
Features of Peripheral Blood Th-Cell Subset Composition and Serum Cytokine Level in Patients with Activity-Driven Ankylosing Spondylitis
 , ,
, ,
Abstract
:1. Introduction
2. Results
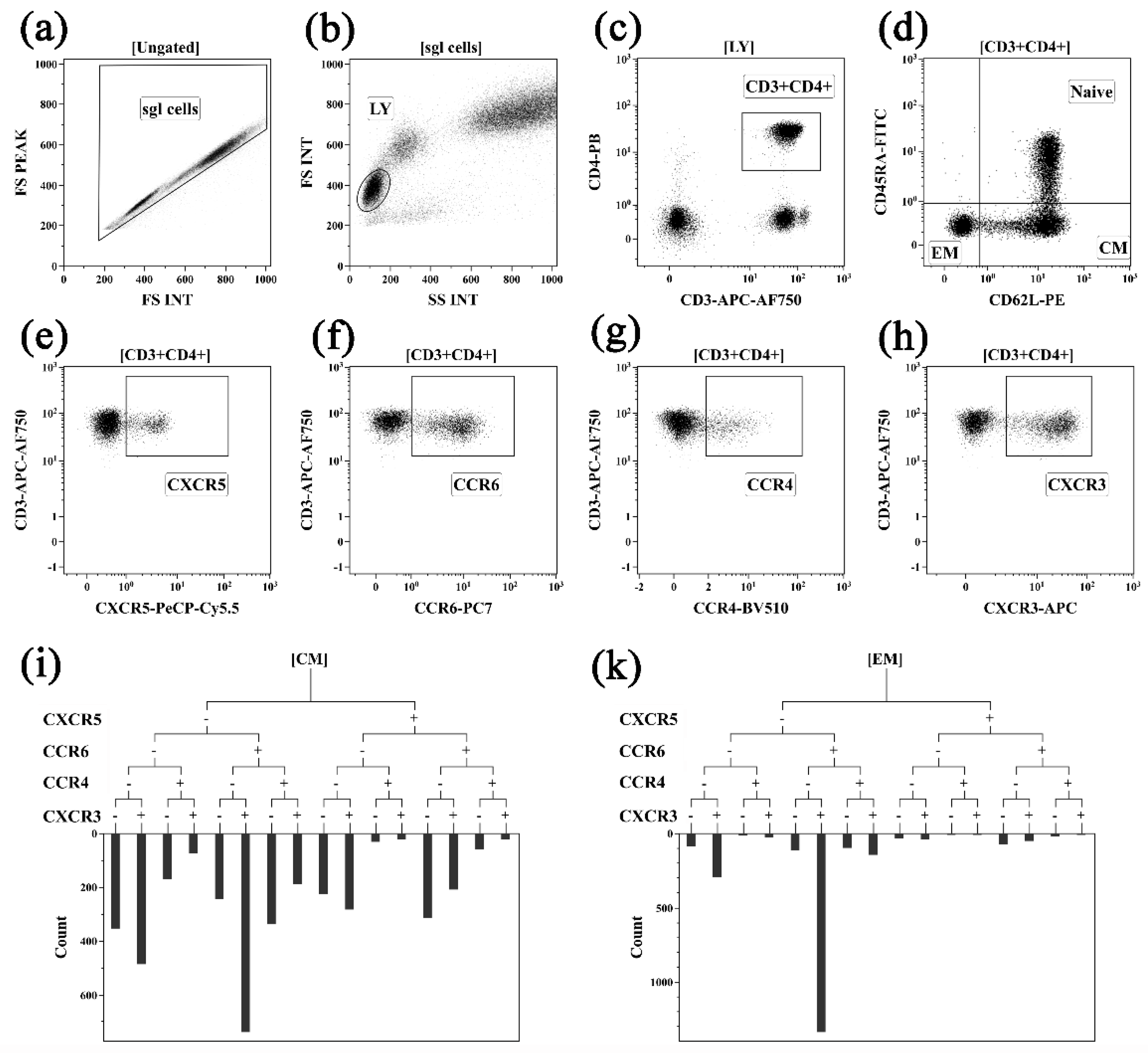
2.1. Assessing Peripheral Blood Th Cell Subset Composition in Activity-Driven AS Patients
2.2. Serum Cytokine Profile in Activity-Driven AS Patients
2.3. Features of Associations between Th Cell Subsets and Serum Cytokine Levels in Activity-Driven AS Patients
3. Discussion
4. Materials and Methods
4.1. Patients
4.2. Sample Collection
4.3. Th Cell Flow Cytometry Immunophenotyping
4.4. Multiplex Assay
4.5. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chen, B.; Li, J.; He, C.; Li, D.; Tong, W.; Zou, Y.; Xu, W. Role of HLA-B27 in the pathogenesis of ankylosing spondylitis (Review). Mol. Med. Rep. 2017, 15, 1943–1951. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dahmani, C.A.; Benzaoui, A.; Amroun, H.; Mecabih, F.; Sediki, F.Z.; Zemani-Fodil, F.; Fodil, M.; Boughrara, W.; Mecheti, B.; Attal, N.; et al. Association of the HLA-B27 antigen and the CTLA4 gene CT60/rs3087243 polymorphism with ankylosing spondylitis in Algerian population: A case-control study. Int. J. Immunogenet. 2018, 45, 109–117. [Google Scholar] [CrossRef] [PubMed]
- Hu, N.; Liu, D.; Zhao, N.; Wang, X.; Bai, Y.; Sun, H. Lack of association between TNF polymorphism and ankylosing spondylitis susceptibility in HLA-B27-positive population: A meta-analysis. Eur. Spine J. 2021, 30, 2401–2408. [Google Scholar] [CrossRef] [PubMed]
- Bridgewood, C.; Sharif, K.; Sherlock, J.; Watad, A.; McGonagle, D. Interleukin-23 pathway at the enthesis: The emerging story of enthesitis in spondyloarthropathy. Immunol. Rev. 2020, 294, 27–47. [Google Scholar] [CrossRef]
- Chisălău, B.A.; Crînguș, L.I.; Vreju, F.A.; Pârvănescu, C.D.; Firulescu, S.C.; Dinescu, Ș.C.; Ciobanu, D.A.; Tica, A.A.; Sandu, R.E.; Siloși, I.; et al. New insights into IL-17/IL-23 signaling in ankylosing spondylitis (Review). Exp. Ther. Med. 2020, 20, 3493–3497. [Google Scholar] [CrossRef]
- Rezaiemanesh, A.; Abdolmaleki, M.; Abdolmohammadi, K.; Aghaei, H.; Pakdel, F.D.; Fatahi, Y.; Soleimanifar, N.; Zavvar, M.; Nicknam, M.H. Immune cells involved in the pathogenesis of ankylosing spondylitis. Biomed. Pharmacother. 2018, 100, 198–204. [Google Scholar] [CrossRef]
- Blair, H.A. Secukinumab: A Review in Ankylosing Spondylitis. Drugs 2019, 79, 433–443. [Google Scholar] [CrossRef] [Green Version]
- Pedersen, S.J.; Maksymowych, W.P. The Pathogenesis of Ankylosing Spondylitis: An Update. Curr. Rheumatol. Rep. 2019, 21, 58. [Google Scholar] [CrossRef]
- Mauro, D.; Thomas, R.; Guggino, G.; Lories, R.; Brown, M.A.; Ciccia, F. Ankylosing spondylitis: An autoimmune or autoinflammatory disease? Nat. Rev. Rheumatol. 2021, 17, 387–404. [Google Scholar] [CrossRef]
- Šućur, A.; Jajić, Z.; Ikić Matijašević, M.; Stipić Marković, A.; Flegar, D.; Lukač, N.; Kelava, T.; Kovačić, N.; Grčević, D. Combined manual and automated immunophenotypisation identified disease-specific peripheral blood immune subpopulations in rheumatoid arthritis, ankylosing spondylitis and psoriatic arthritis. Clin. Exp. Rheumatol. 2020, 38, 903–916. [Google Scholar] [PubMed]
- Hayashi, E.; Chiba, A.; Tada, K.; Haga, K.; Kitagaichi, M.; Nakajima, S.; Kusaoi, M.; Sekiya, F.; Ogasawara, M.; Yamaji, K.; et al. Involvement of Mucosal-associated Invariant T cells in Ankylosing Spondylitis. J. Rheumatol. 2016, 43, 1695–1703. [Google Scholar] [CrossRef] [PubMed]
