
Characterisation of Antiviral Activity of Cathelicidins from Naked Mole Rat and Python bivittatus on Human Herpes Simplex Virus 1

 and
and
Abstract
:1. Introduction
2. Results
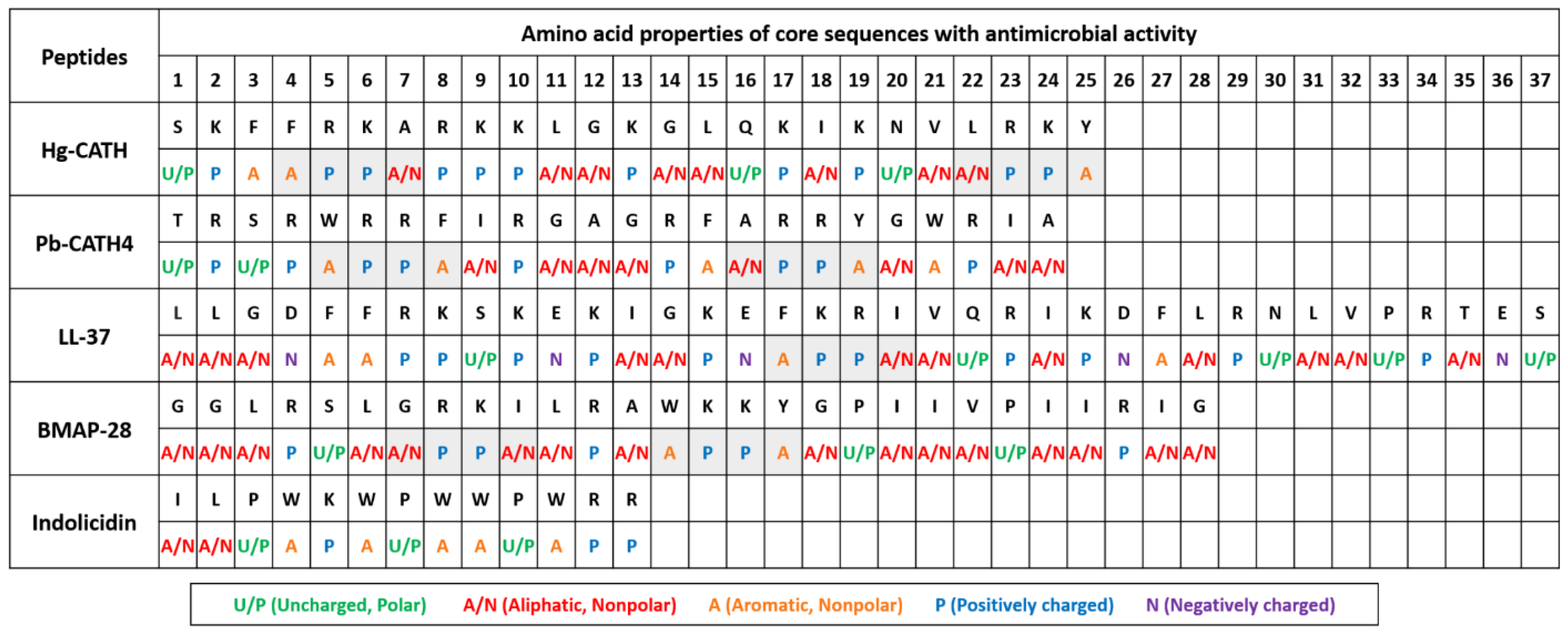
2.1. Comparison of Amino Acid Sequences and Peptide Structures of Hg-CATH and Pb-CATH4
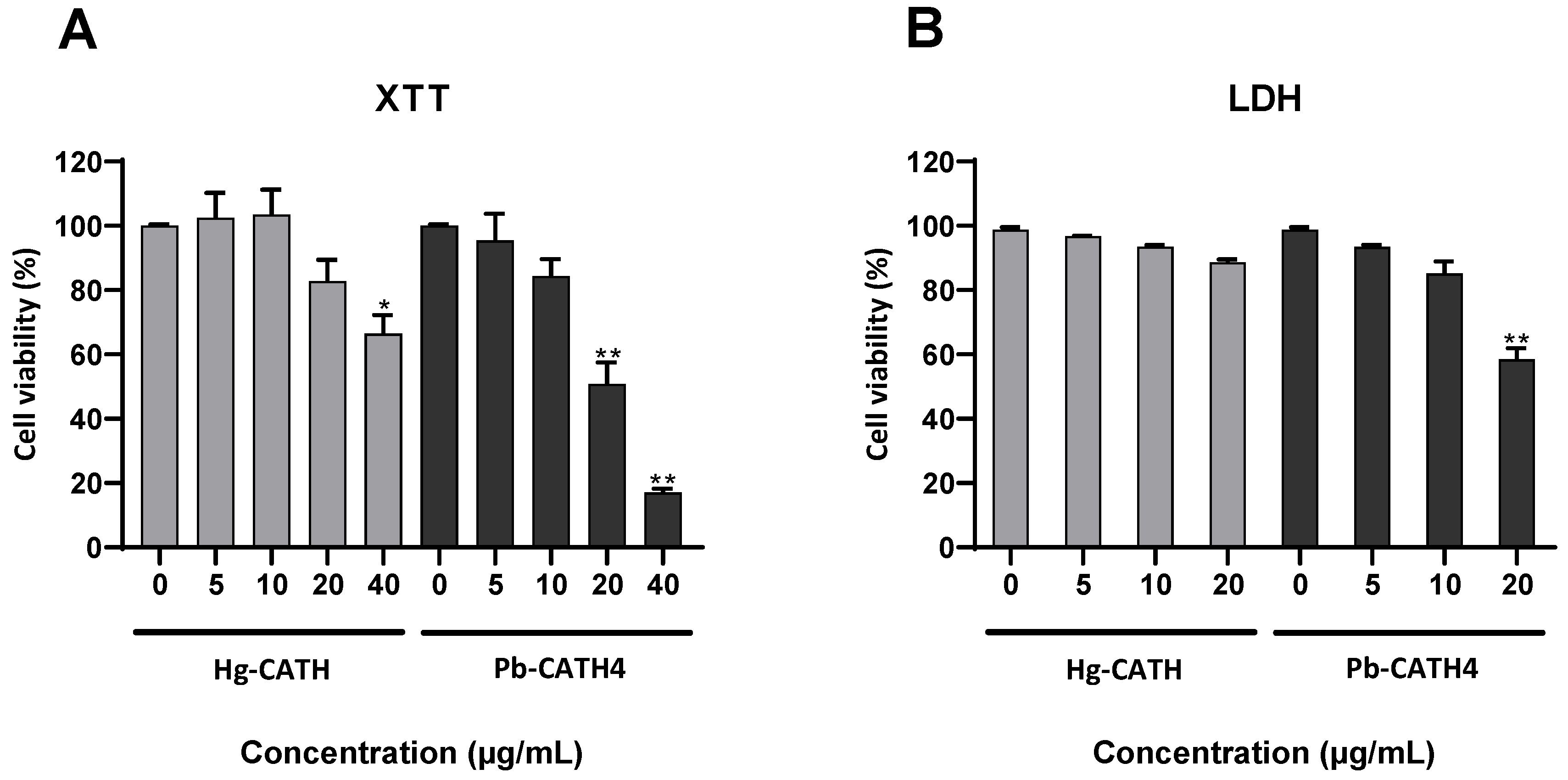
2.2. Cytotoxicity of Hg-CATH and Pb-CATH4 on Primary Human Keratinocytes
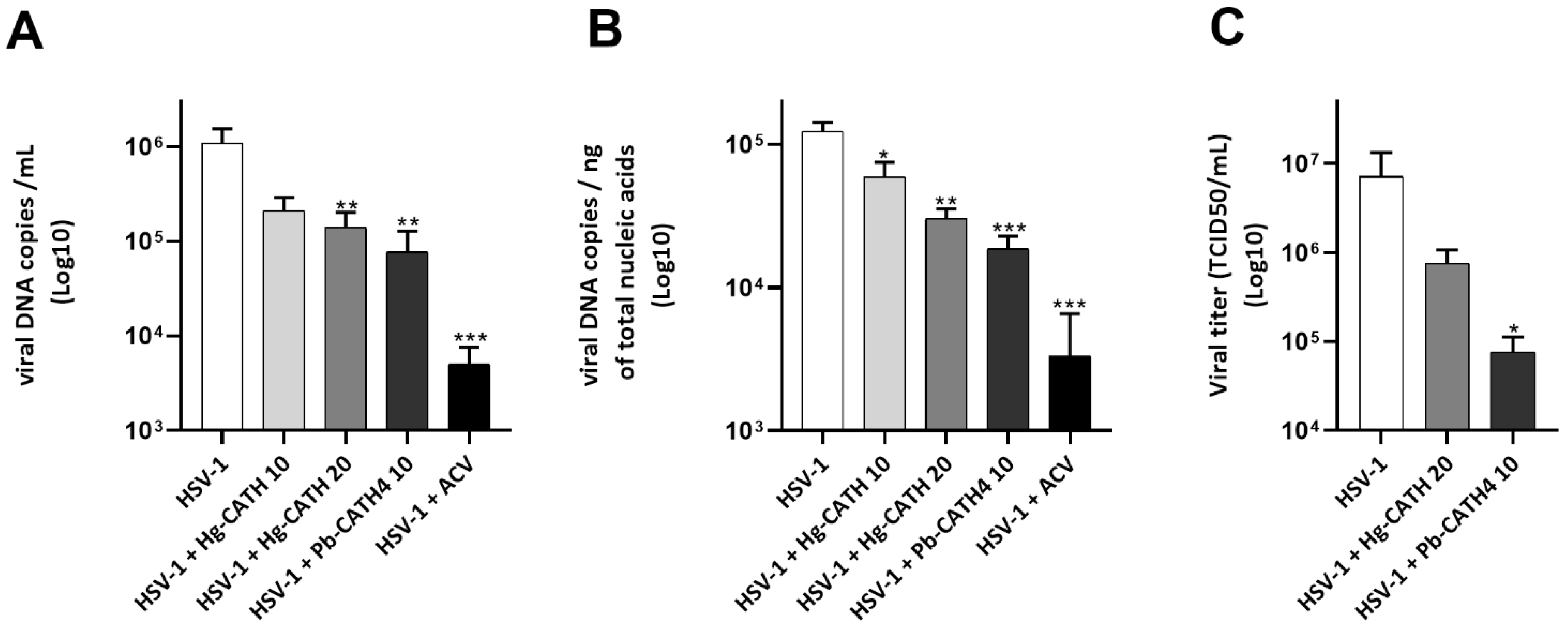
2.3. Hg-CATH and Pb-CATH4 Reduce HSV-1 Replication in Primary Human Keratinocytes
2.4. Hg-CATH and Pb-CATH4 Have No Virucidal Activity against HSV-1
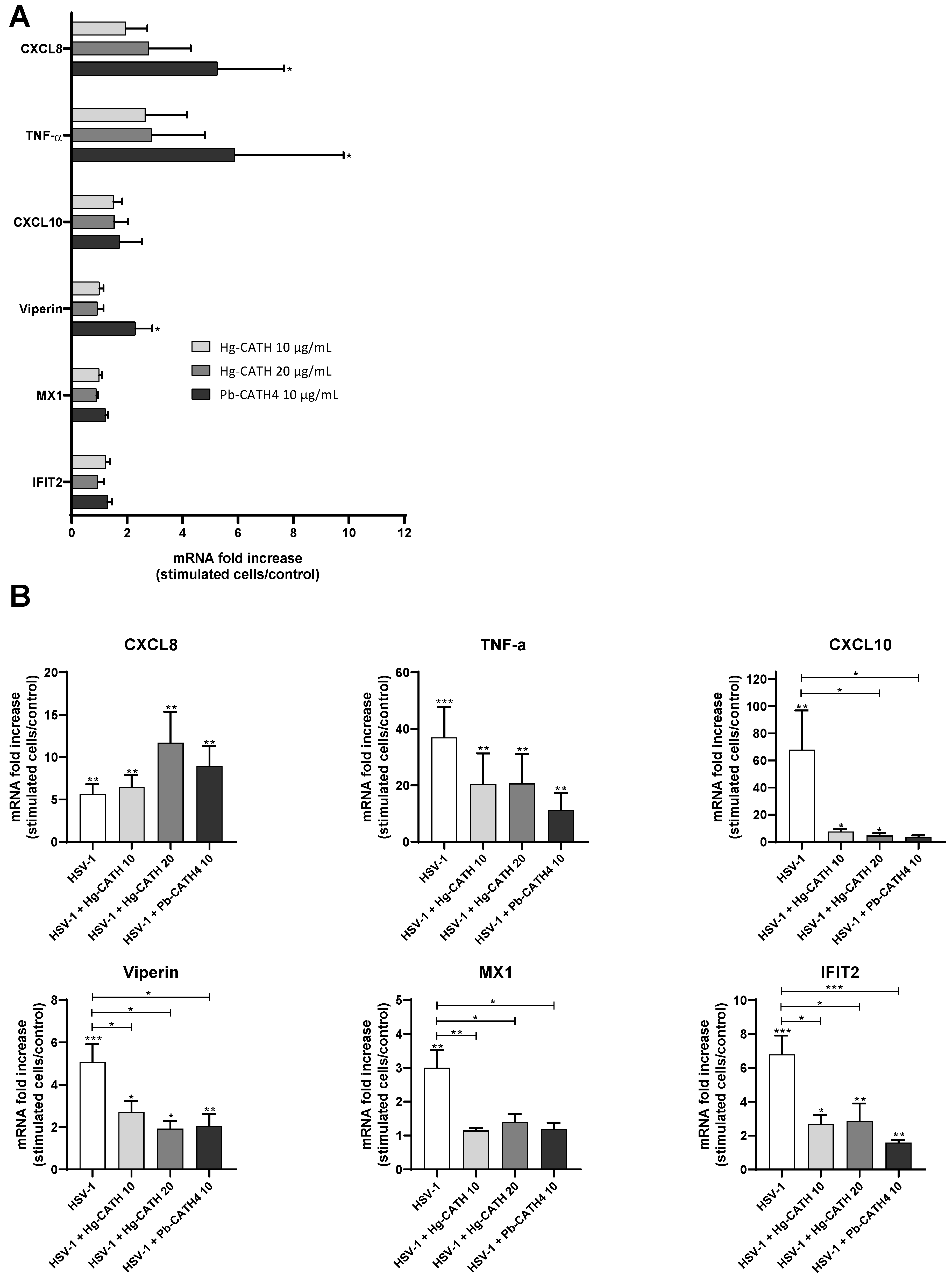
2.5. Modulation of the Innate Immune Response of Keratinocytes by Hg-CATH and Pb-CATH4
3. Discussion
4. Materials and Methods
4.1. Peptides
4.2. Virus Strain and Production
4.3. Isolation and Culture of Normal Human Epidermal Keratinocytes from Skin Samples
4.4. Cell Viability Assay
4.5. Evaluation of the Antiviral Effects of the AMPs
4.6. Nucleic Acid Extraction
4.7. Viral DNA Quantification by Real-Time Polymerase Chain Reaction (qPCR)
4.8. Transcriptomic Analysis of the Inflammatory and Antiviral Response in Keratinocytes
4.9. Viral Titration
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Aurelian, L.; Wagner, E.; Fiume, G.C. Herpes simplex viruses. In Encyclopedia of Virology; Elsevier: Amsterdam, The Netherlands, 1994; pp. 587–626. [Google Scholar]
- Faccin-Galhardi, L.C.; Linhares, R.E.C.; Nozawa, C. Herpesvirus: An underestimated virus. Folia Microbiol. 2017, 62, 151–156. [Google Scholar] [CrossRef]
- Burrel, S.; Boutolleau, D. Virus Herpes Simplex. In Traité de Virologie Médicale; Société Française de Microbiologie (SFM): Paris, France, 2019; pp. 209–224. [Google Scholar]
