physiology, biochemistry and relevance to NSAID therapy. Br. J. Pharmacol. 1999, 128, 1121–1132. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, J.A.; Akarasereenont, P.; Thiemermann, C.; Flower, R.J.; Vane, J.R. Selectivity of nonsteroidal antiinflammatory drugs as inhibitors of constitutive and inducible cyclooxygenase. Proc. Natl. Acad. Sci. USA 1993, 90, 11693–11697. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mitchell, J.A.; Evans, T.W. Cyclooxygenase-2 as a therapeutic target. Inflamm. Res. 1998, 47 (Suppl. S2), S88–S92. [Google Scholar] [CrossRef] [PubMed]
- Naesdal, J.; Brown, K. NSAID-associated adverse effects and acid control aids to prevent them: A review of current treatment options. Drug Saf. 2006, 29, 119–132. [Google Scholar] [CrossRef] [PubMed]
- Solomon, S.D. Cyclooxygenase-2 inhibitors and cardiovascular risk. Curr. Opin. Cardiol. 2006, 21, 613–617. [Google Scholar] [CrossRef] [PubMed]
- Chrubasik, J.E.; Roufogalis, B.D.; Chrubasik, S. Evidence of effectiveness of herbal antiinflammatory drugs in the treatment of painful osteoarthritis and chronic low back pain. Phytother. Res. 2007, 21, 675–683. [Google Scholar] [CrossRef] [PubMed]
- Ghasemian, M.; Owlia, S.; Owlia, M.B. Review of Anti-Inflammatory Herbal Medicines. Adv. Pharmacol. Sci. 2016, 2016, 9130979. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taylor, J.L.S.; van Staden, J.; Jäger, A.K. COX-1 and COX-2 inhibitory activity in extracts prepared from Eucomis species, with further reference to extracts from E. autumnalis autumnalis. S. A. J. Botany 2002, 68, 80–85. [Google Scholar]
- Snodgrass, R.G.; Brüne, B. Regulation and Functions of 15-Lipoxygenases in Human Macrophages. Front. Pharmacol. 2019, 10, 719. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arita, M. Eosinophil polyunsaturated fatty acid metabolism and its potential control of inflammation and allergy. Allergol. Int. 2016, 65, S2–S5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gaffney, B.J. Lipoxygenases: Structural principles and spectroscopy. Annu. Rev. Biophys. Biomol. Struct. 1996, 25, 431–459. [Google Scholar] [CrossRef]
- Yamamoto, S. Mammalian lipoxygenases: Molecular structures and functions. Biochim. Biophys. Acta 1992, 1128, 117–131. [Google Scholar] [CrossRef]
- Kuhn, H.; Humeniuk, L.; Kozlov, N.; Roigas, S.; Adel, S.; Heydeck, D. The evolutionary hypothesis of reaction specificity of mammalian ALOX15 orthologs. Prog. Lipid Res. 2018, 72, 55–74. [Google Scholar] [CrossRef]
- Chedea, V.; Jisaka, M. Inhibition of Soybean Lipoxygenases Structural and Activity Models for the Lipoxygenase Isoenzymes Family. In Recent Trends for Enhancing the Diversity and Quality of Soybean Products; IntechOpen Limited: London, UK, 2011. [Google Scholar]
- Serhan, C.N.; Brain, S.D.; Buckley, C.D.; Gilroy, D.W.; Haslett, C.; O’Neill, L.A.J.; Perretti, M.; Rossi, A.G.; Wallace, J.L. Resolution of inflammation: State of the art, definitions and terms. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2007, 21, 325–332. [Google Scholar] [CrossRef]
- Golan, D.E.; Armstrong, E.; Armstrong, A. Principles of Pharmacology: The Pathophysiologic Basis of Drug Therapy, 4th ed.; Lippincott Williams & Wilkins: Philadelphia, PA, USA, 2016; pp. 1–1024. [Google Scholar]
- Bhattacharya, S.; Mathew, G.; Jayne, D.G.; Pelengaris, S.; Khan, M. 15-Lipoxygenase-1 in Colorectal Cancer: A Review. Tumour Biol. 2009, 30, 185–199. [Google Scholar] [CrossRef] [PubMed]
- Pidgeon, G.P.; Lysaght, J.; Krishnamoorthy, S.; Reynolds, J.V.; O’Byrne, K.; Nie, D.; Honn, K.V. Lipoxygenase metabolism: Roles in tumor progression and survival. Cancer Metastasis Rev. 2007, 26, 503–524. [Google Scholar] [CrossRef]
- Orafaie, A.; Matin, M.M.; Sadeghian, H. The importance of 15-lipoxygenase inhibitors in cancer treatment. Cancer Metastasis Rev. 2018, 37, 397–408. [Google Scholar] [CrossRef] [PubMed]
- Barooni, A.B.; Ghorbani, M.; Salimi, V.; Alimohammadi, A.; Khamseh, M.E.; Akbari, H.; Imani, M.; Nourbakhsh, M.; Sheikhi, A.; Shirian, F.I.; et al. Up-regulation of 15-lipoxygenase enzymes and products in functional and non-functional pituitary adenomas. Lipids Health Dis. 2019, 18, 152. [Google Scholar] [CrossRef] [Green Version]
- Il Lee, S.; Zuo, X.; Shureiqi, I. 15-Lipoxygenase-1 as a tumor suppressor gene in colon cancer: Is the verdict in? Cancer Metastasis Rev. 2011, 30, 481–491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rouzer, C.A.; Marnett, L.J. Cyclooxygenases: Structural and functional insights. J. Lipid Res. 2009, 50, S29–S34. [Google Scholar] [CrossRef] [Green Version]