- Ciccia, F.; Guggino, G.; Rizzo, A.; Saieva, L.; Peralta, S.; Giardina, A.; Cannizzaro, A.; Sireci, G.; De Leo, G.; Alessandro, R.; et al. Type 3 innate lymphoid cells producing IL-17 and IL-22 are expanded in the gut, in the peripheral blood, synovial fluid and bone marrow of patients with ankylosing spondylitis. Ann. Rheum. Dis. 2015, 74, 1739–1747. [Google Scholar] [CrossRef] [Green Version]
- Mauro, D.; Macaluso, F.; Fasano, S.; Alessandro, R.; Ciccia, F. ILC3 in Axial Spondyloarthritis: The Gut Angle. Curr. Rheumatol. Rep. 2019, 21, 37. [Google Scholar] [CrossRef] [PubMed]
- Min, H.K.; Moon, J.; Lee, S.Y.; Lee, A.R.; Lee, C.R.; Lee, J.; Kwok, S.K.; Cho, M.L.; Park, S.H. Expanded IL-22+ Group 3 Innate Lymphoid Cells and Role of Oxidized LDL-C in the Pathogenesis of Axial Spondyloarthritis with Dyslipidaemia. Immune Netw. 2021, 21, e43. [Google Scholar] [CrossRef] [PubMed]
- Long, S.; Ma, L.; Wang, D.; Shang, X. High frequency of circulating follicular helper T cells is correlated with B cell subtypes in patients with ankylosing spondylitis. Exp. Ther. Med. 2018, 15, 4578–4586. [Google Scholar] [CrossRef] [Green Version]
- Wilbrink, R.; Spoorenberg, A.; Verstappen, G.M.P.J.; Kroese, F.G.M. B Cell Involvement in the Pathogenesis of Ankylosing Spondylitis. Int. J. Mol. Sci. 2021, 22, 13325. [Google Scholar] [CrossRef]
- Subrahmanyam, P.B.; Maecker, H.T. Mass Cytometry Analysis of T-Helper Cells. Methods Mol. Biol. 2021, 2285, 49–63. [Google Scholar] [CrossRef]
- Zhu, J. T Helper Cell Differentiation, Heterogeneity, and Plasticity. Cold Spring Harb. Perspect. Biol. 2018, 10, a030338. [Google Scholar] [CrossRef] [Green Version]
- Li, M.; Zhou, X.; Zhou, L.; Yu, Z.; Fu, L.; Yang, P. Meta-Analysis of Changes in the Number and Proportion of Regulatory T Cells in Patients with Ankylosing Spondylitis. Biomed. Res. Int. 2020, 2020, 8709804. [Google Scholar] [CrossRef]
- An, H.; Li, X.; Li, F.; Gao, C.; Li, X.; Luo, J. The absolute counts of peripheral T lymphocyte subsets in patient with ankylosing spondylitis and the effect of low-dose interleukin-2. Medicine 2019, 98, e15094. [Google Scholar] [CrossRef]
- Jiang, Y.; Yang, M.; Zhang, Y.; Huang, Y.; Wu, J.; Xie, Y.; Wei, Q.; Liao, Z.; Gu, J. Dynamics of Adaptive Immune Cell and NK Cell Subsets in Patients With Ankylosing Spondylitis After IL-17A Inhibition by Secukinumab. Front. Pharmacol. 2021, 12, 738316. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.; Lv, Q.; Wei, Q.; Jiang, Y.; Qi, J.; Xiao, M.; Fang, L.; Xie, Y.; Cao, S.; Lin, Z.; et al. TNF-α inhibitor therapy can improve the immune imbalance of CD4+ T cells and negative regulatory cells but not CD8+ T cells in ankylosing spondylitis. Arthritis Res. Ther. 2020, 22, 149. [Google Scholar] [CrossRef] [PubMed]
- Kwon, O.C.; Park, J.H.; Park, M.C. Tumour necrosis factor inhibitor tapering in patients with ankylosing spondylitis at low disease activity: Factors associated with flare. Ther. Adv. Musculoskelet. Dis. 2021, 13, 1759720X20986732. [Google Scholar] [CrossRef] [PubMed]
- Tsukazaki, H.; Kaito, T. The Role of the IL-23/IL-17 Pathway in the Pathogenesis of Spondyloarthritis. Int. J. Mol. Sci. 2020, 21, 6401. [Google Scholar] [CrossRef] [PubMed]
- Deveci, H.; Turk, A.C.; Ozmen, Z.C.; Demir, A.K.; Say Coskun, S.U. Biological and genetic evaluation of IL-23/IL-17 pathway in ankylosing spondylitis patients. Cent. Eur. J. Immunol. 2019, 44, 433–439. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.; Liu, B.; Lin, C.; Gu, J. Imbalance of Peripheral Lymphocyte Subsets in Patients With Ankylosing Spondylitis: A Meta-Analysis. Front. Immunol. 2021, 12, 696973. [Google Scholar] [CrossRef]
