Genotype-Specific Antioxidant Responses and Assessment of Resistance Against Sclerotinia sclerotiorum Causing Sclerotinia Rot in Indian Mustard

 , , ,
, , ,  , ,
, ,
Abstract
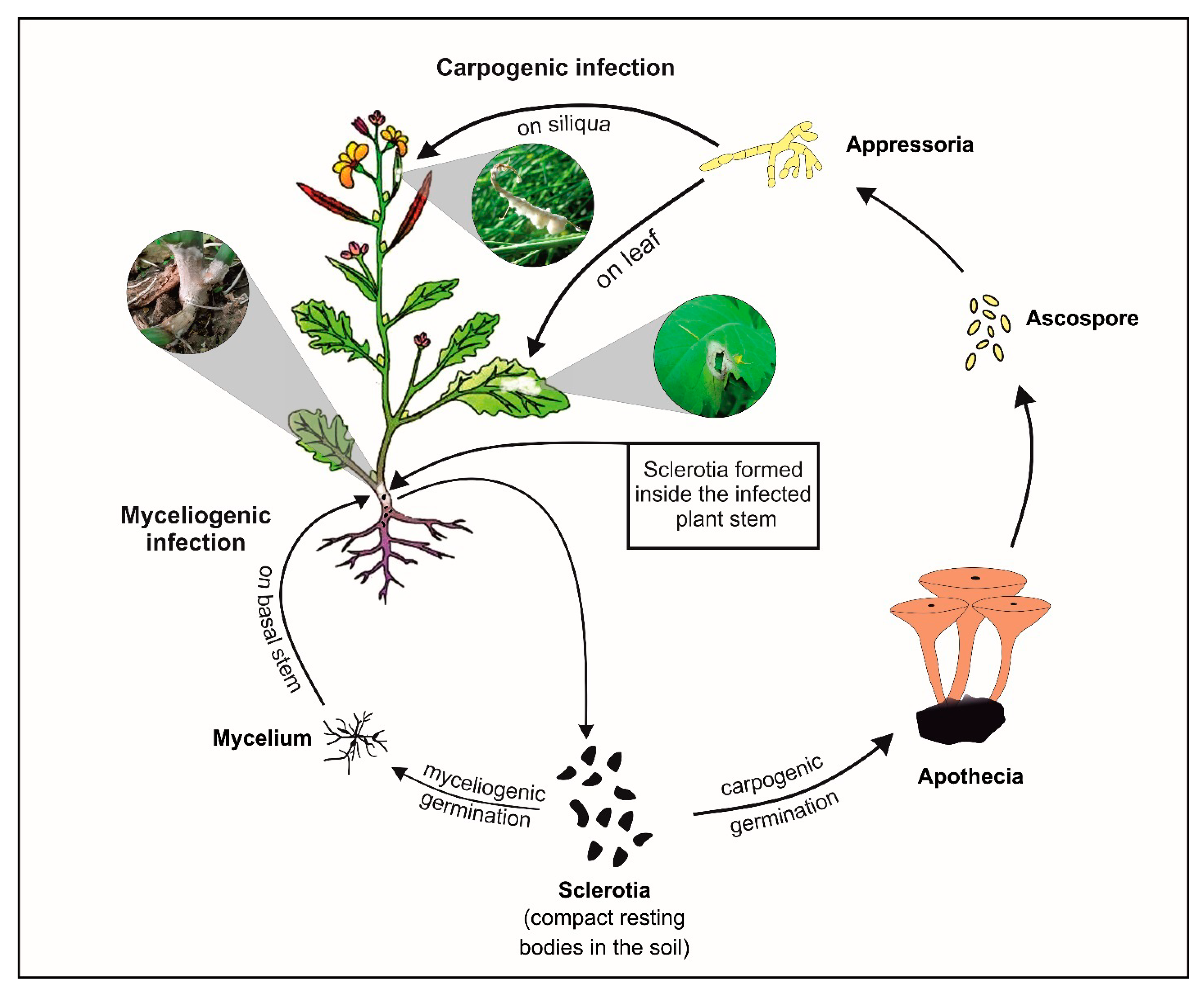
:1. Introduction
2. Results
2.1. Disease Assessment
2.2. Biochemical Analysis
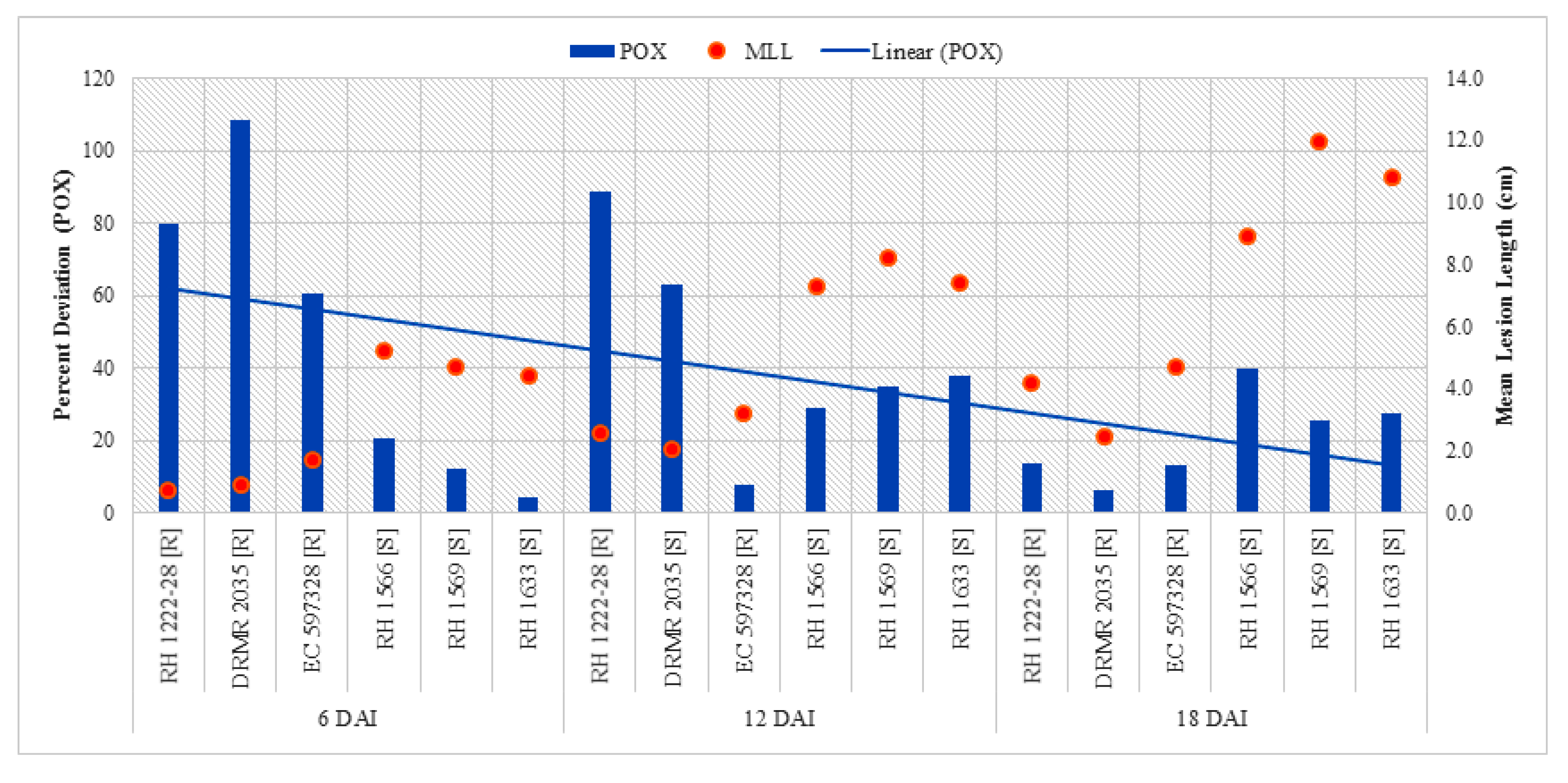
2.2.1. Peroxidase (POX) Activity
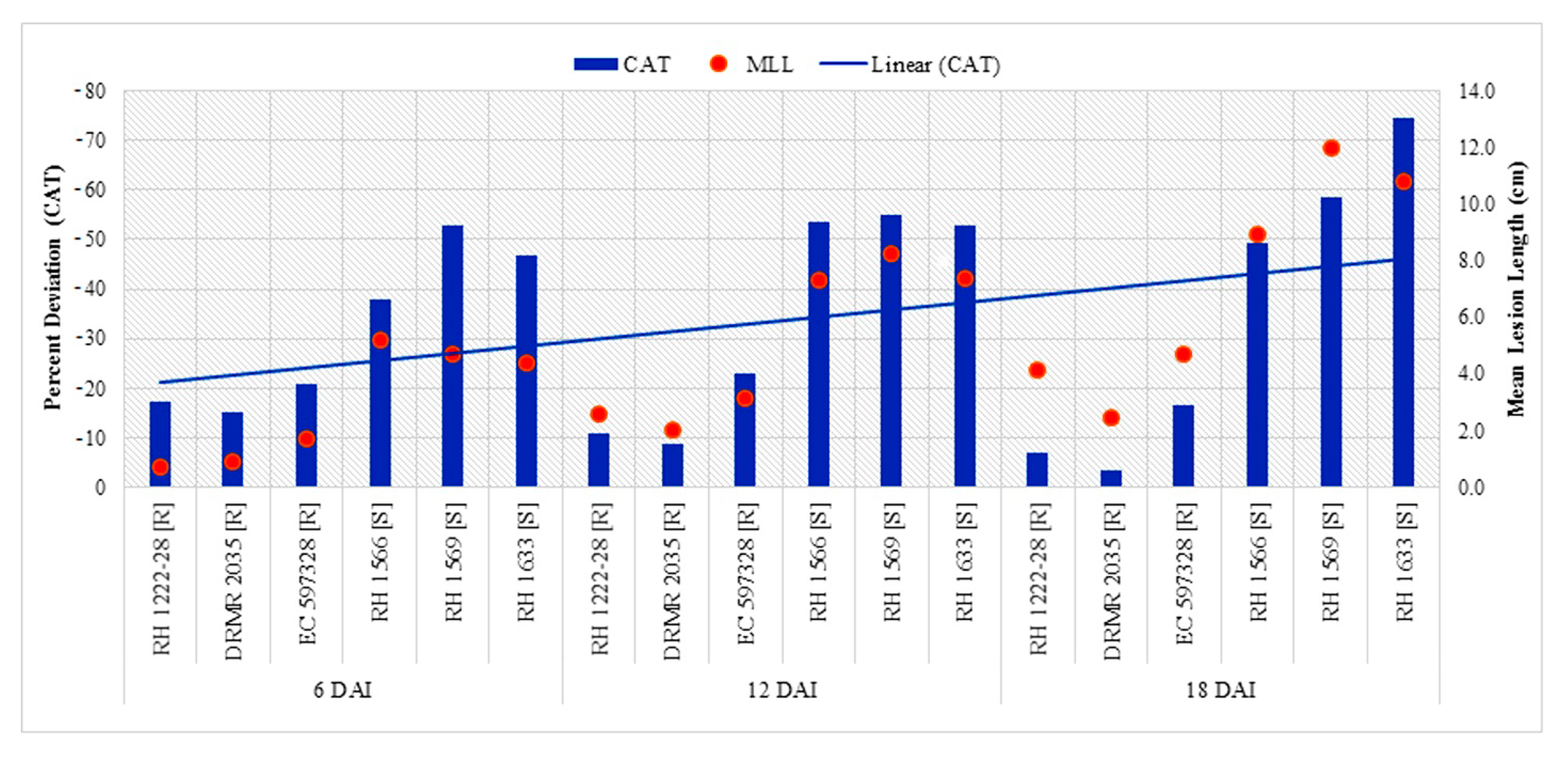
2.2.2. Catalase (CAT) Activity
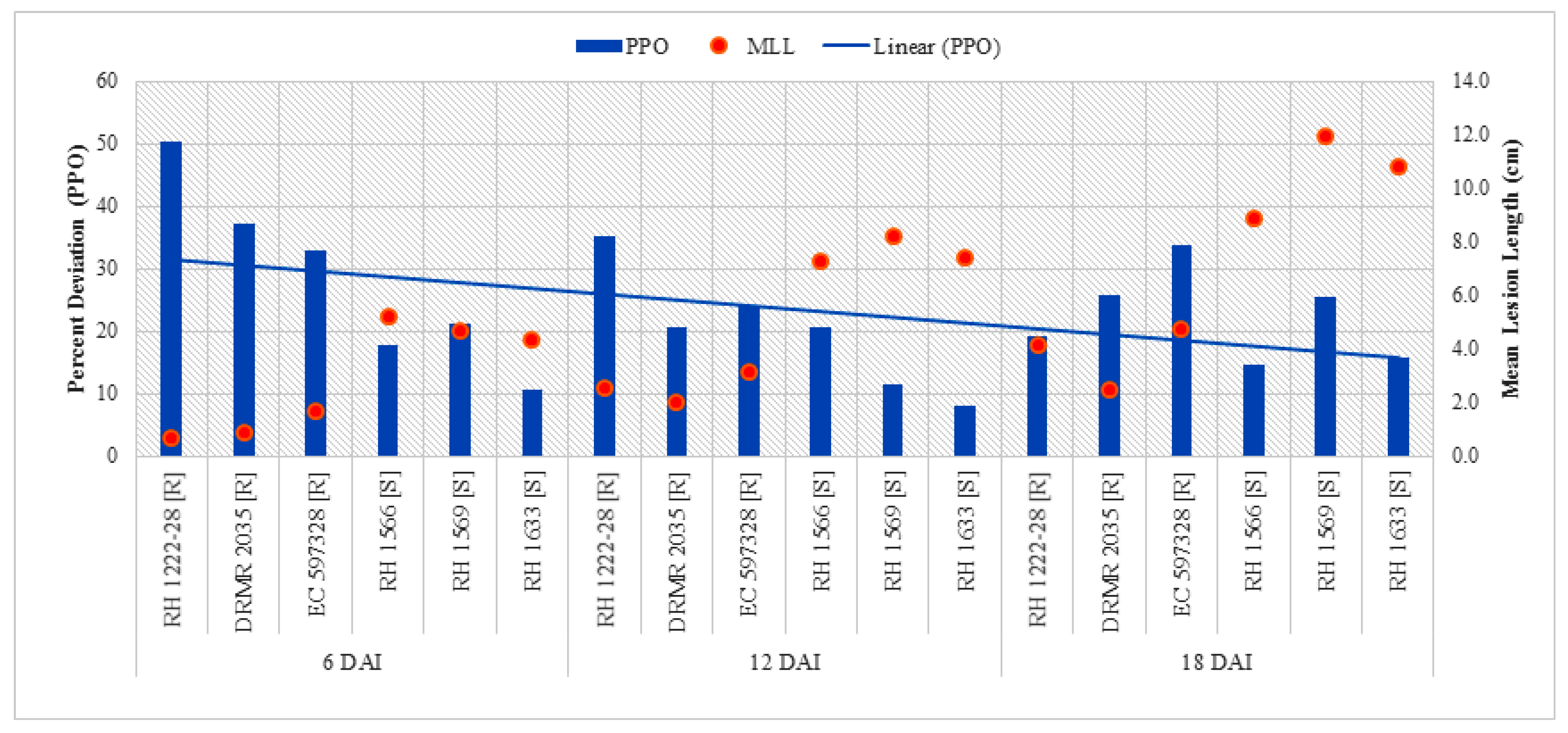
2.2.3. Polyphenol Oxidase (PPO) Activity
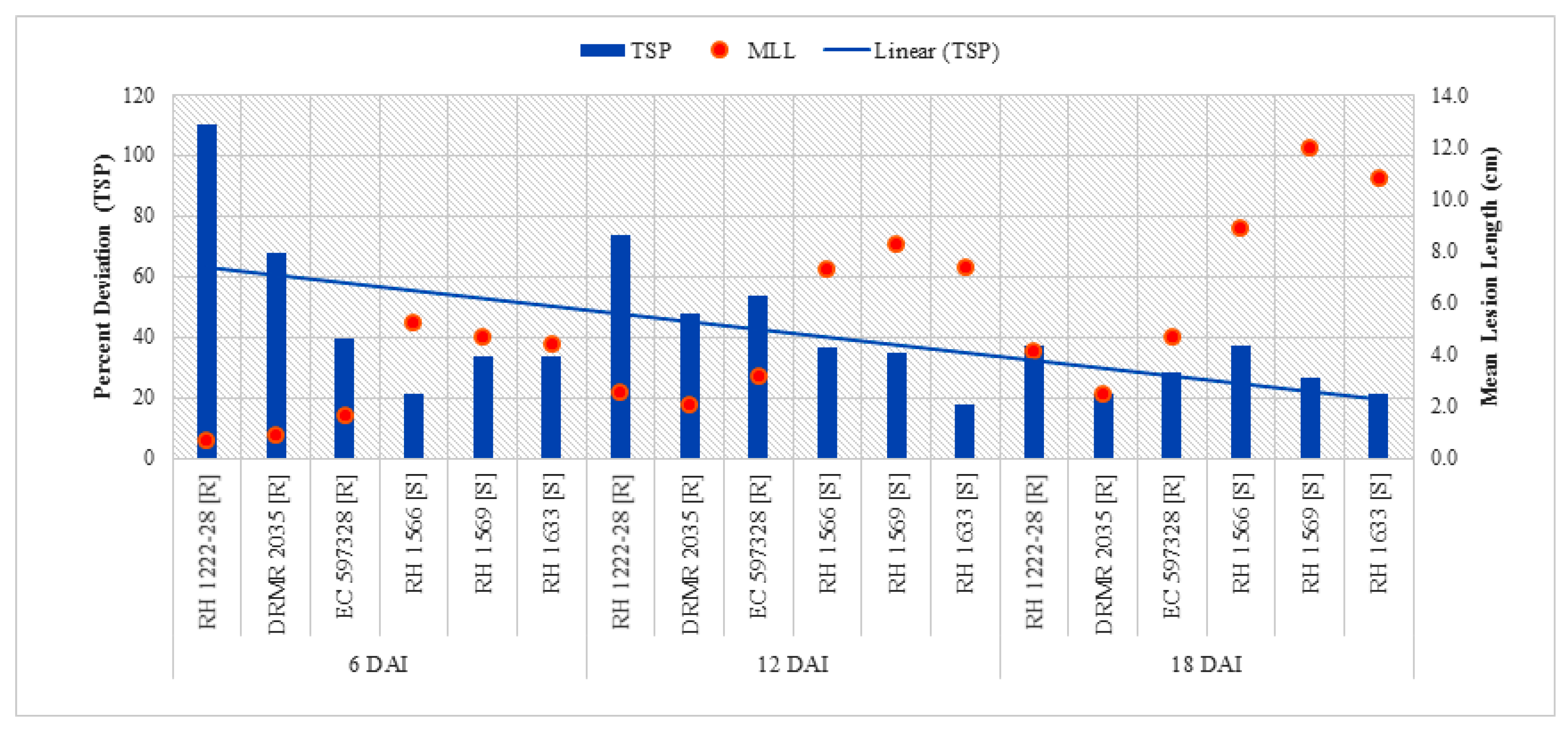
2.2.4. Total Soluble Phenolics (TSP)
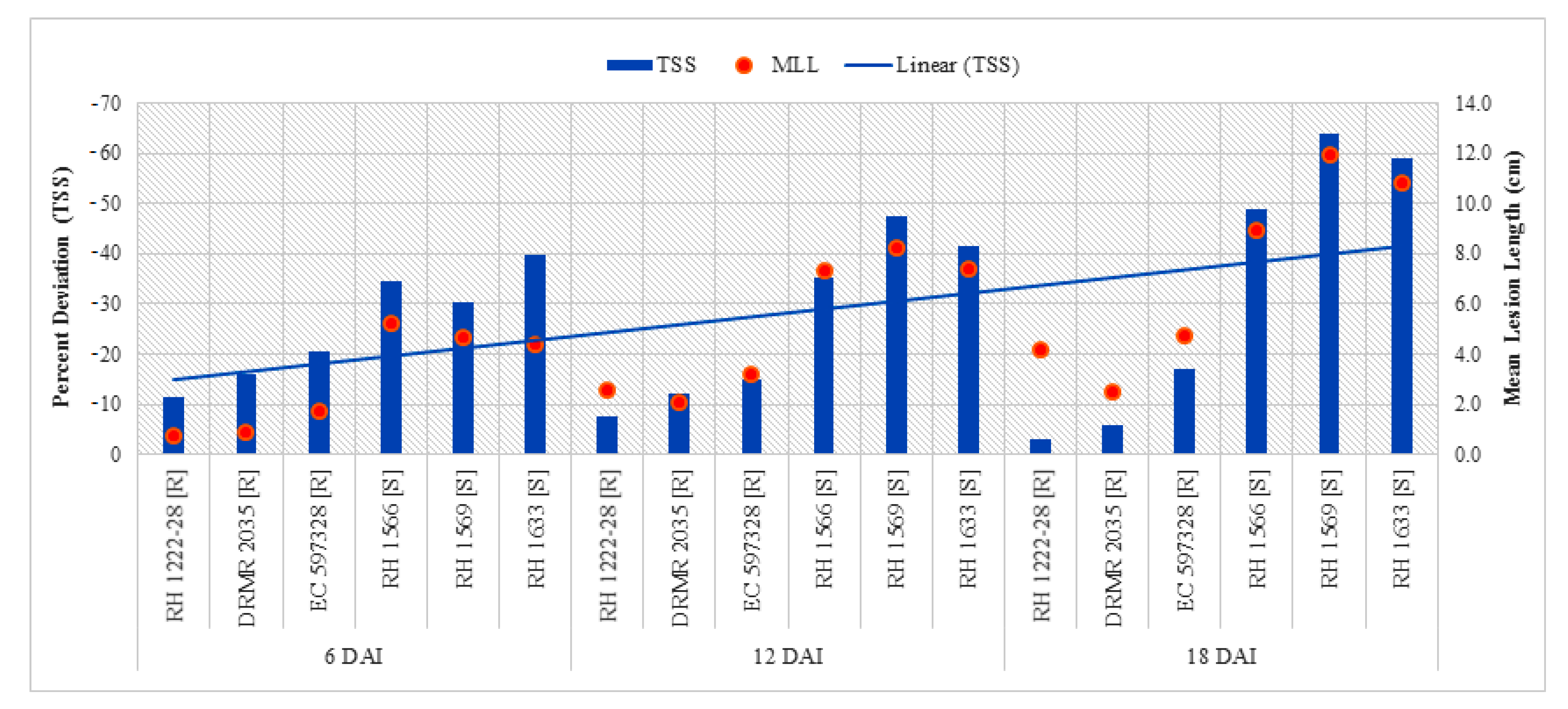
2.2.5. Total Soluble Sugar (TSS)
3. Discussion
4. Materials and Methods
4.1. Plant Materials
4.2. Sclerotinia Sclerotiorum Inoculum Preparation and Disease Assessment
4.3. Sample Collection and Biochemical Analysis
4.4. Determination of Defense-Related Antioxidant Enzymatic Activities
4.5. Quantification of Total Soluble Phenolics (TSP) and Total Soluble Sugar (TSS)
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Renjini, V.R.; Jha, G.K. Oilseeds sector in India: A trade policy perspective. Indian J. Agric. Sci. 2019, 89, 73–80. [Google Scholar]
- Jat, R.S.; Singh, V.V.; Sharma, P.; Rai, P.K. Oilseed brassica in India: Demand, supply, policy perspective and future potential. OCL 2019, 26, 8. [Google Scholar] [CrossRef] [Green Version]
- Chattopadhyay, C.; Kolte, S.J. Diseases. In Brassica Oilseeds: Breeding and Management; Kumar, A., Banga, S.S., Meena, P.D., Kumar, P.R., Eds.; CABI: Croydon, UK, 2015; pp. 115–147. [Google Scholar]
- Rana, K.; Atri, C.; Gupta, M.; Akhatar, J.; Sandhu, P.S.; Kumar, N.; Jaswal, R.; Barbetti, M.J.; Banga, S.S. Mapping resistance responses to Sclerotinia infestation in introgression lines of Brassica juncea carrying genomic segments from wild Brassicaceae B. fruticulosa. Sci. Rep. 2017, 7, 1–12. [Google Scholar] [CrossRef]
- Mahalingam, T.; Chen, W.; Rajapakse, C.S.; Somachandra, K.P.; Attanayake, R.N. Genetic Diversity and Recombination in the Plant Pathogen Sclerotinia sclerotiorum Detected in Sri Lanka. Pathogens 2020, 9, 306. [Google Scholar] [CrossRef] [PubMed]
- Saharan, G.S.; Mehta, N. The disease and symptoms. In Sclerotinia Diseases of Crop Plants: Biology, Ecology and Disease Management, 1st ed.; Springer: Dordrecht, The Netherlands, 2008; pp. 47–70. [Google Scholar]
- Sharma, P.; Samkumar, A.; Rao, M.; Singh, V.V.; Prasad, L.; Mishra, D.C.; Bhattacharya, R.; Gupta, N.C. Genetic diversity studies based on morphological variability, pathogenicity and molecular phylogeny of the Sclerotinia sclerotiorum population from Indian mustard (Brassica juncea). Front. Microbiol. 2018, 9, 1169. [Google Scholar] [CrossRef] [PubMed]
- Xia, S.; Xu, Y.; Hoy, R.; Zhang, J.; Qin, L.; Li, X. The Notorious Soilborne Pathogenic Fungus Sclerotinia sclerotiorum: An Update on Genes Studied with Mutant Analysis. Pathogens 2020, 9, 27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kabbage, M.; Yarden, O.; Dickman, M.B. Pathogenic attributes of Sclerotinia sclerotiorum: Switching from a biotrophic to necrotrophic lifestyle. Plant Sci. 2015, 233, 53–60. [Google Scholar] [CrossRef]
- Hegedus, D.D.; Rimmer, S.R. Sclerotinia sclerotiorum: When “to be or not to be” a pathogen? FEMS Microbiol. Lett. 2005, 251, 177–184. [Google Scholar] [CrossRef] [Green Version]