- Knights, K.M.; Mangoni, A.A.; Miners, J.O. Defining the COX inhibitor selectivity of NSAIDs: Implications for understanding toxicity. Expert Rev. Clin. Pharmacol. 2010, 3, 769–776. [Google Scholar] [CrossRef] [PubMed]
- Hawash, M.; Jaradat, N.; Hameedi, S.; Mousa, A. Design, synthesis and biological evaluation of novel benzodioxole derivatives as COX inhibitors and cytotoxic agents. BMC Chem. 2020, 14, 54. [Google Scholar] [CrossRef] [PubMed]
- Soberón, J.R.; Sgariglia, M.A.; Sampietro, D.A.; Quiroga, E.N.; Vattuone, M.A. Free radical scavenging activities and inhibition of inflammatory enzymes of phenolics isolated from Tripodanthus acutifolius. J. Ethnopharmacol. 2010, 130, 329–333. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.H.; Yoon, K.D.; Chin, Y.-W.; Park, J.H.; Kim, J. Phenolic compounds with radical scavenging and cyclooxygenase-2 (COX-2) inhibitory activities from Dioscorea opposita. Bioorg. Med. Chem. 2009, 17, 2689–2694. [Google Scholar] [CrossRef]
- Schultz, F.; Anywar, G.; Tang, H.; Chassagne, F.; Lyles, J.T.; Garbe, L.A.; Quave, C.L. Targeting ESKAPE pathogens with anti-infective medicinal plants from the Greater Mpigi region in Uganda. Nat. Sci. Rep. 2020, 10, 11935. [Google Scholar] [CrossRef]
- Jeruto, P.; Nyangacha, R.; Mutai, C. In Vitro and In Vivo Antiplasmodial Activity of Extracts of Selected Kenyan Medicinal Plants. Afr. J. Pharm. Pharmacol. 2015, 9, 505. [Google Scholar]
- Nyambati, G.; Osiemo, Z.; Maranga, R.; Samuel, M.A.; Ozwara, H. In Vitro Antiplasmodial Activity of Rubia Cardifolia, Harizonnia Abyssinica, Leucas Calostachys Olive, Sanchus Schweinfurthii Medicinal Plants. J. Appl. Pharm. Sci. 2013, 3, 57–62. [Google Scholar]
- Muregi, F.W.; Chhabra, S.C.; Njagi, E.N.; Lang’at-Thoruwa, C.C.; Njue, W.M.; Orago, A.S.; Omar, S.A.; Ndiege, I.O. Anti-plasmodial activity of some Kenyan medicinal plant extracts singly and in combination with chloroquine. Phytother. Res. 2004, 18, 379–384. [Google Scholar] [CrossRef]
- Kigen, G.; Some, F.; Kibosia, J.; Rono, H.; Kiprop, E.; Wanjohi, B.; Kigen, P.; Kipkore, W. Ethnomedicinal Plants Traditionally Used by the Keiyo Community in Elgeyo Marakwet County, Kenya. J. Biodivers. Biopros. Dev. 2014, 1, 132. [Google Scholar]
- Kimutai, N.; Ariya, O.; Mutai, C.; Jeruto, P. Ethnobotanical study of selected medicinal plants used against bacterial infections in Nandi county, Kenya. J. oMed. Plants Stud. 2019, 7, 103–108. [Google Scholar]
- Douglas, K. Review of Traditionally Used Medicinal Plants by the Kipsigis Community in Kenya. Br. J. Pharm. Res. 2016, 12, 1–6. [Google Scholar] [CrossRef]
- Jeruto, P.; Lukhoba, C.; Ouma, G.; Otieno, D.; Mutai, C. An ethnobotanical study of medicinal plants used by the Nandi people in Kenya. J. Ethnopharmacol. 2008, 116, 370–376. [Google Scholar] [CrossRef] [PubMed]
- Mukungu, N.; Abuga, K.; Okalebo, F.; Ingwela, R.; Mwangi, J. Medicinal plants used for management of malaria among the Luhya community of Kakamega East sub-County, Kenya. J. Ethnopharmacol. 2016, 194, 98–107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johns, T.; Faubert, G.M.; Kokwaro, J.O.; Mahunnah, R.L.A.; Kimanani, E.K. Anti-giardial activity of gastrointestinal remedies of the Luo of East Africa. J. Ethnopharmacol. 1995, 46, 17–23. [Google Scholar] [CrossRef]
- Analo, C. Local indigenous knowledge about some medicinal plants in and around Kakamega forest in western Kenya. F1000Research 2012, 1. [Google Scholar] [CrossRef]
- Okach, D.O.; Nyunja, A.R.O.; Opande, G. Phytochemical screening of some wild plants from Lamiaceae and their role in traditional medicine in Uriri District—Kenya. Int. J. Herb. Med. 2013, 5, 135–143. [Google Scholar]
- Kigen, G.; Maritim, A.; Some, F.; Kibosia, J.; Rono, H.; Chepkwony, S.; Kipkore, W.; Wanjohi, B. Ethnopharmacological survey of the medicinal plants used in Tindiret, Nandi County, Kenya. Afr. J. Tradit. Complementary Altern. Med. 2016, 13, 156. [Google Scholar] [CrossRef] [Green Version]
- Jaeger, P.M.L. Systematic Studies in the Genus Solanum in Africa. Ph.D. Thesis, University of Birmingham, Birmingham, UK, 1985. [Google Scholar]
- Priya, T.T.; Sabu, M.C.; Jolly, C.I. Free radical scavenging and anti-inflammatory properties of Lagerstroemia speciosa (L). Inflammopharmacology 2008, 16, 182–187. [Google Scholar] [CrossRef] [PubMed]
