
Aberrant Expression of microRNA Clusters in Head and Neck Cancer Development and Progression: Current and Future Translational Impacts



1
Genomics Research Center, Academia Sinica, Taipei 115, Taiwan
2
School of Oral Hygiene, College of Oral Medicine, Taipei Medical University, Taipei 110, Taiwan
3
Department of Biochemistry, College of Medicine, Kaohsiung Medical University, Kaohsiung 807, Taiwan
*
Author to whom correspondence should be addressed.
†
These authors contributed equally to this work.
Pharmaceuticals 2021, 14(3), 194; https://doi.org/10.3390/ph14030194
Submission received: 7 January 2021
/
Revised: 14 February 2021
/
Accepted: 23 February 2021
/
Published: 27 February 2021
(This article belongs to the Special Issue MiRNA-Based Therapeutics in Cancer)
Abstract
:MicroRNAs are small non-coding RNAs known to negative regulate endogenous genes. Some microRNAs have high sequence conservation and localize as clusters in the genome. Their coordination is regulated by simple genetic and epigenetic events mechanism. In cells, single microRNAs can regulate multiple genes and microRNA clusters contain multiple microRNAs. MicroRNAs can be differentially expressed and act as oncogenic or tumor suppressor microRNAs, which are based on the roles of microRNA-regulated genes. It is vital to understand their effects, regulation, and various biological functions under both normal and disease conditions. Head and neck squamous cell carcinomas are some of the leading causes of cancer-related deaths worldwide and are regulated by many factors, including the dysregulation of microRNAs and their clusters. In disease stages, microRNA clusters can potentially control every field of oncogenic function, including growth, proliferation, apoptosis, migration, and intercellular commutation. Furthermore, microRNA clusters are regulated by genetic mutations or translocations, transcription factors, and epigenetic modifications. Additionally, microRNA clusters harbor the potential to act therapeutically against cancer in the future. Here, we review recent advances in microRNA cluster research, especially relative to head and neck cancers, and discuss their regulation and biological functions under pathological conditions as well as translational applications.
1. microRNA Biogenesis
Small non-coding RNAs (sncRNAs) are single strand RNA usually < 200 nucleotides in length and include but are not limited to microRNAs, small nucleolar RNAs (snoRNAs), small nuclear RNAs (snRNAs), Piwi-interacting RNAs (piRNAs), endogenous short interfering RNAs (siRNAs), small rDNA-derived RNAs (srRNAs), ribosomal RNA derived fragments (rRFs), transfer RNAs (tRNAs), and their derived small RNAs (tsRNAs) by their origin, length, and functions [1,2,3,4,5,6]. Most sncRNAs are transcripted by RNA polymerase II. Both snoRNAs and snRNAs are larger sncRNAs which length are between 60–140 nucleotides and about 150 nucleotides, respectively. SnoRNAs contribute to RNA modification and snRNA are components in spliceosome. TsRNAs and srRNAs are by-product from tRNA or rRNA and have diverse functions in cell biology. PiRNAs and microRNAs have similar manner to protein coding genes but have different size and interaction proteins. Usually, piRNAs are 26–31 nucleotides in length which interact with piwi-subfamily Argonaute protein and serve as RNA-mediated adaptive immunity against transposon expansions and invasions [7]. MicroRNAs are created by Drosha and Dicer endoribonuclease as 21–24 nucleotides length, that can act as endogenous negative gene regulators in the cells via translational inhibition or mRNA degradation [8]. MicroRNAs have both canonical (Drosha-Dicer and DGCR8 dependent) and noncanonical (Drosha-DGCR8 and Dicer independent but TUTase dependent) pathways to produce mature-form, single-strand microRNAs [9]. The mature-form microRNA enters an RNA-induced silencing complex (RISC) and act as a guidance RNA to regulate the expression of target genes by seed region sequence. RISC-microRNA complex can use microRNA seed region to target microRNA binding elements (MREs) target gene’s 3′-untranslation region (3′-UTR). MicroRNAs act by fine-tuning gene expression through a post-transcriptional mechanism. Due to the multiple-target regulation functions of microRNAs, microRNAs have critical roles in diverse biological functions such as embryogenesis, development, and physiological processes [10] (Figure 1).
2. microRNA Clusters
Evolutionary conservation is not only observed in microRNA–target interactions in diverse species [11] but also in genetic order, forming specific microRNA clusters during genomic organization [12]. Approximately 25% of human and 50% of Drosophila melanogaster microRNAs are clustered on chromosomes. MicroRNA clusters contain at least two or more microRNA genes, which are chromosomal adjacent sequences in the same orientation that are not separated by transcriptional units and may be encoded in opposite directions. Usually, cluster microRNAs are also belonged to the microRNA families with the same seed region sequence which may target the same set of genes and may functional redundancy and cooperation among different miRNAs [13,14] Cluster microRNAs are transcriptionally regulated by the same transcriptional factors or single cellular signaling pathways which can be regulated by single and simple activator or repressor protein. Cluster microRNAs are transcribed into large single RNA transcript which are rapidly spliced and processed by the Drosha-DGCR8 complex into individual primary microRNAs [15,16]. Then Dicer and RISC guide mature microRNA on their target genes.
According to miRBase (Version 22), the human genome has 1971 precursor microRNAs [17]. Among the human microRNAs, 468 microRNAs belong to 153 microRNA clusters [18]. These microRNA clusters are distributed across different chromosomes, and chromosome X, which has the highest microRNA clusters numbers among human chromosomes. There are 18 microRNA clusters on chromosome X. The largest microRNA cluster is on chromosome 19. It has 46 microRNAs. The second large microRNA cluster is on chromosome 14q32.2, which has 42 microRNAs [19]. Based on the microRNA distribution in the genomic region, microRNAs are categorized into intergenic, intronic, and exonic microRNA in the genome. About half microRNAs are intragenic microRNAs including one or few intronic and exonic microRNAs which embedded in intron or exon region of host gene on the same strand [20]. Intragenic microRNAs may either share the same promoter with host genes or have independent transcription by itself. During microRNA maturation process, the intragenic microRNAs are byproduct of host gene transcription which may compete to splicing nucleotide sequence elements of precursor mRNA or few in protein coding sequence and also protein–mRNA complex such as spliceosome and the microprocessor in mature mRNA formation [21,22]. These competitions may reduce the intragenic microRNA cluster formation. There are 76 intergenic clusters that have more than 65 intronic and exonic clusters in human genome. In contrast to regulation of intragenic microRNA cluster, the intergenic microRNA clusters have exclusive regulation mechanisms and tightly control in embryogenesis [23], signifying that intergenic microRNA clusters have more important roles in the control of gene expression and biological functions [18].
MicroRNA clusters not only have multiple crucial physiological roles under normal conditions related to the maintenance of cell hemostasis, but they also play roles in pathological conditions when they are dysregulated. microRNA clusters are involved in biological functions, including differentiation and development, nervous system regulation, immunity balance, DNA repair, cell junction and adhesion molecule regulation, intercellular commutation or as inner cellular secondary messengers, mammalian or cellular reproduction, metabolism homeostasis, cell reprogramming and the mesenchymal to epithelial transition or vice versa, organellar biogenesis and function, and responses to stressful conditions which we have summarized physiological microRNA cluster function in Table 1.
Specifically, cluster miR-17/92 has multiple roles from basic cell biology to whole physiological systems, such as DNA repair, secondary messenger, embryogenesis, developments, inter-cell commutation and immunity response [24]; cluster miR-23/24 is involved in stress response, acting as secondary messenger metabolism, organellar biogenesis and function, feedback loop, controlling immune response, and cell junction and adhesion [25,26]. Cluster miR-106b/25 is important in differentiation and development, cell reprogramming and as secondary messengers in stress and viral infection response [27]. Cluster miR-302b/367 has roles in differentiation and development, organellar biogenesis and function, metabolism, reproduction, cell reprogramming and mesenchymal to epithelial transition (MET). Cluster miR-439/136 and 379/656 are belonged to same genetic imprinting domain and they have critical function in differentiation, development, and nervous system regulation, metabolism, and cell to cell commutation [19,28]. Cluster miR-424/450b controls differentiation, development, MET, metabolism, and stress response [29]. Cluster miR-34b/34c is important in differentiation, development, stress response and reproduction [30]. Cluster miR-212/132 is important in nervous system development and regulation [31].
Both cluster miR-301b/130b and miR-508/513a are important in DNA repair system [32,33]; cluster miR-1/133a, miR-143/145, miR-15a/16-1 in cardiac development and disease and as secondary messengers; cluster miR-371a/373 in mammalian and cellular reproduction; cluster miR-106b/25 in metabolism and organelle biogenesis and function; cluster miR-512/519 in development and cell commutation. Both cluster miR-181a/181b and miR-183/182 are important in immunity response and they have their unique role in nervous system regulation and metabolism, respectively. Moreover, for the cluster miR-183/182, it is missed in the scheme of miR-96 that is part of the cluster and it is missed their roles in development and differentiation in the visual, auditory, vestibular, and olfactory systems [34,35,36]. The cluster miR-181 family, that is an evolutionary conserved family microRNAs organized in 3 clusters that have several different roles in central nervous system and immune system development, neurodegeneration and cancer [37,38,39]. Cluster miR-200b/429 is involved in feed-back loop regulation; cluster miR-199a/214 and miR-27a/27b contribute to organellar biogenesis and functions; cluster miR-29b/29a is crucial in nervous system regulation; cluster miR-193/365a controls metabolism. Cluster miR-206/133b contributes to differentiation and development and cluster miR-211/222 acts as cellular secondary messengers.
3. Regulation of microRNA Clusters
MicroRNA clusters have important roles in the maintenance of normal cellular hemostasis and commutation to surrounding microenvironments. MicroRNA cluster expression is controlled by multiple mechanisms by genetic or epigenetic events and the microRNAs. Mutation or single nucleotide polymorphisms (SNP) on microRNA cluster promoter region changes the transcription factor binding ability which regulate cluster microRNAs expression [41]. About 50% of microRNAs are located at fragile chromosome sites where the genomic regions suffer from copy-number changes in the mitosis [42]. Cancer cells are quickly dividing cells that easily cause replication stress which can promote accumulation of genetic mutation such as chromosome recombination, or translocation, amplification, deletion, or loss of heterozygosity upon alignments of homologous chromosome.
HNSCC is not common cancer model for mechanism studies of microRNA cluster regulation. Thus, we summary the potential mechanisms from other cancer types. In ovarian cancer [43,44,45], Burkitt’s lymphoma [46], and diffuse large B-cell lymphoma, miR-200b/429 [47] or the miR-17/92 cluster [24] undergo copy number amplification to stimulate mature-form microRNA expression. In contrast, miR-143/145 [48,49] and the miR-15a/16-1 [50] cluster are suppressed by chromosome deletion in leukemia. Genetic mutations or translocations also lead to the downregulation of miR-15a/16-1 in chronic lymphocytic leukemia (CLL) or miR-17/92 cluster expression in T-cell acute lymphoblastic leukemia [51,52] (Figure 2A). In addition to genomic translocation and mutation regulation of microRNA clusters, transcription factors (TFs) act on microRNA promoters and collaborate with RNA polymerase II (Pol II) to control microRNA cluster expression in cell-, tissue- or disease-specific manners. The aberrant expression of cancer specific transcription factors and mutation on microRNA cluster promoters contribute to cancer-specific microRNA cluster expression [53,54]. For example, MYC proto-oncogene controls miR-17/92 cluster expression [55] (Figure 2B). Epigenetic modifications are another important regulation mechanism for endogenous gene expression. De novo DNA methyltransferase (DNMT) 3A and 3B transfer adds a methyl group to the C5 positions of cytosine in CpG dinucleotides [56]. About 70% of gene promoters have a CpG island near the transcription factor start site [57,58]. DNA hypermethylation on the promoter CpG island leads to the recruitment of PRC complex repressors, which modifies specific histone methylation (H3K27me3) [59,60]. Eventually, HP1 joins the catalytic protein complex and converts euchromatin into heterochromatin [61,62]. In oral cancer, hypermethylation occurs on the miR-379/656 cluster promoter, where the genomic imprint for the non-coding RNA Meg3 differential methylation region (Meg3-DMR) also inhibits down-stream noncoding RNA and microRNA cluster expression [8,63] (Figure 2C). The abundance of single microRNA targets, like core transcription factors or key metabolic enzymes, can activate oncogenic signaling or change the intracellular metabolite balance to promote cancer progression [64,65]. Compare with single microRNA regulation, there are few studies focus on the regulation of microRNA clusters in cancer. Cluster microRNAs are evolution-conservation and usually coordinately expression of high homologue microRNAs which are usually belonged the same microRNA family. The coordinately expression of cluster microRNAs can not only accumulate contents of microRNAs but also the differential expression of microRNA targets. Minor regulation of the microRNA cluster has a huge impact on cancer tumorigenesis.
4. microRNA in Cancer
MicroRNAs was first discovered in Caenorhabditis elegans in 1993 [66,67]. Aberrant microRNA expression has been found in many diseases like cancer [68,69], neuron degeneration diseases [70], and developmental disabilities [71]. Cancer cells harbor various chromosome instabilities and mutations [72,73] which prompt the development of aberrant transcriptomes in both coding and non-coding genes. Aberrant microRNAs can be signaling transduction brokers between initiation genetic or epigenetic events and down-stream effector genes such as enzymes, signaling molecules, and traffic proteins for maintaining intracellular functions and intercellular commutations. Competing endogenous RNA (ceRNA) is a mechanism that RNA regulates other RNA transcripts by competing for shared microRNAs and microRNA binding elements [74,75,76,77] and include various products from transcriptome, such as protein coding genes, pseudogenes [78], long noncoding RNAs (lncRNAs) [79], and circular RNAs (circRNAs) [80,81]. Up- or down-regulation of one target may change the expression of other cognate targets by sequestering or releasing their shared microRNA molecules. Dysregulation of ceRNA network is important in cancer and other diseases [82,83,84]. MicroRNA families harbor seeding region sequence homologue, which is critical in regulation of ceRNA [85,86]. Some microRNA clusters are comprised of family microRNAs [87] and coordinate-expression [88] which hint microRNA clusters are important in ceRNA regulation. In ceRNA theory, microRNAs could act as hub molecules that promote cross-talk between different signaling pathways or biological functions. In cancer cells, aberrant microRNA molecules can be divided into master or driver molecules that can promote oncogenic cascades from a small signaling initiator toward a completely out of control situation. Differentially expressed microRNAs can also be defined as oncogenic microRNAs (OncomiRs) or tumor suppressor microRNAs (TSmiRs), based on the roles of microRNA regulation genes [89]. MicroRNAs that inhibit the expression of oncogenes, cell proliferation genes, or cancer progression genes can be called tumor suppressor microRNAs (Figure 1) [90,91,92]. In contrast to TSmiRs, microRNAs that target tumor suppressor genes or gene function while maintaining their epithelial status, like tight junction proteins on cell–cell hinge proteins, are called oncogenic microRNAs [93,94]. However, each type of microRNA can have different functions in different cell types [95].