- Mangare, C.; Tischer-Zimmermann, S.; Riese, S.B.; Dragon, A.C.; Prinz, I.; Blasczyk, R.; Maecker-Kolhoff, B.; Eiz-Vesper, B. Robust Identification of Suitable T-Cell Subsets for Personalized CMV-Specific T-Cell Immunotherapy Using CD45RA and CD62L Microbeads. Int. J. Mol. Sci. 2019, 20, 1415. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vanikova, S.; Koladiya, A.; Musil, J. OMIP-080: 29-Color flow cytometry panel for comprehensive evaluation of NK and T cells reconstitution after hematopoietic stem cells transplantation. Cytometry A 2022, 101, 21–26. [Google Scholar] [CrossRef]
- Jameson, S.C. The Naming of Memory T-Cell Subsets. Cold Spring. Harb. Perspect. Biol. 2021, 13, a037788. [Google Scholar] [CrossRef]
- Meraviglia, S.; Di Carlo, P.; Pampinella, D.; Guadagnino, G.; Presti, E.L.; Orlando, V.; Marchetti, G.; Dieli, F.; Sergi, C. T-Cell Subsets (TCM, TEM, TEMRA) and Poly-Functional Immune Response in Patients with Human Immunodeficiency Virus (HIV) Infection and Different T-CD4 Cell Response. Ann. Clin. Lab. Sci. 2019, 49, 519–528. [Google Scholar]
- Al Barashdi, M.A.; Ali, A.; McMullin, M.F.; Mills, K. Protein tyrosine phosphatase receptor type C (PTPRC or CD45). J. Clin. Pathol. 2021, 74, 548–552. [Google Scholar] [CrossRef] [PubMed]
- Courtney, A.H.; Shvets, A.A.; Lu, W.; Griffante, G.; Mollenauer, M.; Horkova, V.; Lo, W.L.; Yu, S.; Stepanek, O.; Chakraborty, A.K.; et al. CD45 functions as a signaling gatekeeper in T cells. Sci. Signal. 2019, 12, eaaw8151. [Google Scholar] [CrossRef] [PubMed]
- Su, Z.; Huang, D. Alternative Splicing of Pre-mRNA in the Control of Immune Activity. Genes 2021, 12, 574. [Google Scholar] [CrossRef] [PubMed]
- Sani, M.M.; Ashari, N.S.M.; Abdullah, B.; Wong, K.K.; Musa, K.I.; Mohamud, R.; Tan, H.T. Reduced CD4+ terminally differentiated effector memory T cells in moderate-severe house dust mites sensitized allergic rhinitis patients. Asian Pac. J. Allergy Immunol. 2019, 3, 138–146. [Google Scholar] [CrossRef] [PubMed]
- Golubovskaya, V.; Wu, L. Different Subsets of T Cells, Memory, Effector Functions, and CAR-T Immunotherapy. Cancers 2016, 8, 36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johansen, K.H.; Golec, D.P.; Thomsen, J.H.; Schwartzberg, P.L.; Okkenhaug, K. PI3K in T Cell Adhesion and Trafficking. Front. Immunol. 2021, 12, 708908. [Google Scholar] [CrossRef] [PubMed]
- Amarnani, A.A.; Poladian, K.R.; Marciano, B.E.; Daub, J.R.; Williams, S.G.; Livinski, A.A.; Hsu, A.P.; Palmer, C.L.; Kenney, C.M.; Avila, D.N.; et al. A Panoply of Rheumatological Manifestations in Patients with GATA2 Deficiency. Sci. Rep. 2020, 10, 8305. [Google Scholar] [CrossRef]
- Dennehy, K.M.; Löll, E.; Dhillon, C.; Classen, J.M.; Warm, T.D.; Schuierer, L.; Hyhlik-Dürr, A.; Römmele, C.; Gosslau, Y.; Kling, E.; et al. Comparison of the Development of SARS-Coronavirus-2-Specific Cellular Immunity, and Central Memory CD4+ T-Cell Responses Following Infection versus Vaccination. Vaccines 2021, 9, 1439. [Google Scholar] [CrossRef]
- Matyushenko, V.; Isakova-Sivak, I.; Kudryavtsev, I.; Goshina, A.; Chistyakova, A.; Stepanova, E.; Prokopenko, P.; Sychev, I.; Rudenko, L. Detection of IFNγ-Secreting CD4+ and CD8+ Memory T Cells in COVID-19 Convalescents after Stimulation of Peripheral Blood Mononuclear Cells with Live SARS-CoV-2. Viruses 2021, 13, 1490. [Google Scholar] [CrossRef] [PubMed]
