In-Site and Ex-Site Date Palm Exposure to Heavy Metals Involved Infra-Individual Biomarkers Upregulation

 and
and
Abstract
:1. Introduction
2. Material and Methods
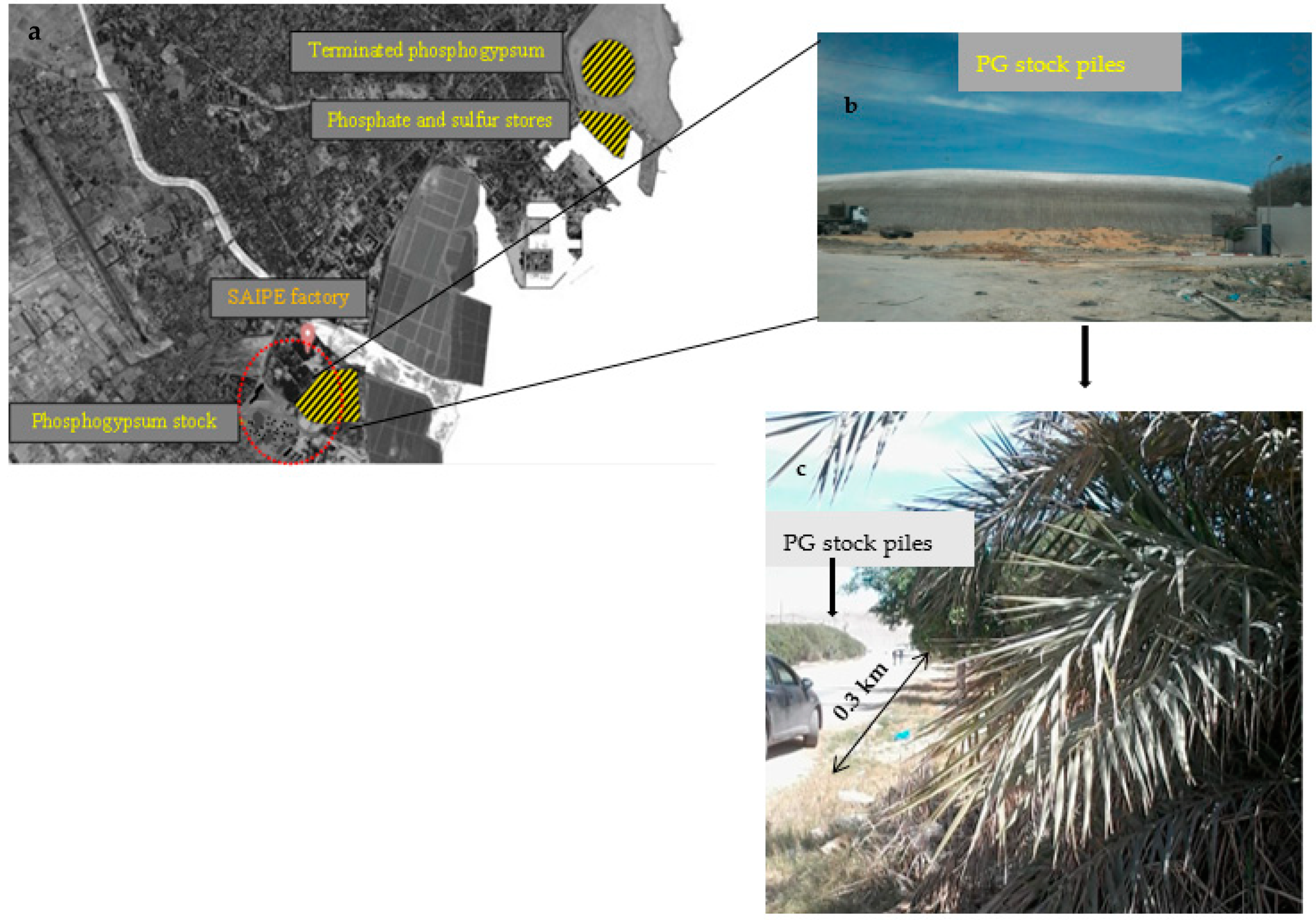
2.1. Soils and Plants Sampling from the Study Site
2.2. Ex-Situ Soil Contamination and Plant Transfer
2.3. Soil and Plant Digestion for Metal Spectroscopic Analysis and Data Processing
2.4. Biochemical Analysis
2.5. Real Time qPCR Amplification
2.6. Statistical Analysis
3. Results
3.1. Soil Contamination and Metal Concentrations in Field
3.2. Metal Concentrations in Plants
3.3. Accumulation of Oxidants in Plant under HMs Stress
3.4. Enzymatic Antioxidant and Secondary Metabolites Potential in Date Palm under HMs Stress
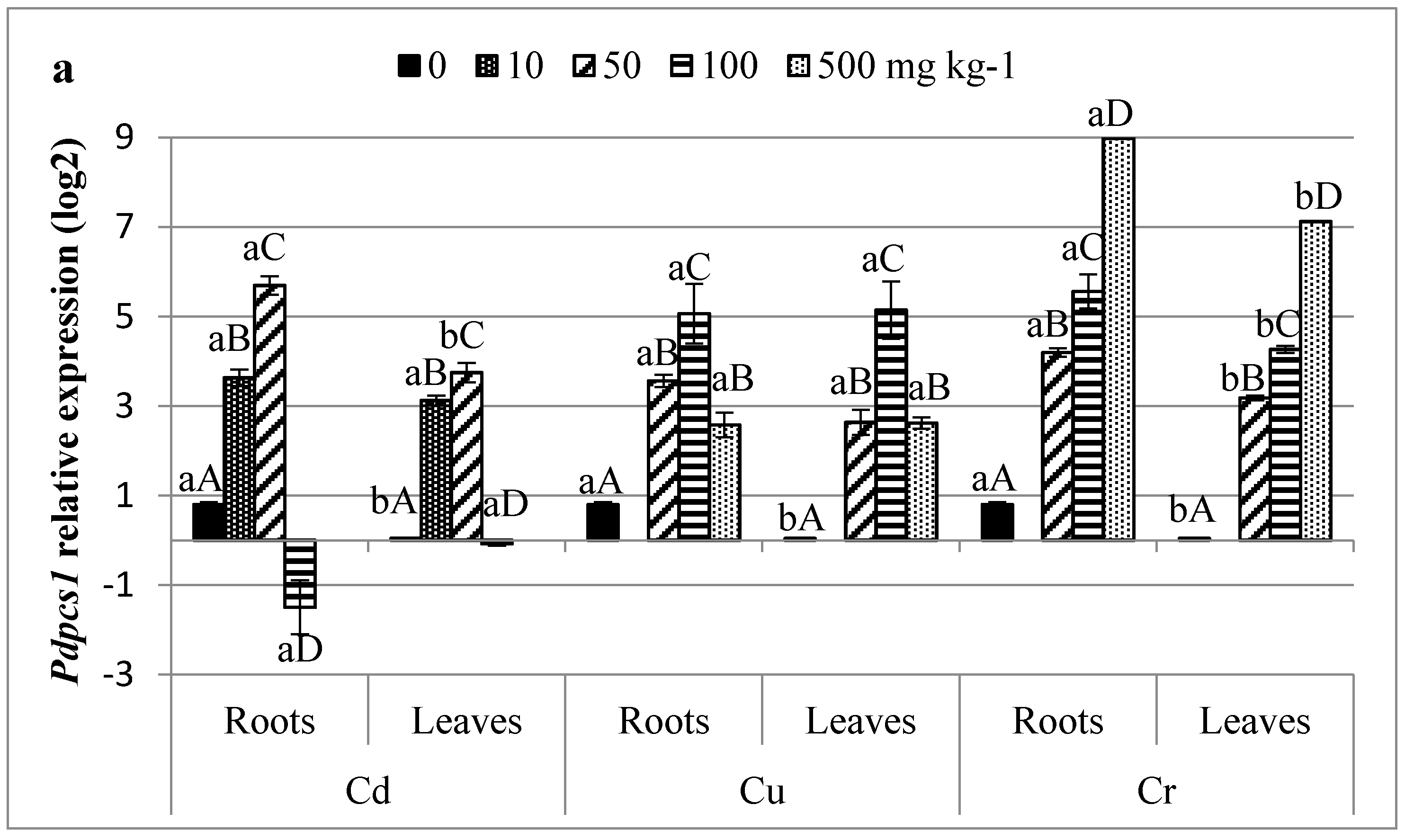
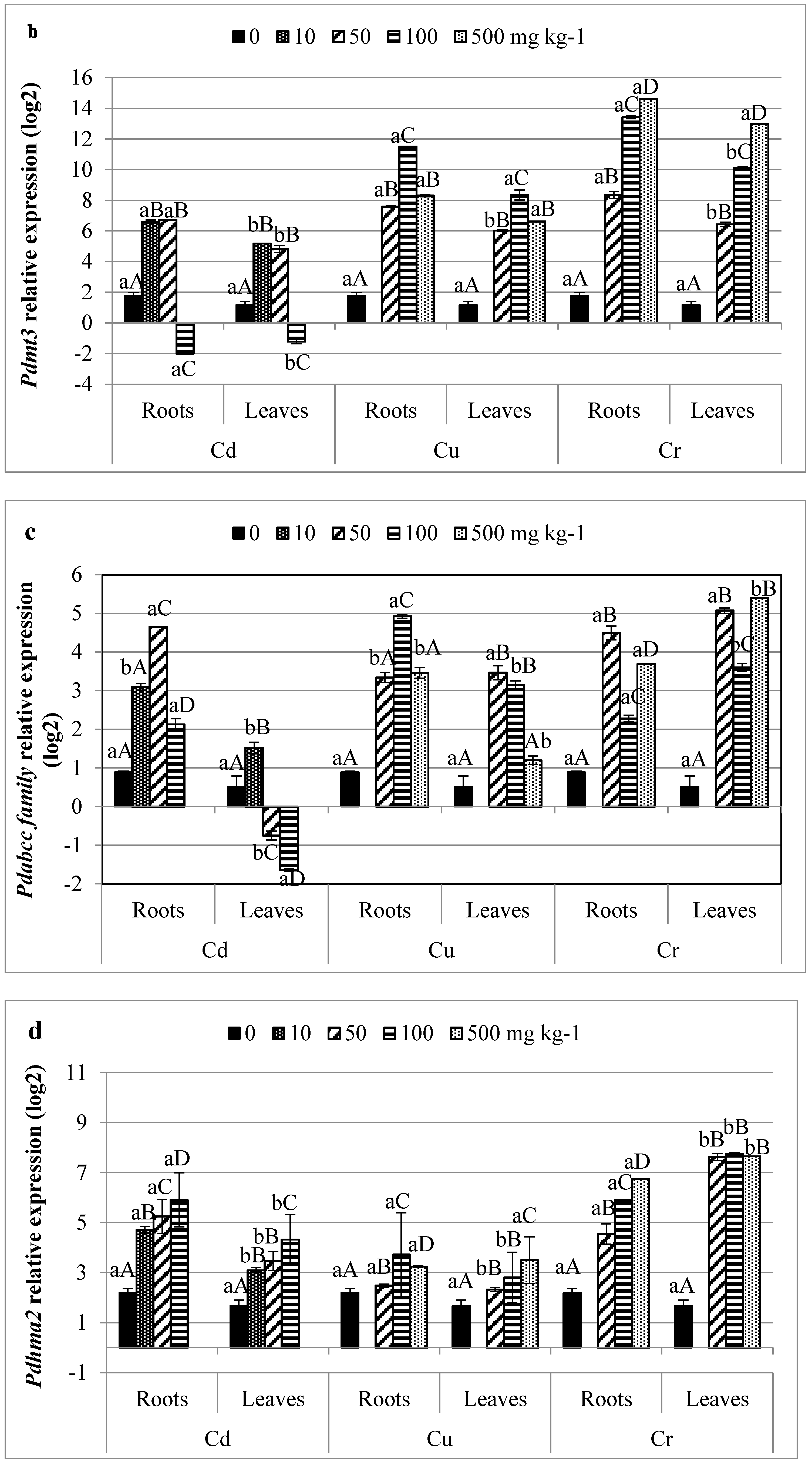
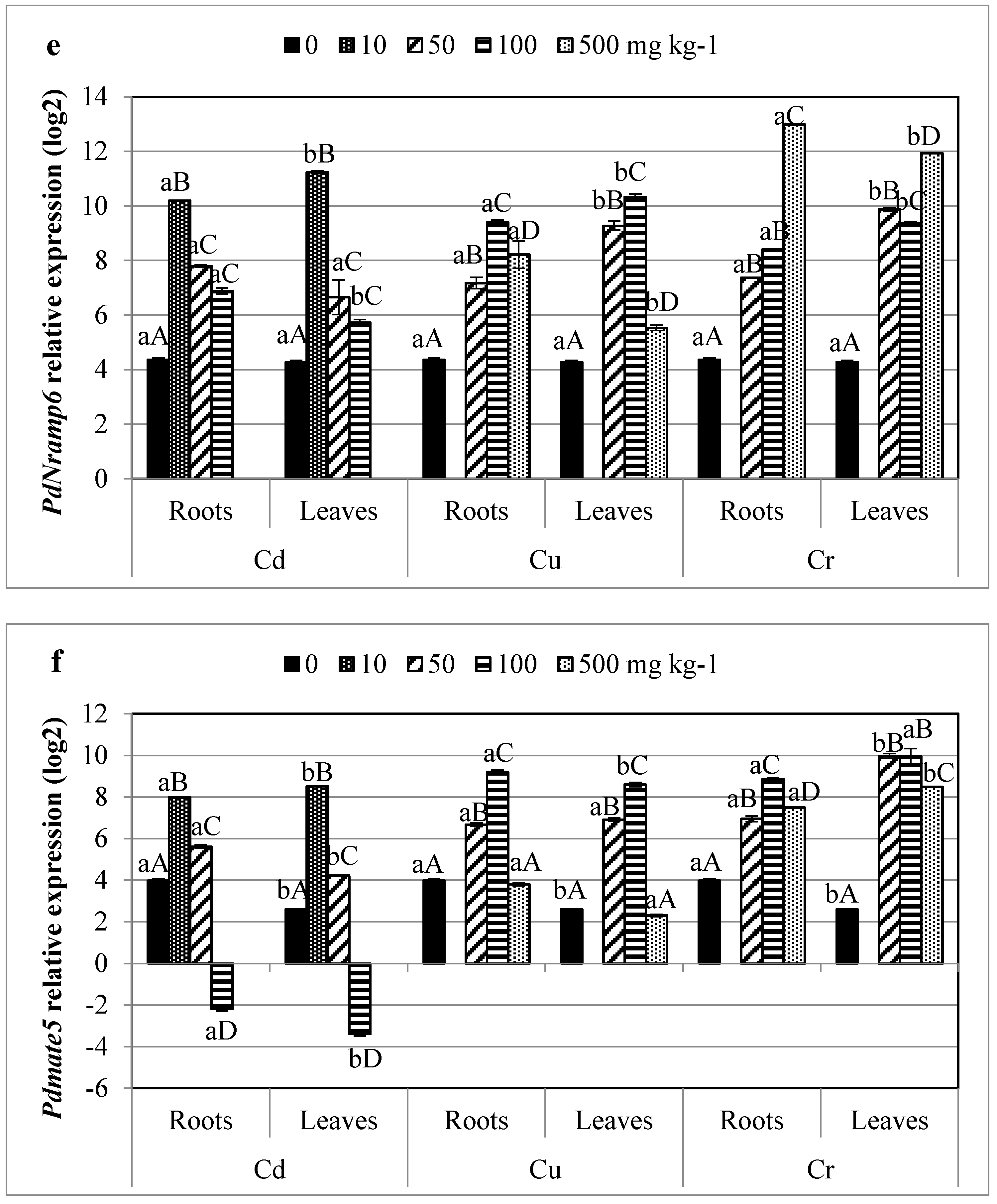
3.5. Profiling of HMs Related Gene Transcripts in Date Palm
4. Discussion
4.1. Metals Concentration and Their Interactions with Plant Nutrients
4.2. Impact of Non-Essential Metal Uptake on Biochemical Integrative Biomarkers in Date Palm
4.3. Essential Metal Uptake Effects on Biochemical Integrative Biomarkers in Date Palm
4.4. Infra-Individual Expression Biomarkers for HMs Stress on Date Palm
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Peng, F.; Yin, H.; Cao, S.L.; Lou, W.Y. Chapter 6: Enzyme Nanocarries. In Advances in Enzyme Technology Biomass, Biofuels, Biochemicals; Elsevier: Amsterdam, The Netherlands, 2019; pp. 153–168. [Google Scholar]
- Flora, S. Structural, chemical and biological aspects of antioxidants for strategies against metal and mettaloid exposure. Oxid Med. Cell Longev. 2009, 2, 191–206. [Google Scholar] [CrossRef] [PubMed]
- Bielen, A.; Remans, T.; Vangronsveld, J.; Cuypers, A. The influence of metal stress on the availability and redox state of ascorbate, and possible interference with its cellular functions. Int. J. Mol. Sci. 2013, 14, 6382–6413. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maresca, V.; Lettieri, G.; Sorbo, S.; Piscopo, M.; Basile, A. Biological Responses to Cadmium Stress in Liverwort Conocephalum conicum (Marchantiales). Int. J. Mol. Sci. 2020, 21, 6485. [Google Scholar] [CrossRef] [PubMed]
- Toxicol, J.E.A.; Bansal, S.L.; Asthana, S. Environmental & Analytical Toxicology Biologically Essential and Non-Essential Elements Causing Toxicity in Environment. J. Environ. Anal. Toxicol. 2018, 8. [Google Scholar] [CrossRef]
- Guo, S.; Li, Q.; Ou-Yang, C.; Chen, L.; Wang, S.; Chen, F. Lead toxicity induced antioxidant enzyme and phenylalanine ammonia lyase activities in Jatropha curcas L. radicles. Fresenius Environ. Bull. 2009, 5, 811–815. [Google Scholar]
- Festa, R.A.; Thiele, D.J. Magasine; Elsevier: Amsterdam, The Netherlands, 2011; pp. R877–R883. [Google Scholar]
- Aghaz, M.; Bandehagh, A. Effect of lead stress on some leaf characteristics in dill (Anethom graveolens) ecotypes. Int. J. Agron. Plant Prod. 2013, 4, 147–150. [Google Scholar]
- Palevsky, H.I.; Ribalet, F.; Swalwell, J.E.; Cosca, C.E.; Cokelet, E.D.; Feely, R.A.; Armbrust, E.V.; Quay, P.D. The influence of net community production and phytoplankton community structure on CO2 uptake in the Gulf of Alaska. Glob. Biogeochem. Cycles 2013, 27, 664–676. [Google Scholar] [CrossRef]