5. microRNA in Cancer Microenvironment
Cancer cells are energy-hungry cells which demand endless nutrition for cell growth. Cancer cells refit surrounding normal cells into cancer-supporting fertile soil through secretion proteins such as cytokines, chemokines, and growth factors or exosomal microRNAs. Secretion proteins are paracrine ligands which bind directly to receptors on the surrounding cell surface and promote cell reorganization, migration, and proliferation. Tumor microenvironment comprises a mass of heterogeneous cell types, including, pericytes [96], endothelial cells [97], tumor-associated macrophages (TAMs) [98], and cancer-associated fibroblasts (CAFs) [99] all of which surround tumor cells as the core and cross-talk among surrounding normal cells. Angiogenesis is an essential process to provide oxygen and nutrient levels for tumor growth and metastasis [100]. During angiogenesis, blood vessels and stromal components, such as pericytes and endothelial cells, produces pro- and antiangiogenic factors and conduct many signaling pathways that allow vascular remodeling, neovascularization and communication with tumor cells. Vascular endothelial growth factor (VEGF) or basic fibroblast growth factor (bFGF) can stimulate pericytes and vascular endothelial cells reorganization into angiogenic vessels which can support nutrition and demand energy for cancer cell growth [101]. Unlike secretion proteins which are usually unstable in the serum and cancer microenvironments. Angiogenic genes may be regulated at the post-transcriptional level by microRNAs [100]. Exosomal microRNAs which are embedded in the small extracellular vesicle (sEV) and shaded by lipid bilayer have high stability and can act as paracrine or endocrine in the receipt cells through membrane-fusion. Exosomal microRNAs play critical role in interaction of tumor cells and tumor microenvironment and thereby modulates tumor progression and development [102]. Involvements of microRNAs in tumor cells, pericytes and endothelial cells during angiogenesis has been described in two comprehensive reviews by Salinas-Vera et al. [103] and Orso et al. [104].
Tumor-associated macrophages (TAMs) are the most abundant immune cells in the tumor microenvironment [105]. MicroRNAs notably influence the phenotype of TAMs through various targets and signal pathways during cancer progression and functions as important regulators in macrophage differentiation, functional polarization, and cellular crosstalk, which has been clearly described by Chen et al. [106]. Especially, microRNAs can be transported between tumor cells and macrophages via microvesicles and exosomes and facilitate crosstalk between tumor cells and macrophages, which is essential for tumor microenvironment formation and tumor progression [106].
Cancer-associated fibroblasts (CAFs) are a key component of the tumor microenvironment with diverse functions, including matrix deposition and remodeling, extensive reciprocal signaling interactions with cancer cells and crosstalk with other stromal cells by secreting various pro-inflammatory factors [107]. Changes of microRNAs expression in CAFs can be induced both by cancer cells and other stromal cells, which result from through direct interaction or by secreted paracrine factors or even by secreted microRNAs. MicroRNAs dysregulation contributes to CAFs phenotype and functions and assists their cancer promotion ability. CAFs interact with cancer cells by secreted microRNAs packaged in extracellular vesicles and exosomes and by directly releasing microRNAs into extracellular fluid. Wang Z. et al. provide the latest research about the relevance of microRNAs in the interaction between cancer cells and the CAFs. MicroRNAs from CAFs modulated cancer cells in various aspects, including cancer progression, metastasis, cancer metabolism, stemness and drug sensitivity [108].
MicroRNAs can circulate between various types of cells and regulate biologic effects, including supportive, inhibitory for tumor growth or dissemination. MicroRNAs can be released from cancer cells or microenvironment cells in various forms of vesicles or as “free” molecules secreted by active mechanisms [109]. Active participation of microRNAs in cancer cells and tumor microenvironment can be as therapeutic response predictors or therapeutic targets. By using sensitive detection technologies, microRNAs in body fluids or blood from patients with cancer can be detected and predicted a distinct microRNA expression profile [110]. MicroRNA is a critical mediator among tumor cells and tumor microenvironment and targeting microRNAs can be as promising therapeutic application.
6. Head and Neck Squamous Cell Carcinoma (HNSCC)
Head and neck squamous cell carcinomas (HNSCCs) represent the top lethal cancer type in the world. Although there have been great improvements in cancer management and treatment strategies in recent decades, which have extended the life quality and lifespan expectancy of most cancer patients. The HNSCC patient survival time or 5-year conditional survival time didn’t have a great improvement in the twentieth century [111]. The new target therapy, advanced radiotherapy, immune-checkpoint inhibitors treatment, and improvement surgeon skills could improve the HNSCC patient survival time [112]. HNSCCs exhibit a gender difference, with the male to female incidence ratio being 2.7:1. As for the mortality rate, males are again the most affected, with the ratio being 3.8:1. HNSCCs still cause close to 3000 deaths in Taiwan and 10,000 deaths in the United States per year [113]. Ninety percent of HNSCC cases originate in the oral cavity and are considered oral to squamous cell carcinomas. HNSCCs can be considered locoregional diseases; however, distant metastasis and local recurrence are major determinant factors in treatment management and cancer prognosis [114]. Cancers, including HNSCCs, are regarded as multifactorial diseases caused by various genetic or epigenetic modifications that induce the silencing of tumor suppressor genes, the activation of oncogenes, the escape of host immune detection and responses, and commutation with external microenvironments [115]. The major risk factors for the development of HNSCCs are alcohol consumption, habitual tobacco use, and betel nut chewing, as is common in southeast Asia. DNA damage by cigarette carcinogens such as tobacco-specific nitrosamines or arecoline from betel chewing are the major carcinogens responsible for HNSCC initiation [116,117]. Globally, smoking is the most preventable single carcinogen which cause of head and neck cancer with the highest users in the WHO European region (75.3%). Alcohol drinking is the second most common preventable factor which cause of HNSCC [118]. Chronic alcohol consumption and tobacco act synergistically in the development of head and neck cancer, while recent human papillomavirus (HPV) infection operates as an independent risk factor and is considered to have a large role in the development of oropharyngeal carcinogenesis [119,120,121]. Changes in tumorigenesis components including coding and non-coding genes modulate every step of cancer progression from tumorigenesis to angiogenesis, recurrence, and distant metastasis formation [112].
7. microRNA Deregulation in HNSCC Carcinogenesis
Abnormal microRNA expression has been reported during head and neck squamous carcinoma cancer (HNSCC) tumorigenesis and progression. Most clinical, experimental, and mechanistic studies of microRNA have focused on global patterns [122,123] of microRNA signatures which are not genetically linked or single microRNA functions in HNSCC. The functions of single oncogenic microRNAs (OncomiRs) or tumor suppressor microRNAs (TSmiR) are summarized in Table 2. miR-450a and miR-455 serve as OncomiRs which contribute tumor progression by targeting TMEM182 and UBE2B, respectively [124,125]. miR-31 has multiple targets, such as SIRT3, FIH, and ARID1A, and also can be cancer biomarkers in the plasma or serum [64,126,127,128]. miR-155 inhibits tumor suppressors and cell division cycle 73 (CDC73) in OSCC [129], and the up-regulation of miR-155 promotes OSCC cervical lymph node metastasis and is associated with a poor overall survival rate in patients [130,131,132]. In contrast to OncomiRs, miR-376c suppresses the expression of the transcription factor RUNX2 and inhibits the growth and metastasis of OSCC cells [63]. miR-379 and miR-30a regulate the expression of DNA methyltransferase 3B (DNMT3B) and the OSCC epigenome, preventing cancer progression [133]. miR-486 targets the cell membrane tyrosine kinase receptor DDR1 and prevents OSCC cell growth [134]. miR-99a governs insulin-like growth factor I receptor signaling [135]. However, most current microRNA studies focus on the association between one-microRNA and one target gene function in cancer. Although single microRNA may be sufficient in tumor biology, the cluster microRNAs have several coordinately regulated microRNAs which can accumulate contents of family microRNAs and aberrant expression of target genes from individual microRNAs in the cluster. Dysregulation microRNA cluster is an energy-saving and broad-impact procedure than single microRNA in cancer development or progression.
8. microRNA Cluster Deregulation in HNSCC
Maintaining life is an energy-consuming process. Cancer cells usually use all of their energy to grow or resist the external environment. Cluster microRNAs usually belong to a single microRNA family, which can enhance their regulation efficiency, and their coordinated regulation by a single genetic or epigenetic effect can save cellular energy for tumorigenesis and cancer progression. Regulation of microRNA cluster is an energy-saving and robust way to change cancer cell fate. Our previous studies focused on the expression, regulation mechanisms, and subsequent functions of aberrant microRNA clusters in HNSCC cancer tumorigenesis and progression [8,188]. DLK1-MEG3 imprinting locus has a huge miR-379/656 cluster which is epigenetic silenced in Taiwan OSCC patents by betel nut exposure. Arecoline, an active alkaloid from betel nut, cause DNA hypermethylation on differentially methylated region (DMR) on DLK1-MEG3 locus and silence whole imprinting locus expressions [8,63,133]. The miR-17/92 cluster has bi-functional role in OSCC growth and migration. The miR-17/92 cluster promotes OSCC growth by targeting FOXP1 [196] but it also suppresses cell migration and invasion by targeting integrin β8 [197]. Activin A is growth promoting autocrine [65]. Down regulation of miR-143/145 cluster synergistic promotes Activin A expression and serves as a poor prognostic marker in oral cancer patients [198]. Another tumor-suppressive microRNA cluster, microRNA-23b/27b, regulates oncogenic MET receptor tyrosine kinase expression and cell migration in OSCC cells [199]. In HNSCC, oncogenic miR-106b/25 cluster including miR-106b, miR-93 and miR-25 [200,201,202] affect cell proliferation [200] via targeting tumor suppressor p21/CDKN1A pathway [203] Upregulation of the miR-371/372/373 microRNA cluster enhances oral cancer oncogenicity and drug resistance [204]. Furthermore, the miR-503/450b cluster on the chromosome X promotes oral cancer progression and this cluster is controlled by inflammation cytokine interleukin 8 and signal transducer and activator of transcription 5 (STAT5) pathway [124,188]. Aberrant microRNA in HNSCC is listed in Table 2.
9. Translational Application of the miR-503/450b Cluster
Exosomes are small cell-membrane-like extracellular vesicles that contain cellular biomolecules such as proteins, mRNAs, non-coding RNAs, and microRNAs for intercellular communication from donor to recipient cells via target cell membrane fusion [205,206,207]. The simplest way for exosome cargo loading to occur is by simple diffusion, and the high abundance of biomolecules in donor cells can allow cargo to diffuse into exosomes by a concentration gradient [208]. The lipid-bilayer structure of exosomes can protect the inner cargo, preventing degradation by serum protease or ribonuclease and creating an extremely high level of stability for cargo molecules [209]. The high stability of exosomal biomolecules makes them suitable for liquid biopsy in clinical diagnosis [210]. microRNAs on the miR-503/450b cluster are the top-ranking expression microRNAs in OSCC tumors [8,211,212,213,214]. Furthermore, these microRNAs can be found in exosomes from T cells [215], dendritic cells [215], endothelial cells [216], and serum [217,218] in non-cancer diseases, such as early onset type 1 diabetes [219] and ischemia [220]. Taken together, these studies reveal the potential roles of the miR-503/450b cluster in immune and cancer microenvironment regulation, and microRNAs within this cluster are stable in the serum and circulation system so they can be delivered to recipient cells or can be detected by current detection technologies. Recently, the miR-503-containing exosome has been used in endothelial-derived breast cancer antitumor neoadjuvant chemotherapy [221]. Thus, microRNAs within the miR-503/450b cluster not only have diagnosis or prognosis functions, but also have potential for use in future cancer therapies.
10. Summary and Perspectives
According to previous microRNA studies, the microRNA cluster is a missing link in cancer research fields. Regulation of the microRNA cluster is an energy-efficient procedure involving tumorigenesis, cell growth, metastasis, and cancer progression. A tiny change in the microRNA cluster can tremendously alter cell functions. Usually, microRNAs in a cluster belong to the same family with high sequence homology. Thus, they can increase the regulation intensity in cancer cells or can be easily amplified in liquid biopsy. Previously, small RNA molecules could only work as cancer biomarkers and were detected by liquid biopsy. Improvements in nucleotide chemistry and modification technology have made RNAs much more stable in modern pharmaceuticals [222,223,224]. Inclisiran is the first U.S. Food and Drug Administration (FDA)-approved small interfering RNA drug that can inhibit PCSK9 translation to treat atherosclerotic cardiovascular disease (ASCVD) including hypercholesterolemia or mixed dyslipidemia [225,226,227,228]. The distribution of extracellular microRNA by antisense oligonucleotides may prevent intercellular communication among cancer microenvironments and cancer progression. microRNA research may have exciting prospects for cancer diagnostics, prognostics, and therapeutics. In the past few decades, there has been a tremendous change from the profiling of altered microRNA expression in cancers to clinical trials with microRNAs as therapy candidates. In the near future, we hope that microRNA replacement and inhibition therapies will enter pre-clinical and clinical pilot studies and potentially be used as personalized medicines.
Author Contributions
Preparation of original draft by L.-J.L. and W.-M.C. W.-M.C. and M.H. participated in the conceptualization. L.-J.L. and W.-M.C. contributed to critical editing of manuscript. Funding was provided by W.-M.C. and M.H. Data acquisition was done by L.-J.L. and W.-M.C. Supervision, M.H. All authors have read and agreed to the published version of the manuscript.
Funding
The research was financially supported by Genomics Research Center to Michael Hsiao and by Taipei Medical University [TMU108-AE1-B52] to Wei-Min Chang.
Conflicts of Interest
The authors declare that they have no competing interests.