- Kasatskaya, S.A.; Ladell, K.; Egorov, E.S.; Miners, K.L.; Davydov, A.N.; Metsger, M.; Staroverov, D.B.; Matveyshina, E.K.; Shagina, I.A.; Mamedov, I.Z.; et al. Functionally specialized human CD4+ T-cell subsets express physicochemically distinct TCRs. Elife 2020, 9, e57063. [Google Scholar] [CrossRef]
- Scheu, S.; Ali, S.; Ruland, C.; Arolt, V.; Alferink, J. The C-C Chemokines CCL17 and CCL22 and Their Receptor CCR4 in CNS Autoimmunity. Int. J. Mol. Sci. 2017, 18, 2306. [Google Scholar] [CrossRef] [PubMed]
- Yoshie, O. CCR4 as a Therapeutic Target for Cancer Immunotherapy. Cancers 2021, 13, 5542. [Google Scholar] [CrossRef] [PubMed]
- Lee, A.Y.; Körner, H. CC chemokine receptor 6 (CCR6) in the pathogenesis of systemic lupus erythematosus. Immunol. Cell Biol. 2020, 98, 845–853. [Google Scholar] [CrossRef] [PubMed]
- Meitei, H.T.; Jadhav, N.; Lal, G. CCR6-CCL20 axis as a therapeutic target for autoimmune diseases. Autoimmun. Rev. 2021, 20, 102846. [Google Scholar] [CrossRef]
- Karin, N. CXCR3 Ligands in Cancer and Autoimmunity, Chemoattraction of Effector T Cells, and Beyond. Front. Immunol. 2020, 11, 976. [Google Scholar] [CrossRef] [PubMed]
- Levesque, L.A.; Roy, S.; Salazar, N. CXCR3 Expression and Genome-Wide 3’ Splice Site Selection in the TCGA Breast Cancer Cohort. Life 2021, 11, 746. [Google Scholar] [CrossRef]
- Magnusen, A.F.; Rani, R.; McKay, M.A.; Hatton, S.L.; Nyamajenjere, T.C.; Magnusen, D.N.A.; Köhl, J.; Grabowski, G.A.; Pandey, M.K. C-X-C Motif Chemokine Ligand 9 and Its CXCR3 Receptor Are the Salt and Pepper for T Cells Trafficking in a Mouse Model of Gaucher Disease. Int. J. Mol. Sci. 2021, 22, 12712. [Google Scholar] [CrossRef]
- Hsieh, C.H.; Jian, C.Z.; Lin, L.I.; Low, G.S.; Ou, P.Y.; Hsu, C.; Ou, D.L. Potential Role of CXCL13/CXCR5 Signaling in Immune Checkpoint Inhibitor Treatment in Cancer. Cancers 2022, 14, 294. [Google Scholar] [CrossRef]
- Hussain, M.; Adah, D.; Tariq, M.; Lu, Y.; Zhang, J.; Liu, J. CXCL13/CXCR5 signaling axis in cancer. Life Sci. 2019, 227, 175–186. [Google Scholar] [CrossRef]
- Kazanietz, M.G.; Durando, M.; Cooke, M. CXCL13 and Its Receptor CXCR5 in Cancer: Inflammation, Immune Response, and Beyond. Front. Endocrinol. 2019, 10, 471. [Google Scholar] [CrossRef] [Green Version]
- Aldridge, J.; Ekwall, A.H.; Mark, L.; Bergström, B.; Andersson, K.; Gjertsson, I.; Lundell, A.C.; Rudin, A. T helper cells in synovial fluid of patients with rheumatoid arthritis primarily have a Th1 and a CXCR3+Th2 phenotype. Arthritis Res. Ther. 2020, 22, 245. [Google Scholar] [CrossRef] [PubMed]
- O’Neil, T.R.; Hu, K.; Truong, N.R.; Arshad, S.; Shacklett, B.L.; Cunningham, A.L.; Nasr, N. The Role of Tissue Resident Memory CD4 T Cells in Herpes Simplex Viral and HIV Infection. Viruses 2021, 13, 359. [Google Scholar] [CrossRef] [PubMed]
- Pandya, J.M.; Lundell, A.C.; Hallström, M.; Andersson, K.; Nordström, I.; Rudin, A. Circulating T helper and T regulatory subsets in untreated early rheumatoid arthritis and healthy control subjects. J. Leukoc. Biol. 2016, 100, 823–833. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wacleche, V.S.; Landay, A.; Routy, J.P.; Ancuta, P. The Th17 Lineage: From Barrier Surfaces Homeostasis to Autoimmunity, Cancer, and HIV-1 Pathogenesis. Viruses 2017, 9, 303. [Google Scholar] [CrossRef] [PubMed]
- Kuwabara, T.; Ishikawa, F.; Kondo, M.; Kakiuchi, T. The Role of IL-17 and Related Cytokines in Inflammatory Autoimmune Diseases. Mediators Inflamm. 2017, 2017, 3908061. [Google Scholar] [CrossRef] [Green Version]