- Cunningham, A.L.; Diefenbach, R.; Miranda-Saksena, M.; Bosnjak, L.; Kim, M.; Jones, C.; Douglas, M.W. The cycle of human herpes simplex virus infection: Virus transport and immune control. J. Infect. Dis. 2006, 194 (Suppl. 1), S11–S18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parra-Sanchez, M. Genital ulcers caused by herpes simplex virus. Enferm. Infecc. Microbiol. Clin. 2019, 37, 260–264. [Google Scholar] [CrossRef] [PubMed]
- Samies, N.L.; James, S.H. Prevention and treatment of neonatal herpes simplex virus infection. Antiviral Res. 2020, 176, 104721. [Google Scholar] [CrossRef] [PubMed]
- Spear, P.G.; Longnecker, R. Herpesvirus entry: An update. J. Virol. 2003, 77, 10179–10185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Whitley, R.; Baines, J. Clinical management of herpes simplex virus infections: Past, present, and future. F1000Res. 2018, 7, 1726. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Damour, A.; Garcia, M.; Seneschal, J.; Lévêque, N.; Bodet, C. Eczema Herpeticum: Clinical and Pathophysiological Aspects. Clin. Rev. Allergy Immunol. 2019, 59, 1–18. [Google Scholar] [CrossRef]
- Izzedine, H.; Launay-Vacher, V.; Deray, G. Antiviral drug-induced nephrotoxicity. Am. J. Kidney Dis. 2005, 45, 804–817. [Google Scholar] [CrossRef]
- Piret, J.; Boivin, G. Resistance of herpes simplex viruses to nucleoside analogues: Mechanisms, prevalence, and management. Antimicrob. Agents Chemother. 2011, 55, 459–472. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piret, J.; Boivin, G. Antiviral drug resistance in herpesviruses other than cytomegalovirus. Rev. Med. Virol. 2014, 24, 186–218. [Google Scholar] [CrossRef]