- Ali, H.; Khan, E.; Ilahi, I. Environmental chemistry and ecotoxicology of hazardous heavy metals: Environmental persistence, toxicity, and bioaccumulation. J. Chem. 2019, 2019. [Google Scholar] [CrossRef] [Green Version]
- Ismael, M.A.; Elyamine, A.M.; Moussa, M.G.; Cai, M.; Zhao, X.; Hu, C. Cadmium in plants: Uptake, toxicity, and its interactions with selenium fertilizers. Metallomics 2019, 11, 255–277. [Google Scholar] [CrossRef]
- Zhang, X.; Zhang, D.; Sun, W.; Wang, T. The adaptive mechanism of plants to iron deficiency via iron uptake, transport, and homeostasis. Int. J. Mol. Sci. 2019, 20, 2424. [Google Scholar] [CrossRef] [Green Version]
- Farhat, N.; Elkhouni, A.; Zorrig, W.; Smaoui, A.; Abdelly, C.; Rabhi, M. Effects of magnesium deficiency on photosynthesis and carbohydrate partitioning. Acta Physiol. Plant. 2016, 38. [Google Scholar] [CrossRef]
- Wang, Y.X.; Hu, Y.; Zhu, Y.F.; Baloch, A.W.; Jia, X.M.; Guo, A.X. Transcriptional and physiological analyses of short-term Iron deficiency response in apple seedlings provide insight into the regulation involved in photosynthesis. BMC Genom. 2018, 19, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Xie, Y.; Ye, S.; Wang, Y.; Xu, L.; Zhu, X.; Yang, J. Transcriptome-based gene profiling provides novel insights into the characteristics of radish root response to Cr stress with next-generation sequencing. Front. Plant Sci. 2015, 6, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Sihag, S.; Brar, B.; Jashi, U. Salicylic acid induced amelioration of chromium toxicity and affects antioxydant enzyme activity in Sorghum biocolo L. Int. J. Phytoremediat. 2019, 21, 293–304. [Google Scholar] [CrossRef]
- Singh, S.; Parihar, P.; Singh, R.; Singh, V.P.; Prasad, S.M. Heavy Metal Tolerance in Plants: Role of Transcriptomics, Proteomics, Metabolomics, and Ionomics. Front. Plant Sci. 2016, 6, 1–36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boyd, R.S. Elemental Defenses of Plants by Metals. Nat. Educ. Knowl. 2010, 3, 57. [Google Scholar]
- Rekik, I.; Chaâbene, Z.; Kriaa, W.; Rorat, A.; Franck, V.; Hafedh, M.; Elleuch, A. Transcriptome assembly and abiotic related gene expression analysis of date palm reveal candidate genes involved in response to cadmium stress. Comp. Biochem. Physiol. Part C 2019, 225, 108569. [Google Scholar] [CrossRef] [PubMed]
- Dalcorso, G.; Farinati, S.; Furini, A. Regulatory networks of cadmium stress in plants. Plant Signal. Behav. 2010, 5, 663–667. [Google Scholar] [CrossRef]
- Cobbett, C.S. Phytochelatin biosynthesis and function in heavy-metal detoxification. Curr. Opin. Plant Biol. 2000, 3, 211–216. [Google Scholar] [CrossRef]
- Cobbett, C.; Goldsbrough, P. Phytochelatins and metallothineins: Roles in Heavy Metal Detoxification and Homeostasis. Annu. Rev. Plant Biol. 2002, 53, 159–182. [Google Scholar] [CrossRef] [Green Version]
- Hassinen, V.; Vallinkoski, V.; Issakainen, S.; Tervahauta, A.; Ka, S.; Servomaa, K. Correlation of foliar MT2b expression with Cd and Zn concentrations in hybrid aspen (Populus tremula  tremuloides) grown in contaminated soil. Environ. Pollut. 2009, 157, 922–930. [Google Scholar] [CrossRef] [PubMed]
