
Zingerone Targets Status Epilepticus by Blocking Hippocampal Neurodegeneration via Regulation of Redox Imbalance, Inflammation and Apoptosis

 ,
,
Abstract
:1. Introduction
2. Results
2.1. Effect of Zingerone and Sodium Valproate on Seizure Activity against LiCl-and-Pilocarpine-induced Epilepsy
2.1.1. Effect on Latency to Start of Seizures
2.1.2. Effect on Percentage Convulsion
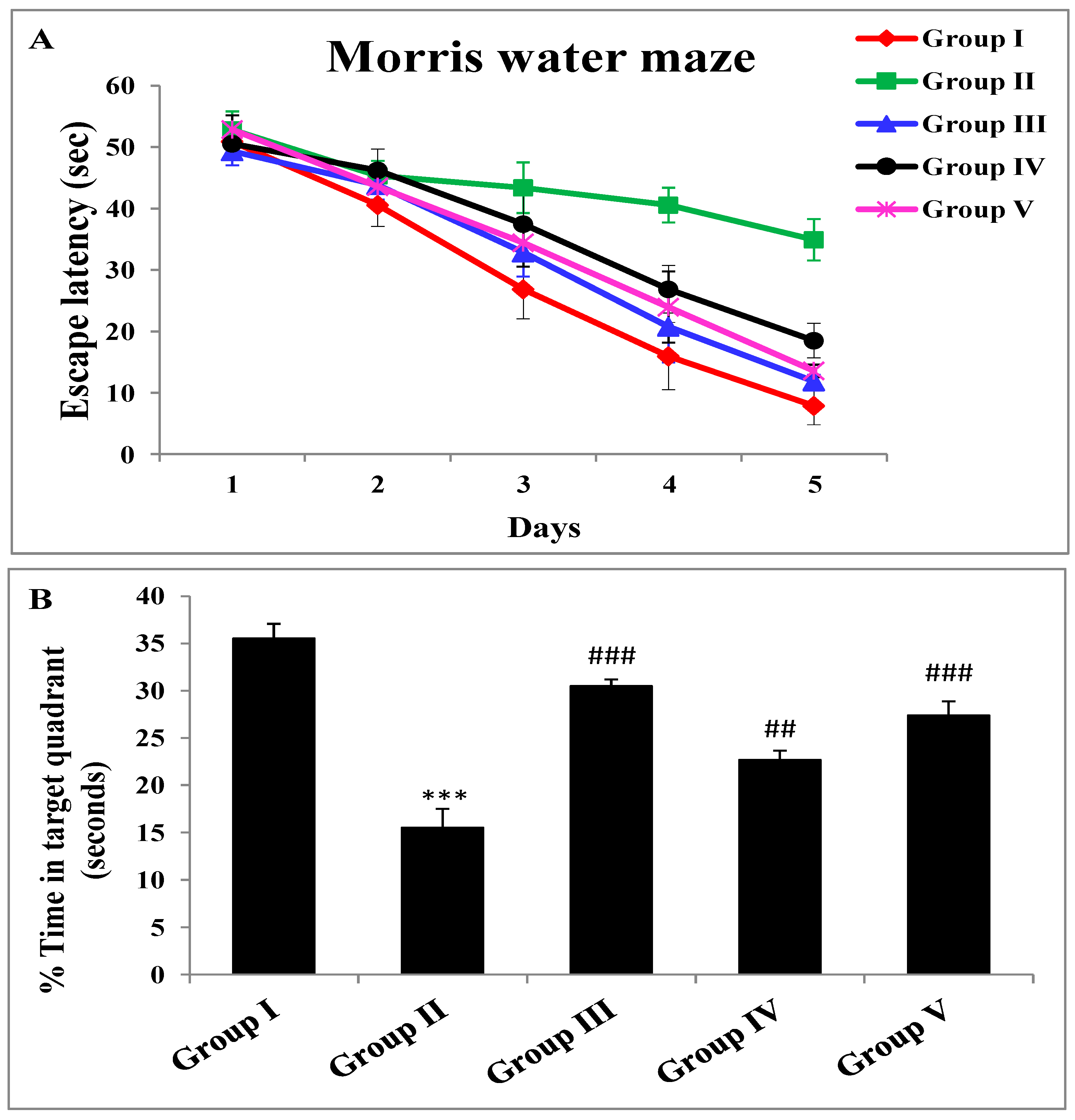
2.2. Effect of Zingerone and Sodium Valproate on Escape Latency and Probe Trial of Morris Water Maze
2.3. Zingerone and Sodium Valproate Effects on Different Biochemical Parameters against LiCl-and-pilocarpine-induced Epilepsy
2.3.1. Lipid Peroxidation (MDA)
