Targeting Cytokinin Homeostasis in Rapid Cycling Brassica rapa with Plant Growth Regulators INCYDE and TD-K

 , , and
, , and
Abstract
:1. Introduction
2. Results
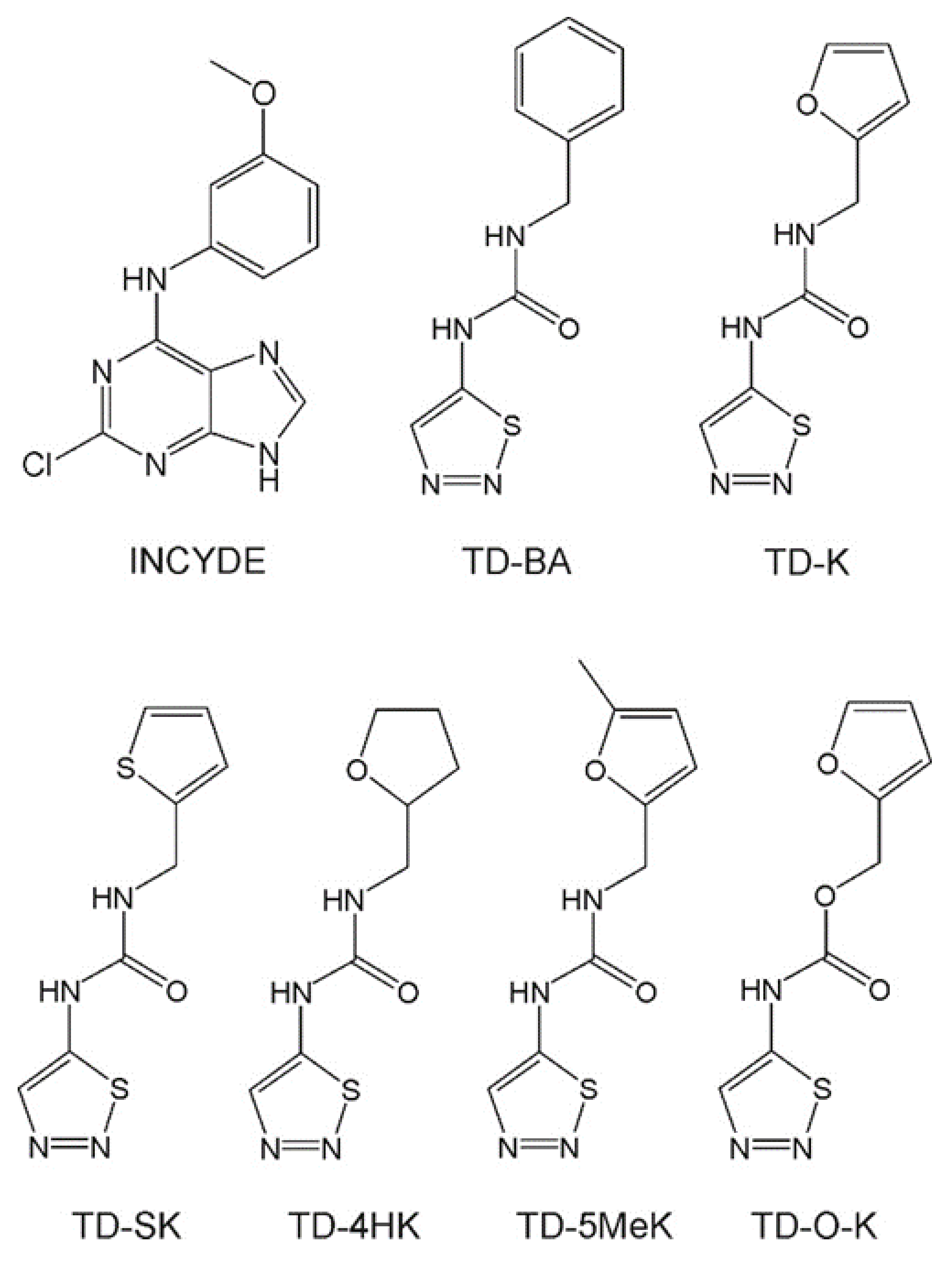
2.1. Design and Chemistry of Compounds
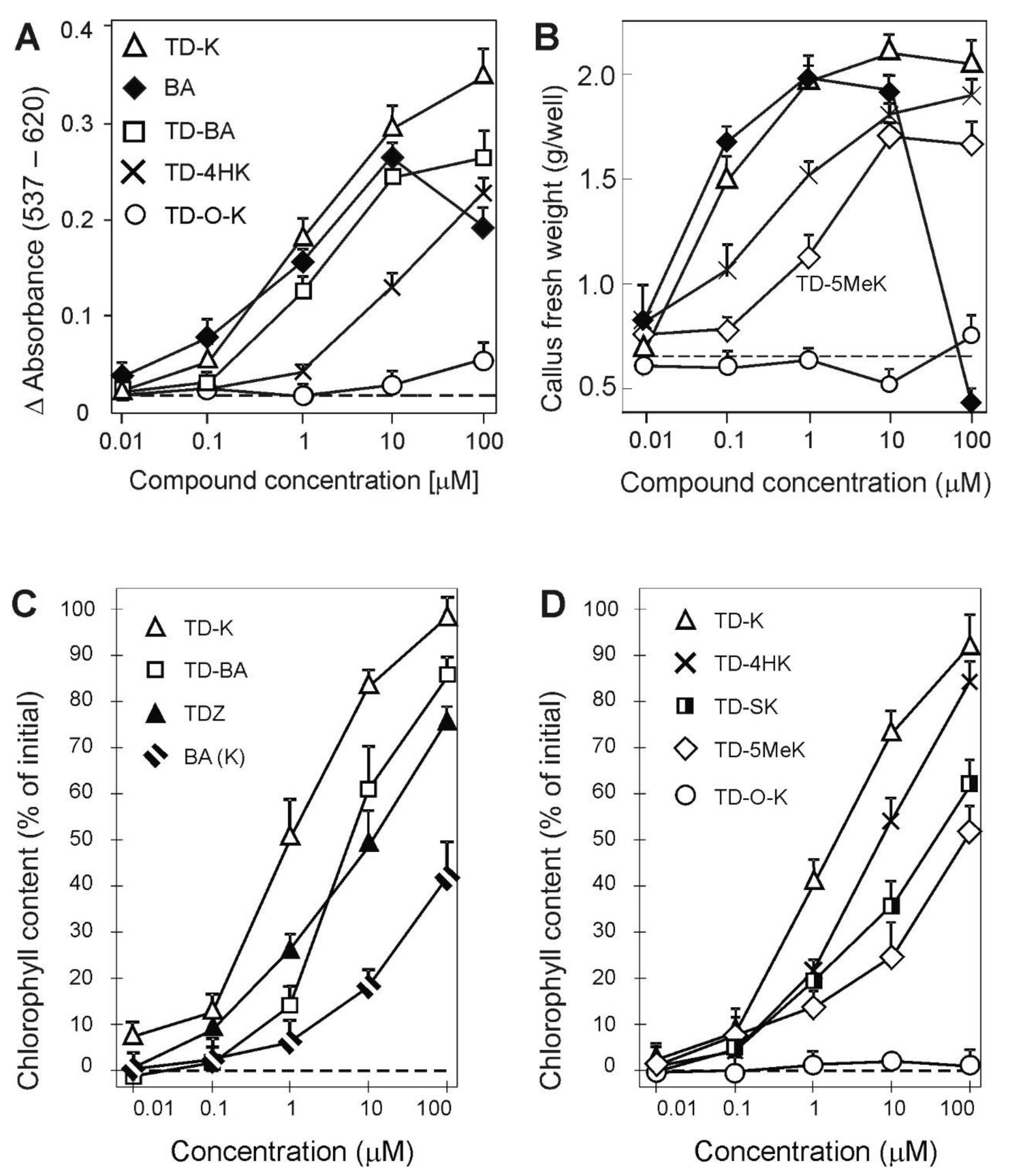
2.2. Activity in Cytokinin Bioassays
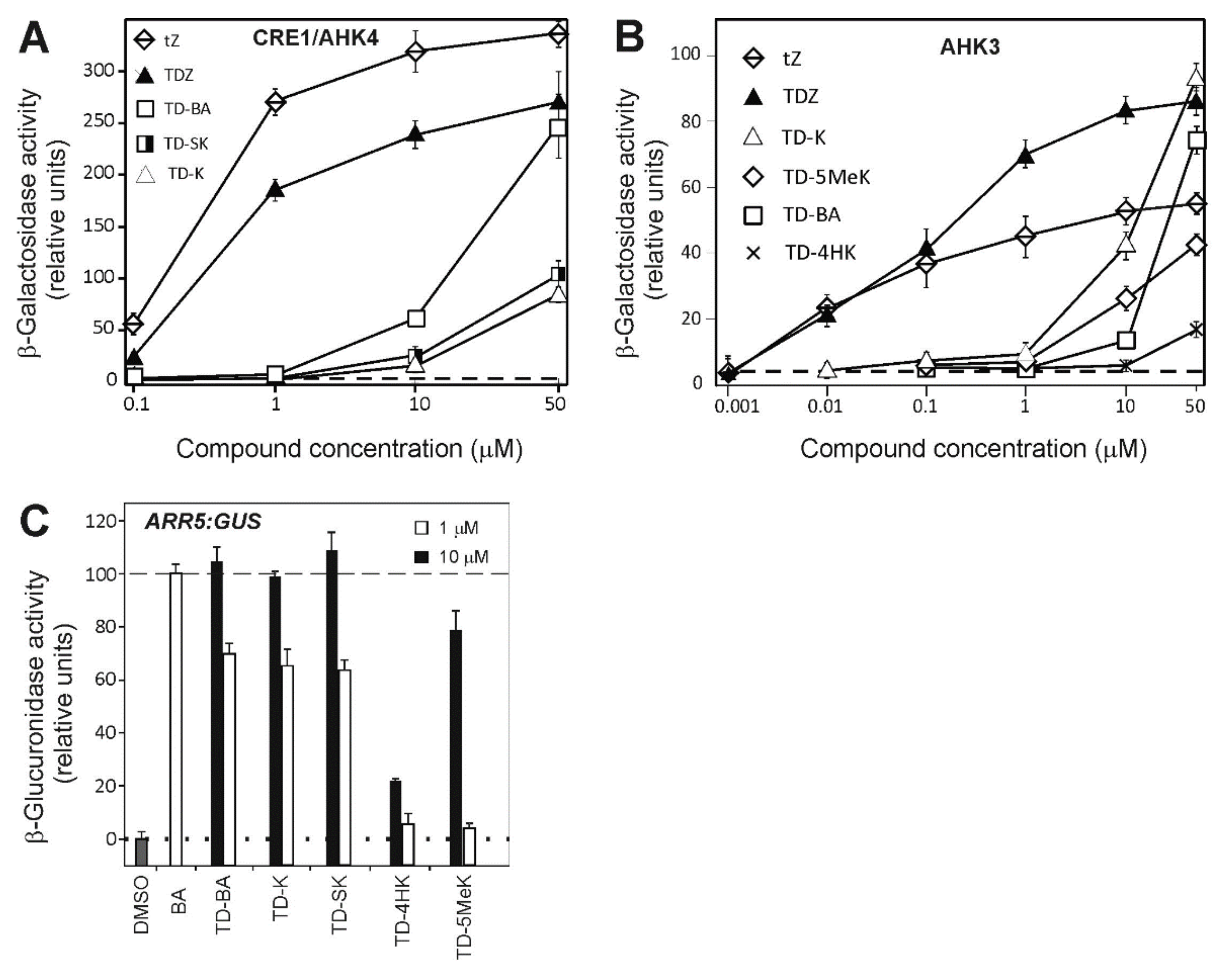
2.3. Activation of Arabidopsis Receptors AHK3 and CRE1/AHK4 and the Cytokinin Primary Response Gene ARR5
2.4. PGR-Treated RCBr
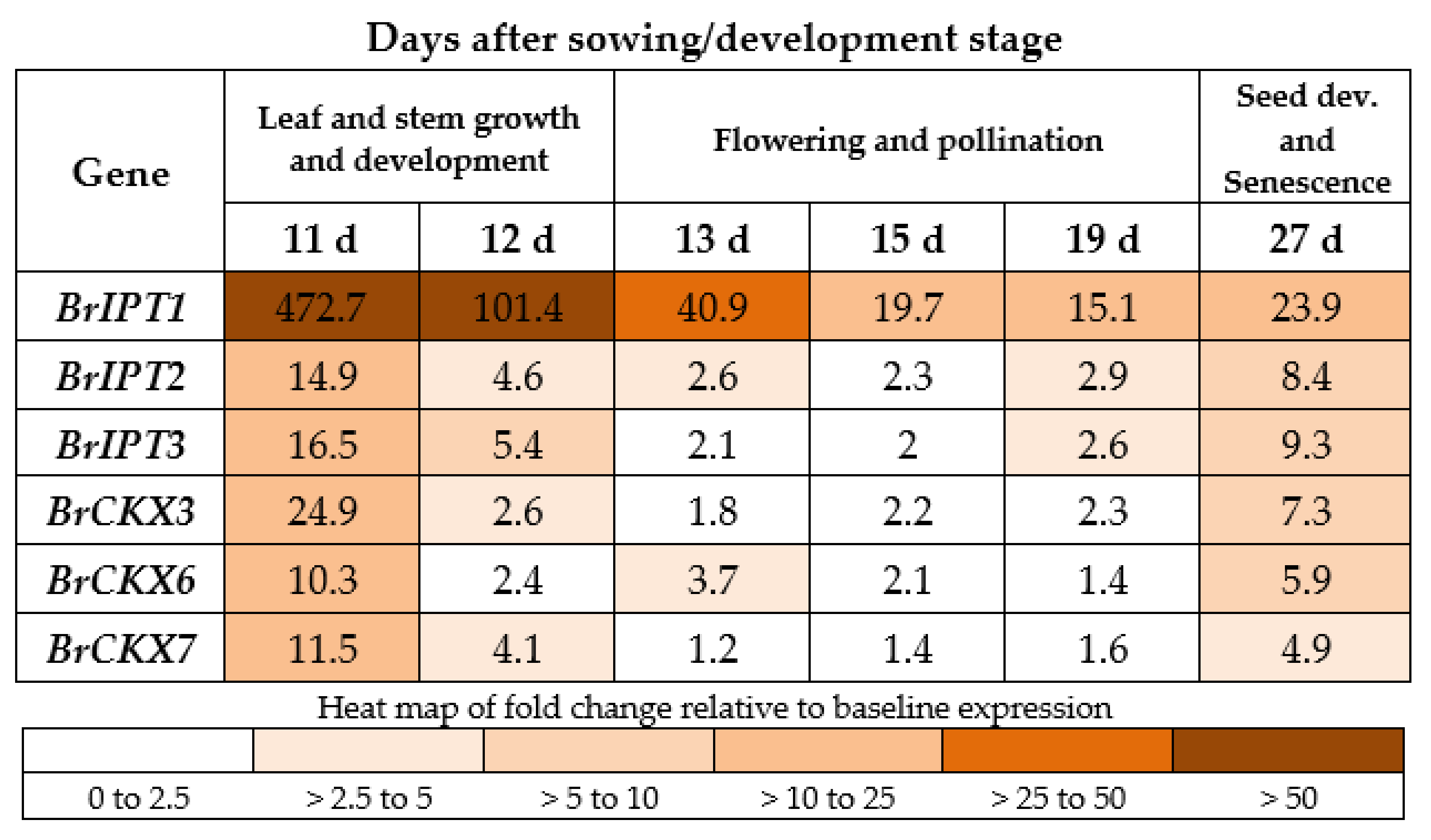
2.5. BrCKX and BrIPT Expression Following True Leaf Appearance
2.6. BrCKX and BrIPT Expression Following INCYDE Treatment
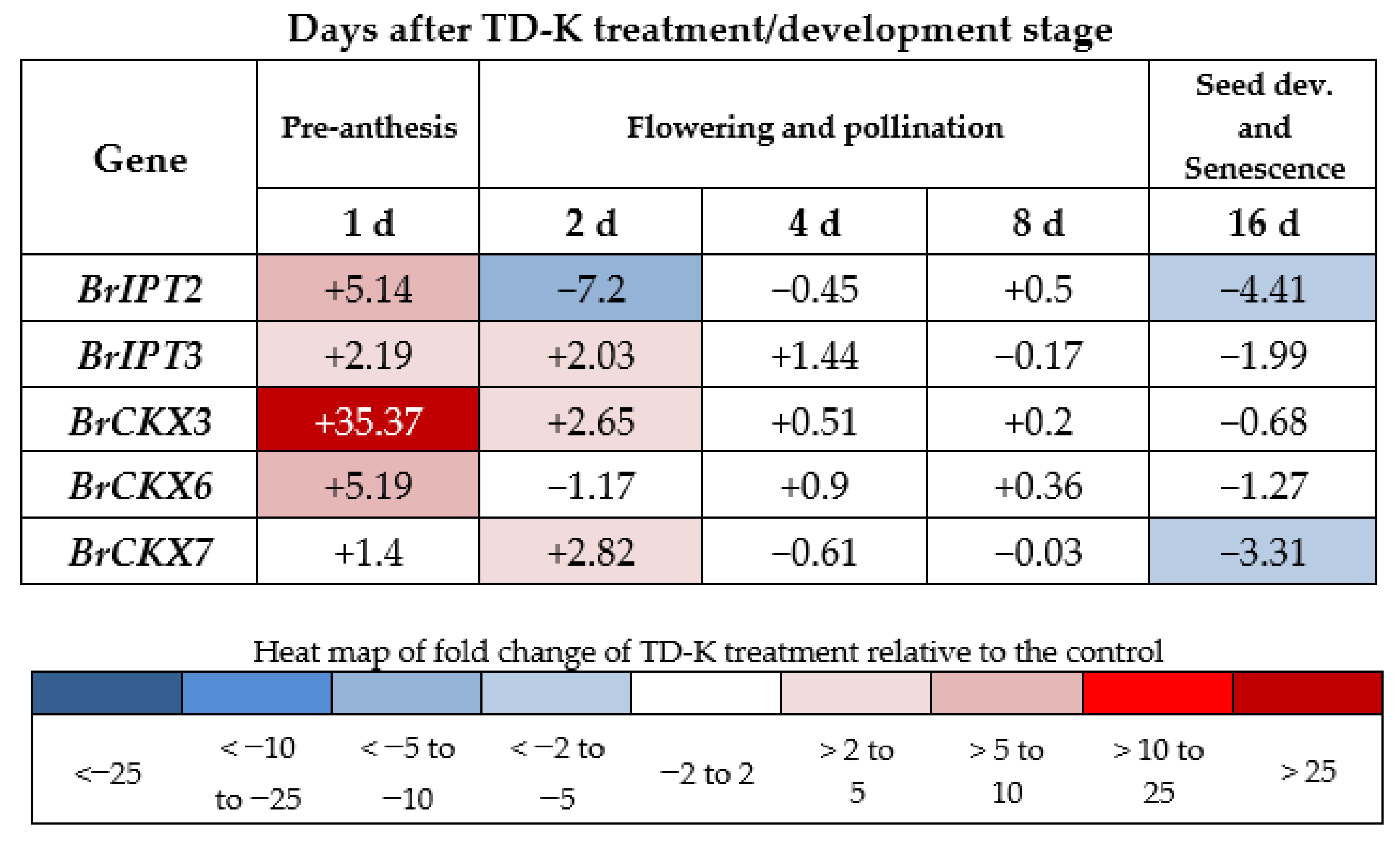
2.7. BrCKX and BrIPT Expression Following TD-K Treatment
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Chemical Synthesis
5.2. Cytokinin Bioassays, Cytokinin Receptor Activation Assay and Arabidopsis ARR5:GUS Reporter Gene Assay
5.3. Plant Material
5.4. Gene Expression
5.4.1. RNA Isolation
5.4.2. cDNA Synthesis
5.4.3. Reverse Transcriptase Quantitative PCR
5.4.4. Expression Analyses
5.5. Experimental Design and Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Agbogidi, O.M. Global Climate Change: A Threat to Food Security and Environmental Conservation. Br. J. Environ. Clim. Chang. 2011, 1, 74–89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manning, D.A. How will minerals feed the world in 2050? Proc. Geol. Assoc. 2015, 126, 14–17. [Google Scholar] [CrossRef]
- Jameson, P.E.; Song, J. Cytokinin: A key driver of seed yield. J. Exp. Bot. 2016, 67, 593–606. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Werner, T.; Motyka, V.; Laucou, V.; Smets, R.; Van Onckelen, H.; Schmülling, T. Cytokinin-deficient transgenic Arabidopsis plants show multiple developmental alterations indicating opposite functions of cytokinins in the regulation of shoot and root meristem activity. Plant Cell 2003, 15, 2532–2550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barbier, F.F.; Dun, E.A.; Kerr, S.C.; Chabikwa, T.G.; Beveridge, C.A. An update on the signals controlling shoot branching. Trends Plant Sci. 2019, 24, 220–236. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Gan, S.S. Translational researches on leaf senescence for enhancing plant productivity and quality. J. Exp. Bot. 2014, 65, 3901–3913. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hönig, M.; Plíhalová, L.; Husičková, A.; Nisler, J.; Doležal, K. Role of cytokinins in senescence, antioxidant defence and photosynthesis. Int. J. Mol. Sci. 2018, 19, 4045. [Google Scholar] [CrossRef] [Green Version]
- Wybouw, B.; De Rybel, B. Cytokinin—A Developing Story. Trends Plant. Sci. 2019, 24, 177–185. [Google Scholar] [CrossRef]
- Cortleven, A.; Leuendorf, J.E.; Frank, M.; Pezzetta, D.; Bolt, S.; Schmülling, T. Cytokinin action in response to abiotic and biotic stresses in plants. Plant Cell Environ. 2019, 42, 998–1018. [Google Scholar] [CrossRef]