- Schinocca, C.; Rizzo, C.; Fasano, S.; Grasso, G.; La Barbera, L.; Ciccia, F.; Guggino, G. Role of the IL-23/IL-17 Pathway in Rheumatic Diseases: An Overview. Front. Immunol. 2021, 12, 637829. [Google Scholar] [CrossRef]
- Chatzileontiadou, D.S.M.; Sloane, H.; Nguyen, A.T.; Gras, S.; Grant, E.J. The Many Faces of CD4+ T Cells: Immunological and Structural Characteristics. Int. J. Mol. Sci. 2020, 22, 73. [Google Scholar] [CrossRef]
- Sehgal, A.; Irvine, K.M.; Hume, D.A. Functions of macrophage colony-stimulating factor (CSF1) in development, homeostasis, and tissue repair. Semin Immunol. 2021, 54, 101509. [Google Scholar] [CrossRef]
- Sinha, S.K.; Miikeda, A.; Fouladian, Z.; Mehrabian, M.; Edillor, C.; Shih, D.; Zhou, Z.; Paul, M.K.; Charugundla, S.; Davis, R.C.; et al. Local M-CSF (Macrophage Colony-Stimulating Factor) Expression Regulates Macrophage Proliferation and Apoptosis in Atherosclerosis. Arterioscler. Thromb. Vasc. Biol. 2021, 41, 220–233. [Google Scholar] [CrossRef]
- Trus, E.; Basta, S.; Gee, K. Who’s in charge here? Macrophage colony stimulating factor and granulocyte macrophage colony stimulating factor: Competing factors in macrophage polarization. Cytokine 2020, 127, 154939. [Google Scholar] [CrossRef]
- Song, J.E.; Kim, J.S.; Shin, J.H.; Moon, K.W.; Park, J.K.; Park, K.S.; Lee, E.Y. Role of Synovial Exosomes in Osteoclast Differentiation in Inflammatory Arthritis. Cells 2021, 10, 120. [Google Scholar] [CrossRef] [PubMed]
- Dankers, W.; den Braanker, H.; Paulissen, S.M.J.; van Hamburg, J.P.; Davelaar, N.; Colin, E.M.; Lubberts, E. The heterogeneous human memory CCR6+ T helper-17 populations differ in T-bet and cytokine expression but all activate synovial fibroblasts in an IFNγ-independent manner. Arthritis Res. Ther. 2021, 23, 157. [Google Scholar] [CrossRef] [PubMed]
- Den Braanker, H.; Razawy, W.; Wervers, K.; Mus, A.C.; Davelaar, N.; Kok, M.R.; Lubberts, E. Characterizing memory T helper cells in patients with psoriasis, subclinical, or early psoriatic arthritis using a machine learning algorithm. Arthritis Res. Ther. 2022, 24, 28. [Google Scholar] [CrossRef] [PubMed]
- Kotake, S.; Yago, T.; Kobashigawa, T.; Nanke, Y. The Plasticity of Th17 Cells in the Pathogenesis of Rheumatoid Arthritis. J. Clin. Med. 2017, 6, 67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leipe, J.; Pirronello, F.; Klose, A.; Schulze-Koops, H.; Skapenko, A. Increased plasticity of non-classic Th1 cells toward the Th17 phenotype. Mod. Rheumatol. 2020, 30, 930–936. [Google Scholar] [CrossRef] [PubMed]
- Van Hamburg, J.P.; Tas, S.W. Molecular mechanisms underpinning T helper 17 cell heterogeneity and functions in rheumatoid arthritis. J. Autoimmun. 2018, 87, 69–81. [Google Scholar] [CrossRef]
- Cerboni, S.; Gehrmann, U.; Preite, S.; Mitra, S. Cytokine-regulated Th17 plasticity in human health and diseases. Immunology 2021, 163, 3–18. [Google Scholar] [CrossRef]
- Schinnerling, K.; Aguillón, J.C.; Catalán, D.; Soto, L. The role of interleukin-6 signalling and its therapeutic blockage in skewing the T cell balance in rheumatoid arthritis. Clin. Exp. Immunol. 2017, 189, 12–20. [Google Scholar] [CrossRef] [Green Version]
- Beurier, P.; Ricard, L.; Eshagh, D.; Malard, F.; Siblany, L.; Fain, O.; Mohty, M.; Gaugler, B.; Mekinian, A. TFH cells in systemic sclerosis. J. Transl Med. 2021, 19, 375. [Google Scholar] [CrossRef]
- Law, H.; Venturi, V.; Kelleher, A.; Munier, C.M.L. Tfh Cells in Health and Immunity: Potential Targets for Systems Biology Approaches to Vaccination. Int. J. Mol. Sci. 2020, 21, 8524. [Google Scholar] [CrossRef]