2.3.2. Superoxide Dismutase Activity (SOD)
2.3.3. Catalase Activity (CAT)
2.3.4. Glutathione Reductase Levels (GR)
2.3.5. Reduced Glutathione Levels (GSH)
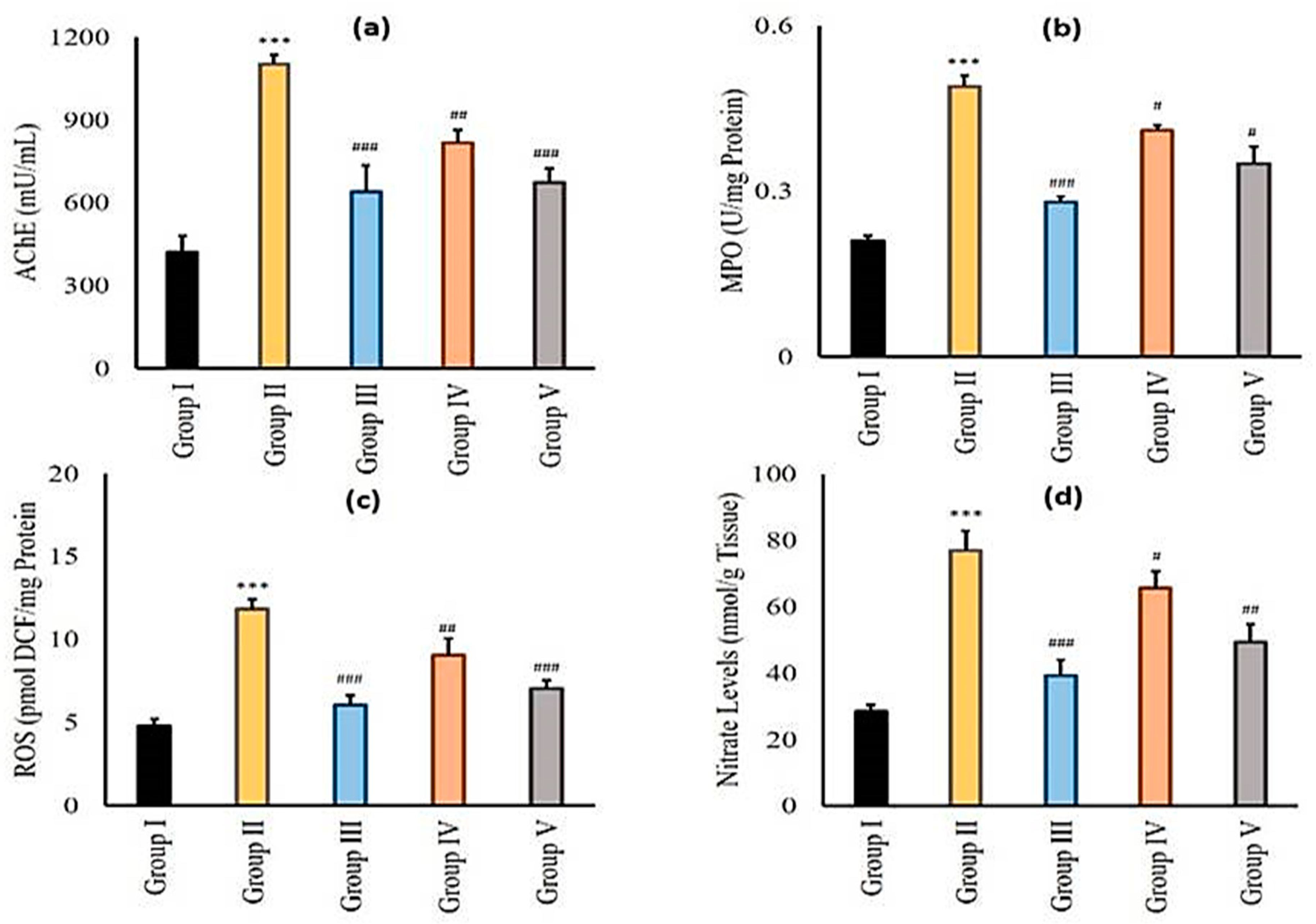
2.3.6. Effect of Zingerone and Sodium Valproate on Acetylcholine Esterase (AChE), Myeloperoxidase (MPO), Reactive Oxygen Species (ROS), and Nitrite (NO) Levels in LiCl-and-pilocarpine-induced Epilepsy
2.3.7. Effect of Zingerone and Sodium Valproate on Inflammatory Mediators (NFκB, TNF-α, IL-6 and IL-1β) in LiCl-and-pilocarpine-induced Epilepsy
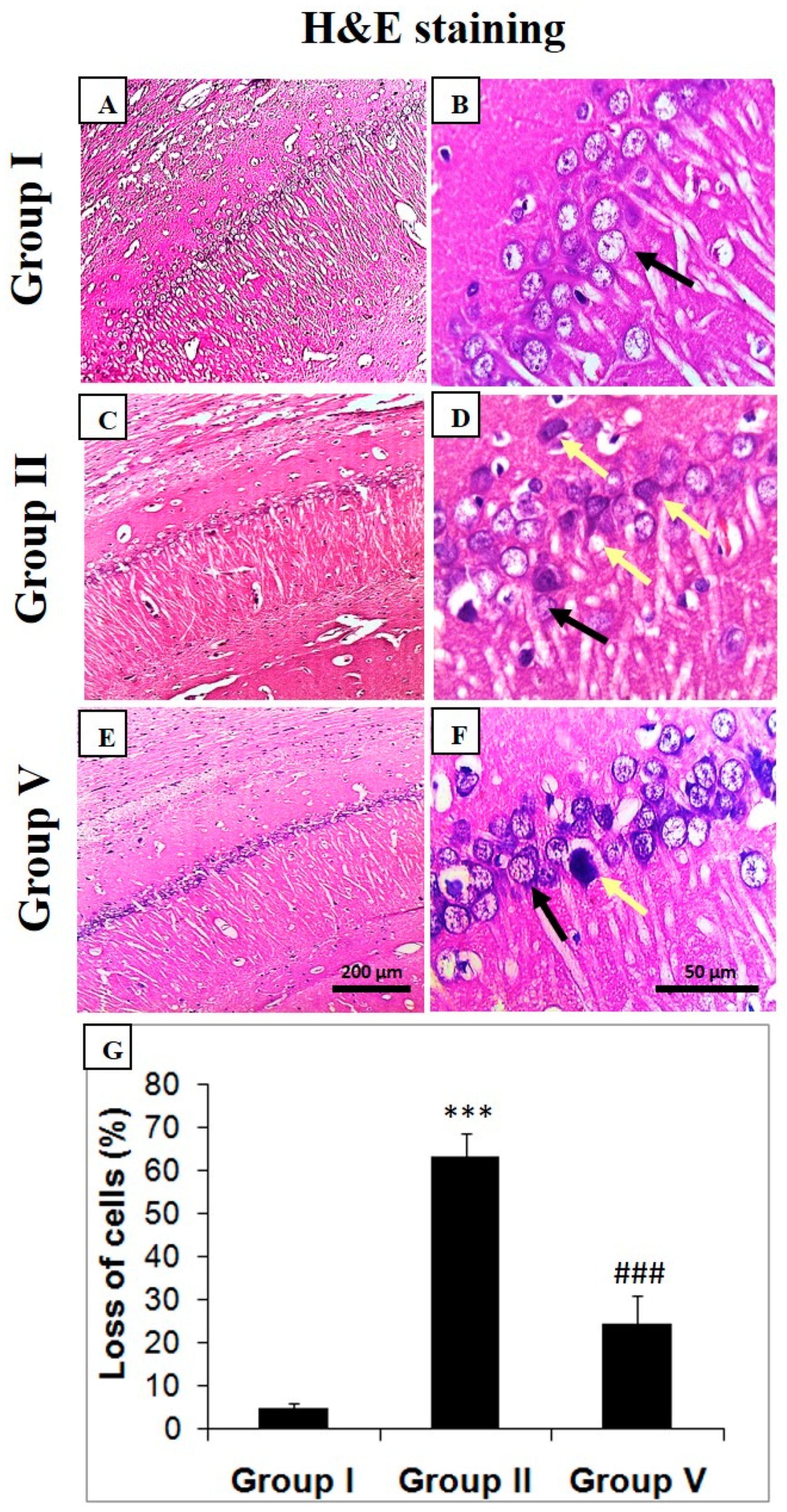
2.4. Histopathological Analysis of Zingerone and Sodium Valproate Treatment in LiCl-and-pilocarpine-induced Epilepsy