- Haney, E.F.; Mansour, S.C.; Hancock, R.E. Antimicrobial Peptides: An Introduction. Methods Mol. Biol. 2017, 1548, 3–22. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Li, X.; Wang, Z. APD3: The antimicrobial peptide database as a tool for research and education. Nucleic Acids Res. 2016, 44, D1087–D1093. [Google Scholar] [CrossRef] [Green Version]
- Wilson, S.S.; Wiens, M.E.; Smith, J.G. Antiviral mechanisms of human defensins. J. Mol. Biol. 2013, 425, 4965–4980. [Google Scholar] [CrossRef]
- Kościuczuk, E.M.; Lisowski, P.; Jarczak, J.; Strzałkowska, N.; Jóźwik, A.; Horbańczuk, J.O.; Krzyżewski, J.; Zwierzchowski, L.; Bagnicka, E. Cathelicidins: Family of antimicrobial peptides. A review. Mol. Biol. Rep. 2012, 39, 10957–10970. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chessa, C.; Bodet, C.; Jousselin, C.; Wehbe, M.; Lévêque, N.; Garcia, M. Antiviral and Immunomodulatory Properties of Antimicrobial Peptides Produced by Human Keratinocytes. Front. Microbiol. 2020, 11, 1155. [Google Scholar] [CrossRef] [PubMed]
- Lehrer, R.I.; Ganz, T. Antimicrobial peptides in mammalian and insect host defence. Curr. Opin. Immunol. 1999, 11, 23–27. [Google Scholar] [CrossRef]
- Roy, M.; Lebeau, L.; Chessa, C.; Damour, A.; Ladram, A.; Oury, B.; Boutolleau, D.; Bodet, C.; Lévêque, N. Comparison of Anti-Viral Activity of Frog Skin Anti-Microbial Peptides Temporin-Sha and [K(3)]SHa to LL-37 and Temporin-Tb against Herpes Simplex Virus Type 1. Viruses 2019, 11, 77. