
Paenibacillus alvei K165 and Fusarium oxysporum F2: Potential Biocontrol Agents against Phaeomoniella chlamydospora in Grapevines

 , , and
, , and {kind=link}
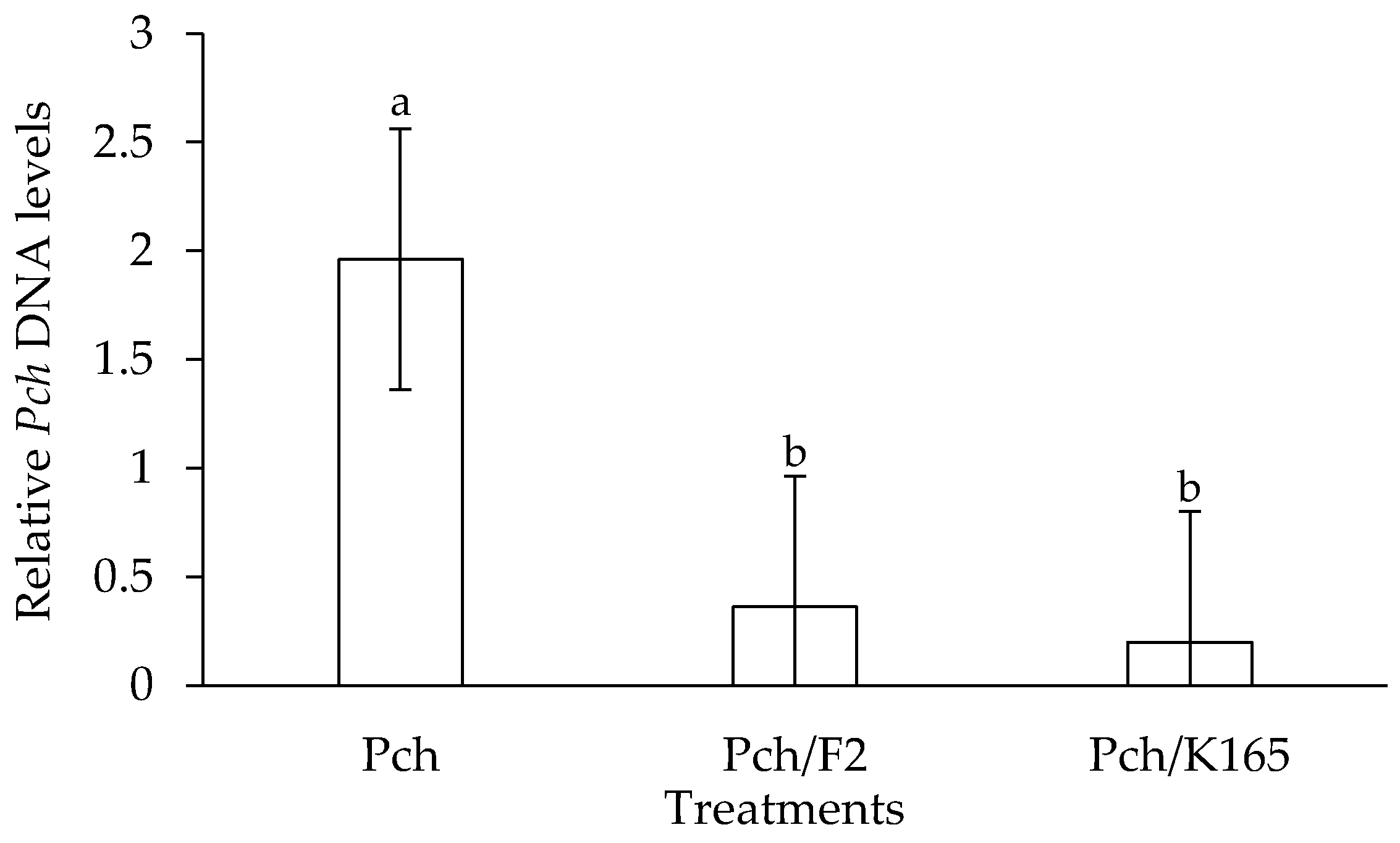{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
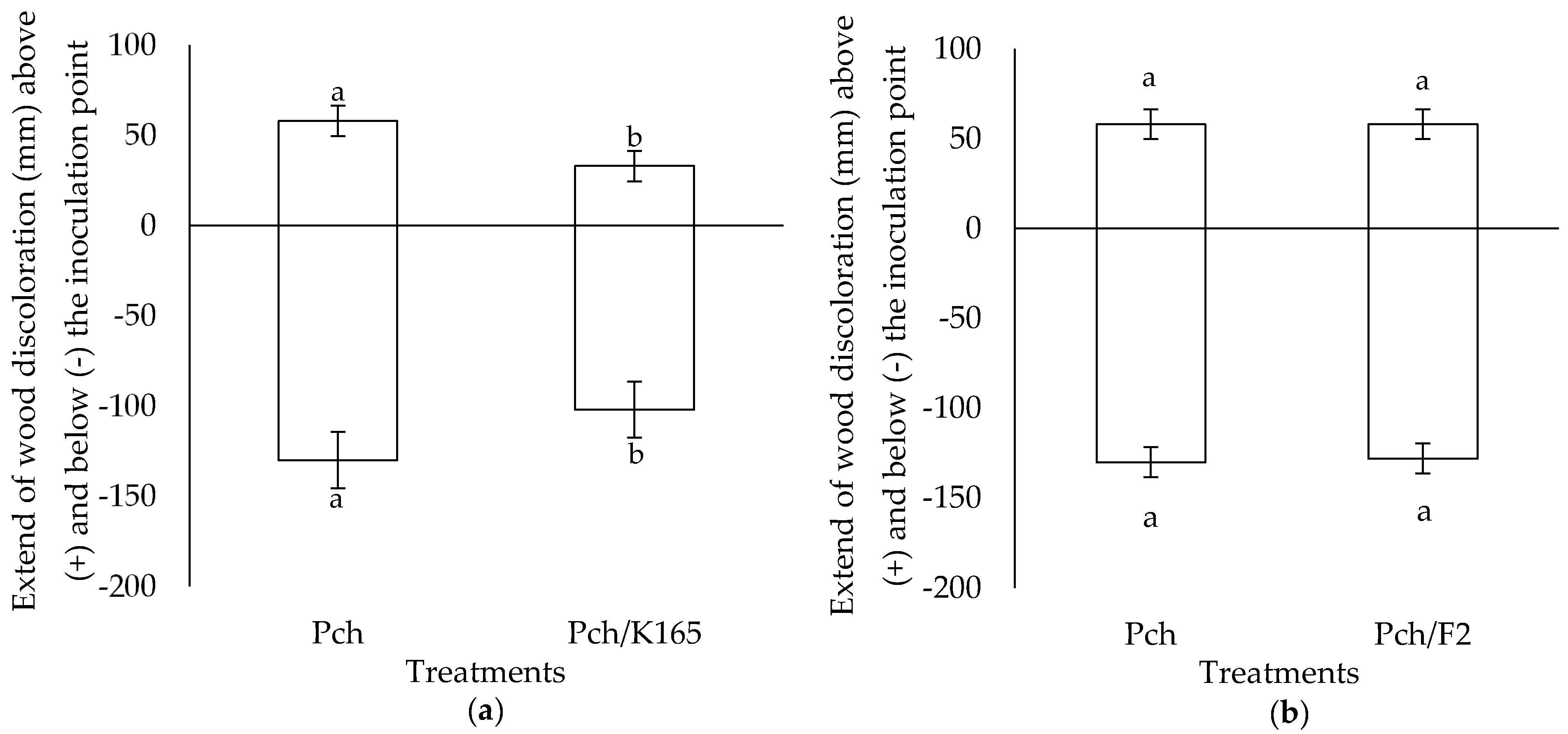
2.1. Evaluation of the Plant Protective Activity of K165 and F2 against Phaeomoniella chlamydospora
2.2. Lignin Quantification
3. Discussion
4. Materials and Methods
4.1. Plant Material
4.2. Microbial Preparation
4.3. In Vitro Activity of K165 and F2 against Phaeomoniella chlamydospora
4.4. In Planta Bioassays to Evaluate the Activity of K165 and F2 against Phaeomoniella chlamydospora
4.5. Disease Assessment
4.6. Colonization Capacity of K165 and F2 in Vines
4.7. qPCR Quantification of Phaeomoniella chlamydospora
4.8. Lignin Quantification
4.9. Statistics
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gramaje, D.; Úrbez-Torres, J.R.; Sosnowski, M.R. Managing grapevine trunk diseases with respect to etiology and epidemiology: Current strategies and future prospects. Plant Dis. 2018, 102, 12–39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mugnai, L.; Granini, A.; Surico, G. Esca (black measles) and brown wood-streaking: Two old and elusive diseases of grapevines. Plant Dis. 1999, 83, 404–418. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Larignon, P.; Dubos, B. Fungi associated with esca disease in grapevine. Eur. J. Plant Pathol. 1997, 103, 147–157. [Google Scholar] [CrossRef]
- Gimenez-Jaime, A.; Aroca, A.; Raposo, R.; Garcia-Jimenez, J.; Armengol, J. Occurrence of fungal pathogens associated with grapevine nurseries and the decline of young vines in Spain. J. Phytopathol. 2006, 154, 598–602. [Google Scholar] [CrossRef]
- Scheck, H.; Vasquez, S.; Fogle, D.; Gubler, W.D. Grape growers report losses to black foot and grapevine decline. Calif. Agric. 1998, 52, 19–23. [Google Scholar] [CrossRef]
- Edwards, J.; Pascoe, I.G. Occurrence of Phaeomoniella chlamydospora and Phaeoacremonium aleophilum associated with Petri disease and esca in Australian grapevines. Australas. Plant Pathol. 2004, 33, 273–279. [Google Scholar] [CrossRef]