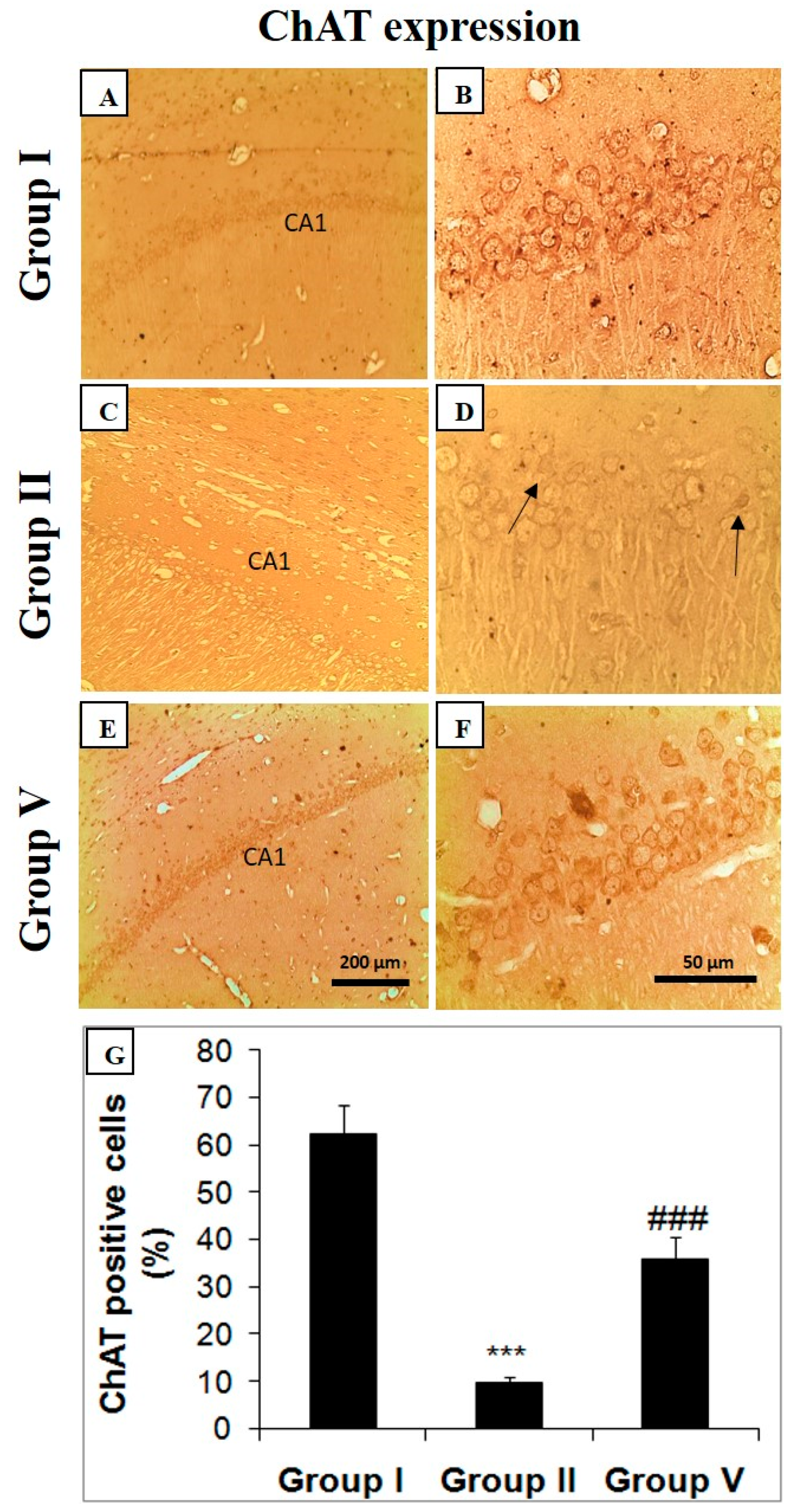
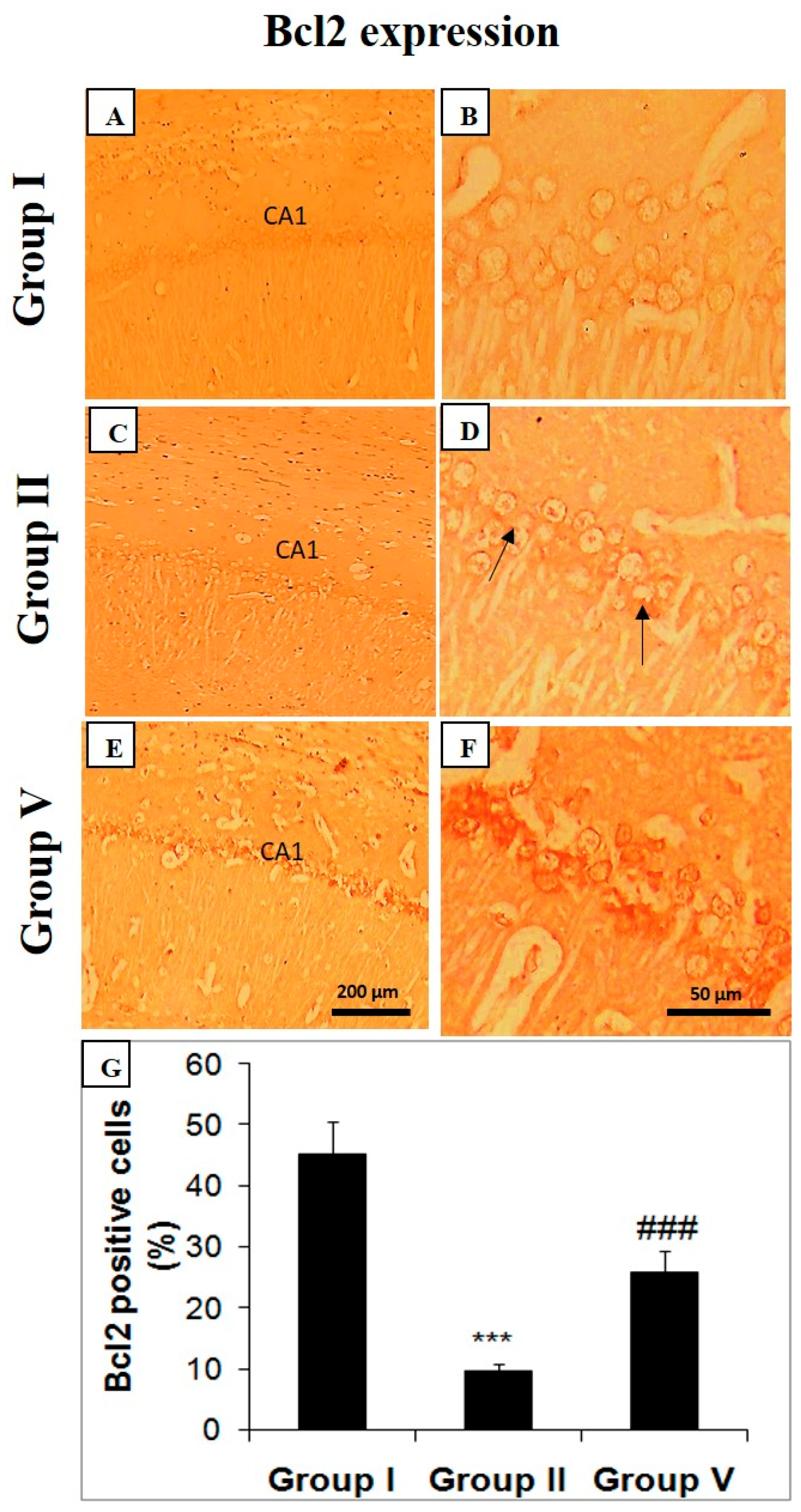
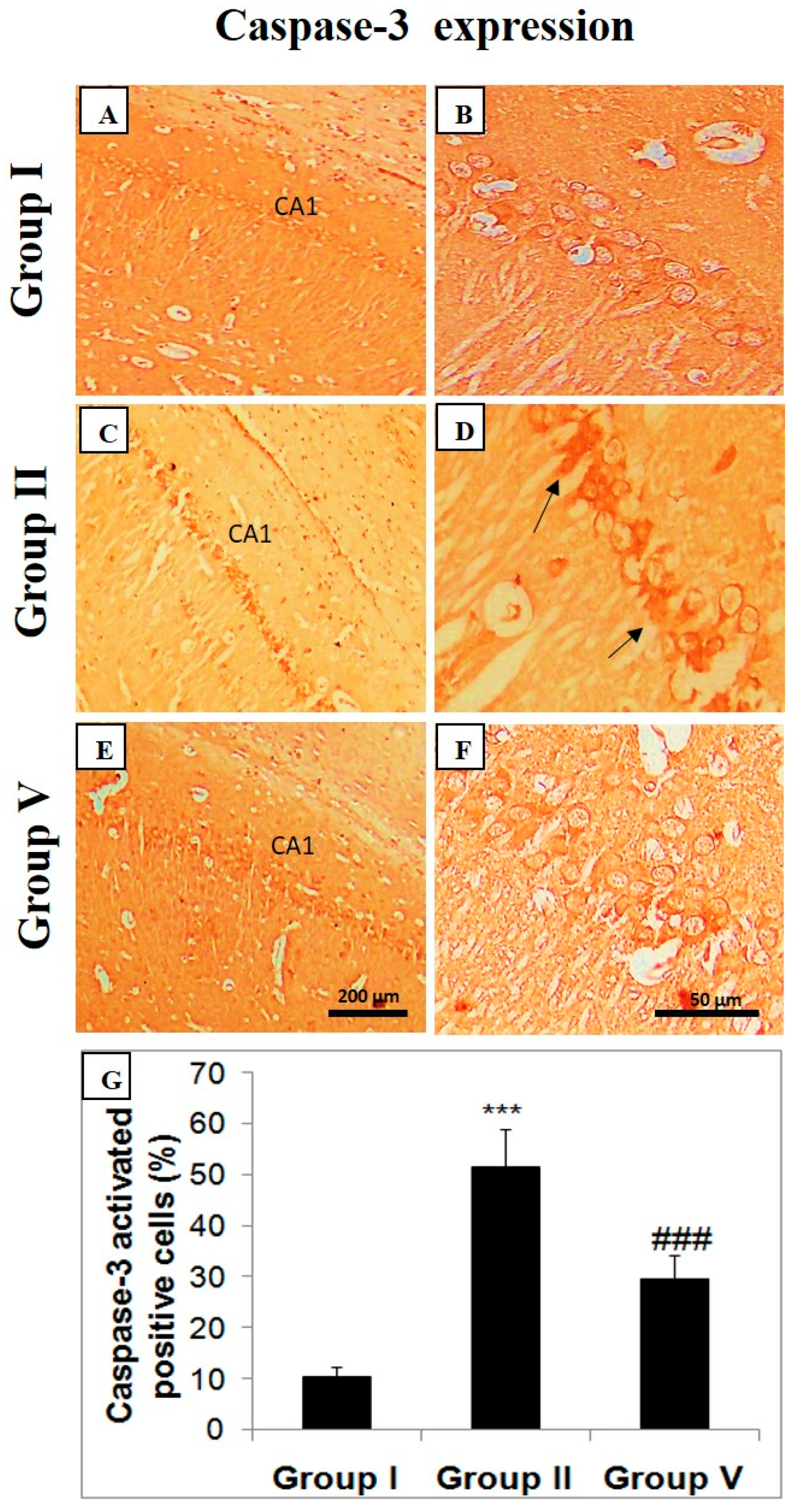
2.5. Effect of Zingerone on Choline Acetyl Transferase (ChAT), Bcl-2, and Caspase Expression
3. Discussion
4. Materials and Methods
4.1. Chemicals
4.2. Animal Study
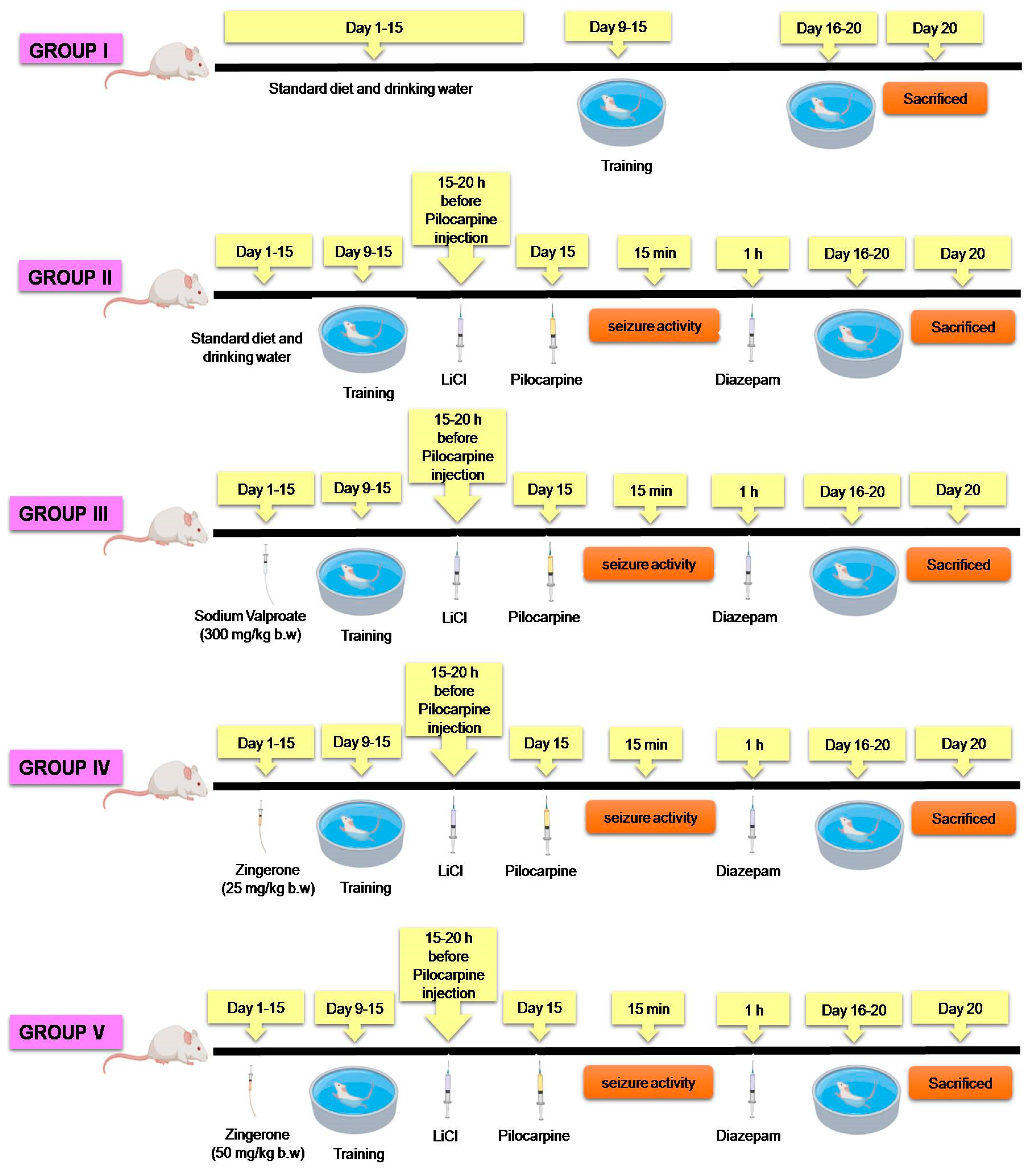
4.3. Preparation of Drugs and Treatment Protocol
Experimental Regimen
4.4. Morris Water Maze (MWM)
4.5. Tissue Homogenization
4.5.1. Assessment of Lipid Peroxidation (LPO)
4.5.2. Estimation of Superoxide Dismutase Activity (SOD)
4.5.3. Estimation of Catalase Activity (CAT)
4.5.4. Estimation of Glutathione Reductase [GR] Activity
4.5.5. Estimation of Reduced Glutathione (GSH)
4.5.6. Assay for Activity of AChE
4.5.7. Assay for Myeloperoxidase Activity
4.5.8. Reactive Oxygen Species (ROS) Assessment
4.5.9. Assay for Nitric Oxide (NO)
4.5.10. Estimation of Nuclear Factor kappa B (NFκ-B), Tumor Necrosis Factor alpha (TNF-α), Interleukin 6 (IL-6) and Interleukin 1 beta (IL-1β)
4.5.11. Assay for Protein Measurement
4.6. Histopathology and Counting of Neuronal Loss
4.7. Immunohistochemical Staining of ChAT, Bcl-2, and Caspase-3
4.8. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- World Health Organization (WHO). 2019. Available online: https://www.who.int/health-topics/epilepsy#tab=tab_1 (accessed on 4 July 2019).
