
Molecular Mimicry between SARS-CoV-2 and Human Endocrinocytes: A Prerequisite of Post-COVID-19 Endocrine Autoimmunity?



Abstract
:1. Introduction
2. Materials and Methods
3. Results
4. Discussion
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mereschkowski, C. Über Natur und Ursprung der Chromatophoren im Pflanzenreiche. Biol. Centralbl. 1905, 25, 593–604. [Google Scholar]
- Mereschkowsky, K. Theorie der zwei Plasmaarten als Grundlage der Symbiogenesis, einer neuen Lehre von der Ent-stehung der Organismen. Biol. Centralbl. 1910, 30, 353–367. [Google Scholar]
- Kaplan, M.H.; Meyeserian, M. An immunologic cross-reaction between group A streptococcal cells and human heart. Lancet 1962, 1, 706–710. [Google Scholar] [CrossRef]
- Damian, R.T. Molecular mimicry: Antigen sharing by parasite and host and its consequences. Am. Nat. 1964, 98, 129–149. Available online: https://www.jstor.org/stable/2459352 (accessed on 1 June 2020). [CrossRef]
- Fujinami, R.S.; Oldstone, M.B. Amino acid homology between the encephalitogenic site of myelin basic protein and virus: Mechanism for autoimmunity. Science 1985, 230, 1043–1045. [Google Scholar] [CrossRef] [PubMed]
- Vermelho, A.B.; de Meirelles, M.N.; Pereira, M.C.; Pohlentz, G.; BarretoBergter, E. Heart muscle cells share common neutral glycosphingolipids with Trypanosoma cruzi. Acta Trop. 1997, 64, 131–143. [Google Scholar] [CrossRef]
- Blank, M.; Barzilai, O.; Shoenfeld, Y. Molecular mimicry and auto-immunity. Clin. Rev. Allergy Immunol. 2007, 32, 111–118. [Google Scholar] [CrossRef] [PubMed]
- Eisenbarth, G.S. Immunoendocrinology: Scientific and Clinical Aspects; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2011. [Google Scholar]
- Plotz, P.H. Autoantibodies are anti-idiotype antibodies to antiviral antibodies. Lancet 1983, 2, 824–826. [Google Scholar] [CrossRef]
- Heyma, P.; Harrison, L.C.; Robins-Browne, R. Thyrotrophin (TSH) binding sites on Yersinia enterocolitica recognized by immunoglobulins from humans with Graves’ disease. Clin. Exp. Immunol. 1986, 64, 249–254. [Google Scholar]
- Matzinger, P. The danger model: A renewed sense of self. Science 2002, 296, 301–305. [Google Scholar] [CrossRef]
- Cheung, R.; Karjalainen, J.; Vandermeulen, J.; Singal, D.P.; Dosch, H.M. T cells from children with IDDM are sensitized to bovine serum albumin. Scand. J. Immunol. 1994, 40, 623–628. [Google Scholar] [CrossRef] [PubMed]
- Chunder, R.; Weier, A.; Mäurer, H.; Luber, N.; Enders, M.; Luber, G.; Heider, T.; Spitzer, A.; Tacke, S.; Becker-Gotot, J.; et al. Antibody cross-reactivity between casein and myelin-associated glycoprotein results in central nervous system demyelination. Proc. Natl. Acad. Sci. USA 2022, 119, e2117034119. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Aoki, V.; Liu, Z.; Prisayanh, P.; Valenzuela, J.G.; Diaz, L.A. From Insect Bites to a Skin Autoimmune Disease: A Conceivable Pathway to Endemic Pemphigus Foliaceus. Front. Immunol. 2022, 13, 907424. [Google Scholar] [CrossRef] [PubMed]
- Vojdani, A.; Rahimian, P.; Kalhor, H.; Mordechai, E. Immunological cross reactivity between Candida albicans and human tissue. J. Clin. Lab. Immunol. 1996, 48, 1–15. [Google Scholar] [PubMed]
- Ferré, E.M.N.; Schmitt, M.M.; Lionakis, M.S. Autoimmune Polyendocrinopathy-Candidiasis-Ectodermal Dystrophy. Front. Pediatr. 2021, 9, 723532. [Google Scholar] [CrossRef]
- Worldometer. Available online: https://www.worldometers.info/coronavirus/ (accessed on 21 July 2022).