- Rumbos, I.; Rumbou, A. Fungi associated with esca and young grapevine decline in Greece. Phytopathol. Mediterr. 2001, 40, S330–S335. [Google Scholar]
- Halleen, F.; Crous, P.W.; Petrini, O. Fungi associated with healthy grapevine cuttings in nursery, with special reference to pathogens involved in the decline of young vines. Australas. Plant Pathol. 2003, 32, 47–52. [Google Scholar] [CrossRef]
- Murolo, S.; Romanazzi, G. Effects of grapevine cultivar, rootstock and clone on esca disease. Australas. Plant Pathol. 2014, 43, 215–221. [Google Scholar] [CrossRef]
- Sparapano, L.; Bruno, G.; Granini, A. Effect on plants of metabolites produced in culture by Phaeoacremonium aleophilum, Phaeoacremonium chlamydosporum and Fomitiporia punctata. Phytopathol. Mediterr. 2000, 39, 169–177. [Google Scholar]
- Abreo, E.; Lupo, S. Phaeomoniella chlamydospora and Phaeoacremonium spp. in grapevines from Uruguay. Phytopathol. Mediterr. 2011, 50, S77. [Google Scholar]
- Cloete, M.; Fischer, M.; Mostert, L.; Halleen, F. Hymenochaetales associated with esca-related wood rots on grapevine with a special emphasis on the status of esca in South African vineyards. Phytopathol. Mediterr. 2015, 54, 299–312. [Google Scholar]
- Lorrain, B.; Ky, I.; Pasquier, G.; Jourdes, M.; Dubrana, L.G.; Gény, L.; Rey, P.; Donèche, B.; Teissedre, P.L. Effect of Esca disease on the phenolic and sensory attributes of Cabernet Sauvignon grapes, musts and wines. Aust. J. Grape Wine R. 2012, 18, 64–72. [Google Scholar] [CrossRef]
- Sidoti, A.; Buonocore, E.; Serges, T.; Mugnai, L. Decline of young grapevines associated with Phaeoacremonium chlamydosporum in Sicily (Italy). Phytopathol. Mediterr. 2000, 39, 87–91. [Google Scholar]
- Eskalen, A.; Gubler, W.D. Association of spores of Phaeomoniella chlamydospora, Phaeoacremonium inflatipes, and Pm. aleophilum with grapevine cordons in California. Phytopathol. Mediterr. 2001, 40, 429–431. [Google Scholar]
- Fourie, P.H.; Halleen, F. Occurrence of grapevine trunk disease pathogens in rootstock mother plants in South Africa. Australas. Plant Path. 2004, 33, 313–315. [Google Scholar] [CrossRef]