References
- Chen, Q.; Yan, M.; Cao, Z.; Li, X.; Zhang, Y.; Shi, J.; Feng, G.H.; Peng, H.; Zhang, X.; Zhang, Y.; et al. Sperm tsRNAs contribute to intergenerational inheritance of an acquired metabolic disorder. Science 2016, 351, 397–400. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Green, D.; Fraser, W.D.; Dalmay, T. Transfer RNA-derived small RNAs in the cancer transcriptome. Pflug. Arch. 2016, 468, 1041–1047. [Google Scholar] [CrossRef] [Green Version]
- Mattick, J.S.; Makunin, I.V. Non-coding RNA. Hum. Mol. Genet. 2006, 15 (Suppl. 1), R17–R29. [Google Scholar] [CrossRef] [Green Version]
- Kirchner, S.; Ignatova, Z. Emerging roles of tRNA in adaptive translation, signalling dynamics and disease. Nat. Rev. Genet. 2015, 16, 98–112. [Google Scholar] [CrossRef]
- Wei, J.W.; Huang, K.; Yang, C.; Kang, C.S. Non-coding RNAs as regulators in epigenetics (Review). Oncol. Rep. 2017, 37, 3–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lambert, M.; Benmoussa, A.; Provost, P. Small Non-Coding RNAs Derived from Eukaryotic Ribosomal RNA. Non-Coding RNA 2019, 5, 16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goriaux, C.; Theron, E.; Brasset, E.; Vaury, C. History of the discovery of a master locus producing piRNAs: The flamenco/COM locus in Drosophila melanogaster. Front. Genet. 2014, 5, 257. [Google Scholar] [CrossRef] [Green Version]
- Shiah, S.G.; Hsiao, J.R.; Chang, W.M.; Chen, Y.W.; Jin, Y.T.; Wong, T.Y.; Huang, J.S.; Tsai, S.T.; Hsu, Y.M.; Chou, S.T.; et al. Downregulated miR329 and miR410 promote the proliferation and invasion of oral squamous cell carcinoma by targeting Wnt-7b. Cancer Res. 2014, 74, 7560–7572. [Google Scholar] [CrossRef] [Green Version]
- Lee, H.; Han, S.; Kwon, C.S.; Lee, D. Biogenesis and regulation of the let-7 miRNAs and their functional implications. Protein Cell 2016, 7, 100–113. [Google Scholar] [CrossRef] [Green Version]
- Gross, N.; Kropp, J.; Khatib, H. MicroRNA Signaling in Embryo Development. Biology 2017, 6, 34. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.T.; Risom, T.; Strauss, W.M. Evolutionary conservation of microRNA regulatory circuits: An examination of microRNA gene complexity and conserved microRNA-target interactions through metazoan phylogeny. DNA Cell Biol. 2007, 26, 209–218. [Google Scholar] [CrossRef] [PubMed]
- Altuvia, Y.; Landgraf, P.; Lithwick, G.; Elefant, N.; Pfeffer, S.; Aravin, A.; Brownstein, M.J.; Tuschl, T.; Margalit, H. Clustering and conservation patterns of human microRNAs. Nucleic Acids Res. 2005, 33, 2697–2706. [Google Scholar] [CrossRef] [PubMed]
- Krol, J.; Loedige, I.; Filipowicz, W. The widespread regulation of microRNA biogenesis, function and decay. Nat. Rev. Genet. 2010, 11, 597–610. [Google Scholar] [CrossRef]
- Nie, M.; Deng, Z.L.; Liu, J.; Wang, D.Z. Noncoding RNAs, Emerging Regulators of Skeletal Muscle Development and Diseases. Biomed. Res. Int. 2015, 2015, 676575. [Google Scholar] [CrossRef] [Green Version]
- Bortolin-Cavaille, M.L.; Dance, M.; Weber, M.; Cavaille, J. C19MC microRNAs are processed from introns of large Pol-II, non-protein-coding transcripts. Nucleic Acids Res. 2009, 37, 3464–3473. [Google Scholar] [CrossRef] [PubMed]
- Mong, E.F.; Yang, Y.; Akat, K.M.; Canfield, J.; VanWye, J.; Lockhart, J.; Tsibris, J.C.M.; Schatz, F.; Lockwood, C.J.; Tuschl, T.; et al. Chromosome 19 microRNA cluster enhances cell reprogramming by inhibiting epithelial-to-mesenchymal transition. Sci. Rep. 2020, 10, 3029. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kozomara, A.; Birgaoanu, M.; Griffiths-Jones, S. miRBase: From microRNA sequences to function. Nucleic Acids Res. 2019, 47, D155–D162. [Google Scholar] [CrossRef]
- Kabekkodu, S.P.; Shukla, V.; Varghese, V.K.; D’Souza, J.; Chakrabarty, S.; Satyamoorthy, K. Clustered miRNAs and their role in biological functions and diseases. Biol. Rev. Camb. Philos. Soc. 2018, 93, 1955–1986. [Google Scholar] [CrossRef]
- Kabekkodu, S.P.; Shukla, V.; Varghese, V.K.; Adiga, D.; Vethil Jishnu, P.; Chakrabarty, S.; Satyamoorthy, K. Cluster miRNAs and cancer: Diagnostic, prognostic and therapeutic opportunities. Wiley Interdiscip. Rev. RNA 2020, 11, e1563. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Shyr, Y.; Cai, J.; Liu, Q. Interplay between miRNAs and host genes and their role in cancer. Brief. Funct. Genomics 2018, 18, 255–266. [Google Scholar] [CrossRef] [Green Version]
- Melamed, Z.; Levy, A.; Ashwal-Fluss, R.; Lev-Maor, G.; Mekahel, K.; Atias, N.; Gilad, S.; Sharan, R.; Levy, C.; Kadener, S.; et al. Alternative splicing regulates biogenesis of miRNAs located across exon-intron junctions. Mol. Cell 2013, 50, 869–881. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.K.; Kim, V.N. Processing of intronic microRNAs. EMBO J. 2007, 26, 775–783. [Google Scholar] [CrossRef] [Green Version]
- Mathieu, J.; Ruohola-Baker, H. Regulation of stem cell populations by microRNAs. Adv. Exp. Med. Biol. 2013, 786, 329–351. [Google Scholar] [CrossRef] [Green Version]
- Mogilyansky, E.; Rigoutsos, I. The miR-17/92 cluster: A comprehensive update on its genomics, genetics, functions and increasingly important and numerous roles in health and disease. Cell Death Differ. 2013, 20, 1603–1614. [Google Scholar] [CrossRef]
- Wu, C.J.; Cho, S.; Huang, H.Y.; Lu, C.H.; Russ, J.; Cruz, L.O.; da Cunha, F.F.; Chen, M.C.; Lin, L.L.; Warner, L.M.; et al. MiR-23~27~24-mediated control of humoral immunity reveals a TOX-driven regulatory circuit in follicular helper T cell differentiation. Sci. Adv. 2019, 5, eaaw1715. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bang, C.; Fiedler, J.; Thum, T. Cardiovascular importance of the microRNA-23/27/24 family. Microcirculation 2012, 19, 208–214. [Google Scholar] [CrossRef]
- Mehlich, D.; Garbicz, F.; Wlodarski, P.K. The emerging roles of the polycistronic miR-106b approximately 25 cluster in cancer—A comprehensive review. Biomed. Pharmacother. 2018, 107, 1183–1195. [Google Scholar] [CrossRef]
- Jishnu, P.V.; Jayaram, P.; Shukla, V.; Varghese, V.K.; Pandey, D.; Sharan, K.; Chakrabarty, S.; Satyamoorthy, K.; Kabekkodu, S.P. Prognostic role of 14q32.31 miRNA cluster in various carcinomas: A systematic review and meta-analysis. Clin. Exp. Metastasis 2020, 37, 31–46. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Liang, R.; Tandon, N.; Matthews, E.R.; Shrestha, S.; Yang, J.; Soibam, B.; Yang, J.; Liu, Y. H19X-encoded miR-424(322)/-503 cluster: Emerging roles in cell differentiation, proliferation, plasticity and metabolism. Cell. Mol. Life Sci. 2019, 76, 903–920. [Google Scholar] [CrossRef]
- Wu, J.; Bao, J.; Kim, M.; Yuan, S.; Tang, C.; Zheng, H.; Mastick, G.S.; Xu, C.; Yan, W. Two miRNA clusters, miR-34b/c and miR-449, are essential for normal brain development, motile ciliogenesis, and spermatogenesis. Proc. Natl. Acad. Sci. USA 2014, 111, E2851–E2857. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wanet, A.; Tacheny, A.; Arnould, T.; Renard, P. miR-212/132 expression and functions: Within and beyond the neuronal compartment. Nucleic Acids Res. 2012, 40, 4742–4753. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramalho-Carvalho, J.; Graca, I.; Gomez, A.; Oliveira, J.; Henrique, R.; Esteller, M.; Jeronimo, C. Downregulation of miR-130b~301b cluster is mediated by aberrant promoter methylation and impairs cellular senescence in prostate cancer. J. Hematol. Oncol. 2017, 10, 43. [Google Scholar] [CrossRef] [Green Version]
- Yoshida, K.; Yokoi, A.; Kato, T.; Ochiya, T.; Yamamoto, Y. The clinical impact of intra- and extracellular miRNAs in ovarian cancer. Cancer Sci. 2020, 111, 3435–3444. [Google Scholar] [CrossRef]
- Fan, J.; Jia, L.; Li, Y.; Ebrahim, S.; May-Simera, H.; Wood, A.; Morell, R.J.; Liu, P.; Lei, J.; Kachar, B.; et al. Maturation arrest in early postnatal sensory receptors by deletion of the miR-183/96/182 cluster in mouse. Proc. Natl. Acad. Sci. USA 2017, 114, E4271–E4280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carrella, S.; Banfi, S.; Karali, M. Sophisticated gene regulation for a complex physiological system: The role of non-coding RNAs in photoreceptor cells. Front. Cell Dev. Biol. 2020, 8, 1825. [Google Scholar]
- Karali, M.; Banfi, S. Non-coding RNAs in retinal development and function. Hum. Genet. 2019, 138, 957–971. [Google Scholar] [CrossRef]
- Indrieri, A.; Carrella, S.; Carotenuto, P.; Banfi, S.; Franco, B. The Pervasive Role of the miR-181 Family in Development, Neurodegeneration, and Cancer. Int. J. Mol. Sci. 2020, 21, 2092. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grewers, Z.; Krueger, A. MicroRNA miR-181-A Rheostat for TCR Signaling in Thymic Selection and Peripheral T-Cell Function. Int. J. Mol. Sci. 2020, 21, 6200. [Google Scholar] [CrossRef]
- Henao-Mejia, J.; Williams, A.; Goff, L.A.; Staron, M.; Licona-Limon, P.; Kaech, S.M.; Nakayama, M.; Rinn, J.L.; Flavell, R.A. The microRNA miR-181 is a critical cellular metabolic rheostat essential for NKT cell ontogenesis and lymphocyte development and homeostasis. Immunity 2013, 38, 984–997. [Google Scholar] [CrossRef] [Green Version]
- Kandettu, A.; Radhakrishnan, R.; Chakrabarty, S.; Sriharikrishnaa, S.; Kabekkodu, S.P. The emerging role of miRNA clusters in breast cancer progression. Biochim. Biophys. Acta Rev. Cancer 2020, 1874, 188413. [Google Scholar] [CrossRef]
- Berindan-Neagoe, I.; Monroig Pdel, C.; Pasculli, B.; Calin, G.A. MicroRNAome genome: A treasure for cancer diagnosis and therapy. CA Cancer J. Clin. 2014, 64, 311–336. [Google Scholar] [CrossRef] [PubMed]
- Calin, G.A.; Sevignani, C.; Dumitru, C.D.; Hyslop, T.; Noch, E.; Yendamuri, S.; Shimizu, M.; Rattan, S.; Bullrich, F.; Negrini, M.; et al. Human microRNA genes are frequently located at fragile sites and genomic regions involved in cancers. Proc. Natl. Acad. Sci. USA 2004, 101, 2999–3004. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, L.; Huang, J.; Yang, N.; Greshock, J.; Megraw, M.S.; Giannakakis, A.; Liang, S.; Naylor, T.L.; Barchetti, A.; Ward, M.R.; et al. microRNAs exhibit high frequency genomic alterations in human cancer. Proc. Natl. Acad. Sci. USA 2006, 103, 9136–9141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.W.; Kim, J.W.; Kim, Y.T.; Kim, J.H.; Kim, S.; Yoon, B.S.; Nam, E.J.; Kim, H.Y. Analysis of chromosomal changes in serous ovarian carcinoma using high-resolution array comparative genomic hybridization: Potential predictive markers of chemoresistant disease. Genes Chromosomes Cancer 2007, 46, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Guan, W.; Cui, H.; Huang, P.; Chun, W.J.; Lee, J.W.; Kim, H.; Zou, H. miR-200b/200a/429 Cluster Stimulates Ovarian Cancer Development by Targeting ING5. J. Oncol. 2020, 2020, 3404059. [Google Scholar] [CrossRef]
- Robaina, M.C.; Faccion, R.S.; Mazzoccoli, L.; Rezende, L.M.; Queiroga, E.; Bacchi, C.E.; Thomas-Tikhonenko, A.; Klumb, C.E. miR-17-92 cluster components analysis in Burkitt lymphoma: Overexpression of miR-17 is associated with poor prognosis. Ann. Hematol. 2016, 95, 881–891. [Google Scholar] [CrossRef]
- Lin, Z.; Wang, X.; Fewell, C.; Cameron, J.; Yin, Q.; Flemington, E.K. Differential expression of the miR-200 family microRNAs in epithelial and B cells and regulation of Epstein-Barr virus reactivation by the miR-200 family member miR-429. J. Virol. 2010, 84, 7892–7897. [Google Scholar] [CrossRef] [Green Version]
- Poli, V.; Secli, L.; Avalle, L. The Microrna-143/145 Cluster in Tumors: A Matter of Where and When. Cancers 2020, 12, 708. [Google Scholar] [CrossRef] [Green Version]
- Almeida, M.I.; Calin, G.A. The miR-143/miR-145 cluster and the tumor microenvironment: Unexpected roles. Genome Med. 2016, 8, 29. [Google Scholar] [CrossRef] [Green Version]