- Ryabkova, V.A.; Churilov, L.P.; Shoenfeld, Y. Influenza infection, SARS, MERS and COVID-19: Cytokine storm—The common denominator and the lessons to be learned. Clin. Immunol. 2021, 223, 108652. [Google Scholar] [CrossRef]
- Dotan, A.; Muller, S.; Kanduc, D.; David, P.; Halpert, G.; Shoenfeld, Y. The SARS-CoV-2 as an instrumental trigger of autoimmunity. Autoimmun. Rev. 2021, 20, 102792. [Google Scholar] [CrossRef]
- Shoenfeld, Y. Corona (COVID-19) time musings: Our involvement in COVID-19 pathogenesis, diagnosis, treatment and vaccine planning. Autoimmun. Rev. 2020, 19, 102538. [Google Scholar] [CrossRef]
- Cappello, F. Is COVID-19 a proteiform disease inducing also molecular mimicry phenomena? Cell Stress Chaperones 2020, 25, 381–382. [Google Scholar] [CrossRef]
- Kanduc, D.; Shoenfeld, Y. On the molecular determinants of the SARS-CoV-2 attack. Clin. Immunol. 2020, 215, 108426. [Google Scholar] [CrossRef]
- Lucchese, G.; Flöel, A. Molecular mimicry between SARS-CoV-2 and respiratory pacemaker neurons. Autoimmun. Rev. 2020, 19, 102556. [Google Scholar] [CrossRef] [PubMed]
- Angileri, F.; Legare, S.; Marino Gammazza, A.; Conway de Macario, E.; Jl Macario, A.; Cappello, F. Molecular mimicry may explain multi-organ damage in COVID-19. Autoimmun. Rev. 2020, 19, 102591. [Google Scholar] [CrossRef] [PubMed]
- Frara, S.; Allora, A.; Castellino, L.; di Filippo, L.; Loli, P.; Giustina, A. COVID-19 and the pituitary. Pituitary 2021, 24, 465–481. [Google Scholar] [CrossRef] [PubMed]
- Ehrenfeld, M.; Tincani, A.; Andreoli, L.; Cattalini, M.; Greenbaum, A.; Kanduc, D.; Alijotas-Reig, J.; Zinserling, V.; Semenova, N.; Amital, H.; et al. COVID-19 and autoimmunity. Autoimmun. Rev. 2020, 19, 102597. [Google Scholar] [CrossRef]
- Churilov, L.P.; Kanduc, D.; Ryabkova, V.A. COVID-19: Adrenal response and molecular mimicry. IMAJ 2021, 23, 618–619. [Google Scholar]
- Kanduc, D. Homology, similarity, and identity in peptide epitope immunodefinition. J. Pept. Sci. 2012, 18, 487–494. [Google Scholar] [CrossRef]
- UniProt Consortium. UniProt: A worldwide hub of protein knowledge. Nucleic Acids Res. 2019, 47, D506–D515. [Google Scholar] [CrossRef]
- Immune Epitope Database and Analysis Resource, IEDB. Available online: https://www.iedb.org (accessed on 21 July 2022).
- National Center of Biotechnology Information (NCBI). Available online: https://www.ncbi.nlm.nih.gov (accessed on 21 July 2022).