- Yacoub, A.; Gerbore, J.; Magnin, N.; Chambon, P.; Dufour, M.C.; Corio-Costet, M.F.; Guyoneaud, R.; Rey, P. Ability of Pythium oligandrum to protect Vitis vinifera L., by inducing plant resistance against Phaeomoniella chlamydospora, a pathogen involved in Esca, a grapevine trunk disease. Biol. Control. 2016, 92, 7–16. [Google Scholar]
- Yacoub, A.; Magnin, N.; Gerbore, J.; Haidar, R.; Bruez, E.; Compant, S.; Guyoneaud, R.; Rey, P. The biocontrol root-oomycete, Pythium oligandrum, triggers grapevine resistance and shifts in the transcriptome of the trunk pathogenic fungus, Phaeomoniella chlamydospora. Int. J. Mol. Sci. 2020, 21, 6876. [Google Scholar] [CrossRef]
- Fourie, P.H.; Halleen, F. Chemical and biological protection of grapevine propagation material from trunk disease pathogens. Eur. J. Plant Pathol. 2006, 116, 255–265. [Google Scholar] [CrossRef]
- Kotze, C.; van Niekerk, J.; Mostert, L.; Halleen, F.; Fourie, P. Evaluation of biocontrol agents for grapevine pruning wound protection against trunk pathogen infection. Phytopathol. Mediterr. 2011, 50, S247–S263. [Google Scholar]
- Alfonzo, A.; Ventorino, V.; Torta, L.; Burruano, S.; Moschetti, G. In vitro antagonism of a grapevine endophytic Bacillus subtilis strain towards “esca” fungi. Integr. Prot. Vitic. IOBC/wprs Bull. 2008, 36, 19–24. [Google Scholar]
- Andreolli, M.; Zapparoli, G.; Angelini, E.; Lucchetta, G.; Lampis, S.; Vallini, G. Pseudomonas protegens MP12: A plant growth-promoting endophytic bacterium with broad-spectrum antifungal activity against grapevine phytopathogens. Microbiol. Res. 2019, 219, 123–131. [Google Scholar] [CrossRef] [PubMed]
- Niem, J.M.; Billones-Baaijens, R.; Stodart, B.; Savocchia, S. Diversity profiling of grapevine microbial endosphere and antagonistic potential of endophytic Pseudomonas against grapevine trunk diseases. Front. Microbiol. 2020, 11, 477. [Google Scholar] [CrossRef] [PubMed]
- Gizi, D.; Stringlis, I.A.; Tjamos, S.E.; Paplomatas, E.J. Seedling vaccination by stem injecting a conidial suspension of F2, a non-pathogenic Fusarium oxysporum strain, suppresses Verticillium wilt of eggplant. Biol. Control. 2011, 58, 387–392. [Google Scholar] [CrossRef]