- Klein, M.; Burla, B.; Martinoia, E. The multidrug resistance-associated protein (MRP/ABCC) subfamily of ATP-binding cassette transporters in plants. FEBS Lett. 2006, 580, 1112–1122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kretzschmar, T.; Burla, B.; Lee, Y.; Martinoia, E.; Nagy, R. Functions of ABC transporters in plants. Essays Biochem. 2011, 50, 145–160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palmer, C.; Guerinot, M. Lou A question of balance: Facing the challenges of Cu, Fe and Zn homeostasis. Nat. Chem. Biol. 2009, 5, 333–340. [Google Scholar] [CrossRef] [Green Version]
- Burkhead, J.L.; Gogolin Reynolds, K.A.; Abdel-Ghany, S.E.; Cohu, C.M.; Pilon, M. Copper homeostasis. New Phytol. 2009, 182, 799–816. [Google Scholar] [CrossRef]
- Lanquar, V.; Lelièvre, F.; Bolte, S.; Hamès, C.; Alcon, C.; Neumann, D.; Vansuyt, G.; Curie, C.; Schröder, A.; Krämer, U.; et al. Mobilization of vacuolar iron by AtNRAMP3 and AtNRAMP4 is essential for seed germination on low iron. EMBO J. 2005, 24, 4041–4051. [Google Scholar] [CrossRef]
- Al-Busaidi, A.; Al-Yahyai, R.; Al-Said, F.; Ahmed, M. Tracking Heavy Metal Concentrations in Soils and Date Palms. Pak. J. Agric. Sci. 2015, 52, 129–134. [Google Scholar]
- Chaâbene, Z.; Rorat, A.; Rekik Hakim, I.; Bernard, F.; Douglas, G.C.; Elleuch, A.; Vandenbulcke, F.; Mejdoub, H. Insight into the expression variation of metal-responsive genes in the seedling of date palm (Phoenix dactylifera). Chemosphere 2018, 197, 123–134. [Google Scholar] [CrossRef]
- Galfati, I.; Bilal, E.; Sassi, A.B.; Abdallah, H.; Zaier, A. Accumulation of heavy metals in native plants growing near the phosphate treatment industry, Tunisia. Carpathian J. Earth Environ. Sci. 2011, 6, 85–100. [Google Scholar]
- Wali, A.; Colinet, G.; Khadhraoui, M.; Ksibi, M. Trace Metals in Surface Soil Contaminated by Release of Phosphate Industry in the Surroundings of Sfax-Tunisia. Environ. Res. Eng. Manag. 2013, 65, 20–30. [Google Scholar] [CrossRef] [Green Version]
- Béjaoui, I.; Hadj, M.B.E.L. Cadmium contamination of local soils and vegetal in a tunisian phosphate plant environment. J. New Sci. 2016, 26, 1445–1453. [Google Scholar]
- Azri, C.; Maalej, A.; Tlili, A.; Medhioub, K. Characterization of the atmospheric pollution level in Sfax City (Tunisia): Influence of sources and meteorological factors. Tech. Sci. Méthodes Génie Urbain Génie Rural 2002, 1, 78–92. [Google Scholar]
- Yanqun, Z.; Yuan, L.; Jianjun, C.; Haiyan, C.; Li, Q.; Schvartz, C. Hyperaccumulation of Pb, Zn and Cd in herbaceous grown on lead—zinc mining area in Yunnan, China. Environ. Int. 2005, 31, 755–762. [Google Scholar] [CrossRef] [PubMed]
- Bu-Olayan, A.H.; Thomas, B.V. Environmental Sciences. Res. J. Environ. Sci. 2009, 3, 581–587. [Google Scholar] [CrossRef] [Green Version]
- Elleuch, A.; Chaâbene, Z.; Grubb, D.C.; Drira, N.; Mejdoub, H.; Khemakhem, B. Morphological and biochemical behavior of fenugreek (Trigonella foenum-graecum) under copper stress. Ecotoxicol. Environ. Saf. 2013, 98, 46–53. [Google Scholar] [CrossRef] [PubMed]