- Uloth, M.B.; Clode, P.L.; You, M.P.; Barbetti, M.J. Attack modes and defence reactions in pathosystems involving Sclerotinia sclerotiorum, Brassica carinata, B. juncea and B. napus. Ann. Bot. 2016, 117, 79–95. [Google Scholar] [CrossRef] [Green Version]
- Peltier, A.J.; Hatfield, R.D.; Grau, C.R. Soybean stem lignin concentration relates to resistance to Sclerotinia sclerotiorum. Plant Dis. 2009, 93, 149–154. [Google Scholar] [CrossRef] [Green Version]
- Sharma, P.; Jha, A.B.; Dubey, R.S.; Pessarakli, M. Reactive oxygen species, oxidative damage, and antioxidative defense mechanism in plants under stressful conditions. J. Bot. 2012, 2012, 1–26. [Google Scholar] [CrossRef] [Green Version]
- Paiva, C.N.; Bozza, M.T. Are reactive oxygen species always detrimental to pathogens? Antioxid. Redox Signal. 2014, 20, 1000–1037. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, X.; He, Z.; Chen, N.; Tang, Z.; Wang, Q.; Cai, Y. The roles of environmental factors in regulation of oxidative stress in plant. Biomed. Res. Int. 2019, 2019, 1–15. [Google Scholar] [CrossRef]
- Caverzan, A.; Casassola, A.; Brammer, S.P. Reactive oxygen species and antioxidant enzymes involved in plant tolerance to stress. In Abiotic and Biotic Stress in Plants-Recent Advances and Future Perspectives; Shanker, A.K., Shanker, C., Eds.; InTechOpen: Rijeka, Croatia, 2016; pp. 463–480. [Google Scholar]
- Ali, M.; Cheng, Z.; Ahmad, H.; Hayat, S. Reactive oxygen species (ROS) as defenses against a broad range of plant fungal infections and case study on ROS employed by crops against Verticillium dahliae wilts. J. Plant Interact. 2018, 13, 353–363. [Google Scholar] [CrossRef] [Green Version]
- Boeckx, T.; Winters, A.L.; Webb, K.J.; Kingston-Smith, A.H. Polyphenol oxidase in leaves: Is there any significance to the chloroplastic localization? J. Exp. Bot. 2015, 66, 3571–3579. [Google Scholar] [CrossRef] [Green Version]
- Taheri, P.; Kakooee, T. Reactive oxygen species accumulation and homeostasis are involved in plant immunity to an opportunistic fungal pathogen. J. Plant Physiol. 2017, 216, 152–163. [Google Scholar] [CrossRef]
- Couee, I.; Sulmon, C.; Gouesbet, G.; and El Amrani, A. Involvement of soluble sugars in reactive oxygen species balance and responses to oxidative stress in plants. J. Exp. Bot. 2006, 57, 449–459. [Google Scholar] [CrossRef]
- Jiang, S.; Han, S.; He, D.; Cao, G.; Fang, K.; Xiao, X.; Yi, J.; Wan, X. The accumulation of phenolic compounds and increased activities of related enzymes contribute to early defense against walnut blight. Physiol. Mol. Plant Pathol. 2019, 108, 101433. [Google Scholar] [CrossRef]
- Wen, L. Cell death in plant immune response to necrotrophs. J. Plant Biochem. Physiol. 2013, 1, 1–3. [Google Scholar] [CrossRef] [Green Version]
- Garg, H.; Atri, C.; Sandhu, P.S.; Kaur, B.; Renton, M.; Banga, S.K.; Singh, H.; Singh, C.; Barbetti, M.J.; Banga, S.S. High level of resistance to Sclerotinia sclerotiorum in introgression lines derived from hybridization between wild crucifers and the crop Brassica species B. napus and B. juncea. Field Crops Res. 2010, 117, 51–58. [Google Scholar] [CrossRef]
- Andersen, E.J.; Ali, S.; Byamukama, E.; Yen, Y.; Nepal, M.P. Disease resistance mechanisms in plants. Genes 2018, 9, 339. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, J.; Cao, L.; Li, H.; Wang, G.; Wang, S.; Li, F.; Zou, X.; Wang, J. Early responses given distinct tactics to infection of Peronophythora litchii in susceptible and resistant litchi cultivar. Sci. Rep. 2019, 9, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mehta, N. Epidemiology and forecasting for the management of rapeseed-mustard diseases. J. Mycol. Plant Pathol. 2014, 44, 131–147. [Google Scholar]