- Tutal, E.; Ozaras, R.; Leblebicioglu, H. Systematic review of COVID-19 and autoimmune thyroiditis. Travel Med. Infect. Dis. 2022, 47, 102314. [Google Scholar] [CrossRef]
- Boddu, S.K.; Aurangabadkar, G.; Kuchay, M.S. New onset diabetes, type 1 diabetes and COVID-19. Diabetes Metab. Syndr. 2020, 14, 2211–2217. [Google Scholar] [CrossRef]
- Steenblock, C.; Schwarz, P.E.H.; Ludwig, B.; Linkermann, A.; Zimmet, P.; Kulebyakin, K.; Tkachuk, V.A.; Markov, A.G.; Lehnert, H.; de Angelis, M.H.; et al. COVID-19 and metabolic disease: Mechanisms and clinical management. Lancet Diabetes Endocrinol. 2021, 9, 786–798. [Google Scholar] [CrossRef]
- Tang, X.; He, B.; Liu, Z.; Zhou, Z.; Li, X. Fulminant type 1 diabetes after COVID-19 vaccination. Diabetes Metab. 2022, 48, 101324. [Google Scholar] [CrossRef] [PubMed]
- Siejka, A.; Barabutis, N. Adrenal insufficiency in the COVID-19 era. Am. J. Physiol. Endocrinol. Metab. 2021, 320, E784–E785. [Google Scholar] [CrossRef] [PubMed]
- Zinserling, V.A.; Semenova, N.Y.; Markov, A.G.; Rybalchenko, O.V.; Wang, J.; Rodionov, R.N.; Bornstein, S.R. Inflammatory Cell Infiltration of Adrenals in COVID-19. Horm. Metab. Res. 2020, 52, 639–641. [Google Scholar] [CrossRef] [PubMed]
- Joshi, M.; Gunawardena, S.; Goenka, A.; Ey, E.; Kumar, G. Post COVID-19 Lymphocytic Hypophysitis: A Rare Presentation. Child Neurol. Open 2022, 9, 2329048X221103051. [Google Scholar] [CrossRef]
- Misgar, R.A.; Rasool, A.; Wani, A.I.; Bashir, M.I. Central diabetes insipidus (infundibuloneurohypophysitis): A late complication of COVID-19 infection. J. Endocrinol. Investig. 2021, 44, 2855–2856. [Google Scholar] [CrossRef]
- Bogdanos, D.P.; Baum, H.; Gunsar, F.; Arioli, D.; Polymeros, D.; Ma, Y.; Burroughs, A.K.; Vergani, D. Extensive homology between the major immunodominant mitochondrial antigen in primary biliary cirrhosis and Helicobacter pylori does not lead to immunological cross-reactivity. Scand. J. Gastroenterol. 2004, 39, 981–987. [Google Scholar] [CrossRef]
- Bogdanos, D.P.; Pares, A.; Baum, H.; Caballeria, L.; Rigopoulou, E.I.; Ma, Y.; Burroughs, A.K.; Rodes, J.; Vergani, D. Disease-specific cross-reactivity between mimicking peptides of heat shock protein of Mycobacterium gordonae and dominant epitope of E2 subunit of pyruvate dehydrogenase is common in Spanish but not British patients with primary biliary cirrhosis. J. Autoimmun. 2004, 22, 353–362. [Google Scholar] [CrossRef]