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, S.; Mele, J.; Wu, Y.; Buffenstein, R.; Hornsby, P.J. Resistance to experimental tumorigenesis in cells of a long-lived mammal, the naked mole-rat (Heterocephalus glaber). Aging Cell 2010, 9, 626–635. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cho, H.-S.; Soundrarajan, N.; Le Van Chanh, Q.; Jeon, H.; Cha, S.-Y.; Kang, M.; Ahn, B.; Hong, K.; Song, H.; Kim, J.-H.; et al. The novel cathelicidin of naked mole rats, Hg-CATH, showed potent antimicrobial activity and low cytotoxicity. Gene 2018, 676, 164–170. [Google Scholar] [CrossRef]
- Kim, D.; Soundrarajan, N.; Lee, J.; Cho, H.-S.; Choi, M.; Cha, S.-Y.; Ahn, B.; Jeon, H.; Le, M.T.; Song, H.; et al. Genomewide Analysis of the Antimicrobial Peptides in Python bivittatus and Characterization of Cathelicidins with Potent Antimicrobial Activity and Low Cytotoxicity. Antimicrob. Agents Chemother. 2017, 61, e00530-17. [Google Scholar] [CrossRef] [Green Version]
- Combet, C.; Blanchet, C.; Geourjon, C.; Deléage, G. NPS@: Network protein sequence analysis. Trends Biochem. Sci. 2000, 25, 147–150. [Google Scholar] [CrossRef]
- Benincasa, M.; Skerlavaj, B.; Gennaro, R.; Pellegrini, A.; Zanetti, M. In vitro and in vivo antimicrobial activity of two alpha-helical cathelicidin peptides and of their synthetic analogs. Peptides 2003, 24, 1723–1731. [Google Scholar] [CrossRef]
- Yasin, B.; Pang, M.; Turner, J.S.; Cho, Y.; Dinh, N.; Waring, A.J.; Lehrer, R.I.; Wagar, E.A. Evaluation of the inactivation of infectious Herpes simplex virus by host-defense peptides. Eur. J. Clin. Microbiol. Infect. Dis. 2000, 19, 187–194. [Google Scholar] [CrossRef] [PubMed]