- Schinella, G.R.; Tournier, H.A.; Prieto, J.M.; Mordujovich de Buschiazzo, P.; Ríos, J.L. Antioxidant activity of anti-inflammatory plant extracts. Life Sci. 2002, 70, 1023–1033. [Google Scholar] [CrossRef]
- Eom, T.; Kim, E.; Kim, J.S. In Vitro Antioxidant, Antiinflammation, and Anticancer Activities and Anthraquinone Content from Rumex crispus Root Extract and Fractions. Antioxidants 2020, 9, 726. [Google Scholar] [CrossRef] [PubMed]
- Allegra, M. Antioxidant and Anti-Inflammatory Properties of Plants Extract. Antioxidants 2019, 8, 549. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koduru, S.; Grierson, D.S.; Aderogba, M.; Eloff, J.; Afolayan, A. Antioxidant Activity of Solanum aculeastrum (Solanaceae) berries. Int. J. Pharmacol. 2006, 2, 262–264. [Google Scholar] [CrossRef] [Green Version]
- Koduru, S.; Grierson, D.S.; Afolayan, A. Antimicrobial Activity of Solanum aculeastrum. Pharm. Biol. 2008, 44, 283–286. [Google Scholar] [CrossRef]
- Steenkamp, V.; Fernandes, A.C.; Van Rensburg, C.E.J. Screening of Venda medicinal plants for antifungal activity against Candida albicans. S. Afr. J. Bot. 2007, 73, 256–258. [Google Scholar] [CrossRef] [Green Version]
- Koduru, S.; Grierson, D.S.; Van de Venter, M.; Afolayan, A. In vitro Antitumour Activity of Solanum aculeastrum Berries on Three Carcinoma Cells. Int. J. Cancer Res. 2006, 2, 397–402. [Google Scholar]
- Aboyade, O.; Yakubu, M.; Grierson, D.; Afolayan, A. Safety evaluation of aqueous extract of unripe berries of Solanum aculeastrum in male Wistar rats. Afr. J. Pharm. Pharmacol. 2010, 4, 90–97. [Google Scholar]
- Aboyade, O.; Yakubu, M.; Grierson, D.; Afolayan, A. Studies on the toxicological effect of the aqueous extract of the fresh, dried and boiled berries of Solanum aculeastrum Dunal in male Wistar rats. Hum. Exp. Toxicol. 2009, 28, 765–775. [Google Scholar] [CrossRef] [PubMed]
- Koduru, S.; Grierson, D.S.; Van de Venter, M.; Afolayan, A. Anticancer Activity of Steroid Alkaloids Isolated from Solanum aculeastrum. Pharm. Biol. 2008, 45, 613–618. [Google Scholar] [CrossRef]
- Koduru, S.; Jimoh, F.; Grierson, D.S.; Afolayan, A. Antioxidant Activity of two Steroid Alkaloids extracted from Solanum aculeastrum. J. Pharmacol. Toxicol. 2007, 2, 160–167. [Google Scholar] [CrossRef] [Green Version]
- Wanyonyi, A.W.; Chhabra, S.C.; Mkoji, G.; Eilert, U.; Njue, W.M. Bioactive steroidal alkaloid glycosides from Solanum aculeastrum. Phytochemistry 2002, 59, 79–84. [Google Scholar] [CrossRef]
- Wanyonyi, A.W.; Chhabra, S.C.; Mkoji, G.; Njue, W.; Tarus, P.K. Molluscicidal and antimicrobial activity of Solanum aculeastrum. Fitoterapia 2003, 74, 298–301. [Google Scholar] [CrossRef]
- Burger, T.; Mokoka, T.; Fouche, G.; Steenkamp, P.; Steenkamp, V.; Cordier, W. Solamargine, a bioactive steroidal alkaloid isolated from Solanum aculeastrum induces non-selective cytotoxicity and P-glycoprotein inhibition. BMC Complement. Altern. Med. 2018, 18, 137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ochwang’i, D.O.; Kimwele, C.N.; Oduma, J.A.; Gathumbi, P.K.; Mbaria, J.M.; Kiama, S.G. Medicinal plants used in treatment and management of cancer in Kakamega County, Kenya. J. Ethnopharmacol. 2014, 151, 1040–1055. [Google Scholar] [CrossRef]
- Koduru, S.; Grierson, D.; Afolayan, A. Ethnobotanical information of medicinal plants used for treatment of cancer in the Eastern Cape Province, South Africa. Curr. Sci. 2007, 92, 906–908. [Google Scholar]
- Mongalo, N.I.; Makhafola, T.J. Ethnobotanical knowledge of the lay people of Blouberg area (Pedi tribe), Limpopo Province, South Africa. J. Ethnobiol. Ethnomed. 2018, 14, 46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mogale, M.M.P.; Raimondo, D.C.; VanWyk, B.E. The ethnobotany of Central Sekhukhuneland, South Africa. S. Afr. J. Bot. 2019, 122, 90–119. [Google Scholar] [CrossRef]
- Komoreng, L.; Thekisoe, O.; Lehasa, S.; Tiwani, T.; Mzizi, N.; Mokoena, N.; Khambule, N.; Ndebele, S.; Mdletshe, N. An ethnobotanical survey of traditional medicinal plants used against lymphatic filariasis in South Africa. S. Afr. J. Bot. 2017, 111, 12–16. [Google Scholar] [CrossRef]
- Ihlenfeldt, H.D. Flora Zambesiaca—FZ Volume:8 Part:3. Available online: http://apps.kew.org/efloras/namedetail.do?qry=namelist&flora=fz&taxon=6482&nameid=16417 (accessed on 15 August 2020).