- Aqeilan, R.I.; Calin, G.A.; Croce, C.M. miR-15a and miR-16-1 in cancer: Discovery, function and future perspectives. Cell Death Differ. 2010, 17, 215–220. [Google Scholar] [CrossRef] [Green Version]
- Kasar, S.; Underbayev, C.; Hassan, M.; Ilev, I.; Degheidy, H.; Bauer, S.; Marti, G.; Lutz, C.; Raveche, E.; Batish, M. Alterations in the mir-15a/16-1 Loci Impairs Its Processing and Augments B-1 Expansion in De Novo Mouse Model of Chronic Lymphocytic Leukemia (CLL). PLoS ONE 2016, 11, e0149331. [Google Scholar] [CrossRef]
- Ye, F. MicroRNA expression and activity in T-cell acute lymphoblastic leukemia. Oncotarget 2018, 9, 5445–5458. [Google Scholar] [CrossRef] [PubMed]
- Ji, M.; Rao, E.; Ramachandrareddy, H.; Shen, Y.; Jiang, C.; Chen, J.; Hu, Y.; Rizzino, A.; Chan, W.C.; Fu, K.; et al. The miR-17-92 microRNA cluster is regulated by multiple mechanisms in B-cell malignancies. Am. J. Pathol. 2011, 179, 1645–1656. [Google Scholar] [CrossRef]
- Yan, Y.; Hanse, E.A.; Stedman, K.; Benson, J.M.; Lowman, X.H.; Subramanian, S.; Kelekar, A. Transcription factor C/EBP-beta induces tumor-suppressor phosphatase PHLPP2 through repression of the miR-17-92 cluster in differentiating AML cells. Cell Death Differ. 2016, 23, 1232–1242. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Choi, P.S.; Casey, S.C.; Dill, D.L.; Felsher, D.W. MYC through miR-17-92 suppresses specific target genes to maintain survival, autonomous proliferation, and a neoplastic state. Cancer Cell 2014, 26, 262–272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Z.M.; Lu, R.; Wang, P.; Yu, Y.; Chen, D.; Gao, L.; Liu, S.; Ji, D.; Rothbart, S.B.; Wang, Y.; et al. Structural basis for DNMT3A-mediated de novo DNA methylation. Nature 2018, 554, 387–391. [Google Scholar] [CrossRef]
- Saxonov, S.; Berg, P.; Brutlag, D.L. A genome-wide analysis of CpG dinucleotides in the human genome distinguishes two distinct classes of promoters. Proc. Natl. Acad. Sci. USA 2006, 103, 1412–1417. [Google Scholar] [CrossRef] [Green Version]
- Deaton, A.M.; Bird, A. CpG islands and the regulation of transcription. Genes Dev. 2011, 25, 1010–1022. [Google Scholar] [CrossRef] [Green Version]
- Chan, H.L.; Morey, L. Emerging Roles for Polycomb-Group Proteins in Stem Cells and Cancer. Trends Biochem. Sci. 2019, 44, 688–700. [Google Scholar] [CrossRef]
- Richly, H.; Aloia, L.; Di Croce, L. Roles of the Polycomb group proteins in stem cells and cancer. Cell Death Dis. 2011, 2, e204. [Google Scholar] [CrossRef] [Green Version]
- Tamaru, H. Confining euchromatin/heterochromatin territory: Jumonji crosses the line. Genes Dev. 2010, 24, 1465–1478. [Google Scholar] [CrossRef] [Green Version]
- Hathaway, N.A.; Bell, O.; Hodges, C.; Miller, E.L.; Neel, D.S.; Crabtree, G.R. Dynamics and memory of heterochromatin in living cells. Cell 2012, 149, 1447–1460. [Google Scholar] [CrossRef] [Green Version]
- Chang, W.M.; Lin, Y.F.; Su, C.Y.; Peng, H.Y.; Chang, Y.C.; Lai, T.C.; Wu, G.H.; Hsu, Y.M.; Chi, L.H.; Hsiao, J.R.; et al. Dysregulation of RUNX2/Activin-A Axis upon miR-376c Downregulation Promotes Lymph Node Metastasis in Head and Neck Squamous Cell Carcinoma. Cancer Res. 2016, 76, 7140–7150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kao, Y.Y.; Chou, C.H.; Yeh, L.Y.; Chen, Y.F.; Chang, K.W.; Liu, C.J.; Fan Chiang, C.Y.; Lin, S.C. MicroRNA miR-31 targets SIRT3 to disrupt mitochondrial activity and increase oxidative stress in oral carcinoma. Cancer Lett. 2019, 456, 40–48. [Google Scholar] [CrossRef]
- Chang, W.M.; Lin, Y.F.; Su, C.Y.; Peng, H.Y.; Chang, Y.C.; Hsiao, J.R.; Chen, C.L.; Chang, J.Y.; Shieh, Y.S.; Hsiao, M.; et al. Parathyroid Hormone-Like Hormone is a Poor Prognosis Marker of Head and Neck Cancer and Promotes Cell Growth via RUNX2 Regulation. Sci. Rep. 2017, 7, 41131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, R.C.; Feinbaum, R.L.; Ambros, V. The C. elegans heterochronic gene lin-4 encodes small RNAs with antisense complementarity to lin-14. Cell 1993, 75, 843–854. [Google Scholar] [CrossRef]
- Wightman, B.; Ha, I.; Ruvkun, G. Posttranscriptional regulation of the heterochronic gene lin-14 by lin-4 mediates temporal pattern formation in C. elegans. Cell 1993, 75, 855–862. [Google Scholar] [CrossRef]
- Adams, B.D.; Kasinski, A.L.; Slack, F.J. Aberrant regulation and function of microRNAs in cancer. Curr. Biol. 2014, 24, R762–R776. [Google Scholar] [CrossRef] [Green Version]
- Koshizuka, K.; Hanazawa, T.; Fukumoto, I.; Kikkawa, N.; Okamoto, Y.; Seki, N. The microRNA signatures: Aberrantly expressed microRNAs in head and neck squamous cell carcinoma. J. Hum. Genet. 2017, 62, 3–13. [Google Scholar] [CrossRef] [PubMed]
- De Paola, E.; Verdile, V.; Paronetto, M.P. Dysregulation of microRNA metabolism in motor neuron diseases: Novel biomarkers and potential therapeutics. Noncoding RNA Res. 2019, 4, 15–22. [Google Scholar] [CrossRef]
- Sehovic, E.; Spahic, L.; Smajlovic-Skenderagic, L.; Pistoljevic, N.; Dzanko, E.; Hajdarpasic, A. Identification of developmental disorders including autism spectrum disorder using salivary miRNAs in children from Bosnia and Herzegovina. PLoS ONE 2020, 15, e0232351. [Google Scholar] [CrossRef] [PubMed]
- Jefford, C.E.; Irminger-Finger, I. Mechanisms of chromosome instability in cancers. Crit. Rev. Oncol. Hematol. 2006, 59, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Fukasawa, K. Centrosome amplification, chromosome instability and cancer development. Cancer Lett. 2005, 230, 6–19. [Google Scholar] [CrossRef]
- Chiu, H.S.; Martinez, M.R.; Bansal, M.; Subramanian, A.; Golub, T.R.; Yang, X.; Sumazin, P.; Califano, A. High-throughput validation of ceRNA regulatory networks. BMC Genom. 2017, 18, 418. [Google Scholar] [CrossRef] [PubMed]
- Salmena, L.; Poliseno, L.; Tay, Y.; Kats, L.; Pandolfi, P.P. A ceRNA hypothesis: The Rosetta Stone of a hidden RNA language? Cell 2011, 146, 353–358. [Google Scholar] [CrossRef] [Green Version]
- Karreth, F.A.; Tay, Y.; Perna, D.; Ala, U.; Tan, S.M.; Rust, A.G.; DeNicola, G.; Webster, K.A.; Weiss, D.; Perez-Mancera, P.A.; et al. In vivo identification of tumor- suppressive PTEN ceRNAs in an oncogenic BRAF-induced mouse model of melanoma. Cell 2011, 147, 382–395. [Google Scholar] [CrossRef] [Green Version]
- Tay, Y.; Kats, L.; Salmena, L.; Weiss, D.; Tan, S.M.; Ala, U.; Karreth, F.; Poliseno, L.; Provero, P.; Di Cunto, F.; et al. Coding-independent regulation of the tumor suppressor PTEN by competing endogenous mRNAs. Cell 2011, 147, 344–357. [Google Scholar] [CrossRef] [Green Version]
- Poliseno, L.; Salmena, L.; Zhang, J.; Carver, B.; Haveman, W.J.; Pandolfi, P.P. A coding-independent function of gene and pseudogene mRNAs regulates tumour biology. Nature 2010, 465, 1033–1038. [Google Scholar] [CrossRef] [Green Version]
- Cesana, M.; Cacchiarelli, D.; Legnini, I.; Santini, T.; Sthandier, O.; Chinappi, M.; Tramontano, A.; Bozzoni, I. A long noncoding RNA controls muscle differentiation by functioning as a competing endogenous RNA. Cell 2011, 147, 358–369. [Google Scholar] [CrossRef] [Green Version]
- Memczak, S.; Jens, M.; Elefsinioti, A.; Torti, F.; Krueger, J.; Rybak, A.; Maier, L.; Mackowiak, S.D.; Gregersen, L.H.; Munschauer, M.; et al. Circular RNAs are a large class of animal RNAs with regulatory potency. Nature 2013, 495, 333–338. [Google Scholar] [CrossRef]
- Hansen, T.B.; Jensen, T.I.; Clausen, B.H.; Bramsen, J.B.; Finsen, B.; Damgaard, C.K.; Kjems, J. Natural RNA circles function as efficient microRNA sponges. Nature 2013, 495, 384–388. [Google Scholar] [CrossRef]
- Qi, X.; Zhang, D.H.; Wu, N.; Xiao, J.H.; Wang, X.; Ma, W. ceRNA in cancer: Possible functions and clinical implications. J. Med. Genet. 2015, 52, 710–718. [Google Scholar] [CrossRef]
- Cardenas, J.; Balaji, U.; Gu, J. Cerina: Systematic circRNA functional annotation based on integrative analysis of ceRNA interactions. Sci. Rep. 2020, 10, 22165. [Google Scholar] [CrossRef] [PubMed]
- Gao, C.; Li, H.; Zhuang, J.; Zhang, H.; Wang, K.; Yang, J.; Liu, C.; Liu, L.; Zhou, C.; Sun, C. The construction and analysis of ceRNA networks in invasive breast cancer: A study based on The Cancer Genome Atlas. Cancer Manag. Res. 2019, 11, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chiu, H.S.; Martinez, M.R.; Komissarova, E.V.; Llobet-Navas, D.; Bansal, M.; Paull, E.O.; Silva, J.; Yang, X.; Sumazin, P.; Califano, A. The number of titrated microRNA species dictates ceRNA regulation. Nucleic Acids Res. 2018, 46, 4354–4369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chiu, H.S.; Llobet-Navas, D.; Yang, X.; Chung, W.J.; Ambesi-Impiombato, A.; Iyer, A.; Kim, H.R.; Seviour, E.G.; Luo, Z.; Sehgal, V.; et al. Cupid: Simultaneous reconstruction of microRNA-target and ceRNA networks. Genome Res. 2015, 25, 257–267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dambal, S.; Shah, M.; Mihelich, B.; Nonn, L. The microRNA-183 cluster: The family that plays together stays together. Nucleic Acids Res. 2015, 43, 7173–7188. [Google Scholar] [CrossRef] [PubMed]
- Guo, L.; Yang, S.; Zhao, Y.; Zhang, H.; Wu, Q.; Chen, F. Global analysis of miRNA gene clusters and gene families reveals dynamic and coordinated expression. Biomed. Res. Int. 2014, 2014, 782490. [Google Scholar] [CrossRef]
- Esquela-Kerscher, A.; Slack, F.J. Oncomirs—MicroRNAs with a role in cancer. Nat. Rev. Cancer 2006, 6, 259–269. [Google Scholar] [CrossRef]
- Zhang, B.; Pan, X.; Cobb, G.P.; Anderson, T.A. microRNAs as oncogenes and tumor suppressors. Dev. Biol. 2007, 302, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Tsai, W.C.; Hsu, S.D.; Hsu, C.S.; Lai, T.C.; Chen, S.J.; Shen, R.; Huang, Y.; Chen, H.C.; Lee, C.H.; Tsai, T.F.; et al. MicroRNA-122 plays a critical role in liver homeostasis and hepatocarcinogenesis. J. Clin. Investig. 2012, 122, 2884–2897. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsu, S.H.; Wang, B.; Kota, J.; Yu, J.; Costinean, S.; Kutay, H.; Yu, L.; Bai, S.; La Perle, K.; Chivukula, R.R.; et al. Essential metabolic, anti-inflammatory, and anti-tumorigenic functions of miR-122 in liver. J. Clin. Investig. 2012, 122, 2871–2883. [Google Scholar] [CrossRef] [Green Version]
- Fabbri, M.; Bottoni, A.; Shimizu, M.; Spizzo, R.; Nicoloso, M.S.; Rossi, S.; Barbarotto, E.; Cimmino, A.; Adair, B.; Wojcik, S.E.; et al. Association of a microRNA/TP53 feedback circuitry with pathogenesis and outcome of B-cell chronic lymphocytic leukemia. JAMA 2011, 305, 59–67. [Google Scholar] [CrossRef]
- Lv, L.; Wang, Q.; Yang, Y.; Ji, H. MicroRNA495 targets Notch1 to prohibit cell proliferation and invasion in oral squamous cell carcinoma. Mol. Med. Rep. 2019, 19, 693–702. [Google Scholar] [CrossRef] [Green Version]
- Oliveto, S.; Mancino, M.; Manfrini, N.; Biffo, S. Role of microRNAs in translation regulation and cancer. World J. Biol. Chem. 2017, 8, 45–56. [Google Scholar] [CrossRef]
- Thijssen, V.L.; Paulis, Y.W.; Nowak-Sliwinska, P.; Deumelandt, K.L.; Hosaka, K.; Soetekouw, P.M.; Cimpean, A.M.; Raica, M.; Pauwels, P.; van den Oord, J.J.; et al. Targeting PDGF-mediated recruitment of pericytes blocks vascular mimicry and tumor growth. J. Pathol. 2018, 246, 447–458. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Wang, X.; Li, Z.; Zhang, Z.; Zhang, Y. Chemokine receptor 7 targets the vascular endothelial growth factor via the AKT/ERK pathway to regulate angiogenesis in colon cancer. Cancer Med. 2019, 8, 5327–5340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarode, P.; Zheng, X.; Giotopoulou, G.A.; Weigert, A.; Kuenne, C.; Gunther, S.; Friedrich, A.; Gattenlohner, S.; Stiewe, T.; Brune, B.; et al. Reprogramming of tumor-associated macrophages by targeting beta-catenin/FOSL2/ARID5A signaling: A potential treatment of lung cancer. Sci. Adv. 2020, 6, eaaz6105. [Google Scholar] [CrossRef]