- Tjamos, E.C.; Tsitsigiannis, D.I.; Tjamos, S.E.; Antoniou, P.P.; Katinakis, P. Selection and screening of endorhizosphere bacteria from solarized soils as biocontrol agents against Verticillium dahliae of solanaceous hosts. Eur. J. Plant Pathol. 2004, 110, 35–44. [Google Scholar] [CrossRef]
- Pantelides, I.S.; Tjamos, S.E.; Striglis, I.A.; Chatzipavlidis, I.; Paplomatas, E.J. Mode of action of a non-pathogenic Fusarium oxysporum strain against Verticillium dahliae using Real Time QPCR analysis and biomarker transformation. Biol. Control. 2009, 50, 30–36. [Google Scholar] [CrossRef]
- Tjamos, S.E.; Flemetakis, E.; Paplomatas, E.J.; Katinakis, P. Induction of resistance to Verticillium dahliae in Arabidopsis thaliana by the biocontrol agent K-165 and pathogenesis-related proteins gene expression. Mol. Plant Microbe Interact. 2005, 18, 555–561. [Google Scholar] [CrossRef] [Green Version]
- Neumann, M.J.; Dobinson, K.F. Sequence tag analysis of gene expression during pathogenic growth and microsclerotia development in the vascular wilt pathogen Verticillium dahliae. Fungal. Genet. Biol. 2003, 38, 54–62. [Google Scholar] [CrossRef]
- Markakis, E.A.; Koubouris, G.C.; Sergentani, C.K.; Ligoxigakis, E.K. Evaluation of Greek grapevine cultivars for resistance to Phaeomoniella chlamydospora. Eur. J. Plant Pathol. 2017, 149, 277–283. [Google Scholar] [CrossRef]
- Sattler, S.E.; Funnell-Harris, D.L. Modifying lignin to improve bioenergy feedstocks: Strengthening the barrier against pathogens? Front. Plant Sci. 2013, 4, 70. [Google Scholar] [CrossRef] [Green Version]
- Díaz, G.A.; Latorre, B.A. Efficacy of paste and liquid fungicide formulations to protect pruning wounds against pathogens associated with grapevine trunk diseases in Chile. Crop Prot. 2013, 46, 106–112. [Google Scholar] [CrossRef]
- Agrios, G. Plant Pathology; Elsevier Academic Press: New York, NY, USA, 2005. [Google Scholar]