- Gu, J.; Li, Z.; Mao, Y.; Struik, P.C.; Zhang, H.; Liu, L.; Wang, Z.; Yang, J. Roles of nitrogen and cytokinin signals in root and shoot communications in maximizing of plant productivity and their agronomic applications. Plant Sci. 2018, 274, 320–331. [Google Scholar] [CrossRef]
- Strnad, M. The aromatic cytokinins. Physiol. Plant. 1997, 101, 674–688. [Google Scholar] [CrossRef]
- Mok, D.W.; Mok, D.C. Cytokinin metabolism and action. Annu. Rev. Plant Biol. 2001, 52, 89–118. [Google Scholar] [CrossRef] [PubMed]
- Lomin, S.N.; Krivosheev, D.M.; Steklov, M.Y.; Arkhipov, D.V.; Osolodkin, D.I.; Schmülling, T.; Romanov, G.A. Plant membrane assays with cytokinin receptors underpin the unique role of free cytokinin bases as biologically active ligands. J. Exp. Bot. 2015, 66, 1851–1863. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nishii, K.; Wright, F.; Chen, Y.Y.; Möller, M. Tangled history of a multigene family: The evolution of ISOPENTENYLTRANSFERASE genes. PLoS ONE 2018, 13, e0201198. [Google Scholar] [CrossRef]
- Gajdošová, S.; Spíchal, L.; Kamínek, M.; Hoyerová, K.; Novák, O.; Dobrev, P.I.; Galuszka, P.; Klíma, P.; Gaudinová, A.; Žižková, E.; et al. Distribution, biological activities, metabolism, and the conceivable function of cis-zeatin-type cytokinins in plants. J. Exp. Bot. 2011, 62, 2827–2840. [Google Scholar] [CrossRef] [Green Version]
- Schäfer, M.; Brütting, C.; Meza-Canales, I.D.; Großkinsky, D.K.; Vankova, R.; Baldwin, I.T.; Meldau, S. The role of cis-zeatin-type cytokinins in plant growth regulation and mediating responses to environmental interactions. J. Exp. Bot. 2015, 66, 4873–4884. [Google Scholar] [CrossRef] [Green Version]
- Spíchal, L. Cytokinins—Recent news and views of evolutionally old molecules. Funct. Plant Biol. 2012, 39, 267–284. [Google Scholar] [CrossRef] [Green Version]
- Spíchal, L.; Rakova, N.Y.; Riefler, M.; Mizuno, T.; Romanov, G.A.; Strnad, M.; Schmülling, T. Two cytokinin receptors of Arabidopsis thaliana, CRE1/AHK4 and AHK3, differ in their ligand specificity in a bacterial assay. Plant Cell Physiol. 2004, 45, 1299–1305. [Google Scholar] [CrossRef] [Green Version]
- Zatloukal, M.; Gemrotová, M.; Doležal, K.; Havlíček, L.; Spíchal, L.; Strnad, M. Novel potent inhibitors of A. thaliana cytokinin oxidase/dehydrogenase. Bioorg. Med. Chem. 2008, 16, 9268–9275. [Google Scholar] [CrossRef]
- Kopečný, D.; Briozzo, P.; Popelková, H.; Šebela, M.; Končitíková, R.; Spíchal, L.; Nisler, J.; Madzak, C.; Frébort, I.; Laloue, M.; et al. Phenyl- and benzylurea cytokinins as competitive inhibitors of cytokinin oxidase/dehydrogenase: A structural study. Biochimie 2010, 92, 1052–1062. [Google Scholar] [CrossRef]