- Peng, L.; Du, W.; Balhuizen, M.; Haagsman, H.P.; de Haan, C.; Veldhuizen, E.J.A. Antiviral Activity of Chicken Cathelicidin B1 Against Influenza A Virus. Front. Microbiol. 2020, 11, 426. [Google Scholar] [CrossRef] [Green Version]
- Cho, H.-S.; Yum, J.; Larivière, A.; Lévêque, N.; Le, Q.V.C.; Ahn, B.; Jeon, H.; Hong, K.; Soundrarajan, N.; Kim, J.-H.; et al. Opossum Cathelicidins Exhibit Antimicrobial Activity Against a Broad Spectrum of Pathogens Including West Nile Virus. Front. Immunol. 2020, 11, 347. [Google Scholar] [CrossRef] [Green Version]
- He, M.; Zhang, H.; Li, Y.; Wang, G.; Tang, B.; Zhao, J.; Huang, Y.; Zheng, J. Cathelicidin-Derived Antimicrobial Peptides Inhibit Zika Virus Through Direct Inactivation and Interferon Pathway. Front. Immunol. 2018, 9, 722. [Google Scholar] [CrossRef] [Green Version]
- Xu, J.; Chen, S.; Jin, J.; Ma, L.; Guo, M.; Zhou, C.; Dou, J. Inhibition of peptide BF-30 on influenza A virus infection in vitro/vivo by causing virion membrane fusion. Peptides 2019, 112, 14–22. [Google Scholar] [CrossRef] [PubMed]
- Xing, M.; Ji, M.; Hu, J.; Zhu, T.; Chen, Y.; Bai, X.; Mwangi, J.; Mo, G.; Lai, R.; Jin, L. Snake Cathelicidin Derived Peptide Inhibits Zika Virus Infection. Front. Microbiol. 2020, 11, 1871. [Google Scholar] [CrossRef]
- Petermann, P.; Thier, K.; Rahn, E.; Rixon, F.J.; Bloch, W.; Özcelik, S.; Krummenacher, C.; Barron, M.; Dixon, M.J.; Scheu, S.; et al. Entry mechanisms of herpes simplex virus 1 into murine epidermis: Involvement of nectin-1 and herpesvirus entry mediator as cellular receptors. J. Virol. 2015, 89, 262–274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hazrati, E.; Galen, B.; Lu, W.; Wang, W.; Ouyang, Y.; Keller, M.J.; Lehrer, R.I.; Herold, B.C. Human alpha- and beta-defensins block multiple steps in herpes simplex virus infection. J. Immunol. 