- Mondello, V.; Songy, A.; Battiston, E.; Pinto, C.; Coppin, C.; Trotel-Aziz, P.; Clément, C.; Mugnai, L.; Fontaine, F. Grapevine Trunk Diseases: A Review of Fifteen Years of Trials for Their Control with Chemicals and Biocontrol Agents. Plant Dis. 2017, 102, 1189–1217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Del Frari, G.; Gobbi, A.; Aggerbeck, M.R.; Oliveira, H.; Hansen, L.H.; Ferreira, R.B. Characterization of the wood mycobiome of Vitis vinifera in a vineyard affected by esca. Spatial distribution of fungal communities and their putative relation with leaf symptoms. Front. Plant Sci. 2019, 10, 910. [Google Scholar] [CrossRef] [Green Version]
- Haidar, R.; Roudet, J.; Bonnard, O.; Dufour, M.C.; Corio-Costet, M.F.; Fert, M.; Gautier, T.; Deschamps, A.; Fermaud, M. Screening and modes of action of antagonistic bacteria to control the fungal pathogen Phaeomoniella chlamydospora involved in grapevine trunk diseases. Microbiol. Res. 2016, 192, 172–184. [Google Scholar] [CrossRef]
- Raaijmakers, J.M.; Vlami, M.; de Souza, J.T. Antibiotic production by bacterial biocontrol agents. Anton. Leeuw. 2002, 81, 537–547. [Google Scholar] [CrossRef]
- Gkizi, D. The Role of Epigenetic Inheritance in the Biological Control of Vetricillium dahliae. Ph.D. Thesis, Agricultural University of Athens, Athens, Greece, 2019. [Google Scholar]
- Evidente, A.; Sparapano, L.; Andolfi, A.; Bruno, G. Two naphtalene pentakides from liquid cultures of Phaeoacremonium aleophilum, a fungus associated with esca of grapevine. Phytopathol. Mediterr. 2000, 39, 162–168. [Google Scholar] [CrossRef]
- Tabacchi, R.; Fkyerat, A.; Poliart, C.; Dubin, G.M. Phytotoxins from fungi of esca grapevine. Phytopathol. Mediterr. 2000, 39, 156–161. [Google Scholar] [CrossRef]
- Bruno, G.; Sparapano, L. Effects of three esca-associated fungi on Vitis vinifera L: I. Characterization of secondary metabolites in culture media and host responses to the pathogens in calli. Physiol. Mol. Plant Pathol. 2006, 69, 209–223. [Google Scholar] [CrossRef]
- Bruno, G.; Sparapano, L. Effects of three esca-associated fungi on Vitis vinifera L. Characterization of biomolecules in xylem sap and leaves of healthy and diseased vines. Physiol. Mol. Plant Pathol. 2006, 69, 195–208. [Google Scholar] [CrossRef]
- Campbell, M.M.; Ellis, B.E. Fungal elicitor-mediated responses in pine cell cultures: I. induction of phenylpropanoid metabolism. Planta 1992, 186, 409–417. [Google Scholar] [CrossRef]