- Berger, R.D.; Filho, A.B.; Amorim, L. Lesion expansion as an epidemic component. Phytopathology 1997, 87, 1005–1013. [Google Scholar] [CrossRef] [PubMed]
- Sharma, P.; Singh, S.; Singh, V.V.; Gupta, N.C.; Prasad, L.; Meena, P.D.; Ambawat, S.; Rai, P.K. Identification and characterization of new resistance sources against Sclerotinia stem rot (Sclerotinia sclerotiorum) in oilseed brassica. Indian J. Mycol. Plant Pathol. 2018, 48, 35–44. [Google Scholar]
- Uloth, M.B.; You, M.P.; Cawthray, G.; Barbetti, M.J. Temperature adaptation in isolates of Sclerotinia sclerotiorum affects their ability to infect Brassica carinata. Plant Pathol. 2015, 64, 1140–1148. [Google Scholar] [CrossRef]
- Denton-Giles, M.; Derbyshire, M.C.; Khentry, Y.; Buchwaldt, L.; Kamphuis, L.G. Partial stem resistance in Brassica napus to highly aggressive and genetically diverse Sclerotinia sclerotiorum isolates from Australia. Can. J. Plant Pathol. 2018, 40, 551–561. [Google Scholar] [CrossRef] [Green Version]
- Gyawali, S.; Harrington, M.; Durkin, J.; Horner, K.; Parkin, I.A.; Hegedus, D.D.; Bekkaoui, D.; Buchwaldt, L. Microsatellite markers used for genome-wide association mapping of partial resistance to Sclerotinia sclerotiorum in a world collection of Brassica napus. Mol. Breeding 2016, 36, 72. [Google Scholar] [CrossRef] [Green Version]
- McCaghey, M.; Willbur, J.; Smith, D.L.; Kabbage, M. The complexity of the Sclerotinia sclerotiorum pathosystem in soybean: Virulence factors, resistance mechanisms, and their exploitation to control Sclerotinia stem rot. Trop. Plant Pathol. 2019, 44, 12–22. [Google Scholar] [CrossRef]
- Moellers, T.C.; Singh, A.; Zhang, J.; Brungardt, J.; Kabbage, M.; Mueller, D.S.; Grau, C.R.; Ranjan, A.; Smith, D.L.; Chowda-Reddy, R.V.; et al. Main and epistatic loci studies in soybean for Sclerotinia sclerotiorum resistance reveal multiple modes of resistance in multi-environments. Sci. Rep. 2017, 7, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Auclair, J.; Boland, G.J.; Cober, E.; Graef, G.L.; Steadman, J.R.; Zilka, J.; Rajcan, I. Development of a new field inoculation technique to assess partial resistance in soybean to Sclerotinia sclerotiorum. Can. J. Plant Sci. 2004, 84, 57–64. [Google Scholar] [CrossRef] [Green Version]
- Amoozadeh, A.L.; Darvishzadeh, R.; Davar, R.; Mandoulakani, B.A.; Haddadi, P.; Basirnia, A. Quantitative trait loci associated with isolate specific and isolate non-specific partial resistance to Sclerotinia sclerotiorum in sunflower. J. Agr. Sci. Tech. Iran 2015, 17, 213–226. [Google Scholar]
- Davar, R.; Darvishzadeh, R.; Majd, A.; Ghosta, Y.; Sarrafi, A. QTL mapping of partial resistance to basal stem rot in sunflower using recombinant inbred lines. Phytopathol. Mediterr. 2010, 49, 330–341. [Google Scholar]
- Liu, R.; Ding, L.N.; Li, M.; Cao, W.; Wang, Y.K.; Wang, W.J.; Yu, Y.K.; Wang, Z.; Zhu, K.M.; Tan, X.L. Characterization of a Rapeseed Anthocyanin-More Mutant with Enhanced Resistance to Sclerotinia sclerotiorum. J. Plant Growth Regul. 2019, 39, 1–14. [Google Scholar] [CrossRef]