- Rustérucci, C.; Stallaert, V.; Milat, M.; Pugin, A.; Ricci, P.; Blein, J. Relationship between active oxygen species, lipid peroxidation, necrosis, and Phytoalexin Production lnduced by Elicitins in Nicofiana. Plant Physiol. 1996, 111, 885–891. [Google Scholar] [CrossRef] [Green Version]
- Gómez-Vidal, S.; Tena, M.; Vicente Lopez-Llorca, L.; Jesús, S. Protein extraction from Phoenix dactylifera L. leaves, a recalcitrant material, for two-dimensional electrophoresis. Electrophoresis 2008, 29, 448–456. [Google Scholar] [CrossRef]
- Bradford, M.M. A Rapid and Sensitive Method for the Quantitation Microgram Quantities of Protein Utilizing the Principle of Protein-Dye Binding. Anal. Biochem. 1976, 254, 248–254. [Google Scholar] [CrossRef]
- Zayneb, C.; Imen, R.H.; Walid, K.; Grubb, C.D.; Bassem, K.; Franck, V.; Hafedh, M.; Amine, E. The phytochelatin synthase gene in date palm (Phoenix dactylifera L.): Phylogeny, evolution and expression. Ecotoxicol. Environ. Saf. 2017, 140, 7–17. [Google Scholar] [CrossRef]
- Beyer, W.F.; Fridovich, I. Assaying for Superoxide Dismutase Activity: Some Large Consequences of Minor Changes in Conditions. Anal. Biochem. 1987, 161, 559–566. [Google Scholar] [CrossRef]
- Aebi, H. Catalase in vitro. Methods Enzymol. 1984, 105, 121–126. [Google Scholar]
- Nakano, Y.; Asada, K. Hydrogen Peroxide is Scavenged by Ascorbate-specific Peroxidase in Spinach Chloroplasts. Plant Cell Physiol. 1981, 22, 867–880. [Google Scholar]
- Pinelo, M.; Rubilar, M.; Sineiro, J.; Nu, M.J. Extraction of antioxidant phenolics from almond hulls (Prunus amygdalus) and pine sawdust (Pinus pinaster). Food Chem. 2004, 85, 267–273. [Google Scholar] [CrossRef]
- Zhishen, J.; Mengcheng, T.; Jianming, W. The determination of flavonoid contents in mulberry and their scavenging effects on superoxide radicals. Food Chem. 1999, 64, 555–559. [Google Scholar] [CrossRef]
- Price, M.L.; Van Scoyoc, S.; Butler, L.G. A Critical Evaluation of the Vanillin Reaction as an Assay for Tannin in Sorghum Grain. J. Agric. Food Chem. 1978, 26, 1214–1218. [Google Scholar] [CrossRef]
- Brulle, F.; Bernard, F.; Vandenbulcke, F.; Cuny, D.; Dumez, S. Identification of suitable qPCR reference genes in leaves of Brassica oleracea under abiotic stresses. Ecotoxicology 2014, 23, 459–471. [Google Scholar] [CrossRef]
- Brulle, F.; Mitta, G.; Cocquerelle, C.; Vieau, D.; Lemiere, S.; Lepretre, A.; Vandenbulcke, F. Cloning and Real-Time PCR Testing of 14 Potential Biomarkers in Eisenia fetida Following Cadmium Exposure. Environ. Sci. Technol. 2006, 40, 2844–2850. [Google Scholar] [CrossRef]
- Vodyanitskii, Y.N. Standards for the contents of heavy metals in soils of some states. Ann. Agrar. Sci. 2016, 14, 257–263. [Google Scholar] [CrossRef] [Green Version]
- Kassir, L.N.; Lartiges, B.; Ouaini, N. Effects of fertilizer industry emissions on local soil contamination: A case study of a phosphate plant on the east Mediterranean coast. Environ. Technol. 2012, 33, 873–885. [Google Scholar] [CrossRef]
- WHO. Permissible Limits of Heavy Metals in Soil and Plants; World Health Organization: Geneva, Switzerland, 1996.