- Chhabra, S.C.; Mahunnah, R.L.A. Plants used in traditional medicine by hayas of the Kagera region, Tanzania. Econ. Bot. 1994, 48, 121–129. [Google Scholar] [CrossRef]
- Kibuuka, M.S.; Anywar, G. Medicinal Plant Species Used in the Management of Hernia by Traditional Medicine Practitioners in Central Uganda. Ethnobot. Res. Appl. 2015, 14, 289–298. [Google Scholar] [CrossRef] [Green Version]
- Hamill, F.A.; Apio, S.; Mubiru, N.K.; Bukenya-Ziraba, R.; Mosango, M.; Maganyi, O.W.; Soejarto, D.D. Traditional herbal drugs of Southern Uganda, II: Literature analysis and antimicrobial assays. J. Ethnopharmacol. 2003, 84, 57–78. [Google Scholar] [CrossRef]
- Tabuti, J.R.; Lye, K.A.; Dhillion, S.S. Traditional herbal drugs of Bulamogi, Uganda: Plants, use and administration. J. Ethnopharmacol. 2003, 88, 19–44. [Google Scholar] [CrossRef]
- Zargoosh, Z.; Ghavam, M.; Bacchetta, G.; Tavili, A. Effects of ecological factors on the antioxidant potential and total phenol content of Scrophularia striata Boiss. Sci. Rep. 2019, 9, 16021. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dharani, N.; Yenesew, A. Medicinal Plants of East Africa; Drongo Editing & Publishing: Nairobi, Kenya, 2010. [Google Scholar]
- Katende, A.B.; Birnie, A.; Tengnas, B. Useful Trees and Shrubs for Uganda—Identification, Propagation, and Management for Agricultural and Pastoral Communities; Nairobi (Kenya) Regional Soil Conservation Unit: Nairobi, Kenya, 1995. [Google Scholar]
- Neuwinger, H.D. African Ethnobotany: Poisons and Drugs: Chemistry, Pharmacology, Toxicology; CRC Press: Boca Raton, FL, USA, 1996. [Google Scholar]
- Erinoso, S.; Aworinde, D. Ethnobotanical survey of some medicinal plants used in traditional health care in Abeokuta areas of Ogun State, Nigeria. Afr. J. Pharm. Pharmacol. 2012, 6, 1352–1362. [Google Scholar] [CrossRef]
- Tjeck, O.P.; Souza, A.; Mickala, P.; Lepengue, A.N.; M’Batchi, B. Bio-efficacy of medicinal plants used for the management of diabetes mellitus in Gabon: An ethnopharmacological approach. J. Intercult. Ethnopharmacol. 2017, 6, 206–217. [Google Scholar] [CrossRef]
- Rakotondrafara, A.; Rakotondrajaona, R.; Rakotoarisoa, M.; Ratsimbason, M.; Rasamison, V.; Rakotonandrasana, S.R. Ethnobotany of medicinal plants used by the Zafimaniry clan in Madagascar. J. Phytopharmacol. 2018, 7, 483–494. [Google Scholar]
- Malan, D.F.; Neuba, D.F.; Kouakou, K.L. Medicinal plants and traditional healing practices in Ehotile people, around the Aby Lagoon (eastern littoral of Cote d’Ivoire). J. Ethnobiol. Ethnomed. 2015, 11, 21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nyamukuru, A.; Tabuti, J.R.S.; Lamorde, M.; Kato, B.; Sekagya, Y.; Aduma, P.R. Medicinal plants and traditional treatment practices used in the management of HIV/AIDS clients in Mpigi District, Uganda. J. Herb. Med. 2017, 7, 51–58. [Google Scholar] [CrossRef]
- Philip, K.; Mwangi, E.; Cheplogoi, P.; Samuel, K. Ethnobotanical Survey of Antimalarial Medicinal Plants Used in Butebo County, Eastern Uganda. Eur. J. Med. Plants 2017, 21, 1–22. [Google Scholar] [CrossRef]
- Perry, R.T.; Halsey, N.A. The Clinical Significance of Measles: A Review. J. Infect. Dis. 2004, 189 (Suppl. S1), S4–S16. [Google Scholar] [PubMed] [Green Version]
- Olila, D.; Olwa, O.; Opuda-Asibo, J. Screening extracts of Zanthoxylum chalybeum and Warburgia ugandensis for activity against measles virus (Swartz and Edmonston strains) in vitro. Afr. Health Sci. 2002, 2, 2–10. [Google Scholar] [PubMed]
- Martin, J.G. Ethnobotany: A Methods Manual; Springer: London, UK, 2019. [Google Scholar]
- Alexiades, M.N.; Sheldon, J.W. Selected Guidelines for Ethnobotanical Research: A Field Manual; New York Botanical Garden: The Bronx, NY, USA, 1996; Volume 10. [Google Scholar]
- Heinrich, M.; Lardos, A.; Leonti, M.; Weckerle, C.; Willcox, M.; Applequist, W.; Ladio, A.; Lin Long, C.; Mukherjee, P.; Stafford, G. Best practice in research: Consensus Statement on Ethnopharmacological Field Studies—ConSEFS. J. Ethnopharmacol. 2018, 211, 329–339. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- SERNEC. SouthEast Regional Network of Expertise and Collections. Available online: http://sernecportal.org/portal/index.php (accessed on 25 January 2021).