- Liu, Y.; Tikunov, Y.; Schouten, R.E.; Marcelis, L.F.; Visser, R.G.; Bovy, A. Anthocyanin biosynthesis and degradation mechanisms in Solanaceous vegetables: A review. Front. Chem. 2018, 6, 52. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Rothstein, S.J. ROS-Induced anthocyanin production provides feedback protection by scavenging ROS and maintaining photosynthetic capacity in Arabidopsis. Plant Signal. Behav. 2018, 13, 1364–1377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calla, B.; Vuong, T.; Radwan, O.; Hartman, G.L.; Clough, S.J. Gene expression profiling soybean stem tissue early response to Sclerotinia sclerotiorum and in silico mapping in relation to resistance markers. Plant Genome 2009, 2, 149–166. [Google Scholar] [CrossRef] [Green Version]
- Glazebrook, J. Contrasting mechanisms of defense against biotrophic and necrotrophic pathogens. Annu. Rev. Phytopathol. 2005, 43, 205–227. [Google Scholar] [CrossRef]
- Vargas, W.A.; Martín, J.M.S.; Rech, G.E.; Rivera, L.P.; Benito, E.P.; Díaz-Mínguez, J.M.; Thon, M.R.; Sukno, S.A. Plant defense mechanisms are activated during biotrophic and necrotrophic development of Colletotricum graminicola in maize. Plant Physiol. 2012, 158, 1342–1358. [Google Scholar] [CrossRef] [Green Version]
- Almagro, L.; Gómez Ros, L.V.; Belchi-Navarro, S.; Bru, R.; Ros Barceló, A.; Pedreno, M.A. Class III peroxidases in plant defence reactions. J. Exp. Bot. 2009, 60, 377–390. [Google Scholar] [CrossRef] [Green Version]
- Shigeto, J.; Tsutsumi, Y. Diverse functions and reactions of class III peroxidases. New Phytol. 2016, 209, 1395–1402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ranjan, A.; Westrick, N.M.; Jain, S.; Piotrowski, J.S.; Ranjan, M.; Kessens, R.; Stiegman, L.; Grau, C.R.; Conley, S.P.; Smith, D.L.; et al. Resistance against Sclerotinia sclerotiorum in soybean involves a reprogramming of the phenylpropanoid pathway and up-regulation of antifungal activity targeting ergosterol biosynthesis. Plant Biotechnol. J. 2019, 17, 1567–1581. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lv, C.-M.; Zhao, Y.; Zhao, X.; Wang, Q.; Meng, X.-X.; Wei, S.-H.; Han, Y.-P.; Li, W.-B.; Zhang, J.-H. Biochemical resistance mechanism of soybean cultivar Maple Arrow to Sclerotinia sclerotiorum. Chin. J. Oil Crop Sci. 2014, 36, 630. [Google Scholar]
- Leite, M.E.; Dos Santos, J.B.; Ribeiro, P.M.; de Souza, D.A.; de Castro Lara, L.A.; de Resende, M.L.V. Biochemical responses associated with common bean defence against Sclerotinia sclerotiorum. Eur. J. Plant Pathol. 2014, 138, 391–404. [Google Scholar] [CrossRef]
- Monazzah, M.; Tahmasebi Enferadi, S.; Rabiei, Z. Enzymatic activities and pathogenesis-related genes expression in sunflower inbred lines affected by Sclerotinia sclerotiorum culture filtrate. J. Appl. Microbiol. 2018, 125, 227–242. [Google Scholar] [CrossRef] [PubMed]
- Na, R.; Luo, Y.; Bo, H.; Jia, R.; Meng, Q.; Zhou, H.; Hao, J.; Zhao, J. Responses of sunflower induced by Sclerotinia sclerotiorum infection. Physiol. Mol. Plant Pathol. 2018, 102, 113–121. [Google Scholar] [CrossRef]
- Liu, J.; Zhang, Y.; Meng, Q.; Shi, F.; Ma, L.; Li, Y. Physiological and biochemical responses in sunflower leaves infected by Sclerotinia sclerotiorum. Physiol. Mol. Plant Pathol. 2017, 100, 41–48. [Google Scholar] [CrossRef]
- Davar, R.; Darvishzadeh, R.; Majd, A. Changes in antioxidant systems in sunflower partial resistant and susceptible lines as affected by Sclerotinia sclerotiorum. Biologia 2013, 68, 821–829. [Google Scholar] [CrossRef]