- Bahloul, M.; Chabbi, I.; Dammak, R.; Amdouni, R.; Medhioub, K.; Azri, C. Geochemical behaviour of PM10 aerosol constituents under the influence of succeeding anticyclonic/cyclonic situations: Case of Sfax City, southern Tunisia. Environ. Monit. Assess. 2015, 187, 757. [Google Scholar] [CrossRef]
- Dong, J.; Wu, F.; Zhang, G. Influence of cadmium on antioxidant capacity and four microelement concentrations in tomato seedlings (Lycopersicon esculentum). Chemosphere 2006, 64, 1659–1666. [Google Scholar] [CrossRef] [PubMed]
- Nazar, R.; Iqbal, N.; Masood, A.; Khan, M.I.R.; Syeed, S.; Khan, N.A. Cadmium Toxicity in Plants and Role of Mineral Nutrients in Its Alleviation. Am. J. Plant Sci. 2012, 03, 1476–1489. [Google Scholar] [CrossRef] [Green Version]
- Millaleo, R.; Reyes-Diaz, M.; Ivanov, A.; Mora, M.; Alberdi, M. Manganese As Essential and Toxic Element for Plants: Transport, Accumulation and Resistance Mechanisms. J. Soil Sci. Plant Nutr. 2010, 10, 470–481. [Google Scholar] [CrossRef] [Green Version]
- Tangahu, B.V.; Rozaimah, S.; Abdullah, S.; Basri, H.; Idris, M.; Anuar, N.; Mukhlisin, M. A Review on Heavy Metals (As, Pb, and Hg) Uptake by Plants through Phytoremediation. Int. J. Chem. Eng. 2011, 31. [Google Scholar] [CrossRef]
- Bright, J.; Desikan, R.; Hancock, J.T.; Weir, I.S.; Neill, S.J.; Lane, C.; Sciences, M.; Lane, C. ABA-induced NO generation and stomatal closure in Arabidopsis are dependent on H2O2 synthesis. Plant J. 2006, 45, 113–122. [Google Scholar] [CrossRef] [PubMed]
- Gill, S.S.; Tuteja, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Biochem. 2010, 48, 909–930. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Nahar, K.; Gill, S.S.; Alharby, H.F.; Razafindrabe, B.H.N.; Fujita, M. Hydrogen Peroxide Pretreatment Mitigates Cadmium-Induced Oxidative Stress in Brassica napus L.: An Intrinsic Study on Antioxidant Defense and Glyoxalase Systems. Front. Plant Sci. 2017, 8, 1–10. [Google Scholar] [CrossRef]
- Romero-Puertas, M.C.; Palma, J.M.; Gómez, M.; Del Río, L.A.; Sandalio, L.M. Cadmium causes the oxidative modification of proteins in pea plants. Plant Cell Environ. 2002, 25, 677–686. [Google Scholar] [CrossRef]
- Rosa, S.B.; Caverzan, A.; Teixeira, F.K.; Lazzarotto, F.; Silveira, J.A.G.; Ferreira-Silva, S.L.; Abreu-Neto, J.; Margis, R.; Margis-Pinheiro, M. Cytosolic APx knockdown indicates an ambiguous redox responses in rice. Phytochemistry 2010, 71, 548–558. [Google Scholar] [CrossRef]
- Gomes-Junior, R.A.; Moldes, C.A.; Delite, F.S.; Pompeu, G.B.; Gratão, P.L.; Mazzafera, P.; Lea, P.J.; Azevedo, R.A. Antioxidant metabolism of coffee cell suspension cultures in response to cadmium. Chemosphere 2006, 65, 1330–1337. [Google Scholar] [CrossRef]
- Ramakrishna, A.; Ravishankar, G.A. Influence of abiotic stress signals on secondary metabolites in plants. Plant Signal. Behav. 2011, 6, 1720–1731. [Google Scholar] [CrossRef]
- Sharma, P.; Jha, A.B.; Dubey, R.S.; Pessarakli, M. Reactive Oxygen Species, Oxidative Damage, and Antioxidative Defense Mechanism in Plants under Stressful Conditions. J. Bot. 2012, 2012, 1–26. [Google Scholar] [CrossRef] [Green Version]
- Ahemad, M. Enhancing phytoremediation of chromium-stressed soils through plant-growth-promoting bacteria. J. Genet. Eng. Biotechnol. 2015, 13, 51–58. [Google Scholar] [CrossRef] [Green Version]
- Chatterjee, J.; Kumar, P.; Nand, P. Chromium toxicity induces oxidative stress in turnip. Indian J. Plant Physiol. 2015, 6, 15–163. [Google Scholar] [CrossRef]
- Dube, B.K.; Tewari, K.; Chatterjee, J.; Chatterjee, C. Excess chromium alters uptake and translocation of certain nutrients in citrullus. Chemosphere 2003, 53, 1147–1153. [Google Scholar] [CrossRef]
- Shanker, A.K.; Djanaguiraman, M.; Sudhagar, R.; Jayaram, K.; Pathmanabhan, G. Expression of metallothionein 3-like protein mRNA in sorghum cultivars under chromium (VI) stress. Curr. Sci. 2004, 86, 901–902. [Google Scholar]
- Singh, S.; Sinha, S. Accumulation of metals and its effects in Brassica juncea (L.) Czern. (cv. Rohini) grown on various amendments of tannery waste. Ecotoxicol. Environ. Saf. 2005, 62, 118–127. [Google Scholar] [CrossRef]
- Dubey, S.; Misra, P.; Dwivedi, S.; Chatterjee, S.; Bag, S.K.; Mantri, S.; Asif, M.H.; Rai, A.; Kumar, S.; Shri, M.; et al. Transcriptomic and metabolomic shifts in rice roots in response to Cr (VI) stress. BMC Genom. 2010, 11, 648. [Google Scholar] [CrossRef] [Green Version]