- Miyawaki, K.; Tarkowski, P.; Matsumoto-Kitano, M.; Kato, T.; Sato, S.; Tarkowska, D.; Tabata, S.; Sandberg, G.; Kakimoto, T. Roles of Arabidopsis ATP/ADP isopentenyltransferases and tRNA isopentenyltransferases in cytokinin biosynthesis. Proc. Natl. Acad. Sci. USA 2006, 103, 16598–16603. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kakimoto, T. Identification of plant cytokinin biosynthetic enzymes as dimethylallyl diphosphate: ATP/ADP isopentenyltransferases. Plant. Cell Physiol. 2001, 42, 677–685. [Google Scholar] [CrossRef] [PubMed]
- Takei, K.; Sakakibara, H.; Tatsuo, S. Identification of genes encoding adenylate isopentenyltransferase, a cytokinin biosynthesis enzyme, in Arabidopsis thaliana. J. Biol. Chem. 2001, 276, 26405–26410. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miyawaki, K.; Matsumoto-Kitano, M.; Kakimoto, T. Expression of cytokinin biosynthetic isopentenyltransferase genes in Arabidopsis: Tissue specificity and regulation by auxin, cytokinin, and nitrate. Plant J. 2004, 37, 128–138. [Google Scholar] [CrossRef] [PubMed]
- Kurakawa, T.; Ueda, N.; Masahiko, M.; Kaoru, K.; Kojima, M.; Nagato, Y.; Sakakibara, H.; Junko, K. Direct control of shoot meristem activity by a cytokinin-activating enzyme. Nature 2007, 445, 652–655. [Google Scholar] [CrossRef] [PubMed]
- Werner, T.; Köllmer, I.; Bartrina, I.; Holst, K.; Schmülling, T. New insights into the biology of cytokinin degradation. Plant Biol. 2006, 8, 371–381. [Google Scholar] [CrossRef]
- Kieber, J.J.; Schaller, G.E. Cytokinin signaling in plant development. Development 2018, 145. [Google Scholar] [CrossRef] [Green Version]
- Galuszka, P.; Popelková, H.; Werner, T.; Frébortová, J.; Pospíšilová, H.; Mik, V.; Köllmer, I.; Schmülling, T.; Frébort, I. Biochemical characterization of cytokinin oxidases/dehydrogenases from Arabidopsis thaliana expressed in Nicotiana tabacum L. J. Plant Growth Regul. 2007, 26, 255–267. [Google Scholar] [CrossRef]
- Zalabák, D.; Galuszka, P.; Mrízová, K.; Podlešáková, K.; Gu, R.; Frébortová, J. Biochemical characterization of the maize cytokinin dehydrogenase family and cytokinin profiling in developing maize plantlets in relation to the expression of cytokinin dehydrogenase genes. Plant. Physiol. Biochem. 2014, 74, 283–293. [Google Scholar] [CrossRef]
- Schmülling, T.; Werner, T.; Riefler, M.; Krupková, E.; Manns, I.B. Structure and function of cytokinin oxidase/dehydrogenase genes of maize, rice, Arabidopsis and other species. J. Plant Res. 2003, 116, 241–252. [Google Scholar] [CrossRef]
- Chen, L.; Zhao, J.; Song, J.; Jameson, P.E. Cytokinin dehydrogenase: A genetic target for yield improvement in wheat. Plant Biotechnol. J. 2020, 18, 614–630. [Google Scholar] [CrossRef] [PubMed]