- Cangkrama, M.; Wietecha, M.; Mathis, N.; Okumura, R.; Ferrarese, L.; Al-Nuaimi, D.; Antsiferova, M.; Dummer, R.; Innocenti, M.; Werner, S. A paracrine activin A-mDia2 axis promotes squamous carcinogenesis via fibroblast reprogramming. EMBO Mol. Med. 2020, 12, e11466. [Google Scholar] [CrossRef]
- Weis, S.M.; Cheresh, D.A. Tumor angiogenesis: Molecular pathways and therapeutic targets. Nat. Med. 2011, 17, 1359–1370. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.; Wang, J.; Deng, X.; Xiong, F.; Zhang, S.; Gong, Z.; Li, X.; Cao, K.; Deng, H.; He, Y.; et al. The role of microenvironment in tumor angiogenesis. J. Exp. Clin. Cancer Res. 2020, 39, 204. [Google Scholar] [CrossRef] [PubMed]
- Denton, A.E.; Roberts, E.W.; Fearon, D.T. Stromal Cells in the Tumor Microenvironment. Adv. Exp. Med. Biol. 2018, 1060, 99–114. [Google Scholar] [CrossRef]
- Salinas-Vera, Y.M.; Marchat, L.A.; Gallardo-Rincon, D.; Ruiz-Garcia, E.; Astudillo-De La Vega, H.; Echavarria-Zepeda, R.; Lopez-Camarillo, C. AngiomiRs: MicroRNAs driving angiogenesis in cancer (Review). Int. J. Mol. Med. 2019, 43, 657–670. [Google Scholar] [CrossRef] [Green Version]
- Orso, F.; Quirico, L.; Dettori, D.; Coppo, R.; Virga, F.; Ferreira, L.C.; Paoletti, C.; Baruffaldi, D.; Penna, E.; Taverna, D. Role of miRNAs in tumor and endothelial cell interactions during tumor progression. Semin. Cancer Biol. 2020, 60, 214–224. [Google Scholar] [CrossRef] [PubMed]
- Noy, R.; Pollard, J.W. Tumor-associated macrophages: From mechanisms to therapy. Immunity 2014, 41, 49–61. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.; Liu, J.M.; Luo, Y.P. MicroRNAs in tumor immunity: Functional regulation in tumor-associated macrophages. J. Zhejiang Univ. Sci. B 2020, 21, 12–28. [Google Scholar] [CrossRef] [PubMed]
- Sahai, E.; Astsaturov, I.; Cukierman, E.; DeNardo, D.G.; Egeblad, M.; Evans, R.M.; Fearon, D.; Greten, F.R.; Hingorani, S.R.; Hunter, T.; et al. A framework for advancing our understanding of cancer-associated fibroblasts. Nat. Rev. Cancer 2020, 20, 174–186. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Tan, Y.; Yu, W.; Zheng, S.; Zhang, S.; Sun, L.; Ding, K. Small role with big impact: miRNAs as communicators in the cross-talk between cancer-associated fibroblasts and cancer cells. Int. J. Biol. Sci. 2017, 13, 339–348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, M.; Chen, J.; Su, F.; Yu, B.; Su, F.; Lin, L.; Liu, Y.; Huang, J.D.; Song, E. Microvesicles secreted by macrophages shuttle invasion-potentiating microRNAs into breast cancer cells. Mol. Cancer 2011, 10, 117. [Google Scholar] [CrossRef] [Green Version]
- Nakamura, K.; Sawada, K.; Yoshimura, A.; Kinose, Y.; Nakatsuka, E.; Kimura, T. Clinical relevance of circulating cell-free microRNAs in ovarian cancer. Mol. Cancer 2016, 15, 48. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.J.; Kim, J.H. Increasing incidence and improving survival of oral tongue squamous cell carcinoma. Sci. Rep. 2020, 10, 7877. [Google Scholar] [CrossRef]
- Chow, L.Q.M. Head and Neck Cancer. N. Engl. J. Med. 2020, 382, 60–72. [Google Scholar] [CrossRef] [PubMed]
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer statistics, 2020. CA Cancer J. Clin. 2020, 70, 7–30. [Google Scholar] [CrossRef]
- Takes, R.P.; Rinaldo, A.; Silver, C.E.; Haigentz, M., Jr.; Woolgar, J.A.; Triantafyllou, A.; Mondin, V.; Paccagnella, D.; de Bree, R.; Shaha, A.R.; et al. Distant metastases from head and neck squamous cell carcinoma. Part I. Basic aspects. Oral Oncol. 2012, 48, 775–779. [Google Scholar] [CrossRef]
- Hanahan, D.; Weinberg, R.A. Hallmarks of cancer: The next generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murphy, S.E.; Carmella, S.G.; Idris, A.M.; Hoffmann, D. Uptake and metabolism of carcinogenic levels of tobacco-specific nitrosamines by Sudanese snuff dippers. Cancer Epidemiol. Biomark. Prev. 1994, 3, 423–428. [Google Scholar]
- Wang, M.; Xiao, C.; Ni, P.; Yu, J.J.; Wang, X.W.; Sun, H. Correlation of Betel Quid with Oral Cancer from 1998 to 2017: A Study Based on Bibliometric Analysis. Chin. Med. J. 2018, 131, 1975–1982. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.; Singhavi, H.; Sathe, P.; Gnanamoorthy, A.; Chaturvedi, P. Addictions causing head-and-neck cancers. Indian J. Med. Paediatr. Oncol. 2020, 41, 510. [Google Scholar]
- Maruyama, R.; Suzuki, H.; Yamamoto, E.; Imai, K.; Shinomura, Y. Emerging links between epigenetic alterations and dysregulation of noncoding RNAs in cancer. Tumour Biol. 2012, 33, 277–285. [Google Scholar] [CrossRef]
- Pelucchi, C.; Gallus, S.; Garavello, W.; Bosetti, C.; La Vecchia, C. Cancer risk associated with alcohol and tobacco use: Focus on upper aero-digestive tract and liver. Alcohol. Res. Health 2006, 29, 193–198. [Google Scholar] [PubMed]
- Gonzalez-Ramirez, I.; Garcia-Cuellar, C.; Sanchez-Perez, Y.; Granados-Garcia, M. DNA methylation in oral squamous cell carcinoma: Molecular mechanisms and clinical implications. Oral Dis. 2011, 17, 771–778. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.A.; Weng, S.L.; Yang, S.F.; Chou, C.H.; Huang, W.C.; Tu, S.J.; Chang, T.H.; Huang, C.N.; Jong, Y.J.; Huang, H.D. A Three-MicroRNA Signature as a Potential Biomarker for the Early Detection of Oral Cancer. Int. J. Mol. Sci. 2018, 19, 758. [Google Scholar] [CrossRef] [Green Version]
- Farah, C.S.; Fox, S.A.; Dalley, A.J. Integrated miRNA-mRNA spatial signature for oral squamous cell carcinoma: A prospective profiling study of Narrow Band Imaging guided resection. Sci. Rep. 2018, 8, 823. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsing, E.W.; Shiah, S.G.; Peng, H.Y.; Chen, Y.W.; Chuu, C.P.; Hsiao, J.R.; Lyu, P.C.; Chang, J.Y. TNF-alpha-induced miR-450a mediates TMEM182 expression to promote oral squamous cell carcinoma motility. PLoS ONE 2019, 14, e0213463. [Google Scholar] [CrossRef]
- Cheng, C.M.; Shiah, S.G.; Huang, C.C.; Hsiao, J.R.; Chang, J.Y. Up-regulation of miR-455-5p by the TGF-beta-SMAD signalling axis promotes the proliferation of oral squamous cancer cells by targeting UBE2B. J. Pathol. 2016, 240, 38–49. [Google Scholar] [CrossRef]
- Liu, C.J.; Lin, S.C.; Yang, C.C.; Cheng, H.W.; Chang, K.W. Exploiting salivary miR-31 as a clinical biomarker of oral squamous cell carcinoma. Head Neck 2012, 34, 219–224. [Google Scholar] [CrossRef]
- Liu, C.J.; Kao, S.Y.; Tu, H.F.; Tsai, M.M.; Chang, K.W.; Lin, S.C. Increase of microRNA miR-31 level in plasma could be a potential marker of oral cancer. Oral Dis. 2010, 16, 360–364. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.J.; Tsai, M.M.; Hung, P.S.; Kao, S.Y.; Liu, T.Y.; Wu, K.J.; Chiou, S.H.; Lin, S.C.; Chang, K.W. miR-31 ablates expression of the HIF regulatory factor FIH to activate the HIF pathway in head and neck carcinoma. Cancer Res. 2010, 70, 1635–1644. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rather, M.I.; Nagashri, M.N.; Swamy, S.S.; Gopinath, K.S.; Kumar, A. Oncogenic microRNA-155 down-regulates tumor suppressor CDC73 and promotes oral squamous cell carcinoma cell proliferation: Implications for cancer therapeutics. J. Biol. Chem. 2013, 288, 608–618. [Google Scholar] [CrossRef] [Green Version]
- Ni, Y.H.; Huang, X.F.; Wang, Z.Y.; Han, W.; Deng, R.Z.; Mou, Y.B.; Ding, L.; Hou, Y.Y.; Hu, Q.G. Upregulation of a potential prognostic biomarker, miR-155, enhances cell proliferation in patients with oral squamous cell carcinoma. Oral Surg. Oral Med. Oral Pathol. Oral Radiol. 2014, 117, 227–233. [Google Scholar] [CrossRef]
- Baba, O.; Hasegawa, S.; Nagai, H.; Uchida, F.; Yamatoji, M.; Kanno, N.I.; Yamagata, K.; Sakai, S.; Yanagawa, T.; Bukawa, H. MicroRNA-155-5p is associated with oral squamous cell carcinoma metastasis and poor prognosis. J. Oral Pathol. Med. 2016, 45, 248–255. [Google Scholar] [CrossRef] [Green Version]
- Lerner, C.; Wemmert, S.; Bochen, F.; Kulas, P.; Linxweiler, M.; Hasenfus, A.; Heinzelmann, J.; Leidinger, P.; Backes, C.; Meese, E.; et al. Characterization of miR-146a and miR-155 in blood, tissue and cell lines of head and neck squamous cell carcinoma patients and their impact on cell proliferation and migration. J. Cancer Res. Clin. Oncol. 2016, 142, 757–766. [Google Scholar] [CrossRef]
- Shiah, S.G.; Hsiao, J.R.; Chang, H.J.; Hsu, Y.M.; Wu, G.H.; Peng, H.Y.; Chou, S.T.; Kuo, C.C.; Chang, J.Y. MiR-30a and miR-379 modulate retinoic acid pathway by targeting DNA methyltransferase 3B in oral cancer. J. Biomed. Sci. 2020, 27, 46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chou, S.T.; Peng, H.Y.; Mo, K.C.; Hsu, Y.M.; Wu, G.H.; Hsiao, J.R.; Lin, S.F.; Wang, H.D.; Shiah, S.G. MicroRNA-486-3p functions as a tumor suppressor in oral cancer by targeting DDR1. J. Exp. Clin. Cancer Res. 2019, 38, 281. [Google Scholar] [CrossRef] [PubMed]
- Yen, Y.C.; Shiah, S.G.; Chu, H.C.; Hsu, Y.M.; Hsiao, J.R.; Chang, J.Y.; Hung, W.C.; Liao, C.T.; Cheng, A.J.; Lu, Y.C.; et al. Reciprocal regulation of microRNA-99a and insulin-like growth factor I receptor signaling in oral squamous cell carcinoma cells. Mol. Cancer 2014, 13, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nohata, N.; Hanazawa, T.; Kikkawa, N.; Sakurai, D.; Sasaki, K.; Chiyomaru, T.; Kawakami, K.; Yoshino, H.; Enokida, H.; Nakagawa, M.; et al. Identification of novel molecular targets regulated by tumor suppressive miR-1/miR-133a in maxillary sinus squamous cell carcinoma. Int. J. Oncol. 2011, 39, 1099–1107. [Google Scholar] [CrossRef]
- Nohata, N.; Sone, Y.; Hanazawa, T.; Fuse, M.; Kikkawa, N.; Yoshino, H.; Chiyomaru, T.; Kawakami, K.; Enokida, H.; Nakagawa, M.; et al. miR-1 as a tumor suppressive microRNA targeting TAGLN2 in head and neck squamous cell carcinoma. Oncotarget 2011, 2, 29–42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koshizuka, K.; Hanazawa, T.; Fukumoto, I.; Kikkawa, N.; Matsushita, R.; Mataki, H.; Mizuno, K.; Okamoto, Y.; Seki, N. Dual-receptor (EGFR and c-MET) inhibition by tumor-suppressive miR-1 and miR-206 in head and neck squamous cell carcinoma. J. Hum. Genet. 2017, 62, 113–121. [Google Scholar] [CrossRef]
- Peng, C.Y.; Liao, Y.W.; Lu, M.Y.; Yu, C.H.; Yu, C.C.; Chou, M.Y. Downregulation of miR-1 enhances tumorigenicity and invasiveness in oral squamous cell carcinomas. J. Formos. Med. Assoc. 2017, 116, 782–789. [Google Scholar] [CrossRef]
- Lu, J.; Zhao, F.P.; Peng, Z.; Zhang, M.W.; Lin, S.X.; Liang, B.J.; Zhang, B.; Liu, X.; Wang, L.; Li, G.; et al. EZH2 promotes angiogenesis through inhibition of miR-1/Endothelin-1 axis in nasopharyngeal carcinoma. Oncotarget 2014, 5, 11319–11332. [Google Scholar] [CrossRef]
- Jiang, L.; Liu, X.; Chen, Z.; Jin, Y.; Heidbreder, C.E.; Kolokythas, A.; Wang, A.; Dai, Y.; Zhou, X. MicroRNA-7 targets IGF1R (insulin-like growth factor 1 receptor) in tongue squamous cell carcinoma cells. Biochem. J. 2010, 432, 199–205. [Google Scholar] [CrossRef]
- Minor, J.; Wang, X.; Zhang, F.; Song, J.; Jimeno, A.; Wang, X.J.; Lu, X.; Gross, N.; Kulesz-Martin, M.; Wang, D.; et al. Methylation of microRNA-9 is a specific and sensitive biomarker for oral and oropharyngeal squamous cell carcinomas. Oral Oncol. 2012, 48, 73–78. [Google Scholar] [CrossRef] [Green Version]
- Yu, T.; Liu, K.; Wu, Y.; Fan, J.; Chen, J.; Li, C.; Yang, Q.; Wang, Z. MicroRNA-9 inhibits the proliferation of oral squamous cell carcinoma cells by suppressing expression of CXCR4 via the Wnt/beta-catenin signaling pathway. Oncogene 2014, 33, 5017–5027. [Google Scholar] [CrossRef] [Green Version]