- Cho, A.K.; Sioutas, C.; Miguel, A.H.; Kumagai, Y.; Schmitz, D.A.; Singh, M.; Eiguren-Fernandez, A.; Froines, J.R. Redox activity of airborne particulate matter at different sites in the Los Angeles Basin. Environ. Res. 2005, 99, 40–47. [Google Scholar] [CrossRef]
- Abdel-Ghany, S.E.; Pilon, M. MicroRNA-mediated systemic down-regulation of copper protein expression in response to low copper availability in Arabidopsis. J. Biol. Chem. 2008, 283, 15932–15945. [Google Scholar] [CrossRef] [Green Version]
- Gielen, H.; Remans, T.; Vangronsveld, J.; Cuypers, A. Toxicity responses of Cu and Cd: The involvement of miRNAs and the transcription factor SPL7. BMC Plant Biol. 2016, 16, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Liu, Y.; Liu, H.; Kang, L.; Geng, J.; Gai, Y.; Ding, Y.; Sun, H.; Li, Y. Identification and expression analysis of MATE genes involved in flavonoid transport in blueberry plants. PLoS ONE 2015, 10, 1–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yadav, P.; Kaur, R.; Kohli, S.K.; Sirhindi, G.; Bhardwaj, R. Castasterone assisted accumulation of polyphenols and antioxidant to increase tolerance of B. juncea plants towards copper toxicity. Cogent Food Agric. 2016, 2, 1–14. [Google Scholar] [CrossRef]
- Mamat, D.D.; Chong, C.S.; Samad, A.A.; Chai, T.T.; Manan, F.A. Effects of copper on total phenolics, flavonoids and mitochondrial properties of Orthosiphon stamineus callus culture. Int. J. Agric. Biol. 2015, 17, 1243–1248. [Google Scholar] [CrossRef]
- Di Toppi, S.L.; Lambardi, M.; Pecchioni, N.; Pazzagli, L.; Durante, M.; Gabbrielli, R. Effects of Cadmium Stress on Hairy Roots of Daucus carota. J. Plant Physiol. 1999, 154, 385–391. [Google Scholar] [CrossRef]
- Degola, F.; De Benedictis, M.; Petraglia, A.; Massimi, A.; Fattorini, L.; Sorbo, S.; Basile, A.; Sanita, L. A Cd/Fe/Zn-Responsive Phytochelatin Synthase is Constitutively Present in the Ancient Liverwort Lunularia cruciata (L.) Dumort. Plant Cell Physiol. 2014, 55, 1884–1891. [Google Scholar] [CrossRef] [Green Version]
- Ramesh, G.; Podila, G.K.; Gay, G.; Marmeisse, R.; Reddy, M.S. Different patterns of regulation for the copper and cadmium Metallothioneins of the ectomycorrhizal fungus Hebeloma cylindrosporum. Appl. Environ. Microbiol. 2009, 75, 2266–2274. [Google Scholar] [CrossRef] [Green Version]
- Finatto, T.; de Oliveira, A.C.; Chaparro, C.; da Maia, L.C.; Farias, D.R.; Woyann, L.G.; Mistura, C.C.; Soares-Bresolin, A.P.; Llauro, C.; Panaud, O.; et al. Abiotic stress and genome dynamics: Specific genes and transposable elements response to iron excess in rice. Rice 2015, 8. [Google Scholar] [CrossRef] [Green Version]
- Tan, J.; Wang, J.; Chai, T.; Zhang, Y.; Feng, S.; Li, Y.; Zhao, H.; Liu, H. Functional analyses of TaHMA2, a P 1B -type ATPase in wheat. Plant Biotechnol. J. 2013, 11, 420–431. [Google Scholar] [CrossRef]
- Liu, W.; Xu, L.; Wang, Y.; Shen, H.; Zhu, X.; Zhang, K.; Chen, Y.; Yu, R.; Limera, C.; Liu, L. Transcriptome-wide analysis of chromium-stress responsive microRNAs to explore miRNA-mediated regulatory networks in radish (Raphanus sativus L.). Sci. Rep. 2015, 5, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Ishimaru, Y.; Takahashi, R.; Bashir, K.; Shimo, H.; Senoura, T.; Sugimoto, K.; Ono, K.; Yano, M.; Ishikawa, S.; Arao, T.; et al. Characterizing the role of rice NRAMP5 in Manganese, Iron and Cadmium Transport. Sci. Rep. 2012, 2, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Durrett, T.P.; Gassmann, W.; Rogers, E.E. The FRD3-Mediated Efflux of Citrate into the Root Vasculature Is Necessary for Efficient Iron Translocation. Plant Physiol. 2007, 144, 197–205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ritter, A.; Dittami, S.M.; Goulitquer, S.; Correa, J.A.; Boyen, C.; Potin, P.; Tonon, T. Transcriptomic and metabolomic analysis of copper stress acclimation in Ectocarpus siliculosus highlights signaling and tolerance mechanisms in brown algae. BMC Plant Biol. 2014, 14, 116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leng, X.; Jia, H.; Sun, X.; Shangguan, L.; Mu, Q.; Wang, B.; Fang, J. Comparative transcriptome analysis of grapevine in response to copper stress. Sci. Rep. 2015, 5, 1–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Doped Soils | |||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Uncontaminated Site | Contaminated Site | Cd (mg kg−1) | Cu (mg kg−1) | Cr (mg kg−1) | |||||||||||||||