- Werner, T.; Nehnevajova, E.; Köllmer, I.; Novák, O.; Strnad, M.; Krämer, U.; Schmülling, T. Root-specific reduction of cytokinin causes enhanced root growth, drought tolerance, and leaf mineral enrichment in Arabidopsis and tobacco. Plant Cell 2010, 22, 3905–3920. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Riefler, M.; Novak, O.; Strnad, M.; Schmülling, T. Arabidopsis cytokinin receptor mutants reveal functions in shoot growth, leaf senescence, seed size, germination, root development, and cytokinin metabolism. Plant Cell 2006, 18, 40–54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gemrotová, M.; Kulkarni, M.G.; Stirk, W.A.; Strnad, M.; Van Staden, J.; Spíchal, L. Seedlings of medicinal plants treated with either a cytokinin antagonist (PI-55) or an inhibitor of cytokinin degradation (INCYDE) are protected against the negative effects of cadmium. Plant Growth Regul. 2013, 71, 137–145. [Google Scholar] [CrossRef]
- Nisler, J.; Kopečný, D.; Pěkná, Z.; Končitíková, R.; Koprna, R.; Murvanidze, N.; Werbrouck, S.P.; Havlíček, L.; De Diego, N.; Kopečná, M.; et al. Diphenylurea-derived cytokinin oxidase/dehydrogenase inhibitors for biotechnology and agriculture. J. Exp. Bot. 2020. [Google Scholar] [CrossRef] [PubMed]
- Bartrina, I.; Otto, E.; Strnad, M.; Werner, T.; Schmülling, T. Cytokinin regulates the activity of reproductive meristems, flower organ size, ovule formation, and thus seed yield in Arabidopsis thaliana. Plant Cell 2011, 23, 69–80. [Google Scholar] [CrossRef] [Green Version]
- Schwarz, I.; Scheirlinck, M.T.; Otto, E.; Bartrina, I.; Schmidt, R.C.; Schmülling, T. Cytokinin regulates the activity of the inflorescence meristem and components of seed yield in oilseed rape. J. Exp. Bot. 2020, 71. [Google Scholar] [CrossRef]
- Ashikari, M.; Sakakibara, H.; Lin, S.; Yamamoto, T.; Takashi, T.; Nishimura, A.; Angeles, E.R.; Qian, Q.; Kitano, H.; Matsuoka, M. Plant science: Cytokinin oxidase regulates rice grain production. Science 2005, 309, 741–745. [Google Scholar] [CrossRef]
- Zalewski, W.; Gasparis, S.; Boczkowska, M.; Rajchel, I.K.; Kała, M.; Orczyk, W.; Nadolska-Orczyk, A. Expression patterns of HvCKX genes indicate their role in growth and reproductive development of barley. PLoS ONE 2014, 9, e115729. [Google Scholar] [CrossRef] [Green Version]
- Holubová, K.; Hensel, G.; Vojta, P.; Tarkowski, P.; Bergougnoux, V.; Galuszka, P. Modification of barley plant productivity through regulation of cytokinin content by reverse-genetics approaches. Front. Plant Sci. 2018, 871, 1676. [Google Scholar] [CrossRef] [Green Version]
- Jabłoński, B.; Ogonowska, H.; Szala, K.; Bajguz, A.; Orczyk, W.; Nadolska-Orczyk, A. Silencing of TaCKX1 mediates expression of other TaCKX genes to increase yield parameters in wheat. Int. J. Mol. Sci. 2020, 21, 4809. [Google Scholar] [CrossRef] [PubMed]
- Gasparis, S.; Przyborowski, M.; Kała, M.; Nadolska-Orczyk, A. Knockout of the HvCKX1 or HvCKX3 gene in barley (Hordeum vulgare L.) by RNA-Guided Cas9 Nuclease affects the regulation of cytokinin metabolism and root morphology. Cells 2019, 8, 782. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fritsche, S.; Poovaiah, C.; MacRae, E.; Thorlby, G. A New Zealand perspective on the application and regulation of gene editing. Front. Plant Sci. 2018, 9, 1323. [Google Scholar] [CrossRef] [PubMed]