- Weckerle, C.S.; de Boer, H.J.; Puri, R.K.; van Andel, T.; Bussmann, R.W.; Leonti, M. Recommended standards for conducting and reporting ethnopharmacological field studies. J. Ethnopharmacol. 2018, 210, 125–132. [Google Scholar] [CrossRef] [PubMed]
- The Angiosperm Phylogeny Group. An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG IV. Bot. J. Linn. Soc. 2016, 181, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Cayman COX-2 ((Human) Inhibitor Screening Assay. Available online: https://www.caymanchem.com/pdfs/701080.pdf (accessed on 21 December 2020).
- Cayman Prostaglandin Screening ELISA Kit. Available online: https://www.caymanchem.com/pdfs/514012.pdf (accessed on 21 December 2020).
- GraphPad KNOWLEDGEBASE-ARTICLE #34 Error Propagandation. Available online: https://www.graphpad.com/support/faq/why-does-prism-report-the-standard-error-for-logec50-but-not-for-the-ec50-itself-when-it-fits-dose-response-curves/ (accessed on 15 October 2020).
- Gierlinski, M. Understanding Statistical Errors—A Primer for Biologists; John Wiley & Sons: Chichester, UK, 2016. [Google Scholar]
- Cayman Lipoxygenase Inhibitor Screening Assay Kit Item No. 760700. Available online: https://www.caymanchem.com/pdfs/760700.pdf (accessed on 5 January 2020).
- Alferez, F.; Singh, S.; Umbach, A.L.; Hockema, B.; Burns, J.K. Citrus abscission and Arabidopsis plant decline in response to 5-chloro-3-methyl-4-nitro-1H-pyrazole are mediated by lipid signalling. Plant Cell Environ. 2005, 28, 1436–1449. [Google Scholar] [CrossRef]
- Jacob, J.; Kumar, P. Dual COX/LOX inhibition: Screening and evaluation of effect of medicinal plants of Kerala as Anti-inflammatory agents. J. Pharmacogn. Phytochem. 2015, 3, 62–66. [Google Scholar]
- Muruganantham, N.; Basavaraj, K.H.; Dhanabal, S.P.; Praveen, T.K.; Shamasundar, N.M.; Rao, K.S. Screening of Caesalpinia bonduc leaves for antipsoriatic activity. J. Ethnopharmacol. 2011, 133, 897–901. [Google Scholar] [CrossRef] [PubMed]
- Kumar, P.; Jacob, J.; Augustine, J. Dual Inhibition of Lipoxygenase/Cyclooxygenase by An Ayurvedic Herb, Tinospora cordifolia (WILLD.); Possible Explanation For Anti-inflammatory Activity. J. Pharm. Res. 2011, 4, 1779–1780. [Google Scholar]
- Sarker, S.D.; Nahar, L. An introduction to natural products isolation. In Natural Products Isolation; Springer: Berlin/Heidelberg, Germany, 2012; pp. 1–25. [Google Scholar]
- Takao, T.; Kitatani, F.; Watanabe, N.; Yagi, A.; Sakata, K. A Simple Screening Method for Antioxidants and Isolation of Several Antioxidants Produced by Marine Bacteria from Fish and Shellfish. Biosci. Biotechnol. Biochem. 1994, 58, 1780–1783. [Google Scholar] [CrossRef] [Green Version]
- Kumarasamy, Y.; Fergusson, M.E.; Nahar, L.; Sarker, S.D. Bioactivity of Moschamindole from Centaurea moschata. Pharm. Biol. 2002, 40, 307–310. [Google Scholar] [CrossRef]
- Sánchez-Rangel, J.C.; Benavides, J.; Heredia, J.B.; Cisneros-Zevallos, L.; Jacobo-Velázquez, D.A. The Folin–Ciocalteu assay revisited: Improvement of its specificity for total phenolic content determination. Anal. Methods 2013, 5, 5990–5999. [Google Scholar] [CrossRef]
- Sarker, S.D.; Nahar, L.; Kumarasamy, Y. Microtitre plate-based antibacterial assay incorporating resazurin as an indicator of cell growth, and its application in the in vitro antibacterial screening of phytochemicals. Methods 2007, 42, 321–324. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Scientific Name | Family | Local Name in Luganda | Plant Part | Voucher Specimen Number and Location | Extraction Solvent | Extract ID |