- Vojdani, A.; Vojdani, E.; Kharrazian, D. Reaction of Human Monoclonal Antibodies to SARS-CoV-2 Proteins With Tissue Antigens: Implications for Autoimmune Diseases. Front. Immunol. 2021, 11, 617089. [Google Scholar] [CrossRef]



{kind=link}
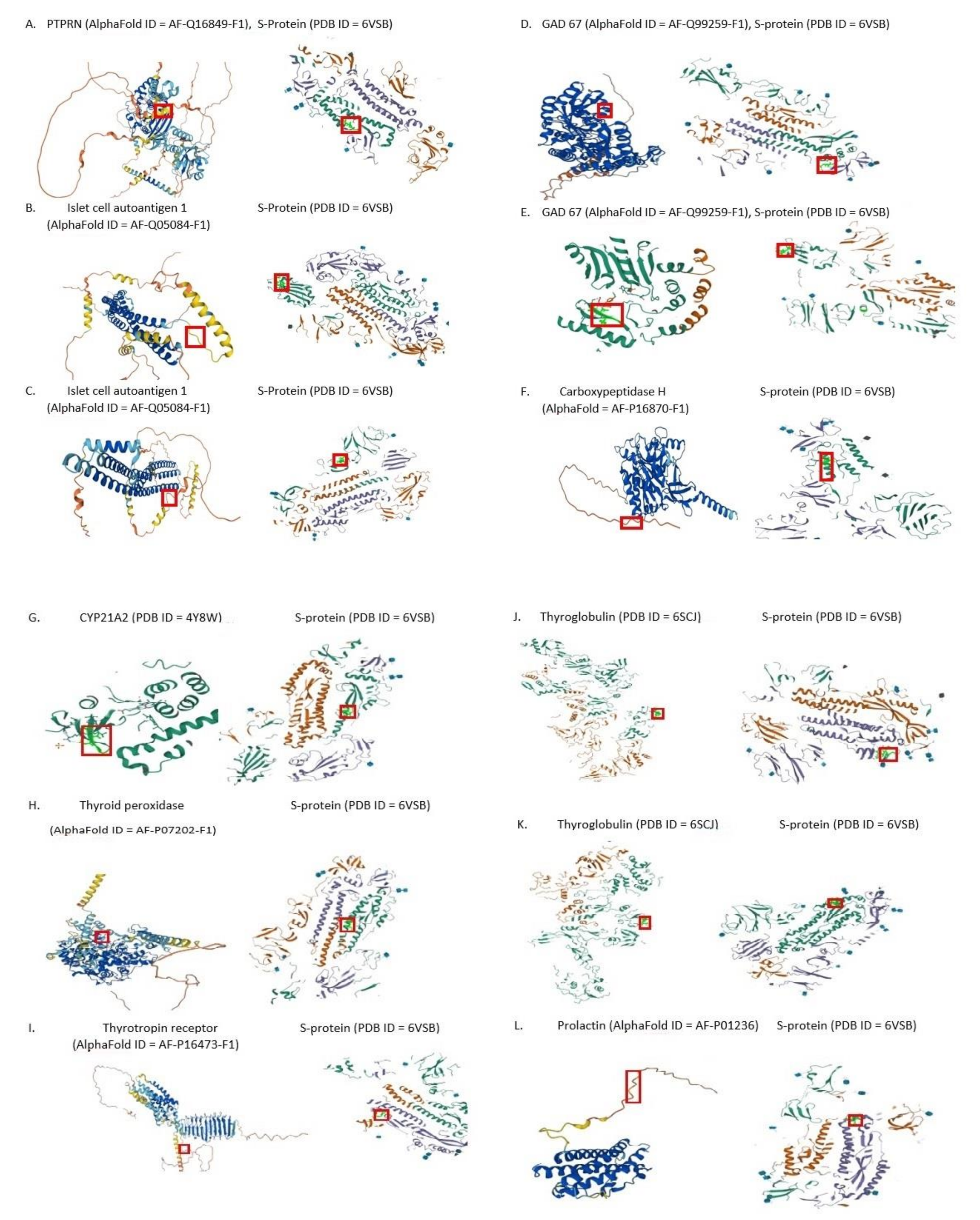{kind=link}
| Langerhans’ Islets β-Cell Autoantigens | Shared Pentapeptides |
|---|---|
| PTPRN (Q16849) | LPPLL |
| Islet cell autoantigen 1 (Q05084) | GYQPY, LDPLS |
| GAD67 (Q99259) | AGAAL, VGYQP |
| Carboxypeptidase H (P16870) | SALLA |
| Autoantigen of Adrenocorticocytes | Shared Pentapeptides |
|---|---|
| CYP21A2 (P08686) | LQDVV |
| Thyroid Autoantigens | Shared Pentapeptides |
|---|---|
| Thyroid peroxidase (P07202) | RAAEI |
| Thyrotropin receptor (P16473) | ICGDS, LLPLV |
| Thyroglobulin (P01266) | FNFSQ, SAIGK, LDSKT |
| Pituitary Autoantigen | Shared Pentapeptide |
|---|---|
| Prolactin (P01236) | SNLLL |
| IEDB ID of an Immunoreactive Epitope | Epitope Sequence |
|---|---|
| 1125063 | gltvLPPLL |
| 1309589 | sygfqptngvGYQPYrvvvl |
| 1074866 | caLDPLSetk |
| 531783 | gAGAALqipfamqma |
| 1310448 | gkLQDVVnqnaqaln |
| 100428 | qliRAAEIrasanlaatk |
| 1310877 | vdctmyICGDStecs |
| 1071273 | LLPLVssqcvnlttr |
| 1087679 | pikdfggFNFSQilpdps |
| 1071651 | nqfnSAIGKiqdsls |
| 1075075 | tLDSKTqsl |
| 1069347 | dstecSNLLLqygsf |
| 1496254 | qytSALLAgtit |
| 1309589 | sygfqptngVGYQPyrvvvl |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Churilov, L.P.; Normatov, M.G.; Utekhin, V.J. Molecular Mimicry between SARS-CoV-2 and Human Endocrinocytes: A Prerequisite of Post-COVID-19 Endocrine Autoimmunity? Pathophysiology 2022, 29, 486-494. https://doi.org/10.3390/pathophysiology29030039
Churilov LP, Normatov MG, Utekhin VJ. Molecular Mimicry between SARS-CoV-2 and Human Endocrinocytes: A Prerequisite of Post-COVID-19 Endocrine Autoimmunity? Pathophysiology. 2022; 29(3):486-494. https://doi.org/10.3390/pathophysiology29030039
Chicago/Turabian StyleChurilov, Leonid P., Muslimbek G. Normatov, and Vladimir J. Utekhin. 2022. "Molecular Mimicry between SARS-CoV-2 and Human Endocrinocytes: A Prerequisite of Post-COVID-19 Endocrine Autoimmunity?" Pathophysiology 29, no. 3: 486-494. https://doi.org/10.3390/pathophysiology29030039