- Suttle, J.C. Involvement of ethylene in the action of the cotton defoliant thidiazuron. Plant Physiol. 1985, 78, 272–276. [Google Scholar] [CrossRef]
- Grossmann, K. Induction of leaf abscission in cotton is a common effect of urea- and adenine-type cytokinins. Plant Physiol. 1991, 95, 234–237. [Google Scholar] [CrossRef] [Green Version]
- Nishijima, T.; Shima, K. Change in flower morphology of Torenia fournieri Lind. induced by forchlorfenuron application. Sci. Hortic. (Amst.) 2006, 109, 254–261. [Google Scholar] [CrossRef]
- Lewis, D.H.; Burge, G.K.; Hopping, M.E.; Jameson, P.E. Cytokinins and fruit development in the kiwifruit (Actinidia deliciosa). II. Effects of reduced pollination and CPPU application. Physiol. Plant 1996, 98, 187–195. [Google Scholar] [CrossRef]
- Stern, R.A.; Ben-Arie, R.; Neria, O.; Flaishman, M. CPPU and BA increase fruit size of ‘Royal Gala’ (Malus domestica) apple in a warm climate. J. Hortic. Sci. Biotechnol. 2003, 78, 297–302. [Google Scholar] [CrossRef]
- Ferrara, G.; Mazzeo, A.; Netti, G.; Pacucci, C.; Matarrese, A.M.S.; Cafagna, I.; Mastrorilli, P.; Vezzoso, M.; Gallo, V. Girdling, gibberellic acid, and forchlorfenuron: Effects on yield, quality, and metabolic profile of table grape cv. Italia. Am. J. Enol. Vitic. 2014, 65, 381–387. [Google Scholar] [CrossRef] [Green Version]
- Nisler, J.; Kopečný, D.; Končitíková, R.; Zatloukal, M.; Bazgier, V.; Berka, K.; Zalabák, D.; Briozzo, P.; Strnad, M.; Spíchal, L. Novel thidiazuron-derived inhibitors of cytokinin oxidase/dehydrogenase. Plant. Mol. Biol. 2016, 92, 235–248. [Google Scholar] [CrossRef]
- Aremu, A.O.; Masondo, N.A.; Sunmonu, T.O.; Kulkarni, M.G.; Zatloukal, M.; Spichal, L.; Doležal, K.; Van Staden, J. A novel inhibitor of cytokinin degradation (INCYDE) influences the biochemical parameters and photosynthetic apparatus in NaCl-stressed tomato plants. Planta 2014, 240, 877–889. [Google Scholar] [CrossRef] [PubMed]
- Reusche, M.; Klásková, J.; Thole, K.; Truskina, J.; Novák, O.; Janz, D.; Strnad, M.; Spíchal, L.; Lipka, V.; Teichmann, T. Stabilization of cytokinin levels enhances Arabidopsis resistance against Verticillium Longisporum. Mol. Plant Microbe Interact. 2013, 26, 850–860. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berková, V.; Kameniarová, M.; Ondrisková, V.; Berka, M.; Menšíková, S.; Kopecká, R.; Luklová, M.; Novák, J.; Spíchal, L.; Rashotte, A.M.; et al. Arabidopsis response to Inhibitor of Cytokinin Degradation INCYDE: Modulations of cytokinin signaling and plant proteome. Plants 2020, 1563. [Google Scholar] [CrossRef]
- Aremu, A.O.; Stirk, W.A.; Masondo, N.A.; Plačková, L.; Novák, O.; Pěnčík, A.; Zatloukal, M.; Nisler, J.; Spíchal, L.; Doležal, K.; et al. Dissecting the role of two cytokinin analogues (INCYDE and PI-55) on in vitro organogenesis, phytohormone accumulation, phytochemical content and antioxidant activity. Plant Sci. 2015, 238, 81–94. [Google Scholar] [CrossRef] [PubMed]