|---|---|---|---|---|---|---|
| Securidaca longipedunculata Fresen. | Polygalaceae | Mukondwe | stem | AG196 (Makerere University herbarium, Uganda) | ethyl acetate 1 | eE001 |
| water 1 | wE001 | |||||
| n-hexane (sox.) 1 | hE001 | |||||
| methanol 1 | mE001 | |||||
| methanol (sox. succ.) 1 | smE001 | |||||
| Microgramma lycopodioides (L.) Copel. | Polypodiaceae | Kukumba | root (rhizomes) | AG639 (Makerere University herbarium, Uganda) | ethyl acetate 1 | eE002 |
| aqueous 1 | wE002 | |||||
| n-hexane (sox.) 1 | hE002 | |||||
| methanol 1 | mE002 | |||||
| methanol 1 (sox. succ.) | smE002 | |||||
| Ficus saussureana DC. | Moraceae | Muwo | stem | AG219 (Makerere University herbarium, Uganda) | ethyl acetate 1 | eE003 |
| aqueous 1 | wE003 | |||||
| n-hexane (sox.) 1 | hE003 | |||||
| methanol 1 | mE003 | |||||
| methanol (sox. succ.) 1 | smE003 | |||||
| Sesamum calycinum subsp. angustifolium (Oliv.) Ihlenf. & Seidenst. | Pedaliaceae | Lutungotungo | leaves | AG205 (Makerere University herbarium, Uganda) 23173 * (Emory University herbarium, USA) | ethyl acetate1 | eE004 |
| water 1 | wE004 | |||||
| n-hexane (sox.) 1 | hE004 | |||||
| methanol 1 | mE004 | |||||
| methanol (sox. succ.) 1 | smE004 | |||||
| ethyl acetate5 | eE004-18 | |||||
| n-hexane (sox.) 5 | hE004-18 | |||||
| Leucas calostachys Oliv. | Lamiaceae | Kakuba musulo | leaves | AG195 (Makerere University herbarium, Uganda) 23175 * (Emory University herbarium, USA) | ethyl acetate 1 | eE005 |
| water 1 | wE005 | |||||
| n-hexane (sox.)1 | hE005 | |||||
| methanol (sox. succ.) 1 | smE005 | |||||
| ethyl acetate 5 | eE005-18 | |||||
| n-hexane (sox.) 5 | hE005-18 | |||||
| methanol 5 | mE005-18 | |||||
| methanol (sox. succ.) 5 | smE005-18 | |||||
| Solanum aculeastrum Dunal | Solanaceae | Kitengo | root | AG193 (Makerere University herbarium, Uganda) | ethyl acetate 1 | eE006 |
| water 1 | wE006 | |||||
| n-hexane (sox.) 1 | hE006 | |||||
| methanol (sox. succ.) 1 | smE006 | |||||
| Albizia coriaria Oliv. | Fabaceae | Mugavu | stem bark | AG203 (Makerere University herbarium, Uganda) | ethyl acetate 2 | eE007 |
| ethanol 2 | etE007 | |||||
| Erythrina abyssinica DC. | Fabaceae | Jjirikiti | stem bark | AG199 (Makerere University herbarium, Uganda) | ethyl acetate 2 | eE008 |
| ethanol 2 | etE008 | |||||
| Zanthoxylum chalybeum Engl. | Rutaceae | Ntaleyaddungu | stem bark | AG204 (Makerere University herbarium, Uganda) | ethyl acetate 2 | eE009 |
| ethanol 2 | etE009 | |||||
| ethanol 3 | etE017 | |||||
| diethyl ether 3 | dietE017 | |||||
| ethanol 4 | etE017a | |||||
| diethyl ether 4 | dietE017a | |||||
| Toddalia asiatica (L.) Lam. | Rutaceae | Kawule | leaves, bark | AG190 (Makerere University herbarium, Uganda) | ethyl acetate 2 | eE010 |
| ethanol2 | etE010 | |||||
| diethyl ether 4 | dietE010 | |||||
| ethanol 4 | etE010a | |||||
| Harungana madagascariensis Lam. ex Poir. | Hypericaceae | Mukabiiransiko | stem bark | AG230 (Makerere University herbarium, Uganda) 23180 * (Emory University herbarium, USA) | ethyl acetate 2 | eE011 |
| ethanol 2 | etE011 | |||||
| diethyl ether 4 | dietE011 | |||||
| ethanol 4 | etE011a | |||||
| ethyl acetate 5 | eE011-18 | |||||
| n-hexane 5 | hE011-18 | |||||
| ethanol 5 | etE011-18 | |||||
| diethyl ether 5 | dietE011-18 | |||||
| Morella kandtiana (Engl.) Verdc. & Polhill | Myricaeae | Mukikimbo | root | AG201 (Makerere University herbarium, Uganda) 23174 * (Emory University herbarium, USA) | ethyl acetate 2 | eE012 |