2006, 177, 8658–8666. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, C.-J.; Buznyk, O.; Kuffova, L.; Rajendran, V.; Forrester, J.V.; Phopase, J.; Islam, M.M.; Skog, M.; Ahlqvist, J.; Griffith, M. Cathelicidin LL-37 and HSV-1 Corneal Infection: Peptide Versus Gene Therapy. Transl. Vis. Sci. Technol. 2014, 3, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Currie, S.M.; Findlay, E.G.; McHugh, B.; Mackellar, A.; Man, T.; Macmillan, D.; Wang, H.; Fitch, P.; Schwarze, J.; Davidson, D.J. The human cathelicidin LL-37 has antiviral activity against respiratory syncytial virus. PLoS ONE 2013, 8, e73659. [Google Scholar] [CrossRef] [PubMed]
- Wong, J.H.; Łęgowska, A.; Rolka, K.; Ng, T.B.; Hui, M.; Cho, C.H.; Lam, W.W.L.; Au, S.W.N.; Gu, O.W.; Wan, D.C.C. Effects of cathelicidin and its fragments on three key enzymes of HIV-1. Peptides 2011, 32, 1117–1122. [Google Scholar] [CrossRef]
- Zheng, W.; Zhang, C.; Wuyun, Q.; Pearce, R.; Li, Y.; Zhang, Y. LOMETS2: Improved meta-threading server for fold-recognition and structure-based function annotation for distant-homology proteins. Nucleic Acids Res. 2019, 47, W429–W436. [Google Scholar] [CrossRef] [Green Version]
- Berendsen, H.; Van Der Spoel, D.; Van Drunen, R. GROMACS: A message-passing parallel molecular dynamics implementation. Comput. Phys. Commun. 1995, 91, 43–56. [Google Scholar] [CrossRef]
- DeLano, W.L. Pymol: An open-source molecular graphics tool. CCP4 Newsl. Protein Crystallogr. 2002, 40, 82–92. [Google Scholar]