| Cd | Cu | Cr | Cd | Cu | Cr | 0 | 10 | 50 | 100 | 0 | 50 | 100 | 500 | 0 | 50 | 100 | 500 | ||
| Metal Concentration (mg kg−1) | Soil | 0.3 (0.1) | 5.4 (0.4) | 0.1 (0.1) | 0.9 (0.1) | 7.5 (0.3) | 28.0 (0.8) | 0.7aA (0.02) | 4.4aB (0.2) | 26.3aC (1.1) | 63.9aD (0.9) | 5.68aA (0.02) | 19.3aB (1.4) | 33.4aC (2.5) | 64.7aD (0.9) | 0.05aA (0.02) | 26.5aB (2.0) | 71.8aC (1.6) | 29.2aD (3.9) |
| Roots | - | - | - | - | - | - | 0.2bA (0.02) | 4.5aAB (0.0) | 16.0aBC (5.04) | 27.1bC (5.1) | 4.85aA (0.02) | 17.2abB (2.9) | 18.8bB (2.1) | 19.6bB (1.9) | 0.64bA (0.02) | 9.8bB (0.6) | 15.4bB (0.8) | 25.8bC (0.2) | |
| Leaves | 0.2 (0.0) | 3.8 (0.7) | 0.1 (0.8) | 1.6 (0.7) | 14.6 (2.8) | 17.9 (1.9) | 0.4cA (0.09) | 2.1aA (1.5) | 12.2bB (1.2) | 15.3abB (3.7) | 3.82bA (0.09) | 9.3bB (1.2) | 12.7bB (1.9) | 13.8bB (0.7) | 0.02aA (0.09) | 7.2bB (0.3) | 8.7bB (0.2) | 14.2cC (2.5) | |
| Zn Concentration (mg kg−1) | Soil | 9.4 (0.2) | 15.4 (0.7) | 19.7aA (0.4) | 34.9aB (0.0) | 36.1aC (0.1) | 41.2aD (0.3) | 19.7aA (0.4) | 23.91aA (1.8) | 29.8aA (5.3) | 32.82aA (2.8) | 19.7aA (0.4) | 25.1aB (0.1) | 36.32aC (0.2) | 29.85aC (0.6) | ||||
| Roots | - | - | 17.50bA (0.6) | 10.47bB (0.6) | 8.4aAB (2.0) | 6.6aAB (2.2) | 17.5bA (0.6) | 16.93bB (0.2) | 11.14aC (0.3) | 10.2aD (0.0) | 17.5bA (0.6) | 14.3bB (0.4) | 9.2bC (3.0) | 8.8bC (4.0) | |||||
| Leaves | 14.0 (1.1) | 36.0 (1.5) | 29.3cA (0.9) | 18.9aA (1.4) | 15.5bA (0.7) | 6.3bB (0.5) | 29.3cA (0.9) | 22.8abB (2.8) | 10.87aC (0.2) | 8.85aC (0.0) | 29.3cA (0.9) | 18.4bB (0.3) | 11.81aC (4.0) | 10.82aC (0.1) | |||||
| Cd (mg kg−1) | Cu (mg kg−1) | Cr (mg kg−1) | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 0 | 10 | 50 | 100 | 0 | 50 | 100 | 500 | 0 | 50 | 100 | 500 | |
| BAF | 0.64 (0.0) | 0.74 (0.2) | 0.46 (0.1) | 0.24 (0.1) | 0.29 (0.0) | 0.49 (0.0) | 0.38 (0.1) | 0.21 (0.1) | 0.38 (0.0) | 0.27 (0.0) | 0.12 (0.0) | 0.04 (0.0) |
| EF | 0.15 (0.4) | 4.94 (0.4) | 4.48 (0.3) | 5.87 (0.2) | 0.64 (0.0) | 1.28 (0.5) | 1.99 (0.1) | 1.39 (0.7) | 5.55 (0.4) | 0.08 (0.0) | 0.1 (0.02) | 0.3 (0.0) |
| TF | 0.23 (0.) | 1.02 (0.0) | 0.60 (0.0) | 0.42 (0.1) | 1.77 (0.) | 0.9 (0.2) | 0.56 (0.0) | 0.30 (0.0) | 1.5 (0.0) | 0.90 (0.1) | 0.56 (0.0) | 0.30 (0.0) |
| BCF | 0.93 (0.1) | 1.72 (0.0) | 1.07 (0.0) | 0.66 (0.1) | 0.19 (0.1) | 1.37 (0.1) | 0.98 (0.3) | 0.51 (0.0) | 1.89 (0.1) | 0.37 (0.0) | 0.21 (0.0) | 0.08 (0.0) |
| Doped Soils | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Uncontaminated Site | Contaminated Site | Cd (mg kg−1) | Cu (mg kg−1) | Cr (mg kg−1) | |||||||||||
| 0 | 10 | 50 | 100 | 0 | 50 | 100 | 500 | 0 | 50 | 100 | 500 | ||||
| Mn Concentration (mg kg−1) | Soil | 9.0 (0.4) | 14.1 (1.8) | 40.7aA (0.3) | 42.2aB (0.3) | 44.9aC (0.0) | 44.9aC (0.1) | 40.7aA (0.3) | 41.8aB (1.5) | 44.0aAB (0.0) | 44.3aB (0.3) | 40.7aA (0.3) | 46.8aA (2.6) | 43.6aA (1.9) | 46.4aA (2.0) |
| Roots | - | - | 4.20bA (0.4) | 6.2bB (0.3) | 4.8bAB (1.0) | 2.2bA (0.3) | 4.2bA (0.4) | 6.1bB (0.1) | 8.4bC (0.7) | 4.0bA (0.5) | 4.2bA (0.4) | 8.4bB (0.5) | 7.8bC (0.2) | 6.8bBC (1.1) | |
| Leaves | 4.5 (0.9) | 16.5 (1.3) | 18.3cA (0.5) | 12.3cB (0.5) | 7.1bC (0.2) | 5.1cD (0.3) | 18.3cA (0.5) | 11.2bB (0.3) | 8.6bC (0.5) | 5.6bD (0.5) | 18.3cA (0.5) | 12.3bB (0.4) | 8.9bC (0.0) | 7.1bD (0.2) | |
| Pairwise Correlation coefficient (r) | |||||||||||||||
| Metal concentration-Mn concentration | 0.914191 | −0.611946 | 0.914191 | −0.48363 | 0.914191 | −0.33352 | |||||||||
| H2O2 µmol/g FW | TBARS nmol/g FW | CAT µmole H2O2/mg Protein | APX | SOD U/mg Protein | Polyphenol % | Flavonoid % | Tannins % | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Root | Leaves | Root | Leaves | Root | Leaves | Root | Leaves | Root | Leaves | Root | Leaves | Root | Leaves | Root | Leaves | ||
| 0 | 156.1aA (4.4) | 244.6bA (5.8) | 44.9aA (2.9) | 56.5bA (4.0) | 1.6aA (0.0) | 1.7aA (0.2) | 8.7aA (0.2) | 9.9bA (0.6) | 444.1aA (6.2) | 550.1bA (28.7) | 100aA (0.00) | 100aA (0.00) | 100aA (0.00) | 100aA (0.00) | 100aA (0.00) | 100aA (0.00) | |
| Cd (mg kg−1) | 10 | 270.4aB (1.3) | 381.9bB (22.0) | 62.5aB (4.7) | 70.4bAB (0.7) | 2.7aB (0.1) | 3.5bB (0.0) | 12.5aB (0.1) | 13.8bB (0.2) | 590.3aB (9.3) | 612.3bB (8.5) | 114.8aB (4.7) | 153.2bB (3.8) | 201.5aB (3.3) | 201.1aB (6.1) | 132.4aB (4.19) | 113.8bB (3.4) |
| 50 | 568.2aC (12.2) | 599.6bC (1.7) | 94.1aC (4.1) | 86.8aC (9.7) | 1.5aA (0.0) | 1.8bC (0.0) | 12.7aB (0.2) | 14.7bB (0.1) | 540.0aB (13.0) | 542.8aA (28.1) | 134.6aC (1.4) | 145.0aB (10.6) | 186.3aB (9.8) | 205.2aB (6.5) | 109.5aA (5.8) | 150.0bC (5.4) | |