- Wen, L.; Tan, T.L.; Shu, J.B.; Chen, Y.; Liu, Y.; Yang, Z.F.; Zhang, Q.P.; Yin, M.Z.; Tao, J.; Guan, C.Y. Using proteomic analysis to find the proteins involved in resistance against Sclerotinia sclerotiorum in adult Brassica napus. Eur. J. Plant Pathol. 2013, 137, 505–523. [Google Scholar] [CrossRef]
- Qi, S.W.; Guan, C.Y.; Liu, C.L. Relationship between some enzyme activity and resistance to Sclerotinia sclerotiorum of rapeseed cultivars. Zuo Wu Xue Bao 2004, 30, 270–273. [Google Scholar]
- Shetty, N.P.; Mehrabi, R.; Lütken, H.; Haldrup, A.; Kema, G.H.; Collinge, D.B.; Jørgensen, H.J.L. Role of hydrogen peroxide during the interaction between the hemibiotrophic fungal pathogen Septoria tritici and wheat. New Phytol. 2007, 174, 637–647. [Google Scholar] [CrossRef] [PubMed]
- Peluffo, L.; Lia, V.; Troglia, C.; Maringolo, C.; Norma, P.; Escande, A.; Hopp, H.E.; Lytovchenko, A.; Fernie, A.R.; Heinz, R.; et al. Metabolic profiles of sunflower genotypes with contrasting response to Sclerotinia sclerotiorum infection. Phytochemistry 2010, 71, 70–80. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Liu, X.; Liu, J.; Wang, H. Changes of protective enzymes activities responding to Sclerotinia sclerotiorum inoculation in non-heading Chinese cabbages. Jiangsu J. Agric. Sci. 2012, 28, 1403–1408. [Google Scholar]
- Taranto, F.; Pasqualone, A.; Mangini, G.; Tripodi, P.; Miazzi, M.M.; Pavan, S.; Montemurro, C. Polyphenol oxidases in crops: Biochemical, physiological and genetic aspects. Int. J. Mol. Sci. 2017, 18, 377. [Google Scholar] [CrossRef]
- Constabel, C.P.; Barbehenn, R. Defensive roles of polyphenol oxidase in plants. In Induced Plant Resistance to Herbivory; Schaller, A., Ed.; Springer: Dordrecht, The Netherlands, 2008; pp. 253–270. [Google Scholar]
- Ni, Y.; Guo, Y.J.; Wang, J.; Xia, R.E.; Wang, X.Q.; Ash, G.; Li, J.N. Responses of physiological indexes and leaf epicuticular waxes of Brassica napus to Sclerotinia sclerotiorum infection. Plant Pathol. 2014, 63, 174–184. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Liu, G.; Zheng, Y.; Wang, X.; Yang, Q. Breeding of the Brassica napus cultivar Zhongshuang 9 with high-resistance to Sclerotinia sclerotiorum and dynamics of its important defense enzyme activity. Sci. Agri. Sin. 2004, 37, 23–28. [Google Scholar]
- Yang, X.; Yang, J.; Li, H.; Niu, L.; Xing, G.; Zhang, Y.; Xu, W.; Zhao, Q.; Li, Q.; Dong, Y. Overexpression of the chitinase gene CmCH1 from Coniothyrium minitans renders enhanced resistance to Sclerotinia sclerotiorum in soybean. Transgenic Res. 2020, 29, 187–198. [Google Scholar] [CrossRef]
- Nicholson, R.L.; Hammerschmidt, R. Phenolic compounds and their role in disease resistance. Annu. Rev. Phytopathol. 1992, 30, 369–389. [Google Scholar] [CrossRef]
- Lin, D.; Xiao, M.; Zhao, J.; Li, Z.; Xing, B.; Li, X.; Kong, M.; Li, L.; Zhang, Q.; Liu, Y.; et al. An overview of plant phenolic compounds and their importance in human nutrition and management of type 2 diabetes. Molecules 2016, 21, 1374. [Google Scholar] [CrossRef]
- Rahaman, M.M.; Zwart, R.S.; Thompson, J.P. Constitutive and Induced Expression of Total Phenol and Phenol Oxidases in Wheat Genotypes Ranging in Resistance/Susceptibility to the Root-Lesion Nematode Pratylenchus thornei. Plants 2020, 9, 485. [Google Scholar] [CrossRef] [Green Version]
- Wallace, G.; Fry, S.C. Phenolic components of the plant cell wall. In International Review of Cytology, 1st ed.; Jeon, K.W., Jarvik, J., Eds.; Elsevier B.V.: Amsterdam, The Netherlands, 1994; Volume 151, pp. 229–267. [Google Scholar]