- Chen, Z.; Jin, Y.; Yu, D.; Wang, A.; Mahjabeen, I.; Wang, C.; Liu, X.; Zhou, X. Down-regulation of the microRNA-99 family members in head and neck squamous cell carcinoma. Oral Oncol. 2012, 48, 686–691. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, Y.; Tymen, S.D.; Chen, D.; Fang, Z.J.; Zhao, Y.; Dragas, D.; Dai, Y.; Marucha, P.T.; Zhou, X. MicroRNA-99 family targets AKT/mTOR signaling pathway in dermal wound healing. PLoS ONE 2013, 8, e64434. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, R.; Mani, R.S.; Russo, N.; Scanlon, C.S.; Tsodikov, A.; Jing, X.; Cao, Q.; Palanisamy, N.; Metwally, T.; Inglehart, R.C.; et al. The tumor suppressor gene rap1GAP is silenced by miR-101-mediated EZH2 overexpression in invasive squamous cell carcinoma. Oncogene 2011, 30, 4339–4349. [Google Scholar] [CrossRef] [Green Version]
- Zheng, M.; Jiang, Y.P.; Chen, W.; Li, K.D.; Liu, X.; Gao, S.Y.; Feng, H.; Wang, S.S.; Jiang, J.; Ma, X.R.; et al. Snail and Slug collaborate on EMT and tumor metastasis through miR-101-mediated EZH2 axis in oral tongue squamous cell carcinoma. Oncotarget 2015, 6, 6797–6810. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, X.R.; Wen, X.; He, Q.M.; Li, Y.Q.; Ren, X.Y.; Yang, X.J.; Zhang, J.; Wang, Y.Q.; Ma, J.; Liu, N. MicroRNA-101 inhibits invasion and angiogenesis through targeting ITGA3 and its systemic delivery inhibits lung metastasis in nasopharyngeal carcinoma. Cell Death Dis. 2017, 8, e2566. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.Y.; Liu, Z.J.; He, H.; Zhang, C.; Wang, Y.L. MicroRNA-101-3p suppresses cell proliferation, invasion and enhances chemotherapeutic sensitivity in salivary gland adenoid cystic carcinoma by targeting Pim-1. Am. J. Cancer Res. 2015, 5, 3015–3029. [Google Scholar]
- Peng, M.; Pang, C. MicroRNA-140-5p inhibits the tumorigenesis of oral squamous cell carcinoma by targeting p21-activated kinase 4. Cell Biol. Int. 2019. [Google Scholar] [CrossRef] [PubMed]
- Qiao, Z.; Zou, Y.; Zhao, H. MicroRNA-140-5p inhibits salivary adenoid cystic carcinoma progression and metastasis via targeting survivin. Cancer Cell Int. 2019, 19, 301. [Google Scholar] [CrossRef]
- Severino, P.; Bruggemann, H.; Andreghetto, F.M.; Camps, C.; Klingbeil Mde, F.; de Pereira, W.O.; Soares, R.M.; Moyses, R.; Wunsch-Filho, V.; Mathor, M.B.; et al. MicroRNA expression profile in head and neck cancer: HOX-cluster embedded microRNA-196a and microRNA-10b dysregulation implicated in cell proliferation. BMC Cancer 2013, 13, 533. [Google Scholar] [CrossRef]
- Li, G.; Wu, Z.; Peng, Y.; Liu, X.; Lu, J.; Wang, L.; Pan, Q.; He, M.L.; Li, X.P. MicroRNA-10b induced by Epstein-Barr virus-encoded latent membrane protein-1 promotes the metastasis of human nasopharyngeal carcinoma cells. Cancer Lett. 2010, 299, 29–36. [Google Scholar] [CrossRef]
- Lu, Y.C.; Chen, Y.J.; Wang, H.M.; Tsai, C.Y.; Chen, W.H.; Huang, Y.C.; Fan, K.H.; Tsai, C.N.; Huang, S.F.; Kang, C.J.; et al. Oncogenic function and early detection potential of miRNA-10b in oral cancer as identified by microRNA profiling. Cancer Prev. Res. 2012, 5, 665–674. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Wang, J.; Chen, Z.; Wang, K.; Shi, L. MicroRNA-1-3p inhibits the proliferation and migration of oral squamous cell carcinoma cells by targeting DKK1. Biochem. Cell Biol. 2018, 96, 355–364. [Google Scholar] [CrossRef]
- Chen, Y.F.; Wei, Y.Y.; Yang, C.C.; Liu, C.J.; Yeh, L.Y.; Chou, C.H.; Chang, K.W.; Lin, S.C. miR-125b suppresses oral oncogenicity by targeting the anti-oxidative gene PRXL2A. Redox Biol. 2019, 22, 101140. [Google Scholar] [CrossRef]
- Jiang, L.; Dai, Y.; Liu, X.; Wang, C.; Wang, A.; Chen, Z.; Heidbreder, C.E.; Kolokythas, A.; Zhou, X. Identification and experimental validation of G protein alpha inhibiting activity polypeptide 2 (GNAI2) as a microRNA-138 target in tongue squamous cell carcinoma. Hum. Genet. 2011, 129, 189–197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jing, P.; Sa, N.; Liu, X.; Liu, X.; Xu, W. MicroR-140-5p suppresses tumor cell migration and invasion by targeting ADAM10-mediated Notch1 signaling pathway in hypopharyngeal squamous cell carcinoma. Exp. Mol. Pathol. 2016, 100, 132–138. [Google Scholar] [CrossRef] [PubMed]
- Kai, Y.; Peng, W.; Ling, W.; Jiebing, H.; Zhuan, B. Reciprocal effects between microRNA-140-5p and ADAM10 suppress migration and invasion of human tongue cancer cells. Biochem. Biophys. Res. Commun. 2014, 448, 308–314. [Google Scholar] [CrossRef] [PubMed]
- Manikandan, M.; Deva Magendhra Rao, A.K.; Rajkumar, K.S.; Rajaraman, R.; Munirajan, A.K. Altered levels of miR-21, miR-125b-2*, miR-138, miR-155, miR-184, and miR-205 in oral squamous cell carcinoma and association with clinicopathological characteristics. J. Oral Pathol. Med. 2015, 44, 792–800. [Google Scholar] [CrossRef]
- Shin, K.H.; Bae, S.D.; Hong, H.S.; Kim, R.H.; Kang, M.K.; Park, N.H. miR-181a shows tumor suppressive effect against oral squamous cell carcinoma cells by downregulating K-ras. Biochem. Biophys. Res. Commun. 2011, 404, 896–902. [Google Scholar] [CrossRef] [PubMed]
- He, Q.; Zhou, X.; Li, S.; Jin, Y.; Chen, Z.; Chen, D.; Cai, Y.; Liu, Z.; Zhao, T.; Wang, A. MicroRNA-181a suppresses salivary adenoid cystic carcinoma metastasis by targeting MAPK-Snai2 pathway. Biochim. Biophys. Acta 2013, 1830, 5258–5266. [Google Scholar] [CrossRef]
- Liu, M.; Wang, J.; Huang, H.; Hou, J.; Zhang, B.; Wang, A. miR-181a-Twist1 pathway in the chemoresistance of tongue squamous cell carcinoma. Biochem. Biophys. Res. Commun. 2013, 441, 364–370. [Google Scholar] [CrossRef]
- Yang, C.C.; Hung, P.S.; Wang, P.W.; Liu, C.J.; Chu, T.H.; Cheng, H.W.; Lin, S.C. miR-181 as a putative biomarker for lymph-node metastasis of oral squamous cell carcinoma. J. Oral Pathol. Med. 2011, 40, 397–404. [Google Scholar] [CrossRef] [PubMed]
- Supic, G.; Zeljic, K.; Rankov, A.D.; Kozomara, R.; Nikolic, A.; Radojkovic, D.; Magic, Z. miR-183 and miR-21 expression as biomarkers of progression and survival in tongue carcinoma patients. Clin. Oral Investig. 2018, 22, 401–409. [Google Scholar] [CrossRef]
- Cervigne, N.K.; Reis, P.P.; Machado, J.; Sadikovic, B.; Bradley, G.; Galloni, N.N.; Pintilie, M.; Jurisica, I.; Perez-Ordonez, B.; Gilbert, R.; et al. Identification of a microRNA signature associated with progression of leukoplakia to oral carcinoma. Hum. Mol. Genet. 2009, 18, 4818–4829. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Carvalho, A.C.; Scapulatempo-Neto, C.; Maia, D.C.; Evangelista, A.F.; Morini, M.A.; Carvalho, A.L.; Vettore, A.L. Accuracy of microRNAs as markers for the detection of neck lymph node metastases in patients with head and neck squamous cell carcinoma. BMC Med. 2015, 13, 108. [Google Scholar] [CrossRef] [Green Version]
- Obayashi, M.; Yoshida, M.; Tsunematsu, T.; Ogawa, I.; Sasahira, T.; Kuniyasu, H.; Imoto, I.; Abiko, Y.; Xu, D.; Fukunaga, S.; et al. microRNA-203 suppresses invasion and epithelial-mesenchymal transition induction via targeting NUAK1 in head and neck cancer. Oncotarget 2016, 7, 8223–8239. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.; Zhang, Y.; Zhou, D.; Cao, G.; Wu, Y. miR-204 enhances p27 mRNA stability by targeting Brd4 in head and neck squamous cell carcinoma. Oncol. Lett. 2018, 16, 4179–4184. [Google Scholar] [CrossRef]
- Yu, C.C.; Chen, P.N.; Peng, C.Y.; Yu, C.H.; Chou, M.Y. Suppression of miR-204 enables oral squamous cell carcinomas to promote cancer stemness, EMT traits, and lymph node metastasis. Oncotarget 2016, 7, 20180–20192. [Google Scholar] [CrossRef] [Green Version]
- Ma, L.; Deng, X.; Wu, M.; Zhang, G.; Huang, J. Down-regulation of miRNA-204 by LMP-1 enhances CDC42 activity and facilitates invasion of EBV-associated nasopharyngeal carcinoma cells. FEBS Lett. 2014, 588, 1562–1570. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Li, F.; Zhou, X. miR-204-5p regulates cell proliferation and metastasis through inhibiting CXCR4 expression in OSCC. Biomed. Pharmacother. 2016, 82, 202–207. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.J.; Shen, W.G.; Liu, C.J.; Chen, Y.W.; Lu, H.H.; Tsai, M.M.; Lin, S.C. miR-221 and miR-222 expression increased the growth and tumorigenesis of oral carcinoma cells. J. Oral Pathol. Med. 2011, 40, 560–566. [Google Scholar] [CrossRef]
- Zhou, L.; Jiang, F.; Chen, X.; Liu, Z.; Ouyang, Y.; Zhao, W.; Yu, D. Downregulation of miR-221/222 by a microRNA sponge promotes apoptosis in oral squamous cell carcinoma cells through upregulation of PTEN. Oncol. Lett. 2016, 12, 4419–4426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, F.; Zhao, W.; Zhou, L.; Liu, Z.; Li, W.; Yu, D. MiR-222 targeted PUMA to improve sensitization of UM1 cells to cisplatin. Int. J. Mol. Sci. 2014, 15, 22128–22141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, X.; Yu, J.; Jiang, L.; Wang, A.; Shi, F.; Ye, H.; Zhou, X. MicroRNA-222 regulates cell invasion by targeting matrix metalloproteinase 1 (MMP1) and manganese superoxide dismutase 2 (SOD2) in tongue squamous cell carcinoma cell lines. Cancer Genomics Proteomics 2009, 6, 131–139. [Google Scholar]
- Tachibana, H.; Sho, R.; Takeda, Y.; Zhang, X.; Yoshida, Y.; Narimatsu, H.; Otani, K.; Ishikawa, S.; Fukao, A.; Asao, H.; et al. Circulating miR-223 in Oral Cancer: Its Potential as a Novel Diagnostic Biomarker and Therapeutic Target. PLoS ONE 2016, 11, e0159693. [Google Scholar] [CrossRef]
- Jiang, L.; Lv, L.; Liu, X.; Jiang, X.; Yin, Q.; Hao, Y.; Xiao, L. MiR-223 promotes oral squamous cell carcinoma proliferation and migration by regulating FBXW7. Cancer Biomark. 2019, 24, 325–334. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Wang, A.; Heidbreder, C.E.; Jiang, L.; Yu, J.; Kolokythas, A.; Huang, L.; Dai, Y.; Zhou, X. MicroRNA-24 targeting RNA-binding protein DND1 in tongue squamous cell carcinoma. FEBS Lett. 2010, 584, 4115–4120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.L.; Tao, B.; Li, J.; Mao, X.; He, W.; Chen, Q.B. Melatonin Inhibits the Progression of Oral Squamous Cell Carcinoma via Inducing miR-25-5p Expression by Directly Targeting NEDD9. Front. Oncol. 2020. [Google Scholar] [CrossRef]
- Jia, L.F.; Wei, S.B.; Gan, Y.H.; Guo, Y.; Gong, K.; Mitchelson, K.; Cheng, J.; Yu, G.Y. Expression, regulation and roles of miR-26a and MEG3 in tongue squamous cell carcinoma. Int. J. Cancer 2014, 135, 2282–2293. [Google Scholar] [CrossRef] [PubMed]
- Wei, Z.; Chang, K.; Fan, C.; Zhang, Y. MiR-26a/miR-26b represses tongue squamous cell carcinoma progression by targeting PAK1. Cancer Cell Int. 2020, 20, 82. [Google Scholar] [CrossRef]
- Fukumoto, I.; Hanazawa, T.; Kinoshita, T.; Kikkawa, N.; Koshizuka, K.; Goto, Y.; Nishikawa, R.; Chiyomaru, T.; Enokida, H.; Nakagawa, M.; et al. MicroRNA expression signature of oral squamous cell carcinoma: Functional role of microRNA-26a/b in the modulation of novel cancer pathways. Br. J. Cancer 2015, 112, 891–900. [Google Scholar] [CrossRef] [Green Version]
- Lu, W.C.; Liu, C.J.; Tu, H.F.; Chung, Y.T.; Yang, C.C.; Kao, S.Y.; Chang, K.W.; Lin, S.C. miR-31 targets ARID1A and enhances the oncogenicity and stemness of head and neck squamous cell carcinoma. Oncotarget 2016, 7, 57254–57267. [Google Scholar] [CrossRef] [Green Version]
- Manikandan, M.; Deva Magendhra Rao, A.K.; Arunkumar, G.; Rajkumar, K.S.; Rajaraman, R.; Munirajan, A.K. Down Regulation of miR-34a and miR-143 May Indirectly Inhibit p53 in Oral Squamous Cell Carcinoma: A Pilot Study. Asian Pac. J. Cancer Prev. 2015, 16, 7619–7625. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, Y.; Wang, X. miR-34a targets BCL-2 to suppress the migration and invasion of sinonasal squamous cell carcinoma. Oncol. Lett. 2018, 16, 6566–6572. [Google Scholar] [CrossRef]