- Stafstrom, C.E.; Carmant, L. Seizures and epilepsy: An overview for neuroscientists. Cold Spring Harb. Perspect. Med. 2015, 5, a022426. [Google Scholar] [CrossRef]
- Trinka, E.; Höfler, J.; Leitinger, M.; Brigo, F. Pharmacotherapy for Status Epilepticus. Drugs 2015, 75, 1499–1521. [Google Scholar] [CrossRef] [Green Version]
- Zhao, F.; Kang, H.; You, L.; Rastogi, P.; Venkatesh, D.; Chandra, M. Neuropsychological deficits in temporal lobe epilepsy: A comprehensive review. Ann. Indian Acad. Neurol. 2014, 17, 374–382. [Google Scholar] [CrossRef]
- Trinka, E.; Cock, H.; Hesdorffer, D.; Rossetti, A.O.; Scheffer, I.E.; Shinnar, S.; Shorvon, S.; Lowenstein, D.H. A definition and classification of status epilepticus--Report of the ILAE Task Force on Classification of Status Epilepticus. Epilepsia 2015, 56, 1515–1523. [Google Scholar] [CrossRef]
- Kinjo, E.R.; Higa, G.S.; Santos, B.A.; de Sousa, E.; Damico, M.V.; Walter, L.T.; Morya, E.; Valle, A.C.; Britto, L.R.; Kihara, A.H. Pilocarpine-induced seizures trigger differential regulation of microRNA-stability related genes in rat hippocampal neurons. Sci. Rep. 2016, 6, 20969. [Google Scholar] [CrossRef] [Green Version]
- Deng, X.; Wang, M.; Hu, S.; Feng, Y.; Shao, Y.; Xie, Y.; Wu, M.; Chen, Y.; Shi, X. The Neuroprotective Effect of Astaxanthin on Pilocarpine-Induced Status Epilepticus in Rats. Front. Cell. Neurosci. 2019, 13, 123. [Google Scholar] [CrossRef] [PubMed]
- Shin, E.J.; Jeong, J.H.; Chung, Y.H.; Kim, W.K.; Ko, K.H.; Bach, J.H.; Hong, J.S.; Yoneda, Y.; Kim, H.C. Role of oxidative stress in epileptic seizures. Neurochem. Int. 2011, 59, 122–137. [Google Scholar] [CrossRef] [Green Version]
- Wang, A.; Si, Z.; Li, X.; Lu, L.; Pan, Y.; Liu, J. FK506 Attenuated Pilocarpine-Induced Epilepsy by Reducing Inflammation in Rats. Front. Neurol. 2019, 10, 971. [Google Scholar] [CrossRef]
- Wang, Z.; Zhou, L.; An, D.; Xu, W.; Wu, C.; Sha, S.; Li, Y.; Zhu, Y.; Chen, A.; Du, Y.; et al. TRPV4-induced inflammatory response is involved in neuronal death in pilocarpine model of temporal lobe epilepsy in mice. Cell Death Dis. 2019, 10, 386. [Google Scholar] [CrossRef]
- Rehman, M.U.; Wali, A.F.; Ahmad, A.; Shakeel, S.; Rasool, S.; Ali, R.; Rashid, S.M.; Madkhali, H.; Ganaie, M.A.; Khan, R. Neuroprotective Strategies for Neurological Disorders by Natural Products: An update. Curr. Neuropharmacol. 2019, 17, 247–267. [Google Scholar] [CrossRef] [PubMed]
- Safhi, M.M. Nephroprotective Effect of Zingerone against CCl4-Induced Renal Toxicity in Swiss Albino Mice: Molecular Mechanism. Oxi. Med. Cell. Longev. 2018, 2474831. [Google Scholar] [CrossRef] [Green Version]
- Ganaie, M.A.; Al Saeedan, A.; Madhkali, H.; Jan, B.L.; Khatlani, T.; Sheikh, I.A.; Rehman, M.U.; Wani, K. Chemopreventive efficacy zingerone (4-[4-hydroxy-3-methylphenyl] butan-2-one in experimental colon carcinogenesis in Wistar rats. Environ. Toxicol. 2019, 34, 610–625. [Google Scholar] [CrossRef]
- Rehman, M.U.; Rashid, S.M.; Rasool, S.; Shakeel, S.; Ahmad, B.; Ahmad, S.B.; Madkhali, H.; Ganaie, M.A.; Majid, S.; Bhat, S.A. Zingerone (4-(4-hydroxy-3-methylphenyl)butan-2-one) ameliorates renal function via controlling oxidative burst and inflammation in experimental diabetic nephropathy. Arch. Physiol. Biochem. 2019, 125, 201–209. [Google Scholar] [CrossRef] [PubMed]
- Min, G.; Ku, S.K.; Lee, T.; Bae, J.S. Suppressive effects of zingerone on TGFBIp-mediated septic responses. Arch. Pharmacol. Res. 2018, 41, 276–287. [Google Scholar] [CrossRef]
- Kim, C.Y.; Seo, Y.; Lee, C.; Park, G.H.; Jang, J.H. Neuroprotective Effect and Molecular Mechanism of [6]-Gingerol against Scopolamine-Induced Amnesia in C57BL/6 Mice. Evid. Based Complement. Altern. Med. eCAM 2018, 8941564. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Safhi, M.M. Zingerone protects the tellurium toxicity in the brain mitochondria of rats. Metabolomics 2015, 5, 156. [Google Scholar] [CrossRef] [Green Version]
- Li, L.L.; Cui, Y.; Guo, X.H.; Ma, K.; Tian, P.; Feng, J.; Wang, J.M. Pharmacokinetics and Tissue Distribution of Gingerols and Shogaols from Ginger (Zingiber officinale Rosc.) in Rats by UPLC⁻Q-Exactive⁻HRMS. Molecules. 2019, 24, 512. [Google Scholar] [CrossRef] [Green Version]
- Kabuto, H.; Yamanushi, T.T. Effects of zingerone [4-(4-hydroxy-3-methoxyphenyl)-2-butanone] and eugenol [2-methoxy-4-(2-propenyl)phenol] on the pathological progress in the 6-hydroxydopamine-induced Parkinson’s disease mouse model. Neurochem. Res. 2011, 36, 2244–2249. [Google Scholar] [CrossRef]
- Vaibhav, K.; Shrivastava, P.; Tabassum, R.; Khan, A.; Javed, H.; Ahmed, M.E.; Islam, F.; Safhi, M.M.; Islam, F. Delayed administration of zingerone mitigates the behavioral and histological alteration via repression of oxidative stress and intrinsic programmed cell death in focal transient ischemic rats. Pharmacol. Biochem. Behavior. 2013, 113, 53–62. [Google Scholar] [CrossRef]
- Al Kury, L.T.; Mahgoub, M.; Howarth, F.C.; Oz, M. Natural Negative Allosteric Modulators of 5-HT₃ Receptors. Molecules 2018, 23, 3186. [Google Scholar] [CrossRef] [Green Version]
- Venkatanarayana, N. Evaluation of anticonvulsant activity of ethanolic extract of Zingiber officinale in Swiss albino rats. J. Chem. Pharm. Res. 2013, 5, 60–64. [Google Scholar]
- Walker, M.C. Pathophysiology of status epilepticus. Neurosci. Lett. 2018, 667, 84–91. [Google Scholar] [CrossRef]
- Trinka, E.; Brigo, F.; Shorvon, S. Recent advances in status epilepticus. Curr. Opin. Neurol. 2016, 29, 189–198. [Google Scholar] [CrossRef]