| 100 | 709.5aD (18.7) | 598.5bC (6.5) | 98.3aC (3.0) | 82.2bBC (2.7) | 0.9aC (0.0) | 0.4bD (0.0) | 6.1aB (0.1) | 11.4bC (0.7) | 310.6aC (10.0) | 373.6bC (22.4) | 72.0aD (4.3) | 83.7bC (2.8) | 85.4aC (5.9) | 60.6bC (8.0) | 55.8aC (4.2) | 64.9bD (1.7) | |
| One-way and Two-way ANOVA main effects | |||||||||||||||||
| Tissues | 0.000 * | 0.000 * | 0.000 * | 0.000 * | 0.000 * | 0.000 * | 0.000 * | 0.000 * | |||||||||
| Concentration | 0.000 * | 0.626 | 0.000 * | 0.000 * | 0.000 * | 0.000 * | 0.520 | 0.000 * | |||||||||
| Tissues x Concentration | 0.000 * | 0.000 * | 0.002 * | 0.003 * | 0.001 * | 0.000 * | 0.000 * | 0.000 * | |||||||||
| Cu (mg kg−1) | 0 | 156.1aA (4.4) | 244.6bA (5.8) | 44.9aA (2.9) | 56.5bA (4.0) | 1.6aA (0.0) | 1.7aA (0.2) | 8.7aA (0.2) | 9.9bA (0.6) | 444.1aA (6.2) | 550.1bA (28.7) | 100aA (0.00) | 100aA (0.00) | 100aA (0.00) | 100aA (0.00) | 100aA (0.00) | 100aA (0.00) |
| 50 | 202.9aB (4.6) | 248.4aA (20.9) | 44.3aA (3.4) | 63.5bAB (3.1) | 2.0aB (0.1) | 2.0aAB (0.1) | 10.9aB (0.1) | 11.8bB (0.2) | 583.8aB (13.8) | 633.5bB (17.7) | 116.7aA (7.0) | 127.2aB (3.8) | 238.1aA (6.6) | 247.8aB (1) | 148.4aB (7) | 151.8aB (6) | |
| 100 | 319.4aC (17.8) | 325.0aB (25.0) | 53.5aA (2.31) | 69.2bBC (3.17) | 2.4aC (0.1) | 2.4bBC (0.0) | 12.5aBC (0.1) | 13.7bC (0.2) | 660.8aC (12.3) | 717.1bC (9.5) | 151.5aB (6.3) | 185.2bC (12.5) | 271.2aA (18.1) | 184.8bC (11.4) | 162.6aC (6.5) | 179.8bC (2.8) | |
| 500 | 429.1aD (20.2) | 464.3aC (15.4) | 79.5aB (7.5) | 88.1aC (0.8) | 2.3aBC (0.2) | 2.8bC (0.1) | 11.9aC (1.4) | 14.1aC (0.1) | 829.6aB (5.5) | 776.0bC (11.) | 100.8aA (12.8) | 110.2aAB (7.68) | 223.7aA (8.2) | 94.5bA (12.9) | 87aA (4.3) | 119.7bD (6.2) | |
| One-way and Two-way ANOVA main effects | |||||||||||||||||
| Tissues | 0.000 * | 0.000 * | 0.000 * | 0.000 * | 0.000 * | 0.000 * | 0.613 | 0.000 * | |||||||||
| Concentration | 0.015 * | 0.000 * | 0.340 * | 0.001 * | 0.000 * | 0.990 | 0.539 | 0.004 * | |||||||||
| Tissues x Concentration | 0.352 | 0.175 | 0.780 | 0.453 | 0.350 | 0.363 | 0.648 | 0.034 * | |||||||||
| Cr (mg kg−1) | 0 | 156.1aA (4.4) | 244.6bA (5.8) | 44.9aA (2.9) | 56.5bA (4.0) | 1.6aA (0.0) | 1.7aA (0.2) | 8.7aA (0.2) | 9.9bA (0.6) | 444.1aA (6.2) | 550.1bA (28.7) | 100aA (0.00) | 100aA (0.00) | 100aA (0.00) | 100aA (0.00) | 100aA (0.00) | 100aA (0.00) |
| 50 | 183.9aA (18.1) | 268.5bA (16.0) | 45.1aA (1.9) | 59.0bA (1.9) | 2.1aAB (0.0) | 1.8aAB (0.1) | 11.0aB (0.1) | 11.4bB (0.1) | 503.5aB (7.6) | 576.0bA (9.7) | 120.2aB (1.2) | 152.6bB (2.6) | 152.5aB (3.6) | 130.7bB (1.3) | 169.6aB (3.5) | 153.0bB (5.9) | |
| 100 | 317.7aB (3.2) | 347.9aB (32.7) | 60.6aB (7.5) | 72.6aB (2.4) | 2.4aBC (0.3) | 2.2aBC (0.1) | 12.6aC (0.3) | 12.2aB (0.2) | 617.1aC (26.8) | 684.5bB (11.1) | 160.6aC (6.7) | 217.8bC (0.4) | 272.8aC (8.3) | 210.2bC (8.9) | 127.0aC (3.5) | 181.3bC (6.2) | |
| 500 | 589.4aC (11.7) | 520.9bC (27.6) | 83.0aC (2.7) | 87.1aC (0.8) | 2.7aC (0.2) | 2.3aC (0.1) | 14.5aD (0.4) | 14.5aC (0.3) | 730.7aD (11.6) | 787bC (16.6) | 130.1aD (1.0) | 113.6bD (9.4) | 199.00aA (4.1) | 151.6bA (1.5) | 103.5aA (3.3) | 112.4aA (6.7) | |
| One-way and Two-way ANOVA main effects | |||||||||||||||||
| Tissues | 0.000 * | 0.000 * | 0.000 * | 0.000 * | 0.000 * | 0.000 * | 0.000 * | 0.000 * | |||||||||
| Concentration | 0.229 | 0.005 * | 0.065 | 0.154 | 0.001 * | 0.342 | 0.519 | 0.177 | |||||||||
| Tissues x Concentration | 0.022 * | 0.440 | 0.219 | 0.149 | 0.640 | 0.029 * | 0.900 | 0.470 | |||||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chaâbene, Z.; Rorat, A.; Kriaa, W.; Rekik, I.; Mejdoub, H.; Vandenbulcke, F.; Elleuch, A. In-Site and Ex-Site Date Palm Exposure to Heavy Metals Involved Infra-Individual Biomarkers Upregulation. Plants 2021, 10, 137. https://doi.org/10.3390/plants10010137
Chaâbene Z, Rorat A, Kriaa W, Rekik I, Mejdoub H, Vandenbulcke F, Elleuch A. In-Site and Ex-Site Date Palm Exposure to Heavy Metals Involved Infra-Individual Biomarkers Upregulation. Plants. 2021; 10(1):137. https://doi.org/10.3390/plants10010137
Chicago/Turabian StyleChaâbene, Zayneb, Agnieszka Rorat, Walid Kriaa, Imen Rekik, Hafedh Mejdoub, Franck Vandenbulcke, and Amine Elleuch. 2021. "In-Site and Ex-Site Date Palm Exposure to Heavy Metals Involved Infra-Individual Biomarkers Upregulation" Plants 10, no. 1: 137. https://doi.org/10.3390/plants10010137