- Kurata, I.; Matsumoto, I.; Sumida, T. T follicular helper cell subsets: A potential key player in autoimmunity. Immunol. Med. 2021, 44, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Morita, R.; Schmitt, N.; Bentebibel, S.E.; Ranganathan, R.; Bourdery, L.; Zurawski, G.; Foucat, E.; Dullaers, M.; Oh, S.; Sabzghabaei, N.; et al. Human blood CXCR5(+)CD4(+) T cells are counterparts of T follicular cells and contain specific subsets that differentially support antibody secretion. Immunity 2011, 34, 108–121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cui, D.; Tang, Y.; Jiang, Q.; Jiang, D.; Zhang, Y.; Lv, Y.; Xu, D.; Wu, J.; Xie, J.; Wen, C.; et al. Follicular Helper T Cells in the Immunopathogenesis of SARS-CoV-2 Infection. Front. Immunol. 2021, 12, 731100. [Google Scholar] [CrossRef] [PubMed]
- Velu, V.; Mylvaganam, G.; Ibegbu, C.; Amara, R.R. Tfh1 Cells in Germinal Centers During Chronic HIV/SIV Infection. Front. Immunol. 2018, 9, 1272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Ji, H.; Zhao, P.; Yan, H.; Cai, Y.; Yu, L.; Hu, X.; Sun, X.; Jiang, Y. Follicular helper T cell and memory B cell immunity in CHC patients. J. Mol. Med. 2019, 97, 397–407. [Google Scholar] [CrossRef] [PubMed]
- Desbois, A.C.; Régnier, P.; Quiniou, V.; Lejoncour, A.; Maciejewski-Duval, A.; Comarmond, C.; Vallet, H.; Rosenzwag, M.; Darrasse-Jèze, G.; Derian, N.; et al. Specific Follicular Helper T Cell Signature in Takayasu Arteritis. Arthritis Rheumatol. 2021, 73, 1233–1243. [Google Scholar] [CrossRef]
- Weinstein, J.S.; Herman, E.I.; Lainez, B.; Licona-Limón, P.; Esplugues, E.; Flavell, R.; Craft, J. TFH cells progressively differentiate to regulate the germinal center response. Nat. Immunol. 2016, 17, 1197–1205. [Google Scholar] [CrossRef] [Green Version]
- Kudryavtsev, I.; Serebriakova, M.; Starshinova, A.; Zinchenko, Y.; Basantsova, N.; Malkova, A.; Soprun, L.; Churilov, L.P.; Toubi, E.; Yablonskiy, P.; et al. Imbalance in B cell and T Follicular Helper Cell Subsets in Pulmonary Sarcoidosis. Sci. Rep. 2020, 10, 1059. [Google Scholar] [CrossRef] [Green Version]
- Cooper, A.M.; Khader, S.A. IL-12p40: An inherently agonistic cytokine. Trends Immunol. 2007, 28, 33–38. [Google Scholar] [CrossRef]
- Li, Z.; Huang, Z.; Bai, L. Cell Interplay in Osteoarthritis. Front. Cell. Dev. Biol. 2021, 9, 720477. [Google Scholar] [CrossRef]
- Paiva, I.A.; Badolato-Corrêa, J.; Familiar-Macedo, D.; de-Oliveira-Pinto, L.M. Th17 Cells in Viral Infections-Friend or Foe? Cells 2021, 10, 1159. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Lu, Q.; Liu, Y.; Shi, Z.; Hu, L.; Zeng, Z.; Tu, Y.; Xiao, Z.; Xu, Q. Th17 Cells in Inflammatory Bowel Disease: Cytokines, Plasticity, and Therapies. J. Immunol. Res. 2021, 2021, 8816041. [Google Scholar] [CrossRef] [PubMed]
- Van der Linden, S.; Valkenburg, H.A.; Cats, A. Evaluation of diagnostic criteria for ankylosing spondylitis. A proposal for modification of the New York criteria. Arthritis Rheum. 1984, 27, 361–368. [Google Scholar] [CrossRef] [PubMed]
- Machado, P.M.; Landewé, R.; Heijde, D.V.; Assessment of SpondyloArthritis international Society (ASAS). Ankylosing Spondylitis Disease Activity Score (ASDAS): 2018 update of the nomenclature for disease activity states. Ann. Rheum. Dis. 2018, 77, 1539–1540. [Google Scholar] [CrossRef]
- Golovkin, A.; Kalinina, O.; Bezrukikh, V.; Aquino, A.; Zaikova, E.; Karonova, T.; Melnik, O.; Vasilieva, E.; Kudryavtsev, I. Imbalanced Immune Response of T-Cell and B-Cell Subsets in Patients with Moderate and Severe COVID-19. Viruses 2021, 13, 1966. [Google Scholar] [CrossRef] [PubMed]