- Soliman, A.F.; Anees, L.M.; Ibrahim, D.M. Cardioprotective effect of zingerone against oxidative stress, inflammation, and apoptosis induced by cisplatin or gamma radiation in rats. Naunyn Schmiedeberg Arch. Pharmacol. 2018, 39, 819–832. [Google Scholar] [CrossRef]
- Phelan, K.D.; Shwe, U.T.; Williams, D.K.; Greenfield, L.J.; Zheng, F. Pilocarpine-induced status epilepticus in mice: A comparison of spectral analysis of electroencephalogram and behavioral grading using the Racine scale. Epilepsy Res. 2015, 117, 90–96. [Google Scholar] [CrossRef] [Green Version]
- Greenfield, L.J., Jr. Molecular mechanisms of antiseizure drug activity at GABAA receptors. Seizure 2013, 22, 589–600. [Google Scholar] [CrossRef] [Green Version]
- Yue, H.Y.; Jiang, C.Y.; Fujita, T.; Kumamoto, E. Zingerone enhances glutamatergic spontaneous excitatory transmission by activating TRPA1 but not TRPV1 channels in the adult rat substantia gelatinosa. J. Neurophysiol. 2013, 110, 658–671. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castle, N.A. Differential inhibition of potassium currents in rat ventricular myocytes by capsaicin. Cardiovasc. Res. 1992, 26, 1137–1144. [Google Scholar] [CrossRef] [PubMed]
- Erdélyi, L. Guaiacol and vanilloid compounds modulate the A-type potassium currents in molluscan neurons. Acta Biol. Hung. 1999, 50, 65–79. [Google Scholar] [PubMed]
- Kim, J.N.; Kim, H.J.; Kim, I.; Kim, Y.T.; Kim, B.J. The Mechanism of Action of Zingerone in the Pacemaker Potentials of Interstitial Cells of Cajal Isolated from Murine Small Intestine. Cell. Physiol. Biochem. Int. J. Exp. Cell. Physiol. Biochem. Pharmacol. 2018, 46, 2127–2137. [Google Scholar] [CrossRef] [Green Version]
- George, K.; Thomas, N.S.; Malathi, R. Modulatory Effect of Selected Dietary Phytochemicals on Delayed Rectifier K+ Current in Human Prostate Cancer Cells. J. Membr. Biol. 2019, 252, 195–206. [Google Scholar] [CrossRef]
- Marafiga, J.R.; Pasquetti, M.V.; Calcagnotto, M.E. GABAergic interneurons in epilepsy: More than a simple change in inhibition. Epilepsy Behav. 2020, 106935. [Google Scholar] [CrossRef] [PubMed]
- Hosseini, A.; Mirazi, N. Acute administration of ginger (Zingiber officinale rhizomes) extract on timed intravenous pentylenetetrazol infusion seizure model in mice. Epilepsy Res. 2014, 108, 411–419. [Google Scholar] [CrossRef]
- Lerche, H.; Shah, M.; Beck, H.; Noebels, J.; Johnston, D.; Vincent, A. Ion channels in genetic and acquired forms of epilepsy. J. Physiol. 2013, 591, 753–764. [Google Scholar] [CrossRef] [PubMed]
- Power, K.N.; Gramstad, A.; Gilhus, N.E.; Hufthammer, K.O.; Engelsen, B.A. Cognitive dysfunction after generalized tonic-clonic status epilepticus in adults. Acta. Neurol. Scand. 2018, 137, 417–424. [Google Scholar] [CrossRef]
- Martinc, B.; Grabnar, I.; Vovk, T. Antioxidants as a preventive treatment for epileptic process: A review of the current status. Curr. Neuropharmacol. 2014, 12, 527–550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kandeda, A.K.; Taiwe, G.S.; Moto, F.C.O.; Ngoupaye, G.T.; Nkantchoua, G.C.N.; Njapdounke, J.S.K.; Omam, J.P.O.; Pale, S.; Kouemou, N.; Bum, E.N. Antiepileptogenic and Neuroprotective Effects of Pergularia daemia on Pilocarpine Model of Epilepsy. Front. Pharmacol. 2017, 8, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Pearson-Smith, J.N.; Patel, M. Metabolic dysfunction and oxidative stress in epilepsy. Int. J. Mol. Sci. 2017, 18, E2365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rajan, I.; Narayanan, N.; Rabindran, R.; Jayasree, P.R.; Kumar, P.R.M. Zingerone protects against stannous chloride-induced and hydrogen peroxide-induced oxidative DNA damage in vitro. Biol. Trace. Elem. Res. 2013, 155, 455–459. [Google Scholar] [CrossRef]
- Diniz, T.C.; Silva, J.C.; de Lima-Saraiva, S.R.; Ribeiro, F.P.; Pacheco, A.G.; de Freitas, R.M.; Quintans-Júnior, L.J.; Quintans, J.; Mendes, R.L.; Almeida, J.R. The role of flavonoids on oxidative stress in epilepsy. Oxi. Med. Cell. Longev. 2015, 171756. [Google Scholar] [CrossRef]
- Mani, V.; Arivalagan, S.; Siddique, A.I.; Namasivayam, N. Antioxidant and anti-inflammatory role of zingerone in ethanol-induced hepatotoxicity. Mol. Cell. Biochem. 2016, 421, 169–181. [Google Scholar] [CrossRef]
- Carmona-Aparicio, L.; Zavala-Tecuapetla, C.; González-Trujano, M.E.; Sampieri, A.I.; Montesinos-Correa, H.; Granados-Rojas, L.; Floriano-Sánchez, E.; Coballase-Urrutía, E.; Cárdenas-Rodríguez, N. Status epilepticus: Using antioxidant agents as alternative therapies. Exp. Ther. Med. 2016, 12, 1957–1962. [Google Scholar] [CrossRef]
- Amin, I.; Hussain, I.; Rehman, M.U.; Mir, B.A.; Ganaie, S.A.; Ahmad, S.B.; Mir, M.; Shanaz, S.; Muzamil, S.; Arafah, A.; et al. Zingerone prevents lead-induced toxicity in liver and kidney tissues by regulating the oxidative damage in Wistar rats. J. Food Biochem. 2020, e13241. [Google Scholar] [CrossRef]
- McElroy, P.B.; Liang, L.P.; Day, B.J.; Patel, M. Scavenging reactive oxygen species inhibits status epilepticus-induced neuroinflammation. Exp. Neurol. 2017, 298, 13–22. [Google Scholar] [CrossRef]
- da Silva, A.P.D.S.; Lopes, J.S.; Vieira, P.D.S.; Pinheiro, E.E.; da Silva, M.L.D.G.; Silva Filho, J.C.C.; da Costa Júnior, J.S.; David, J.M.; de Freitas, R.M. Behavioral and neurochemical studies in mice pretreated with garcinielliptone FC in pilocarpine-induced seizures. Pharmacol. Biochem. Behav. 2014, 124, 305–310. [Google Scholar] [CrossRef] [Green Version]