- Smýkalová, I.; Vrbová, M.; Cvečková, M.; Plačková, L.; Žukauskaitė, A.; Zatloukal, M.; Hrdlička, J.; Plíhalová, L.; Doležal, K.; Griga, M. The effects of novel synthetic cytokinin derivatives and endogenous cytokinins on the in vitro growth responses of hemp (Cannabis sativa L.) explants. Plant Cell Tissue Organ. Cult. 2019, 139, 381–394. [Google Scholar] [CrossRef]
- Aremu, A.O.; Bairu, M.W.; Novák, O.; Plačková, L.; Zatloukal, M.; Doležal, K.; Finnie, J.F.; Strnad, M.; Van Staden, J. Physiological responses and endogenous cytokinin profiles of tissue-cultured “Williams” bananas in relation to roscovitine and an inhibitor of cytokinin oxidase/dehydrogenase (INCYDE) treatments. Planta 2012, 236, 1775–1790. [Google Scholar] [CrossRef] [PubMed]
- Antoniadi, I.; Novák, O.; Gelovád, Z.; Johnson, A.; Plíhal, O.; Vain, T.; Simerský, R.; Mik, V.; Karady, M.; Pernisová, M. Cell-surface receptors enable perception of extracellular cytokinins. Nat. Commun. 2020, 11, 1–10. [Google Scholar] [CrossRef]
- Goh, D.M.; Cosme, M.; Kisiala, A.B.; Mulholland, S.; Said, Z.M.F.; Spíchal, L.; Emery, R.J.N.; Declerck, S.; Guinel, F.C. A stimulatory role for cytokinin in the arbuscular mycorrhizal symbiosis of pea. Front. Plant Sci. 2019, 10, 262. [Google Scholar] [CrossRef]
- Prerostova, S.; Dobrev, P.I.; Kramna, B.; Gaudinova, A.; Knirsch, V.; Spíchal, L.; Zatloukal, M.; Vankova, R. Heat acclimation and inhibition of cytokinin degradation positively affect heat stress tolerance of Arabidopsis. Front. Plant Sci. 2020, 11. [Google Scholar] [CrossRef] [Green Version]
- Nisler, J.; Zatloukal, M.; Spíchal, L.; Koprna, R.; Doležal, K.; Strnad, M. 1, 2, 3-thiadiazol-5yl-urea Derivatives, Use Thereof for Regulating Plant Senescence and Preparations Containing These Derivatives. U.S. Patent 9,993,002, 12 June 2018. [Google Scholar]
- O’Keefe, D.; Song, J.; Jameson, P.E. Isopentenyl transferase and cytokinin oxidase/dehydrogenase gene family members are differentially expressed during pod and seed development in rapid-cycling Brassica. J. Plant Growth Regul. 2011, 30, 92–99. [Google Scholar] [CrossRef]
- Nisler, J.; Zatloukal, M.; Sobotka, R.; Pilný, J.; Zdvihalová, B.; Novák, O.; Strnad, M.; Spíchal, L. New urea derivatives are effective anti-senescence compounds acting most likely via a cytokinin-independent mechanism. Front. Plant Sci. 2018, 9. [Google Scholar] [CrossRef] [PubMed]
- Mok, M.C.; Mok, D.W.S.; Armstrong, D.J.; Shudo, K.; Isogai, Y.; Okamoto, T. Cytokinin activity of N-phenyl-N’-1,2,3-thiadiazol-5-ylurea (Thidiazuron). Phytochemistry 1982, 21, 1509–1511. [Google Scholar] [CrossRef]