- Lattanzio, V.; Lattanzio, V.M.; Cardinali, A. Role of phenolics in the resistance mechanisms of plants against fungal pathogens and insects. In Phytochemistry: Advances in Research; Imperato, F., Ed.; Research Signpost: Trivandrum, India, 2006; pp. 23–67. [Google Scholar]
- Skłodowska, M.; Mikiciński, A.; Wielanek, M.; Kuźniak, E.; Sobiczewski, P. Phenolic profiles in apple leaves and the efficacy of selected phenols against fire blight (Erwinia amylovora). Eur. J. Plant Pathol. 2018, 151, 213–228. [Google Scholar] [CrossRef] [Green Version]
- Mandavia, M.K.; Gajera, H.P.; Andharia, J.H.; Khandar, R.R.; Parameswaran, M. Cell wall degrading enzymes in host-pathogen interaction of Fusarium wilt of chickpea: Inhibitory effects of phenolic compounds. Indian Phytopathol. 1999, 52, 285–288. [Google Scholar]
- Sharma, S.; Harmandeep; Soni, G. Interaction of phenolic compounds with pectinases from Sclerotinia sclerotiorum. Indian Phytopath. 2001, 54, 167–170. [Google Scholar]
- Rodríguez, M.A.; Venedikian, N.; Bazzalo, M.E.; Godeas, A. Histopathology of Sclerotinia sclerotiorum attack on flower parts of Helianthus annuus heads in tolerant and susceptible varieties. Mycopathologia 2004, 157, 291–302. [Google Scholar] [CrossRef] [PubMed]
- Prats, E.; Bazzalo, M.E.; León, A.; Jorrín, J.V. Accumulation of soluble phenolic compounds in sunflower capitula correlates with resistance to Sclerotinia sclerotiorum. Euphytica 2003, 132, 321–329. [Google Scholar] [CrossRef] [Green Version]
- Bazzalo, M.E.; Dimarco, P.; Martinez, F.; Daleo, G.R. Indicators of resistance of sunflower plant to basal stalk rot (Sclerotinia sclerotiorum): Symptomatological, biochemical, anatomical, and morphological characters of the host. Euphytica 1991, 57, 195–205. [Google Scholar] [CrossRef]
- El_Komy, M.H.; Saleh, A.A.; Ibrahim, Y.E.; Molan, Y.Y. Early production of reactive oxygen species coupled with an efficient antioxidant system play a role in potato resistance to late blight. Trop. Plant Pathol. 2020, 45, 44–55. [Google Scholar] [CrossRef]
- Hellmann, H.A.; Smeekens, S. Sugar sensing and signaling in plants. Front. Plant Sci. 2014, 5, 113. [Google Scholar]
- Hura, K.; Hura, T.; Dziurka, K.; Dziurka, M. Carbohydrate, phenolic and antioxidant level in relation to chlorophyll a content in oilseed winter rape (Brassica napus L.) inoculated with Leptosphaeria maculans. Eur. J. Plant Pathol. 2015, 143, 291–303. [Google Scholar] [CrossRef] [Green Version]
- Engelsdorf, T.; Horst, R.J.; Pröls, R.; Pröschel, M.; Dietz, F.; Hückelhoven, R.; Voll, L.M. Reduced carbohydrate availability enhances the susceptibility of Arabidopsis toward Colletotrichum higginsianum. Plant Physiol. 2013, 162, 225–238. [Google Scholar] [CrossRef] [Green Version]
- Ponmurugan, P.; Baby, U.I. Morphological, physiological and biochemical changes in resistant and susceptible cultivars of tea in relation to phomopsis disease. Plant Pathol. J. 2007, 6, 91–94. [Google Scholar]
- Bandara, Y.M.A.Y.; Tesso, T.T.; Zhang, K.; Wang, D.; Little, C.R. Charcoal rot and Fusarium stalk rot diseases influence sweet sorghum sugar attributes. Ind. Crops Prod. 2018, 112, 188–195. [Google Scholar] [CrossRef]
- Qi, H.X.; Wang, T.K.; Qi, Y.S.; Zhang, J.Z.; Liu, Y.J.; Li, S.M. Studies on the leaf biochemical and physiological changes of resistant and susceptible grape cultivars infected by Plasmopara viticola J. Int. J. Fruit Sci. 2006, 23, 73–76. [Google Scholar]