- Gnatek, Y.; Zimmerman, G.; Goll, Y.; Najami, N.; Soreq, H.; Friedman, A. Acetylcholinesterase loosens the brain’s cholinergic anti-inflammatory response and promotes epileptogenesis. Front. Mol. Neurosci. 2012, 5, 66. [Google Scholar] [CrossRef] [Green Version]
- Aronica, E.; Bauer, S.; Bozzi, Y.; Caleo, M.; Dingledine, R.; Gorter, J.A.; Henshall, D.C.; Kaufer, D.; Koh, S.; Löscher, W.; et al. Neuroinflammatory targets and treatments for epilepsy validated in experimental models. Epilepsia 2017, 58, 27–38. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, B.; Rehman, M.U.; Amin, I.; Mir, M.; Ahmad, S.B.; Farooq, A.; Muzamil, S.; Hussain, I.; Masoodi, M.; Fatima, B. Zingerone (4-(4-hydroxy-3-methylphenyl) butan-2-one) protects against alloxan-induced diabetes via alleviation of oxidative stress and inflammation: Probable role of NF-kB activation. Saud. Pharm. J. 2018, 26, 1137–1145. [Google Scholar] [CrossRef]
- Shimada, T.; Takemiya, T.; Sugiura, H.; Yamagata, K. Role of inflammatory mediators in the pathogenesis of epilepsy. Mediators. Inflamm. 2014, 901902. [Google Scholar] [CrossRef]
- Wali, A.F.; Rehman, M.U.; Raish, M.; Kazi, M.; Rao, P.; Alnemer, O.; Ahmad, P.; Ahmad, A. Zingerone [4-(3-Methoxy-4-hydroxyphenyl)-butan-2] Attenuates Lipopolysaccharide-Induced Inflammation and Protects Rats from Sepsis Associated Multi Organ Damage. Molecules 2020, 25, 5127. [Google Scholar] [CrossRef]
- Borges, K.; Gearing, M.; McDermott, D.L.; Smith, A.B.; Almonte, A.G.; Wainer, B.H.; Dingledine, R. Neuronal and glial pathological changes during epileptogenesis in the mouse pilocarpine model. Exp. Neurol. 2003, 182, 21–34. [Google Scholar] [CrossRef]
- Ahmad, B.; Rehman, M.U.; Amin, I.; Arif, A.; Rasool, S.; Bhat, S.A.; Afzal, I.; Hussain, I.; Bilal, S.; Mir, M.U.R. A Review on Pharmacological Properties of Zingerone (4-(4-Hydroxy-3-methoxyphenyl)-2-butanone). Sci. World J. 2015, 816364. [Google Scholar] [CrossRef] [Green Version]
- Džoljić, E.; Grbatinić, I.; Kostić, V. Why is nitric oxide important for our brain? Funct. Neurol. 2015, 30, 159–163. [Google Scholar] [CrossRef]
- Mir, B.; Amin, I.; Rehman, M.U.; Bhat, R.; Ali, A.; Baba, O.K.; Fatima, B.; Ali, R.; Ahmad, S.B.; Muzamil, S.; et al. Chemoprotective potential of zingerone (vanillyl acetone) in cyclophosphamide-induced hepatic toxicity. Pharmacogn. Mag. 2018, 14, 434–439. [Google Scholar] [CrossRef]
- Vasconcelos Rios, E.R.; Moura Rocha, N.F.; Rodrigues Carvalho, A.M.; Freire Vasconcelos, L.; Leite Dias, M.; de Carvalho Lima, C.N.; Soares Lopes, K.; Cavalcante Melo, F.H.; de França Fonteles, M.M. Involvement of the nitric oxide/cyclic guanylate monophosphate pathway in the pilocarpine-induced seizure model in mice. Pharmacology 2013, 91, 131–134. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Seeburg, D.P.; Pulli, B.; Wojtkiewicz, G.R.; Bure, L.; Atkinson, W.; Schob, S.; Iwamoto, Y.; Ali, M.; Zhang, W.; et al. Myeloperoxidase Nuclear Imaging for Epileptogenesis. Radiology 2016, 278, 822–830. [Google Scholar] [CrossRef] [Green Version]
- Rehman, M.U.; Ahmad, B.; Arif, A.; Rasool, S.; Farooq, A.; Razzaq, R.; Bhat, S.A.; Bashir, S.; Shabir, O.; Amin, I.; et al. Zingerone protects against cisplatin-induced oxidative damage in the jejunum of Wistar rats. Orient. Pharm. Exp. Med. 2015, 15, 199–206. [Google Scholar] [CrossRef]
- Marisela, M.A.; Concepción, N.R.; Daniel, J.R.; Erika, R.M.; Petra, Y.G. Oxidative Stress Associated with Neuronal Apoptosis in Experimental Models of Epilepsy. Oxidative Med. Cell. Longev. 2014, 293689, 12. [Google Scholar] [CrossRef]
- Engel, T.; Henshall, D.C. Apoptosis, Bcl-2 family proteins and caspases: The ABCs of seizure-damage and epileptogenesis? Int. J. Physiol. Pathophysiol. Pharmacol. 2009, 1, 97–115. [Google Scholar] [PubMed]
- Mao, X.Y.; Zhou, H.H.; Jin, W.L. Redox-Related Neuronal Death and Crosstalk as Drug Targets: Focus on Epilepsy. Front. Neurosci. 2019, 13, 512. [Google Scholar] [CrossRef] [Green Version]
- Folbergrová, J.; Kunz, W.S. Mitochondrial dysfunction in epilepsy. Mitochondrion 2012, 12, 35–40. [Google Scholar] [CrossRef]
- Cho, I.; Cho, Y.J.; Kim, H.W.; Heo, K.; Lee, B.I.; Kim, W.J. Effect of Androsterone after Pilocarpine-induced Status Epilepticus in Mice. J. Epilepsy Res. 2014, 4, 7–13. [Google Scholar] [CrossRef]
- Ahmed Juvale, I.I.; Che Has, A.T. The evolution of the pilocarpine animal model of status epilepticus. Heliyon 2020, 6, e04557. [Google Scholar] [CrossRef] [PubMed]
- Kandemir, F.M.; Yildirim, S.; Caglayan, C.; Kucukler, S.; Eser, G. Protective effects of zingerone on cisplatin-induced nephrotoxicity in female rats. Environ. Sci. Pollut. Res. 2019, 26, 22562–22574. [Google Scholar] [CrossRef]
- Kaygusuzoglu, E.; Caglayan, C.; Kandemir, F.M.; Yıldırım, S.; Kucukler, S.; Kılınc, M.A.; Saglam, Y.S. Zingerone ameliorates cisplatin-induced ovarian and uterine toxicity via suppression of sex hormone imbalances, oxidative stress, inflammation and apoptosis in female wistar rats. Biomed. Pharmacother. 2018, 102, 517–530. [Google Scholar] [CrossRef]
- Bilal, A.; Insha, A.; Towseef, A.; Muneeb, U.R.; Showkat, A.; Saiema, R.; Ahmad, A.; Adil, F.; Showkeen, M.; Ishraq, H.; et al. Zingerone (4-(4-hydroxy-3-methoxyphenyl)-2-butanone) Protects Against Acetaminophen Induced Hepatotoxicity in Wistar Rats via Alleviation of Oxidative Stress and Inflammation. Asian J. Anim. Vet. Adv. 2016, 11, 832–839. [Google Scholar]
- Shakeel, S.; Rehman, M.U.; Tabassum, N.; Amin, U.; Mir, R. Effect of Naringenin (A naturally occurring flavanone) Against Pilocarpine-induced Status Epilepticus and Oxidative Stress in Mice. Pharmacogn. Mag. 2017, 13, 1–14. [Google Scholar] [CrossRef]