- Yip, W.-K.; Yang, S.F. Effect of thidiazuron, a cytokinin-active urea derivative, in cytokinin-dependent ethylene production systems. Plant Physiol. 1986, 80, 515–519. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bamberger, E.; Mayer, A.M. Effect of kinetin on formation of red pigment in seedlings of Amaranthus retroflexus. Science 1960, 131, 1094–1095. [Google Scholar] [CrossRef]
- Abad, A.; Agulló, C.; Cuñat, A.C.; Jiménez, R.; Vilanova, C. Preparation and promotion of fruit growth in kiwifruit of fluorinated N-phenyl-N’-1,2,3-thiadiazol-5-yl ureas. J. Agric. Food Chem. 2004, 52, 4675–4683. [Google Scholar] [CrossRef]
- Boyes, D.C.; Zayed, A.M.; Ascenzi, R.; McCaskill, A.J.; Hoffman, N.E.; Davis, K.R.; Görlach, J. Growth stage-based phenotypic analysis of Arabidopsis: A model for high throughput functional genomics in plants. Plant Cell 2001, 13, 1499–1510. [Google Scholar] [CrossRef] [Green Version]
- Koprna, R.; De Diego, N.; Dundálková, L.; Spíchal, L. Use of cytokinins as agrochemicals. Bioorg. Med. Chem. 2016, 24, 484–492. [Google Scholar] [CrossRef]
- Zalewski, W.; Galuszka, P.; Gasparis, S.; Orczyk, W.; Nadolska-Orczyk, A. Silencing of the HvCKX1 gene decreases the cytokinin oxidase/dehydrogenase level in barley and leads to higher plant productivity. J. Exp. Bot. 2010, 61, 1839–1851. [Google Scholar] [CrossRef] [Green Version]
- Chatfield, J.M.; Armstrong, D.J. Regulation of cytokinin oxidase activity in callus tissues of Phaseolus vulgaris L. cv Great Northern. Plant Physiol. 1986, 80, 493–499. [Google Scholar] [CrossRef] [Green Version]
- Kamínek, M.; Armstrong, D.J. Genotypic variation in cytokinin oxidase from Phaseolus callus cultures. Plant Physiol. 1990, 93, 1530–1538. [Google Scholar] [CrossRef] [Green Version]
- Motyka, V.; Vaňková, R.; Čapková, V.; Petrášek, J.; Kamínek, M.; Schmülling, T. Cytokinin-induced upregulation of cytokinin oxidase activity in tobacco includes changes in enzyme glycosylation and secretion. Physiol. Plant 2003, 117, 11–21. [Google Scholar] [CrossRef]
- Blagoeva, E.; Dobrev, P.I.; Malbeck, J.; Motyka, V.; Gaudinová, A.; Vaňková, R. Effect of exogenous cytokinins, auxins and adenine on cytokinin N-glucosylation and cytokinin oxidase/dehydrogenase activity in de-rooted radish seedlings. Plant Growth Regul. 2004, 44, 15–23. [Google Scholar] [CrossRef]
- Liu, Z.; Lv, Y.; Zhang, M.; Liu, Y.; Kong, L.; Zou, M.; Lu, G.; Cao, J.; Yu, X. Identification, expression, and comparative genomic analysis of the IPT and CKX gene families in Chinese cabbage (Brassica rapa ssp pekinensis). BMC Genom. 2013, 14, 594. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brugière, N.; Jiao, S.; Hantke, S.; Zinselmeier, C.; Roessler, J.A.; Niu, X.; Jones, R.J.; Habben, J.E. Cytokinin oxidase gene expression in maize is localized to the vasculature, and is induced by cytokinins, abscisic acid, and abiotic stress. Am. Soc. Plant Biol. 2003, 132, 1228–1240. [Google Scholar] [CrossRef] [Green Version]