- Reddy, M.S.; Patrick, Z.A. Colonization of tobacco seedling roots by fluorescent pseudomonad suppressive to black root rot caused by Thielaviopsis basicola. Crop Prot. 1992, 11, 148–154. [Google Scholar] [CrossRef]
- Dellaporta, S.L.; Wood, J.; Hicks, J.B. A plant DNA minipreparation, version II. Plant Mol. Biol. Rep. 1983, 1, 19–21. [Google Scholar] [CrossRef]
- Tegli, S.; Bertelli, E.; Surico, G. Sequence analysis of ITS ribosomal DNA in five Phaeoacremonium species and development of a PCR based assay for the detection of P. chlamydospora and P. aleophilum in grapevine tissue. Phytopathol. Mediterr. 2000, 39, 134–149. [Google Scholar] [CrossRef]
- Dufour, M.C.; Lambert, C.; Bouscaut, J.; Mérillon, J.M.; Corio-Costet, M.F. Benzothiadiazole primed defence responses and enhanced differential expression of defence genes in Vitis vinifera infected with biotrophic pathogens Erysiphe necator and Plasmopara viticola. Plant Pathol. 2013, 62, 370–382. [Google Scholar] [CrossRef]
- Schenk, S.T.; Schikora, A. Lignin extraction and quantification, a tool to monitor defense reaction at the plant cell wall level. Bio-Protocol 2015, 5, e1430. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gkikas, F.-I.; Tako, A.; Gkizi, D.; Lagogianni, C.; Markakis, E.A.; Tjamos, S.E. Paenibacillus alvei K165 and Fusarium oxysporum F2: Potential Biocontrol Agents against Phaeomoniella chlamydospora in Grapevines. Plants 2021, 10, 207. https://doi.org/10.3390/plants10020207
Gkikas F-I, Tako A, Gkizi D, Lagogianni C, Markakis EA, Tjamos SE. Paenibacillus alvei K165 and Fusarium oxysporum F2: Potential Biocontrol Agents against Phaeomoniella chlamydospora in Grapevines. Plants. 2021; 10(2):207. https://doi.org/10.3390/plants10020207
Chicago/Turabian StyleGkikas, Fedon-Ioannis, Alexandros Tako, Danai Gkizi, Christina Lagogianni, Emmanouil A. Markakis, and Sotirios E. Tjamos. 2021. "Paenibacillus alvei K165 and Fusarium oxysporum F2: Potential Biocontrol Agents against Phaeomoniella chlamydospora in Grapevines" Plants 10, no. 2: 207. https://doi.org/10.3390/plants10020207