| ethanol 2 | etE012 | |||||
| ethanol 4 | etE012a | |||||
| diethyl ether 4 | dietE012 | |||||
| ethyl acetate 5 | eE012-18 | |||||
| diethyl ether 5 | dietE012-18 | |||||
| Cassine buchananii Loes. | Celastraceae | Mbaluka | stem bark | AG198 (Makerere University herbarium, Uganda) | ethyl acetate 2 | eE013 |
| ethanol 2 | etE013 | |||||
| ethanol 4 | etE013a | |||||
| Warburgia ugandensis Sprague | Canellaceae | Abasi | stem bark | AG220 (Makerere University herbarium, Uganda) 23181 * (Emory University herbarium, USA) | ethanol 4 | etE014a |
| diethyl ether 4 | dietE014 | |||||
| ethyl acetate 5 | eE014-18 | |||||
| water | wE014-18 | |||||
| diethyl ether 5 | dietE014-18 | |||||
| n-hexane (sox.) 5 | hE014-18 | |||||
| ethanol | etE014-18 | |||||
| Combretum molle R.Br. ex G.Don | Combretaceae | Ndagi | stem bark | AG191 (Makerere University herbarium, Uganda) | ethyl acetate 2 | eE015 |
| ethanol 2 | etE015 | |||||
| Plectranthus hadiensis (Forssk.) Schweinf. ex Sprenger | Lamiaceae | Kibwankulata | leaves | AG210 (Makerere University herbarium, Uganda) | diethyl ether 4 | dietE016 |
| n-hexane 4 | hE016 |
| Extract ID | Plant Species | Type of Extract | IC50 ± SEM | Ratio COX-2 COX-1 | |
|---|---|---|---|---|---|
| COX-2 | COX-1 | ||||
| eE005 | Leucas calostachys | ethyl acetate | 0.66 ± 0.66 | 7.76 ± 1.58 | 0.1 |
| eE006 | Solanum aculeastrum | ethyl acetate | 1.74 ± 0.28 | 9.72 ± 0.28 | 0.2 |
| hE006 | Solanum aculeastrum | n-hexane | 3.19 ± 0.43 | 3.99 ± 3.92 | 0.8 |
| hE004 | Sesamum calycinum subsp. angustifolium | n-hexane | 3.65 ± 0.56 | 8.57 ± 2.03 | 0.4 |
| dietE016 | Plectranthus hadiensis | diethyl ether | 4.55 ± 0.76 | 5.83 ± 3.79 | 0.8 |
| eE004 | Sesamum calycinum subsp. angustifolium | ethyl acetate | 6.05 ± 0.20 | 11.47 ± 2.89 | 0.5 |
| dietE014 | Warburgia ugandensis | diethyl ether | 13.33 ± 4.36 | 11.05 ± 1.43 | 1.2 |
| etE009 | Zanthoxylum chalybeum | ethnanol | 16.07 ± 2.29 | 24.89 ± 4.16 | 0.7 |
| dietE012 | Morella kandtiana | diethyl ether | 17.24 ± 2.79 | 15.01 ± 1.14 | 1.2 |
| positive control | DuP-769 | - (pure compound) | 0.93 ± 0.20 | >100.0 | >0.001 |
| Scientific Name | Extract ID | S. aureus ATCC 25923 | E. coli K12 ATCC 23716 | L. innocua ATCC 33090 | L. monocytogenes ATCC 15313 |
|---|---|---|---|---|---|
| Securidaca longipedunculata | eE001 | 104.17 ± 29.46 | >500 | 500.00 ± 0 | - |
| wE001 | >500 | >500 | >500 | - | |
| hE001 | 125.00 ± 0 | >500 | 250.00 ± 0 | 250.00 ± 0 | |
| mE001 | 83.33 ± 29.46 | >500 | >500 | - | |
| smE001 | 104.17 ± 29.46 | >500 | >500 | - | |
| Microgramma lycopodioides | eE002 | 500.00 ± 0 | >500 | >500 | - |
| wE002 | >500 | >500 | >500 | - | |
| hE002 | - | >500 | >500 | - | |
| mE002 | 250.00 ± 0 | 500.00 ± 0 | 500.00 ± 0 | 250.00 ± 0 | |
| smE002 | 26.04 ± 7.37 | >500 | >500 | - | |
| Ficus saussureana | eE003 | 500 ± 0 | 500.00 ± 0 | >500 | - |
| wE003 | >500 | >500 | >500 | - | |
| hE003 | 500.00 ± 0 | 500.00 ± 0 | >500 | - | |
| mE003 | 20.83 ± 7.37 | 500.00 ± 0 | >500 | - | |
| smE003 | 500 ± 0 | 500.00 ± 0 | >500 | - | |
| Sesamum calycinum subsp. angustifolium | eE004 | 125.00 ± 0 | 500.00 ± 0 | >500 | - |
| wE004 | 500 ± 0 | >500 | >500 | - | |
| hE004 | 125.00 ± 0 | 500.00 ± 0 | >500 | - | |
| mE004 | 250.00 ± 0 | 500.00 ± 0 | >500 | - | |
| smE004 | 250.00 ± 0 | >500 | >500 | - | |
| eE004-18 | 250.00 ± 0 | - | - | - | |
| hE004-18 | 31.25 ± 0 | - | - | - | |
| Leucas calostachys | eE005 | 500.00 ± 0 | >500 | >500 | - |
| wE005 | - | >500 | >500 | - | |
| hE005 | 62.50 ± 0 | 500.00 ± 0 | >500 | - | |