- Vaidyanathan, P.; Appleton, E.; Tran, D.; Vahid, A.; Church, G.; Densmore, D. Algorithms for the selection of fluorescent reporters. Commun. Biol. 2021, 4, 118. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Parameters | Control | Low-Activity AS | High-Activity AS |
|---|---|---|---|
| Th cells, % | 47.8 (44.9–52.7) | 45.3 (40.1–49.4) | 48.6 (40.2–53.0) |
| Th cells, cells/μL | 775.7 (676.0–1053.2) | 901.3 (735.3–1188.7) | 1061.6 (726.4–1217.7) |
| ‘Naïve’ Th, % | 31.4 (25.7–40.1) | 31.9 (23.6–43.5) | 25.6 (18.7–33.3) р1 = 0.037 |
| CM Th, % | 41.5 (37.9–47.3) | 45.4 (37.8–51.5) | 54.0 (46.3–56.1) р1 < 0.001 р2 = 0.032 |
| EM Th, % | 22.9 (17.3–27.3) | 18.5 (15.2–23.2) р1 = 0.048 | 18.8 (14.6–23.0) |
| TEMRA Th, % | 0.59 (0.25–1.89) | 0.86 (0.37–1.79) | 0.78 (0.51–1.50) |
| Parameters | Control | Low-Activity AS | High-Activity AS |
|---|---|---|---|
| CM Th, cells/ μL | 332.5 (266.8–468.8) | 388.4 (331.5–496.0) р1 = 0.017 | 576.5 (337.3–687.5) р1 < 0.001 |
| Th1 cells, % | 12.96 (11.03–16.89) | 10.89 (9.22–13.78) | 11.36 (6.61–13.29) р1 = 0.035 |
| Th2 cells, % | 10.67 (9.29–12.98) | 13.83 (9.83–15.83) | 14.92 (10.99–18.08) р1 = 0.014 |
| Th17 Th, % | 35.78 (32.94–43.69) | 40.98 (35.85–47.36) р1 = 0.007 | 40.41 (36.42–43.87) р1 = 0.048 |
| follicular Th, % | 18.89 (15.68–22.36) | 18.89 (12.28–22.29) | 17.65 (16.54–20.29) |
| CM Th17 Cell Subsets | Control | Low-Activity AS | High-Activity AS |
|---|---|---|---|
| ‘classical’ Th17 | 12.33 (10.35–15.69) | 16.12 (13.39–17.58) р1 = 0.004 | 16.99 (14.62–18.55) р1 < 0.001 |
| DN Th17 | 2.61 (1.72–3.16) | 4.38 (3.46–5.37) р1 < 0.001 | 4.45 (3.56–6.07) р1 < 0.001 |
| DP Th17 | 10.89 (8.58–12.65) | 8.05 (7.20–10.31) р1 = 0.001 | 7.27 (5.70–10.39) р1 < 0.001 |
| Th17.1 | 10.37 (8.16–12.74) | 11.76 (8.63–15.54) | 9.97 (6.52–13.13) |
| CM Tfh Cell Subsets | Control | Low-Activity AS | High-Activity AS |
|---|---|---|---|
| Tfh1 | 34.08 (29.87–36.40) | 24.77 (21.82–28.69) р1 < 0.001 | 24.29 (21.71–30.06) р1 < 0.001 |
| Tfh2 | 20.04 (18.20–24.45) | 22.63 (19.53–28.17) р1 = 0.021 | 27.62 (20.92–33.03) р1 = 0.001 |
| Tfh17 | 28.85 (26.71–35.16) | 36.19 (30.68–40.48) р1 < 0.001 | 36.47 (31.56–39.46) р1 = 0.013 |
| DP Tfh | 13.72 (11.40–18.52) | 14.09 (11.55–15.68) | 11.34 (9.63–14.11) р1 = 0.015 р2 = 0.049 |
| Parameters | Control | Low-Activity AS | High-Activity AS |
|---|---|---|---|
| EM Th, cells/μL | 178.0 (139.5–230.7) | 159.7 (123.4–264.3) | 205.1 (111.0–252.4) |
| Th1 cells, % | 21.24 (16.11–29.20) | 15.29 (13.12–26.35) | 15.00 (11.98–25.13) |
| Th2 cells, % | 1.35 (0.88–1.78) | 1.64 (1.06–2.38) | 2.02 (1.68–3.42) р1 < 0.001 р2 = 0.041 |
| Th17 Th, % | 52.32 (41.39–59.88) | 58.84 (53.06–66.20) р1 = 0.040 | 58.48 (51.36–63.37) |
| follicular Th, % | 8.10 (5.81–12.43) | 7.68 (3.80–10.05) | 9.21 (7.07–11.67) |
| EM Th17 Cell Subset | Control | Low-Activity AS | High-Activity AS |
|---|---|---|---|
| ‘classical’ Th17 | 13.01 (9.30–15.35) | 15.63 (11.13–19.70) р1 = 0.035 | 19.16 (15.07–22.52) р1 < 0.001 |
| DN Th17 | 2.33 (1.28–3.02) | 3.52 (2.44–5.11) р1 = 0.001 | 3.97 (2.24–4.60) р1 = 0.009 |
| DP Th17 | 11.44 (8.07–15.27) | 11.31 (7.90–13.88) | 10.48 (6.69–14.20) |
| Th17.1 | 24.02 (18.80–29.67) | 25.00 (18.18–31.99) | 22.11 (16.41–25.40) |
| EM Tfh Cell Subsets | Control | Low-Activity AS | High-Activity AS |
|---|---|---|---|