- Garcia, M.; Alout, H.; Diop, F.; Damour, A.; Bengue, M.; Weill, M.; Missé, D.; Lévêque, N.; Bodet, C. Innate Immune Response of Primary Human Keratinocytes to West Nile Virus Infection and Its Modulation by Mosquito Saliva. Front. Cell. Infect. Microbiol. 2018, 8, 387. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward (5′ → 3′) | Reverse (5′ → 3′) |
|---|---|---|
| CXCL8 | TTGCCAAGGAGTGCTAAAGAA | AACCCTCTGCACCCAGTTTT |
| CXCL10 | AAGGATGGACCACACAGAGG | TGGAAGATGGGAAAGGTGAG |
| G3PDH | GGCTCTCCAGAACATCATCCCTGC | GGGTGTCGCTGTTGAAGTCAGAGG |
| IFIT2 | GCGTGAGAAGGTGAAGAGG | AATTTGGCAATGCAGGTAGG |
| MX1 | ACCACAGAGGCTCTCAGCAT | ACCACAGAGGCTCTCAGCAT |
| Viperin | GGCAAGTTGGTGAGGTTCTG | ACCACCTCCTCAGCTTTTGA |
| RPS28 | CCGTGTGCAGCCTATCAAG | CAAGCTCAGCGCAACCTC |
| TNF-α | TCACCCACACCATCAGCCGCATCG | GGGAAGGTTGGATGTTCGTCCTCC |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Damour, A.; Garcia, M.; Cho, H.-S.; Larivière, A.; Lévêque, N.; Park, C.; Bodet, C. Characterisation of Antiviral Activity of Cathelicidins from Naked Mole Rat and Python bivittatus on Human Herpes Simplex Virus 1. Pharmaceuticals 2021, 14, 715. https://doi.org/10.3390/ph14080715
Damour A, Garcia M, Cho H-S, Larivière A, Lévêque N, Park C, Bodet C. Characterisation of Antiviral Activity of Cathelicidins from Naked Mole Rat and Python bivittatus on Human Herpes Simplex Virus 1. Pharmaceuticals. 2021; 14(8):715. https://doi.org/10.3390/ph14080715
Chicago/Turabian StyleDamour, Alexia, Magali Garcia, Hye-Sun Cho, Andy Larivière, Nicolas Lévêque, Chankyu Park, and Charles Bodet. 2021. "Characterisation of Antiviral Activity of Cathelicidins from Naked Mole Rat and Python bivittatus on Human Herpes Simplex Virus 1" Pharmaceuticals 14, no. 8: 715. https://doi.org/10.3390/ph14080715