- Yeh, L.Y.; Yang, C.C.; Wu, H.L.; Kao, S.Y.; Liu, C.J.; Chen, Y.F.; Lin, S.C.; Chang, K.W. The miR-372-ZBTB7A Oncogenic Axis Suppresses TRAIL-R2 Associated Drug Sensitivity in Oral Carcinoma. Front. Oncol. 2020, 10, 47. [Google Scholar] [CrossRef] [Green Version]
- Peng, H.Y.; Jiang, S.S.; Hsiao, J.R.; Hsiao, M.; Hsu, Y.M.; Wu, G.H.; Chang, W.M.; Chang, J.Y.; Jin, S.L.; Shiah, S.G. IL-8 induces miR-424-5p expression and modulates SOCS2/STAT5 signaling pathway in oral squamous cell carcinoma. Mol. Oncol. 2016, 10, 895–909. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fei, Y.; Shan, W.; Chen, X. MiR-503-5p functions as an oncogene in oral squamous cell carcinoma by targeting Smad7. Histol. Histopathol. 2020, 35, 893–901. [Google Scholar] [CrossRef] [PubMed]
- Liborio-Kimura, T.N.; Jung, H.M.; Chan, E.K. miR-494 represses HOXA10 expression and inhibits cell proliferation in oral cancer. Oral Oncol. 2015, 51, 151–157. [Google Scholar] [CrossRef] [PubMed]
- He, H.; Liao, X.; Yang, Q.; Liu, Y.; Peng, Y.; Zhong, H.; Yang, J.; Zhang, H.; Yu, Z.; Zuo, Y.; et al. MicroRNA-494-3p Promotes Cell Growth, Migration, and Invasion of Nasopharyngeal Carcinoma by Targeting Sox7. Technol. Cancer Res. Treat. 2018, 17, 1533033818809993. [Google Scholar] [CrossRef]
- He, K.; Tong, D.; Zhang, S.; Cai, D.; Wang, L.; Yang, Y.; Gao, L.; Chang, S.; Guo, B.; Song, T.; et al. miRNA-99b-3p functions as a potential tumor suppressor by targeting glycogen synthase kinase-3beta in oral squamous cell carcinoma Tca-8113 cells. Int. J. Oncol. 2015, 47, 1528–1536. [Google Scholar] [CrossRef]
- Yan, B.; Fu, Q.; Lai, L.; Tao, X.; Fei, Y.; Shen, J.; Chen, Z.; Wang, Q. Downregulation of microRNA 99a in oral squamous cell carcinomas contributes to the growth and survival of oral cancer cells. Mol. Med. Rep. 2012, 6, 675–681. [Google Scholar] [CrossRef] [PubMed]
- Kuo, Y.Z.; Tai, Y.H.; Lo, H.I.; Chen, Y.L.; Cheng, H.C.; Fang, W.Y.; Lin, S.H.; Yang, C.L.; Tsai, S.T.; Wu, L.W. MiR-99a exerts anti-metastasis through inhibiting myotubularin-related protein 3 expression in oral cancer. Oral Dis. 2014, 20, e65–e75. [Google Scholar] [CrossRef]
- Jakymiw, A.; Patel, R.S.; Deming, N.; Bhattacharyya, I.; Shah, P.; Lamont, R.J.; Stewart, C.M.; Cohen, D.M.; Chan, E.K. Overexpression of dicer as a result of reduced let-7 MicroRNA levels contributes to increased cell proliferation of oral cancer cells. Genes Chromosomes Cancer 2010, 49, 549–559. [Google Scholar] [CrossRef] [Green Version]
- Guo, J.; Wen, N.; Yang, S.; Guan, X.; Cang, S. MiR-92a regulates oral squamous cell carcinoma (OSCC) cell growth by targeting FOXP1 expression. Biomed. Pharmacother. 2018, 104, 77–86. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.C.; Yang, Y.J.; Li, Y.J.; Chen, S.T.; Lin, B.R.; Wu, T.S.; Lin, S.K.; Kuo, M.Y.; Tan, C.T. MicroRNA-17/20a functions to inhibit cell migration and can be used a prognostic marker in oral squamous cell carcinoma. Oral Oncol. 2013, 49, 923–931. [Google Scholar] [CrossRef] [PubMed]
- Bufalino, A.; Cervigne, N.K.; de Oliveira, C.E.; Fonseca, F.P.; Rodrigues, P.C.; Macedo, C.C.; Sobral, L.M.; Miguel, M.C.; Lopes, M.A.; Paes Leme, A.F.; et al. Low miR-143/miR-145 Cluster Levels Induce Activin A Overexpression in Oral Squamous Cell Carcinomas, Which Contributes to Poor Prognosis. PLoS ONE 2015, 10, e0136599. [Google Scholar] [CrossRef] [PubMed]
- Fukumoto, I.; Koshizuka, K.; Hanazawa, T.; Kikkawa, N.; Matsushita, R.; Kurozumi, A.; Kato, M.; Okato, A.; Okamoto, Y.; Seki, N. The tumor-suppressive microRNA-23b/27b cluster regulates the MET oncogene in oral squamous cell carcinoma. Int. J. Oncol. 2016, 49, 1119–1129. [Google Scholar] [CrossRef]
- Hui, A.B.; Lenarduzzi, M.; Krushel, T.; Waldron, L.; Pintilie, M.; Shi, W.; Perez-Ordonez, B.; Jurisica, I.; O’Sullivan, B.; Waldron, J.; et al. Comprehensive MicroRNA profiling for head and neck squamous cell carcinomas. Clin. Cancer Res. 2010, 16, 1129–1139. [Google Scholar] [CrossRef] [Green Version]
- Cao, P.; Zhou, L.; Zhang, J.; Zheng, F.; Wang, H.; Ma, D.; Tian, J. Comprehensive expression profiling of microRNAs in laryngeal squamous cell carcinoma. Head Neck 2013, 35, 720–728. [Google Scholar] [CrossRef]
- Ramdas, L.; Giri, U.; Ashorn, C.L.; Coombes, K.R.; El-Naggar, A.; Ang, K.K.; Story, M.D. miRNA expression profiles in head and neck squamous cell carcinoma and adjacent normal tissue. Head Neck 2009, 31, 642–654. [Google Scholar] [CrossRef]
- Ivanovska, I.; Ball, A.S.; Diaz, R.L.; Magnus, J.F.; Kibukawa, M.; Schelter, J.M.; Kobayashi, S.V.; Lim, L.; Burchard, J.; Jackson, A.L.; et al. MicroRNAs in the miR-106b family regulate p21/CDKN1A and promote cell cycle progression. Mol. Cell. Biol. 2008, 28, 2167–2174. [Google Scholar] [CrossRef] [Green Version]
- Lin, S.C.; Wu, H.L.; Yeh, L.Y.; Yang, C.C.; Kao, Y.K.; Chang, K.W. Activation of the miR-371/372/373 miRNA Cluster Enhances Oncogenicity and Drug Resistance in Oral Carcinoma Cells. Int. J. Mol. Sci. 2020, 21, 9442. [Google Scholar] [CrossRef]
- Thery, C.; Witwer, K.W.; Aikawa, E.; Alcaraz, M.J.; Anderson, J.D.; Andriantsitohaina, R.; Antoniou, A.; Arab, T.; Archer, F.; Atkin-Smith, G.K.; et al. Minimal information for studies of extracellular vesicles 2018 (MISEV2018): A position statement of the International Society for Extracellular Vesicles and update of the MISEV2014 guidelines. J. Extracell. Vesicles 2018, 7, 1535750. [Google Scholar] [CrossRef] [Green Version]
- Yanez-Mo, M.; Siljander, P.R.; Andreu, Z.; Zavec, A.B.; Borras, F.E.; Buzas, E.I.; Buzas, K.; Casal, E.; Cappello, F.; Carvalho, J.; et al. Biological properties of extracellular vesicles and their physiological functions. J. Extracell. Vesicles 2015, 4, 27066. [Google Scholar] [CrossRef] [Green Version]
- Behera, J.; Tyagi, N. Exosomes: Mediators of bone diseases, protection, and therapeutics potential. Oncoscience 2018, 5, 181–195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, S.P.; Lin, Z.X.; Jiang, X.Y.; Yu, X.Y. Exosomal cargo-loading and synthetic exosome-mimics as potential therapeutic tools. Acta Pharmacol. Sin. 2018, 39, 542–551. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanz-Rubio, D.; Martin-Burriel, I.; Gil, A.; Cubero, P.; Forner, M.; Khalyfa, A.; Marin, J.M. Stability of Circulating Exosomal miRNAs in Healthy Subjects. Sci. Rep. 2018, 8, 10306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, B.; Xu, K.; Zheng, X.; Chen, T.; Wang, J.; Song, Y.; Shao, Y.; Zheng, S. Application of exosomes as liquid biopsy in clinical diagnosis. Signal Transduct. Target Ther. 2020, 5, 144. [Google Scholar] [CrossRef]
- Stansfield, J.C.; Rusay, M.; Shan, R.; Kelton, C.; Gaykalova, D.A.; Fertig, E.J.; Califano, J.A.; Ochs, M.F. Toward Signaling-Driven Biomarkers Immune to Normal Tissue Contamination. Cancer Inform. 2016, 15, 15–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Avissar, M.; Christensen, B.C.; Kelsey, K.T.; Marsit, C.J. MicroRNA expression ratio is predictive of head and neck squamous cell carcinoma. Clin. Cancer Res. 2009, 15, 2850–2855. [Google Scholar] [CrossRef] [Green Version]
- Christensen, B.C.; Moyer, B.J.; Avissar, M.; Ouellet, L.G.; Plaza, S.L.; McClean, M.D.; Marsit, C.J.; Kelsey, K.T. A let-7 microRNA-binding site polymorphism in the KRAS 3’ UTR is associated with reduced survival in oral cancers. Carcinogenesis 2009, 30, 1003–1007. [Google Scholar] [CrossRef] [Green Version]
- Peng, H.Y.; Hsiao, J.R.; Chou, S.T.; Hsu, Y.M.; Wu, G.H.; Shieh, Y.S.; Shiah, S.G. MiR-944/CISH mediated inflammation via STAT3 is involved in oral cancer malignance by cigarette smoking. Neoplasia 2020, 22, 554–565. [Google Scholar] [CrossRef]
- Mittelbrunn, M.; Gutierrez-Vazquez, C.; Villarroya-Beltri, C.; Gonzalez, S.; Sanchez-Cabo, F.; Gonzalez, M.A.; Bernad, A.; Sanchez-Madrid, F. Unidirectional transfer of microRNA-loaded exosomes from T cells to antigen-presenting cells. Nat. Commun. 2011, 2, 282. [Google Scholar] [CrossRef] [Green Version]
- Van Balkom, B.W.; Eisele, A.S.; Pegtel, D.M.; Bervoets, S.; Verhaar, M.C. Quantitative and qualitative analysis of small RNAs in human endothelial cells and exosomes provides insights into localized RNA processing, degradation and sorting. J. Extracell. Vesicles 2015, 4, 26760. [Google Scholar] [CrossRef]
- Keerthikumar, S.; Chisanga, D.; Ariyaratne, D.; Al Saffar, H.; Anand, S.; Zhao, K.; Samuel, M.; Pathan, M.; Jois, M.; Chilamkurti, N.; et al. ExoCarta: A Web-Based Compendium of Exosomal Cargo. J. Mol. Biol. 2016, 428, 688–692. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taylor, D.D.; Gercel-Taylor, C. MicroRNA signatures of tumor-derived exosomes as diagnostic biomarkers of ovarian cancer. Gynecol. Oncol. 2008, 110, 13–21. [Google Scholar] [CrossRef] [PubMed]
- Kong, Q.; Guo, X.; Guo, Z.; Su, T. Urinary Exosome miR-424 and miR-218 as Biomarkers for Type 1 Diabetes in Children. Clin. Lab. 2019, 65. [Google Scholar] [CrossRef]
- Xie, L.; Zhao, H.; Wang, Y.; Chen, Z. Exosomal shuttled miR-424-5p from ischemic preconditioned microglia mediates cerebral endothelial cell injury through negatively regulation of FGF2/STAT3 pathway. Exp. Neurol. 2020, 333, 113411. [Google Scholar] [CrossRef]
- Bovy, N.; Blomme, B.; Freres, P.; Dederen, S.; Nivelles, O.; Lion, M.; Carnet, O.; Martial, J.A.; Noel, A.; Thiry, M.; et al. Endothelial exosomes contribute to the antitumor response during breast cancer neoadjuvant chemotherapy via microRNA transfer. Oncotarget 2015, 6, 10253–10266. [Google Scholar] [CrossRef] [Green Version]
- Buhr, C.A.; Matteucci, M. 2’modified Nucleoside and Nucleotide Compounds. U.S. Patent 5,466,786, 14 November 1995. [Google Scholar]
- Kurreck, J. Antisense technologies. Improvement through novel chemical modifications. Eur. J. Biochem. 2003, 270, 1628–1644. [Google Scholar] [CrossRef]
- Roberts, T.C.; Langer, R.; Wood, M.J.A. Advances in oligonucleotide drug delivery. Nat. Rev. Drug Discov. 2020, 19, 673–694. [Google Scholar] [CrossRef] [PubMed]
- Khan, S.A.; Naz, A.; Qamar Masood, M.; Shah, R. Meta-Analysis of Inclisiran for the Treatment of Hypercholesterolemia. Am. J. Cardiol. 2020, 134, 69–73. [Google Scholar] [CrossRef]
- Ray, K.K.; Wright, R.S.; Kallend, D.; Koenig, W.; Leiter, L.A.; Raal, F.J.; Bisch, J.A.; Richardson, T.; Jaros, M.; Wijngaard, P.L.J.; et al. Two Phase 3 Trials of Inclisiran in Patients with Elevated LDL Cholesterol. N. Engl. J. Med. 2020, 382, 1507–1519. [Google Scholar] [CrossRef] [PubMed]
- Dyrbus, K.; Gasior, M.; Penson, P.; Ray, K.K.; Banach, M. Inclisiran-New hope in the management of lipid disorders? J. Clin. Lipidol. 2020, 14, 16–27. [Google Scholar] [CrossRef]
- Nishikido, T.; Ray, K.K. Inclisiran for the treatment of dyslipidemia. Expert Opin. Investig. Drugs 2018, 27, 287–294. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
microRNA biogenesis and their biological functions.

Figure 2.
The regulation mechanisms of microRNA clusters in carcinogenesis. (A,B) Expression of microRNA cluster is regulated by genetic events. miR-17/92 cluster is stimulated by chromosome t(8;12) translation (A) or transcriptional activation by transcription factor MYC (B). (C) The epigenetic regulation of miR-379/656 cluster upon DNA hypermethylation on MEG3-DMR.
Figure 2.
The regulation mechanisms of microRNA clusters in carcinogenesis. (A,B) Expression of microRNA cluster is regulated by genetic events. miR-17/92 cluster is stimulated by chromosome t(8;12) translation (A) or transcriptional activation by transcription factor MYC (B). (C) The epigenetic regulation of miR-379/656 cluster upon DNA hypermethylation on MEG3-DMR.


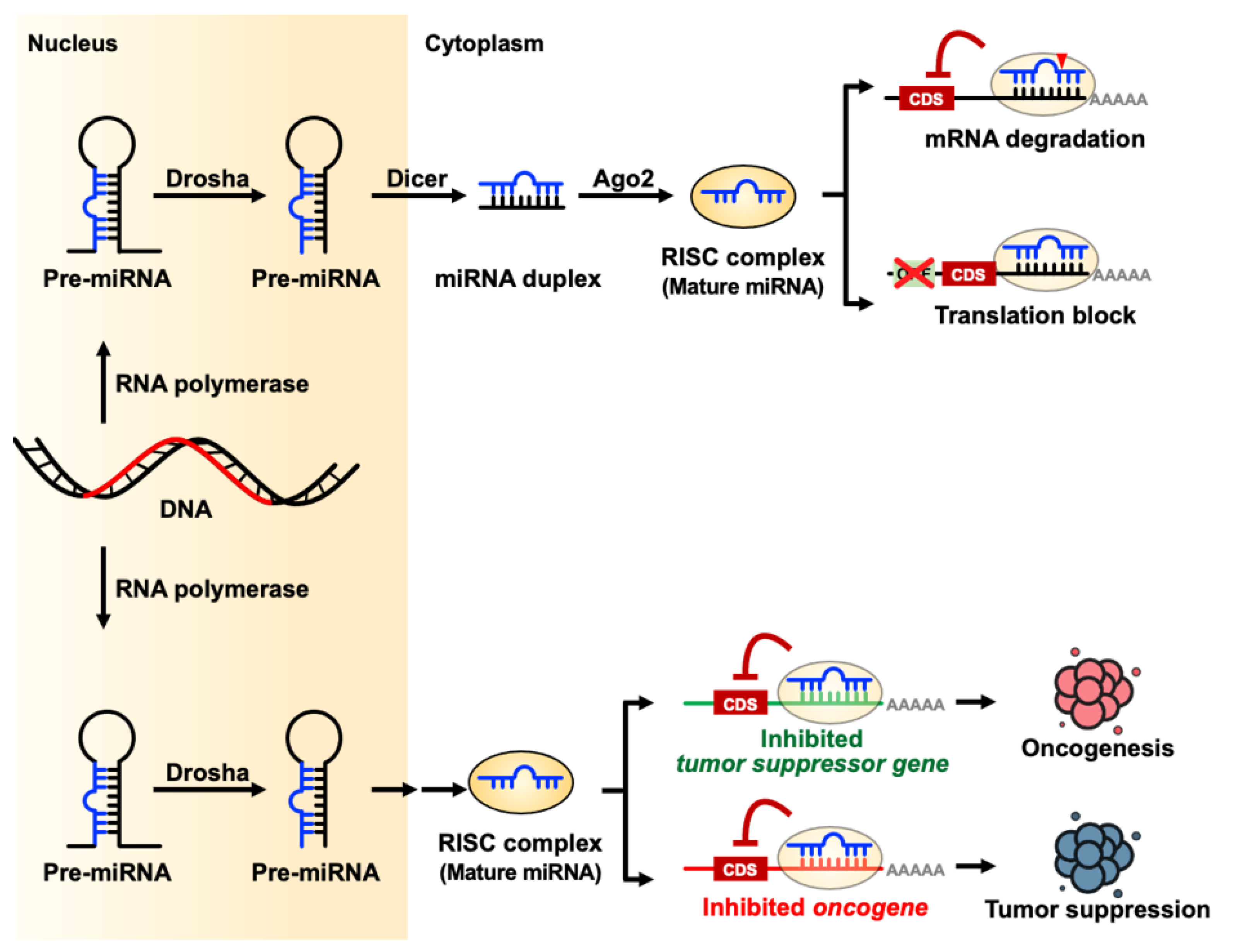{kind=link}
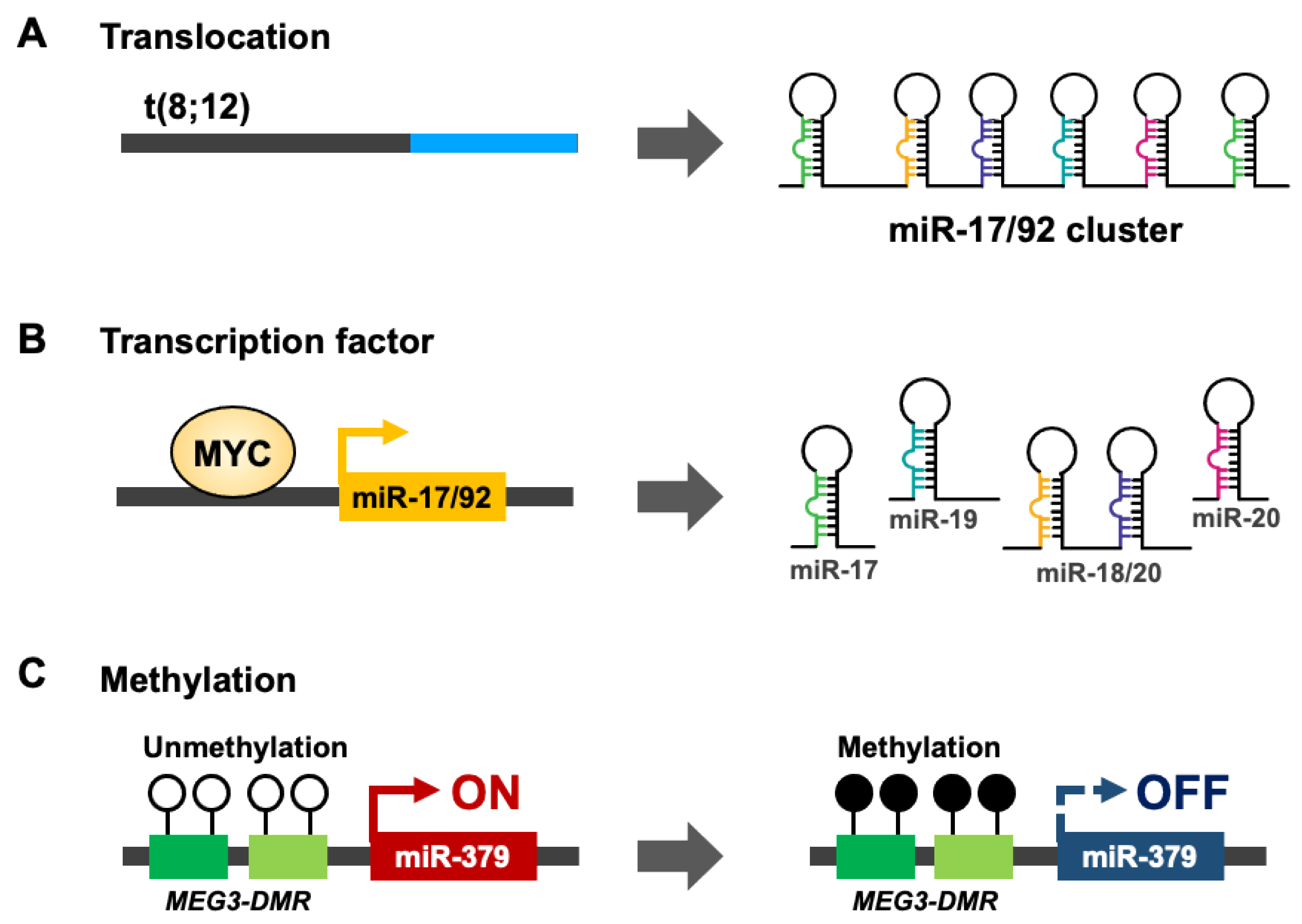{kind=link}
Table 1.