| Tfh1 | 30.54 (23.90–34.47) | 21.38 (17.11–27.08) р1 < 0.001 | 25.89 (18.91–30.03) р1 = 0.016 |
| Tfh2 | 14.90 (10.34–18.45) | 15.38 (11.96–20.60) | 18.96 (13.64–26.33) р1 = 0.031 |
| Tfh17 | 38.47 (32.33–43.07) | 46.48 (40.91–48.40) р1 < 0.001 | 44.16 (35.08–46.68) |
| DP Tfh | 16.20 (12.08–21.25) | 16.14 (12.84–18.72) | 13.12 (10.07–14.55) р1 = 0.019 р2 = 0.037 |
| Parameters | Control | Low-activity AS | High-activity AS |
|---|---|---|---|
| IL-1β | 7.64 (5.01–17.93) | 30.09 (17.97–59.97) р1 < 0.001 | 28.56 (13.51–39.94) р1 < 0.001 |
| IL-2 | 0.44 (0.10–1.05) | 1.05 (0.78–1.46) р1 = 0.016 | 0.98 (0.57–1.88) |
| IL-4 | 0.83 (0.24–1.92) | 1.32 (0.72–2.65) | 1.62 (0.87–3.80) |
| IL-5 | 3.80 (1.77–4.68) | 4.24 (2.93–6.01) | 6.38 (4.00–10.01) р1 = 0.017 |
| IL-17A | 2.22 (1.04–5.48) | 6.17 (5.13–8.19) р1 = 0.003 | 5.48 (3.35–6.85) р1 = 0.048 |
| IL-17E/IL-25 | 177 (99–308) | 290 (207–450) р1 = 0.041 | 253 (216–344) |
| IL-17F | 11.58 (11.58–18.54) | 23.63 (17.53–45.82) р1 < 0.001 | 21.58 (15.53–46.90) р1 = 0.002 |
| IFN-γ | 10.64 (3.63–15.46) | 16.75 (10.67–28.09) р1 = 0.026 | 18.73 (12.47–28.49) р1 = 0.021 |
| TNF-α | 19.12 (14.30–29.27) | 29.27 (23.31–41.10) | 22.41 (16.11–36.97) |
| Parameters | Low-Activity AS | High-Activity AS | р |
|---|---|---|---|
| Age (years), Ме (С25–С75) | 37 (30–45) | 41 (39–48) | 0.117 |
| Gender, n (%) Females/Males | 8 (21.6)/29 (78.4) | 6 (28.6)/15 (71.4) | 0.750 |
| HLA-B27+, n (%) | 29 (78.4) | 19 (90.5) | 0.301 |
| Extra-skeletal signs, n (%) | 13 (36.4) | 7 (33.3) | 1.000 |
| BASDAI, Ме (IQR) | 1.0 (1.0–2.0) | 3.9 (2.6–4.9) | <0.001 |
| ASDAS-CRP, Ме (IQR) | 1.3 (1.1–1.6) | 2.6 (2.1–3.6) | <0.001 |
| BASFI, Ме (IQR) | 1.0 (0.5–2.0) | 3.9 (3.7–4.6) | <0.001 |
| BASFI > 4, n (%) | 0 (0) | 10 (47.6) | <0.001 |
| Leukocytes, 109/L Ме (IQR) | 6.94 (6.16–8.28) | 8.08 (6.77–10.68) | 0.094 |
| Platelets, 109/L Ме (IQR) | 275.0 (23.7.0–318.0) | 296.0 (248.0–332.8) | 0.441 |
| RBCs, 1012/L Ме (IQR) | 4.88 (4.58–5.13) | 4.76 (4.29–5.04) | 0.454 |
| Hemoglobin, g/L Ме (IQR) | 14.60 (13.90–15.50) | 13.70 (13.00–14.50) | 0.052 |
| CRP, mg/L Ме (IQR) | 2.10 (0.79–3.08) | 8.37 (3.71–21.82) | <0.001 |
| ESR, mm/h Ме (IQR) | 5.0 (4.1–6.9) | 10.1 (5.2–21.9) | 0.020 |
| Type of Therapy | Low-Activity AS | High-Activity AS | р |
|---|---|---|---|
| No bDMARDs | 13 (35.2) | 6 (28.6) | 0.0502 |
| Anti-IL17 | 9 (24.3) | 6 (28.6) | 0.4661 |
| Anti-TNFα | 15 (40.5) | 9 (42.8) | 0.1482 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shesternya, P.A.; Savchenko, A.A.; Gritsenko, O.D.; Vasileva, A.O.; Kudryavtsev, I.V.; Masterova, A.A.; Isakov, D.V.; Borisov, A.G. Features of Peripheral Blood Th-Cell Subset Composition and Serum Cytokine Level in Patients with Activity-Driven Ankylosing Spondylitis. Pharmaceuticals 2022, 15, 1370. https://doi.org/10.3390/ph15111370
Shesternya PA, Savchenko AA, Gritsenko OD, Vasileva AO, Kudryavtsev IV, Masterova AA, Isakov DV, Borisov AG. Features of Peripheral Blood Th-Cell Subset Composition and Serum Cytokine Level in Patients with Activity-Driven Ankylosing Spondylitis. Pharmaceuticals. 2022; 15(11):1370. https://doi.org/10.3390/ph15111370
Chicago/Turabian StyleShesternya, Pavel A., Andrei A. Savchenko, Olga D. Gritsenko, Alexandra O. Vasileva, Igor V. Kudryavtsev, Alena A. Masterova, Dmitry V. Isakov, and Alexandr G. Borisov. 2022. "Features of Peripheral Blood Th-Cell Subset Composition and Serum Cytokine Level in Patients with Activity-Driven Ankylosing Spondylitis" Pharmaceuticals 15, no. 11: 1370. https://doi.org/10.3390/ph15111370