- Vorhees, C.V.; Williams, M.T. Morris water maze: Procedures for assessing spatial and related forms of learning and memory. Nat. Protocol. 2006, 1, 848–858. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wright, J.R.; Colby, H.D.; Miles, P.R. Cytosolic factors which affect microsomal lipid peroxidation in lung and liver. Arch. Biochem. Biophys. 1981, 206, 296–304. [Google Scholar] [CrossRef]
- Marklund, S.; Marklund, G. Involvement of the superoxide anion radical in the autoxidation of pyrogallol and a convenient assay for superoxide dismutase. Eur. J. Biochem. 1974, 47, 469–474. [Google Scholar] [CrossRef] [PubMed]
- Claiborne, A. Catalase activity. In CRC Handbook of Methods in Oxygen Radical Research; Greenwald, R.A., Ed.; CRC: Boca Raton, FL, USA, 1985; pp. 283–284. [Google Scholar]
- Rashid, S.; Ali, N.; Nafees, S.; Hasan, S.K.; Sultana, S. Mitigation of 5-Fluorouracil induced renal toxicity by chrysin via targeting oxidative stress and apoptosis in wistar rats. Food. Chem. Toxicol. 2014, 66, 185–193. [Google Scholar] [CrossRef] [PubMed]
- Ellman, G.L.; Courtney, K.D.; Andres, V., Jr.; Featherstone, R.M. A new and rapid colorimetric determination of acetylcholinesterase activity. Biochem. Pharmacol. 1961, 7, 88–95. [Google Scholar] [CrossRef]
- Bradley, P.P.; Priebat, D.A.; Christensen, R.D.; Rothstein, G. Measurement of cutaneous inflammation: Estimation of neutrophil content with an enzyme marker. J. Investig. Dermatol. 1982, 78, 206–209. [Google Scholar] [CrossRef] [Green Version]
- Liua, C.M.; Mab, J.Q.; Suna, J.Q. Quercetin protects the rat kidney against oxidative stress-mediated DNA damage and apoptosis induced by lead. Environ. Toxicol. Pharmacol. 2010, 30, 264–271. [Google Scholar] [CrossRef]
- Green, L.C.; Wagner, D.A.; Glogowski, J.; Skipper, P.L.; Wishnok, J.S.; Tannenbaum, S.R. Analysis of nitrate, nitrite, and [15N] nitrate in biological fluids. Anal. Biochem. 1982, 126, 131–138. [Google Scholar] [CrossRef]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L. Protein measurement with the Folin Phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef]
- Rashid, S.; Nafees, S.; Vafa, A.; Afzal, S.M.; Ali, N.; Rehman, M.U.; Hasan, S.K.; Siddiqi, A.; Barnwal, P.; Majed, F.; et al. Inhibition of precancerous lesions development in kidneys by chrysin via regulating hyperproliferation, inflammation and apoptosis at pre clinical stage. Arch. Biochem. Biophys. 2016, 606, 1–9. [Google Scholar] [CrossRef]
- Szilágyi, T.; Orbán-Kis, K.; Horváth, E.; Metz, J.; Pap, Z.; Pávai, Z. Morphological identification of neuron types in the rat hippocampus. Romanian J. Morphol. Embryol. Rev. Roum. Morphol. Embryol. 2011, 52, 15–20. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Animals/Convulsion Score | Group I | Group II | Group III | Group IV | Group V |
|---|---|---|---|---|---|
| No. of Animals | 14 | 14 | 14 | 14 | 14 |
| Latency to First Convulsion (s) | 0 | 564 ± 44 | 3473 ± 48 *** | 864 ± 55 ** | 1194 ± 56 *** |
| Percentage Convulsion (%) | 0 | 100 | 10 | 100 | 42.6 |
| Parameters | Group I | Group II | Group III | Group IV | Group V |
|---|---|---|---|---|---|
| LPO (n moles MDA formed/g tissue) | 2.92 ± 0.03 | 6.09 ± 0.42 *** | 3.32 ± 0.43 ### | 4.80 ± 0.54 # | 3.43 ± 0.54 ### |
| SOD (U/mg Protein) | 274.8 ± 19.74 | 105.5 ± 10.53 *** | 247.1 ± 13.03 ### | 187.8 ± 12.93 # | 233.51 ± 20.12 ### |
| CAT (n moles/min/mg protein) | 287.2 ± 20.63 | 97.34 ± 9.14 *** | 251.02 ± 23.05 ### | 191.05 ± 12.32 # | 243.37 ± 17.18 ### |
| GR (n moles/min/mg protein) | 232.6 ± 21.02 | 151.7 ± 15.01 *** | 219.2 ± 21.2 ### | 217.8 ± 11.54 ## | 210.60 ± 19.3 ## |
| GSH (n mol/mg Protein) | 292.8 ± 16.32 | 119.64 ± 11.8 *** | 277.98 ± 18.0 ### | 141.37 ± 13.1 # | 219.63 ± 13.7 ## |
| Inflammatory Markers | Group I | Group II | Group III | Group IV | Group V |
|---|---|---|---|---|---|
| NFk-B (pg/mL) | 732.03 ± 53.4 | 1643.0 ± 136.7 *** | 893.02 ± 67.0 ### | 1342.5 ± 92.7 # | 922.11 ± 87.1 ### |
| TNF-α (pg/mL) | 244.82 ± 19.4 | 765.62 ± 44.2 *** | 269.05 ± 20.4 ### | 517.81 ± 31.6 ## | 288.53 ± 19.1 ### |
| IL-6 (pg/mL) | 823.26 ± 47.2 | 1694.0 ± 140.2 *** | 946.03 ± 40.2 ### | 1440.5 ± 87.4 ### | 997.16 ± 73.2 ## |
| IL-1β (pg/mL) | 739.10 ± 51.8 | 1828.9 ± 121.8 *** | 855.45 ± 74.0 ### | 1514.1 ± 103.6 # | 893.73 ± 67.9 ### |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rashid, S.; Wali, A.F.; Rashid, S.M.; Alsaffar, R.M.; Ahmad, A.; Jan, B.L.; Paray, B.A.; Alqahtani, S.M.A.; Arafah, A.; Rehman, M.U. Zingerone Targets Status Epilepticus by Blocking Hippocampal Neurodegeneration via Regulation of Redox Imbalance, Inflammation and Apoptosis. Pharmaceuticals 2021, 14, 146. https://doi.org/10.3390/ph14020146
Rashid S, Wali AF, Rashid SM, Alsaffar RM, Ahmad A, Jan BL, Paray BA, Alqahtani SMA, Arafah A, Rehman MU. Zingerone Targets Status Epilepticus by Blocking Hippocampal Neurodegeneration via Regulation of Redox Imbalance, Inflammation and Apoptosis. Pharmaceuticals. 2021; 14(2):146. https://doi.org/10.3390/ph14020146
Chicago/Turabian StyleRashid, Summya, Adil Farooq Wali, Shahzada Mudasir Rashid, Rana M. Alsaffar, Ajaz Ahmad, Basit L. Jan, Bilal Ahmad Paray, Saeed M. A. Alqahtani, Azher Arafah, and Muneeb U. Rehman. 2021. "Zingerone Targets Status Epilepticus by Blocking Hippocampal Neurodegeneration via Regulation of Redox Imbalance, Inflammation and Apoptosis" Pharmaceuticals 14, no. 2: 146. https://doi.org/10.3390/ph14020146