- Li, C.X.; Li, H.; Sivasithamparam, K.; Fu, T.D.; Li, Y.C.; Liu, S.Y.; Barbetti, M.J. Expression of field resistance under Western Australian conditions to Sclerotinia sclerotiorum in Chinese and Australian Brassica napus and Brassica juncea germplasm and its relation with stem diameter. Aust. J. Agric. Res. 2006, 57, 1131–1135. [Google Scholar] [CrossRef]
- Sen, A.; Alikamanoglu, S. Antioxidant enzyme activities, malondialdehyde, and total phenolic content of PEG-induced hyperhydric leaves in sugar beet tissue culture. In Vitro Cell. Dev. Biol. 2013, 49, 396–404. [Google Scholar] [CrossRef]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar]
- Maehly, A.C. The assay of catalase and peroxidases. In Methods of Biochemical Analysis; Glick, D., Ed.; Interscience Publishers, Inc.: New York, NY, USA, 1954; Volume 1, pp. 357–424. [Google Scholar]
- Aebi, H. Catalase in vitro. In Methods in Enzymology; Packer, L., Ed.; Academic Press: New York, NY, USA, 1984; Volume 105, pp. 121–126. [Google Scholar]
- Gauillard, F.; Richardforget, F.; Nicolas, J. New spectrophotometric assay for polyphenol oxidase activity. Anal. Biochem. 1993, 215, 59–65. [Google Scholar] [CrossRef]
- Bray, H.G.; Thorpe, W.V. Analysis of phenolic compounds of interest in metabolism. In Methods of Biochemical Analysis; Glick, D., Ed.; Interscience Publishers, Inc.: New York, NY, USA, 1954; Volume 1, pp. 27–52. [Google Scholar]
- Dubois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.T.; Smith, F. Colorimetric method for determination of sugars and related substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Source of Variation | df | p Value |
|---|---|---|
| Lesion length (6 DAI) | 5 | <0.0001 |
| Lesion length (12 DAI) | 5 | <0.0001 |
| Lesion length (18 DAI) | 5 | <0.0001 |
| Genotype | Mean Lesion Length (cm) | Rate of Increase in Lesion Length (cm day-1) | AUDPC * | Disease Response ** | ||||
|---|---|---|---|---|---|---|---|---|
| 6 DAI | 12 DAI | 18 DAI | 6 DAI | 12 DAI | 18 DAI | |||
| RH 1222-28 | 0.72 d | 2.58 b | 4.16 c | 0.12 | 0.31 | 0.26 | 32.28 | Resistant |
| DRMR 2035 | 0.91 d | 2.06 b | 2.48 c | 0.15 | 0.19 | 0.07 | 25.26 | Highly Resistant |
| EC 597328 | 1.70 c | 3.19 b | 4.73 c | 0.28 | 0.25 | 0.26 | 43.53 | Resistant |
| RH 1566 | 5.22 a | 7.30 a | 8.91 b | 0.87 | 0.35 | 0.27 | 101.85 | Susceptible |
| RH 1569 | 4.68 b | 8.24 a | 11.95 a | 0.76 | 0.59 | 0.62 | 113.37 | Highly Susceptible |
| RH 1633 | 4.39 b | 7.40 a | 10.81 ab | 0.73 | 0.50 | 0.57 | 103.17 | Highly Susceptible |
| p Values | ||||||
|---|---|---|---|---|---|---|
| Sources of Variation | df | POX | CAT | PPO | TSP | TSS |
| ST | 2 | <0.0001 | <0.0001 | <0.0001 | 0.0283 | <0.0001 |
| PI | 1 | <0.0001 | <0.0001 | <0.0001 | <0.0001 | <0.0001 |
| G | 5 | 0.0002 | <0.0001 | <0.0001 | <0.0001 | <0.0001 |
| ST X PI | 2 | 0.0002 | NS | NS | NS | NS |
| ST X G | 10 | <0.0001 | NS | 0.0077 | NS | 0.0120 |
| PI X G | 5 | 0.0003 | <0.0001 | NS | 0.0001 | <0.0001 |
| ST X PI X G | 10 | <0.0001 | NS | NS | NS | NS |
| DAI/PI/G | POX µmol min −1 mg −1 Protein | CAT µmol min −1 mg −1 Protein | PPO µmol min −1 mg −1 Protein | TSP mg g −1 DW | TSS mg g −1 DW |
|---|---|---|---|---|---|
| 6DAI/UI/R1 | 2.96 e–g | 6.60 ab | 1.03 a−c | 0.98 b−d | 46.37 c |
| 6DAI/UI/R2 | 2.87 e−g | 7.08 a | 1.38 a−c | 1.00 b−d | 48.30 b |
| 6DAI/UI/R3 | 3.21 d−g | 6.22 a−c | 1.21 a−c | 0.94 b−d | 49.74 a |
| 6DAI/UI/S1 | 3.02 d−g | 5.98 b−d | 0.79 bc | 0.79 cd | 48.60 b |
| 6DAI/UI/S2 | 2.86 e−g | 6.05 b−d | 0.69 bc | 0.71 cd | 46.64 c |