- Song, J.; Jiang, L.; Jameson, P.E. Co-ordinate regulation of cytokinin gene family members during flag leaf and reproductive development in wheat. BMC Plant Biol. 2012, 12, 78. [Google Scholar] [CrossRef] [Green Version]
- Liu, P.; Zhang, C.; Ma, J.Q.; Zhang, L.Y.; Yang, B.; Tang, X.Y.; Huang, L.; Zhou, X.T.; Lu, K.; Li, J.N. Genome-wide identification and expression profiling of cytokinin oxidase/dehydrogenase (CKX) genes reveal likely roles in pod development and stress responses in oilseed rape (Brassica napus L.). Genes 2018, 9, 168. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Ma, X.M.; Kojima, M.; Sakakibara, H.; Hou, B.K. N-glucosyltransferase UGT76C2 is involved in cytokinin homeostasis and cytokinin response in Arabidopsis thaliana. Plant Cell Physiol. 2011, 52, 2200–2213. [Google Scholar] [CrossRef] [Green Version]
- Goldschmidt, H.; Bardach, B. Zur Kenntniss der Diazoamidokörper. Berichte der Deutschen Chemischen Gesellschaft 1892, 25, 1347–1378. [Google Scholar] [CrossRef] [Green Version]
- Holub, J.; Hanuš, J.; Hanke, D.E.; Strnad, M. Biological activity of cytokinins derived from ortho- and meta-hydroxybenzyladenine. Plant Growth Regul. 1998, 26, 109–115. [Google Scholar] [CrossRef]
- Song, J.; Jiang, L.; Jameson, P.E. Expression patterns of Brassica napus genes implicate IPT, CKX, sucrose transporter, cell wall invertase, and amino acid permease gene family members in leaf, flower, silique, and seed development. J. Exp. Bot. 2015, 66, 5067–5082. [Google Scholar] [CrossRef] [Green Version]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 2001, 29. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Trait | Control | INCYDE |
|---|---|---|
| Shoot DW (mg) | 183.9 ± 96.9 a | 186.2 ± 91.9 a |
| Branch number | 1.6 ± 0.03 a | 1.8 ± 0.09 a |
| Silique number | 5.2 ± 1.4 a | 7.0 ± 2.3 a |
| Silique length (mm) | 37.2 ± 1.6 a | 39.7 ± 0.8 a |
| Silique mass (mg) | 50.8 ± 2.1 a | 46.6 ± 1.2 a |
| Seeds per silique | 11.9 ± 1.2 a | 12.3 ± 2.6 a |
| Seeds per plant | 54.3 ± 7.0 b | 74.5 ± 4.8 a |
| Seed mass (mg) | 1.6 ± 0.8 a | 2.3 ± 0.4 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
van Voorthuizen, M.J.; Nisler, J.; Song, J.; Spíchal, L.; Jameson, P.E. Targeting Cytokinin Homeostasis in Rapid Cycling Brassica rapa with Plant Growth Regulators INCYDE and TD-K. Plants 2021, 10, 39. https://doi.org/10.3390/plants10010039
van Voorthuizen MJ, Nisler J, Song J, Spíchal L, Jameson PE. Targeting Cytokinin Homeostasis in Rapid Cycling Brassica rapa with Plant Growth Regulators INCYDE and TD-K. Plants. 2021; 10(1):39. https://doi.org/10.3390/plants10010039
Chicago/Turabian Stylevan Voorthuizen, Matthew J., Jaroslav Nisler, Jiancheng Song, Lukáš Spíchal, and Paula E. Jameson. 2021. "Targeting Cytokinin Homeostasis in Rapid Cycling Brassica rapa with Plant Growth Regulators INCYDE and TD-K" Plants 10, no. 1: 39. https://doi.org/10.3390/plants10010039