| smE005 | 500 ± 0 | >500 | >500 | - | |
| eE005-18 | 500.00 ± 0 | - | - | - | |
| hE005-18 | 104.17 ± 29.46 | - | - | - | |
| mE005-18 | 500 ± 0 | - | - | - | |
| smE005-18 | 104.17 ± 29.46 | - | - | - | |
| Solanum aculeastrum | eE006 | 500.00 ± 0 | 500.00 ± 0 | >500 | - |
| wE006 | 500.00 ± 0 | >500 | >500 | - | |
| hE006 | 125.00 ± 0 | >500 | >500 | - | |
| smE006 | 11.72 ± 5.52 | >500 | >500 | - | |
| Albizia coriaria | eE007 | 250.00 ± 0 | 250.00 ± 0 | >500 | - |
| etE007 | 500.00 ± 0 | 500.00 ± 0 | >500 | - | |
| Erythrina abyssinica | eE008 | 83.33 ± 29.46 | 500.00 ± 0 | >500 | - |
| etE008 | 62.50 ± 0 | >500 | >500 | - | |
| Zanthoxylum chalybeum | eE009 | 31.25 ± 0 | - | >500 | - |
| etE009 | 500.00 ± 0 | >500 | >500 | - | |
| etE017 | 500.00 ± 0 | >500 | >500 | - | |
| dietE017 | 250.00 ± 0 | >500 | >500 | - | |
| etE017a | >500 | 250.00 ± 0 | - | - | |
| dietE017a | 13.02 ± 3.62 | >500 | - | - | |
| Toddalia asiatica | eE010 | 31.25 ± 0 | >500 | >500 | - |
| etE010 | 31.25 ± 0 | >500 | >500 | - | |
| dietE010 | 20.83 ± 7.37 | >500 | >500 | - | |
| etE010a | 83.33 ± 29.46 | >500 | >500 | - | |
| Harungana madagascariensis | eE011 | 125.00 ± 0 | 250.00 ± 0 | 500.00 ± 0 | >500 |
| etE011 | 57.29 ± 7.37 | 500.00 ± 0 | 500.00 ± 0 | >500 | |
| dietE011 | 13.02 ± 3.68 | 500.00 ± 0 | 41.67 ± 14.73 | 125 ± 0 | |
| etE011a | 125.00 ± 0 | 250.00 ± 0 | 500.00 ± 0 | >500 | |
| eE011-18 | 52.08 ± 14.73 | - | - | - | |
| hE011-18 | 31.25 ± 0 | - | - | - | |
| etE011-18 | 31.25 ± 0 | - | - | - | |
| dietE011-18 | 52.08 ± 14.73 | - | - | - | |
| Morella kandtiana | eE012 | 250.00 ± 0 | - | - | - |
| etE012 | 500.00 ± 0 | 500.00 ± 0 | >500 | - | |
| etE012a | 500.00 ± 0 | 250.00 ± 0 | >500 | - | |
| dietE012 | 250.00 ± 0 | 500.00 ± 0 | >500 | - | |
| eE012-18 | 500.00 ± 0 | - | - | - | |
| dietE012-18 | 500.00 ± 0 | - | - | - | |
| Cassine buchananii | eE013 | 500.00 ± 0 | 500.00 ± 0 | >500 | - |
| etE013 | 500.00 ± 0 | 500.00 ± 0 | >500 | - | |
| etE013a | 500.00 ± 0 | 500.00 ± 0 | >500 | - | |
| Warburgia ugandensis | etE014a | 500.00 ± 0 | 500.00 ± 0 | >500 | - |
| dietE014 | 31.25 ± 0 | 500.00 ± 0 | 500.00 ± 0 | 125 ± 0 | |
| eE014-18 | 31.25 ± 0 | - | - | - | |
| wE014-18 | >500 | - | - | - | |
| dietE014-18 | 31.25 ± 0 | - | - | - | |
| hE014-18 | 31.25 ± 0 | - | - | - | |
| etE014-18 | 41.67 ± 14.73 | - | - | - | |
| Combretum molle | eE015 | 500.00 ± 0 | 250.00 ± 0 | >500 | - |
| etE015 | 500.00 ± 0 | 500.00 ± 0 | >500 | - | |
| Plectranthus hadiensis | dietE016 | 104.17 ± 29.46 | >500 | >500 | - |
| hE016 | 62.50 ± 0 | >500 | >500 | - | |
| ciprofloxacin | - | 0.19 ± 0.06 | 7.81 ± 0 | 0.12 ± 0.00 | 0.12 ± 0.00 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schultz, F.; Osuji, O.F.; Wack, B.; Anywar, G.; Garbe, L.-A. Antiinflammatory Medicinal Plants from the Ugandan Greater Mpigi Region Act as Potent Inhibitors in the COX-2/PGH2 Pathway. Plants 2021, 10, 351. https://doi.org/10.3390/plants10020351
Schultz F, Osuji OF, Wack B, Anywar G, Garbe L-A. Antiinflammatory Medicinal Plants from the Ugandan Greater Mpigi Region Act as Potent Inhibitors in the COX-2/PGH2 Pathway. Plants. 2021; 10(2):351. https://doi.org/10.3390/plants10020351
Chicago/Turabian StyleSchultz, Fabien, Ogechi Favour Osuji, Barbara Wack, Godwin Anywar, and Leif-Alexander Garbe. 2021. "Antiinflammatory Medicinal Plants from the Ugandan Greater Mpigi Region Act as Potent Inhibitors in the COX-2/PGH2 Pathway" Plants 10, no. 2: 351. https://doi.org/10.3390/plants10020351