| 6DAI/UI/S3 | 2.98 e−g | 5.84 b−f | 0.59 bc | 0.63 d | 47.82 b |
| 6DAI/I/R1 | 5.32 ab | 5.45 c−h | 1.56 ab | 2.06 a | 41.07 f−h |
| 6DAI/I/R2 | 5.98 a | 6.01 b−d | 1.90 a | 1.68 a−d | 40.62 gh |
| 6DAI/I/R3 | 5.16 ab | 4.93 e−i | 1.60 ab | 1.31 a−d | 39.51 i |
| 6DAI/I/S1 | 3.64 c−e | 3.71 k−m | 0.93 a−c | 0.96 b−d | 31.81 p |
| 6DAI/I/S2 | 3.21 d−g | 2.86 mn | 0.83 bc | 0.95 b−d | 32.50 op |
| 6DAI/I/S3 | 3.11 d−g | 3.11 l−n | 0.66 bc | 0.84 b−d | 28.73 q |
| 12DAI/UI/R1 | 2.63 e−g | 5.56 c−g | 0.94 a−c | 1.09 a−d | 41.34 fg |
| 12DAI/UI/R2 | 2.75 e−g | 5.87 b−e | 1.27 a−c | 1.18 a−d | 40.33 hi |
| 12DAI/UI/R3 | 2.98 e−g | 5.35 c−h | 0.92 a−c | 1.03 b−d | 44.08 d |
| 12DAI/UI/S1 | 2.54 fg | 5.13 d−h | 0.80 bc | 0.91 b−d | 42.50 e |
| 12DAI/UI/S2 | 2.70 e−g | 5.48 c−h | 0.63 bc | 0.78 cd | 40.62 gh |
| 12DAI/UI/S3 | 2.89 e−g | 4.93 e−i | 0.66 bc | 0.75 cd | 41.92 ef |
| 12DAI/I/R1 | 4.97 b | 4.96 e−i | 1.26 a−c | 1.89 ab | 38.16 j |
| 12DAI/I/R2 | 4.49 bc | 5.35 c−h | 1.54 ab | 1.74 a−c | 35.41 l |
| 12DAI/I/R3 | 3.21 d-g | 4.12 i−k | 1.14 a−c | 1.58 a−d | 37.45 j |
| 12DAI/I/S1 | 3.28 d−f | 2.39 no | 0.96 a−c | 1.24 a−d | 27.52 r |
| 12DAI/I/S2 | 3.64 c−e | 2.48 no | 0.70 bc | 1.05 a−d | 21.42 t |
| 12DAI/I/S3 | 3.99 cd | 2.33 no | 0.72 bc | 0.88 b−d | 24.46 s |
| 18DAI/UI/R1 | 2.51 fg | 4.86 f−j | 0.78 bc | 1.17 a−d | 34.35 m |
| 18DAI/UI/R2 | 2.39 fg | 5.31 c−h | 0.96 a−c | 1.23 a−d | 36.61 k |
| 18DAI/UI/R3 | 2.43 fg | 4.67 g−k | 0.67 bc | 1.06 a−d | 40.78 gh |
| 18DAI/UI/S1 | 2.19 g | 4.53 h−k | 0.59 bc | 0.98 b−d | 35.33 l |
| 18DAI/UI/S2 | 2.51 fg | 4.12 i−k | 0.46 c | 0.90 b−d | 34.43 m |
| 18DAI/UI/S3 | 2.30 fg | 4.81 g−j | 0.55 bc | 0.84 b−d | 35.32 l |
| 18DAI/I/R1 | 2.85 e−g | 4.51 h−k | 0.93 a−c | 1.60 a−d | 33.28 no |
| 18DAI/I/R2 | 2.53 fg | 5.12 d−h | 1.21 a−c | 1.49 a−d | 34.43 m |
| 18DAI/I/R3 | 2.75 e−g | 3.89 j−l | 0.89 a−c | 1.36 a−d | 33.87 mn |
| 18DAI/I/S1 | 3.06 d−g | 2.29 no | 0.67 bc | 1.34 a−d | 18.11 u |
| 18DAI/I/S2 | 3.16 d−g | 1.71 op | 0.58 bc | 1.14 a−d | 12.43 w |
| 18DAI/I/S3 | 2.94 e−g | 1.23 p | 0.64 bc | 1.02 b−d | 14.48 v |
| Per Cent Deviation (PD) | Mean Lesion Length (cm) | ||
|---|---|---|---|
| 6 DAI | 12 DAI | 18 DAI | |
| POX | −0.943 * | −0.407 NS | 0.801 NS |
| CAT | 0.999 * | 0.976 * | 0.965 * |
| PPO | −0.862 * | −0.742 NS | −0.436 NS |
| TSP | −0.697 NS | −0.768 NS | −0.045 NS |
| TSS | 0.887 * | 0.972 * | 0.976 * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Singh, M.; Avtar, R.; Pal, A.; Punia, R.; Singh, V.K.; Bishnoi, M.; Singh, A.; Choudhary, R.R.; Mandhania, S. Genotype-Specific Antioxidant Responses and Assessment of Resistance Against Sclerotinia sclerotiorum Causing Sclerotinia Rot in Indian Mustard. Pathogens 2020, 9, 892. https://doi.org/10.3390/pathogens9110892
Singh M, Avtar R, Pal A, Punia R, Singh VK, Bishnoi M, Singh A, Choudhary RR, Mandhania S. Genotype-Specific Antioxidant Responses and Assessment of Resistance Against Sclerotinia sclerotiorum Causing Sclerotinia Rot in Indian Mustard. Pathogens. 2020; 9(11):892. https://doi.org/10.3390/pathogens9110892
Chicago/Turabian StyleSingh, Manjeet, Ram Avtar, Ajay Pal, Rakesh Punia, Vivek K. Singh, Mahavir Bishnoi, Anoop Singh, Raju Ram Choudhary, and Shiwani Mandhania. 2020. "Genotype-Specific Antioxidant Responses and Assessment of Resistance Against Sclerotinia sclerotiorum Causing Sclerotinia Rot in Indian Mustard" Pathogens 9, no. 11: 892. https://doi.org/10.3390/pathogens9110892