The physiological functions of microRNA clusters.
| miRNA Function | 1/133a | 106a/363 | 106b/25 | 143/145 | 15a/16-1 | 17/92 | 181a/181b | 183/182 | 193/365a | 199a/241 | 200b/429 | 206/133b/ | 212/132 | 221/222 | 23a/24-2 | 27a/27b | 29b/29a | 301b/130b | 302b/367 | 34b/34c | 371a/373 | 379/656 | 493/136 | 424/450b | 508/513a | 512/519 |
| Differentiation and development | ◎ | ◎ | ◎ | ◎ | ◎ | ◎ | ◎ | ◎ | ◎ | ◎ | ◎ | ◎ | ||||||||||||||
| Secondary messengers | ◎ | ◎ | ◎ | ◎ | ◎ | ◎ | ◎ | ◎ | ◎ | |||||||||||||||||
| microRNA and stress | ◎ | ◎ | ◎ | ◎ | ◎ | ◎ | ||||||||||||||||||||
| Immunity | ◎ | ◎ | ◎ | ◎ | ||||||||||||||||||||||
| Feedback loops | ◎ | ◎ | ◎ | |||||||||||||||||||||||
| Cell junctions and adhesions | ◎ | ◎ | ||||||||||||||||||||||||
| Mammalian reproductions | ◎ | ◎ | ◎ | ◎ | ◎ | |||||||||||||||||||||
| Cardiac development and diseases | ◎ | ◎ | ◎ | ◎ | ||||||||||||||||||||||
| Cellular reprogramming | ◎ | ◎ | ◎ | ◎ | ||||||||||||||||||||||
| Cell to cell communication | ◎ | ◎ | ◎ | ◎ | ||||||||||||||||||||||
| DNA repair | ◎ | ◎ | ◎ | |||||||||||||||||||||||
| Viral infection | ◎ | ◎ | ||||||||||||||||||||||||
| Nervous system regulation | ◎ | ◎ | ◎ | ◎ | ||||||||||||||||||||||
| Metabolism | ◎ | ◎ | ◎ | ◎ | ◎ | ◎ | ||||||||||||||||||||
| Organellar biogenesis and function | ◎ | ◎ | ◎ | ◎ | ◎ | |||||||||||||||||||||
| Mesenchymal to epithelial transition | ◎ | ◎ | ◎ |
Table 2.
Aberrant microRNA in HNSCC.
| microRNA | Function | Targets | Effects on Tumor | Reference |
|---|---|---|---|---|
| miR-1 | TSmiR | TAGLN2, PNP | Proliferation − Apoptosis + | [136] |
| miR-1 | TSmiR | TAGLN2 | Proliferation − Apoptosis + Cell cycle arrest + | [137] |
| miR-1 | TSmiR | EGFR, c-Met | Proliferation − Migration − Invasion − | [138] |
| miR-1 | TSmiR | Slug | Migration − Invasion − Stemness − | [139] |
| miR-1 | TSmiR | ET-1 | Angiogenesis − | [140] |
| miR-7 | TSmiR | IGF1R | Proliferation − Apoptosis + Cell cycle arrest + | [141] |
| miR-9 | TSmiR | PTEN | Proliferation − | [142] |
| CXCR4 | Proliferation − Apoptosis + Cell cycle arrest + Invasion − | [143] | ||
| miR-100 | TSmiR | IGF1R, mTOR | Proliferation − Apoptosis + Migration − | [144] |
| Akt1 | Proliferation − Apoptosis + Migration − | [145] | ||
| miR-101 | TSmiR | EZH2, rap1GAP | Proliferation − Invasion − | [146] |
| EZH2 | Migration − Invasion − EMT − | [147] | ||
| miR-101 | TSmiR | ITGA3 | Migration − Invasion − Angiogenesis − | [148] |
| miR-101-3p | TSmiR | Pim-1 | Proliferation − Apoptosis + Invasion − | [149] |
| miR-104-5p | TSmiR | PAK4 | Proliferation − Apoptosis + Cell cycle arrest + | [150] |
| miR-104-5p | TSmiR | Survivin | Proliferation − Apoptosis + Invasion − | [151] |
| miR-10b | TSmiR | - | Proliferation − Cell cycle arrest + | [152] |
| miR-10b | OncomiR | - | Migration + Invasion + | [153] |
| miR-10b | OncomiR | - | Migration + Invasion + | [154] |
| miR-1-3p | TSmiR | DKK1 | Proliferation − Apoptosis + Cell cycle arrest + Migration − Invasion − | [155] |
| miR-125b | TSmiR | PRXL2A | Oxidative stress + Drug sensitivity + | [156] |
| miR-138 | TSmiR | GNAI2 | Proliferation − Apoptosis + | [157] |
| miR-140-5p | TSmiR | ADAM10 | Migration − Invasion − | [158] |
| miR-140-5p | TSmiR | LAMC1, HDAC7, PAX6 | Migration − Invasion − | [159] |
| miR-146a | TSmiR | - | Proliferation − Migration − | [132] |
| miR-155 | TSmiR | - | Proliferation − Migration − | [132] |
| miR-155 | OncomiR | - | Association with tobacco chewing | [160] |
| miR-155 | OncomiR | CDC73 | Proliferation + Migration + | [129,130,131] |
| miR-181a | TSmiR | K-ras | Proliferation − | [161] |
| miR-181a | TSmiR | MAP2K1, MAPK1, Snai2 | Proliferation − Migration − Invasion − | [162] |
| miR-181a | TSmiR | Twist1 | Drug sensitivity + EMT − Metastasis − | [163] |
| miR-181a/b | OncomiR | - | Migration − Invasion − | [164] |
| miR-183 | OncomiR | - | Prognostic biomarker | [165] |
| miR-196a | OncomiR | AXIN | Proliferation − Cell cycle arrest + | [152] |
| miR-21 | OncomiR | - | Prognostic biomarker | [165] |
| miR-21 | OncomiR | TPM1, PTEN, CDK2AP1, HIF-1A, HIF-2A | Association with tumor progression | [166] |
| miR-203 | OncomiR | - | As diagnostic marker of metastasis | [167] |
| miR-203 | TSmiR | - | Invasion − EMT − | [168] |
| miR-204 | TSmiR | BRD4 | Proliferation − Apoptosis + Cell cycle arrest + | [169] |
| TSmiR | Sox4, Slug | Stemness − EMT − Metastasis − | [170] | |
| TSmiR | CDC2 | Invasion − Metastasis − | [171] | |
| miR-204-5p | TSmiR | CXCR4 | Proliferation − Cell cycle arrest + Migration − Invasion − | [172] |
| miR-205 | OncomiR | - | As diagnostic marker of metastasis | [167] |
| miR-206 | TSmiR | EGFR, c-Met | Proliferation − Migration − Invasion − | [138] |
| miR-221 | OncomiR | p27, p57 | Proliferation + | [173] |
| PTEN | Proliferation + Invasion + Apoptosis − | [174] | ||
| miR-222 | OncomiR | p27, p57 | Proliferation + | [173] |
| PTEN | Proliferation + Invasion + Apoptosis − | [174] | ||
| miR-222 | TSmiR | PUMA | Proliferation − Invasion − Drug sensitivity + | [175] |
| miR-222 | TSmiR | MMP1, SOD2 | Invasion − | [176] |
| miR-223 | OncomiR | - | Proliferation − Apoptosis + | [177] |
| miR-223 | OncomiR | FBXW7 | Proliferation + Migration + | [178] |
| miR-24 | OncomiR | DND1 | Proliferation + Apoptosis − | [179] |
| miR-25 | TSmiR | NEDD9 | Proliferation − Apoptosis + | [180] |
| miR-26a | TSmiR | DNMT3B | Proliferation − Cell cycle arrest + Apoptosis + | [181] |
| miR-26a/b | TSmiR | PAK1 | Cell cycle arrest + Migration − Invasion − Apoptosis + Glycolysis − | [182] |
| miR-26a/b | TSmiR | TMEM184B | Migration − Invasion − | [183] |
| miR-31 | OncomiR | - | As diagnostic biomarker | [126,127] |
| SIRT3 | Migration + Invasion + | [64] | ||
| FIH | Migration + Invasion + | [128] | ||
| ARID1A | Oncogenesis + Stemness + | [184] | ||
| miR-34a | TSmiR | MDM4, SIRT1 | Downregulated in tumors | [185] |
| miR-34a | TSmiR | BCL-2 | Migration − Invasion − | [186] |
| miR-372 | OncomiR | ZBTB7A | Drug sensitivity − | [187] |
| miR-376c | TSmiR | RUNX2 | Migration − Invasion − Lymphatic metastasis − | [63] |
| miR-379 | TSmiR | DNMT3B | Proliferation + | [133] |
| miR-424 | OncomiR | SOCS2 | Migration + Invasion + | [188] |
| miR-450a | OncomiR | TMEM182 | Migration + Invasion + | [124] |
| miR-455 | OncomiR | UBE2B | Proliferation + | [125] |
| miR-486 | TSmiR | DDR1 | Proliferation − Apoptosis + | [134] |
| miR-503 | OncomiR | Smad7 | Proliferation + Migration + Invasion + | [189] |
| miR-494 | TSmiR | HOXA10 | Proliferation − | [190] |
| miR-494-3p | OncomiR | Sox7 | Proliferation + Migration + Invasion + | [191] |
| miR-99 | TSmiR | AKT1 | Proliferation − Apoptosis + Migration − | [145] |
| miR-99-3b | TSmiR | GSK3b | Proliferation − | [192] |
| miR-99a | TSmiR | mTOR | Proliferation − | [193] |
| MTMR3 | Migration − Invasion − | [194] | ||
| miR-99a | TSmiR | IGF1R | Migration − Invasion − | [135] |
| let-7b | TSmiR | Dicer | Proliferation − | [195] |
Both (−) and (+) mean microRNAs show inhibitory or stimulative effect on head and neck cancer, respectively. Abbreviations: ADAM10, ADAM metallopeptidase domain 10; AKT1, AKT serine/threonine kinase 1; ARID1A, AT-rich Interaction domain 1A; AXIN, Axis inhibition protein 2; BCL-2, B-cell lymphoma 2; BRD4, Bromodomain-containing protein 4; CDC42/73, Cell division cycle 42/73; CDK2AP1, Cyclin-dependent kinase 2-associated protein 1; c-MET, Hepatocyte growth factor receptor; CXCR4, C-X-C chemokine receptor type 4; DDR1, Discoidin domain receptor-1; DKK1, Dickkopf wnt signaling pathway inhibitor 1; DND1, RNA-binding protein dead end 1; DNMT3B, DNA (cytosine-5-)-methyltransferase 3 beta; EGFR, Epidermal growth factor receptor; ET-1, Endothelin-1; EZH2, Enhancer of zeste 2 polycomb repressive complex 2; FBXW7, F-Box and wd repeat domain containing 7; FIH, Factor-inhibiting hypoxia-inducible factor; GNAI2, G protein subunit alpha I2; GSK3b, Glycogen synthase kinase 3 beta; HDAC7, Histone deacetylase 7; HIF-1/2A, Hypoxia inducible factor 1/2 alpha; HOXA10, Homeobox A10; IGF1R, Insulin-like growth factor 1 receptor; ITGA3, Integrin subunit alpha 3; K-ras, KRAS proto-oncogene; LAMC1, Laminin subunit gamma 1; MAP2K1, Mitogen-activated protein kinase kinase 1; MDM4, Mouse double minute 4 homolog; MMP1, Matrix metallopeptidase 1; MTMR3, Myotubularin-related protein 3; mTOR, Mammalian target of rapamycin; NEDD9, Neural precursor cell expressed, developmentally down-regulated 9; p27, Cyclin dependent kinase inhibitor 1b; p57, Cyclin dependent kinase inhibitor 1c; PAK1/4, p21-activated kinase 1/4; PAX6, Paired type homeobox 6; Pim-1, Serine/threonine-protein kinase Pim-1; PNP, Purine nucleoside phosphorylase; PRXL2A, Peroxiredoxin like 2a; PTEN, Phosphatase and tensin homolog; PUMA, BCL2 binding component 3; rap1GAP, RAP1 GTPase activating protein; RUNX2, RUNX Family transcription factor 2; SIRT1, Sirtuin 1; SIRT3, Silent information regulator 3; Slug, SNAI2, snail family transcriptional repressor; Smad7, SMAD family member 7; Snai2, Snail family transcriptional repressor 2; SOCS2, Suppressor of cytokine signaling 2; SOD2, Superoxide dismutase 2; Sox4/7, SRY-related HMG-box 4/7; Survivin, BIRC5, baculoviral IAP repeat containing 5; TAGLN2, Transgelin 2; TMEM182/184B, Transmembrane protein 182/184B; TPM1, Tropomyosin 1; Twist1, Twist family BHLH transcription factor 1; UBE2B, Ubiquitin conjugating enzyme E2 B; ZBTB7A, Zinc finger and BTB domain containing 7A.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Li, L.-J.; Chang, W.-M.; Hsiao, M. Aberrant Expression of microRNA Clusters in Head and Neck Cancer Development and Progression: Current and Future Translational Impacts. Pharmaceuticals 2021, 14, 194. https://doi.org/10.3390/ph14030194
AMA Style
Li L-J, Chang W-M, Hsiao M. Aberrant Expression of microRNA Clusters in Head and Neck Cancer Development and Progression: Current and Future Translational Impacts. Pharmaceuticals. 2021; 14(3):194. https://doi.org/10.3390/ph14030194
Chicago/Turabian StyleLi, Li-Jie, Wei-Min Chang, and Michael Hsiao. 2021. "Aberrant Expression of microRNA Clusters in Head and Neck Cancer Development and Progression: Current and Future Translational Impacts" Pharmaceuticals 14, no. 3: 194. https://doi.org/10.3390/ph14030194
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.