High Humidity Causes Abnormalities in the Process of Appressorial Formation of Blumeria graminis f. sp. hordei

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
2.1. Reduction of the Penetration Rate of B. graminis f. sp. hordei under the High Humidity Condition
2.2. Proteinaceous Leakage from the Tip of Appressorial Germ Tube under the High Humidity Condition
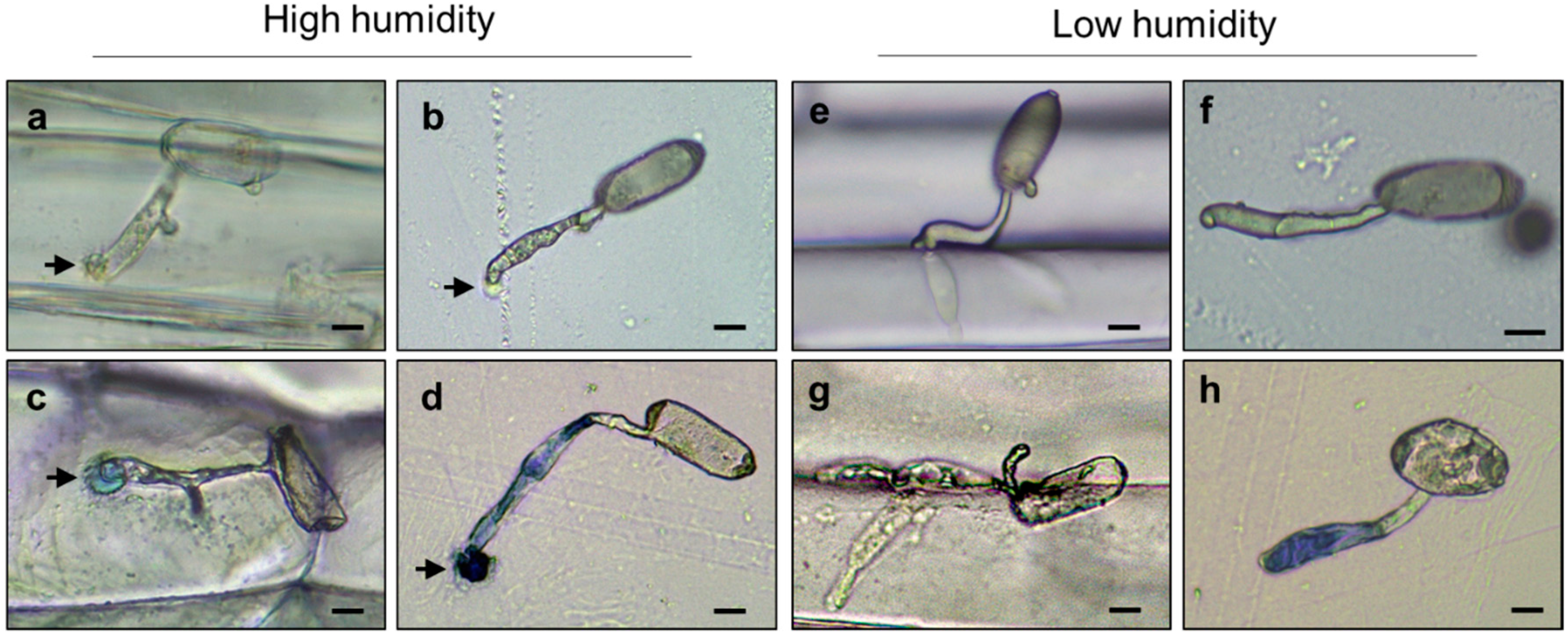
2.3. High Humidity Influences the Formation of the Appressorium
2.4. Conclusions
3. Materials and Methods
3.1. Host Plant and Fungal Material
3.2. Fungal Inoculation
3.3. Histochemical Staining
3.4. Scanning Electron Microscopy Analysis
3.5. Measurement of the Frequency of Conidia with the Leakage and the Length of AGT
Author Contributions
Funding
Conflicts of Interest
References
- Carver, T.L.W.; Bushnell, W.R. The probable role of primary germ tubes in water uptake before infection by Erysiphe graminis. Physiol. Plant Pathol. 1983, 23, 229–240. [Google Scholar] [CrossRef]
- Yamaoka, N.; Matsumoto, I.; Nishiguchi, M. The role of primary germ tubes (PGT) in the life cycle of Blumeria graminis: The stopping of PGT elongation is necessary for the triggering of appressorial germ tube (AGT) emergence. Physiol. Mol. Plant Pathol. 2006, 69, 153–159. [Google Scholar] [CrossRef]
- Yamaoka, N.; Ohta, T.; Danno, N.; Taniguchi, S.; Matsumoto, I.; Nishiguchi, M. The role of primary germ tubes in the life cycle of Blumeria graminis: The primary germ tube is responsible for the suppression of resistance induction of a host plant cell. Physiol. Mol. Plant Pathol. 2007, 71, 184–191. [Google Scholar] [CrossRef]
- Pryce-Jones, E.; Carver, T.; Gurr, S.J. The role of cellulase enzymes and mechanical force in host penetration by Erysiphe graminis f. sp. hordei. Physiol. Mol. Plant Pathol. 1999, 55, 175–182. [Google Scholar] [CrossRef]
- Butt, D. Epidemiology of powdery mildews. In The Powdery Mildews; Spencer, D., Ed.; Academic Press: London, UK, 1978; pp. 51–81. [Google Scholar]
- Carver, T.L.W.; Ingerson-Morris, S.M.; Thomas, B.J.; Gay, A.P. Light-mediated delay of primary haustorium formation by Erysiphe graminis f.sp avenae. Physiol. Mol. Plant Pathol. 1994, 45, 59–79. [Google Scholar] [CrossRef]
- Ward, S.V.; Manners, J.G. Environmental effects on the quantity and viability of conidia produced by Erysiphe graminis. Trans. Br. Mycol. Soc. 1974, 62, 119–128. [Google Scholar] [CrossRef]
- Harris, J.G.; Manners, J.G. Influence of relative humidity on germination and disease development in Erysiphe graminis. Trans. Br. Mycol. Soc. 1983, 81, 605–611. [Google Scholar] [CrossRef]
- Yamaoka, N. Morphogenesis of the powdery mildew fungus in water (1) The effect of liquid on infection process of Erysiphe graminis. Ann. Phytopathol. Soc. Jpn. 1993, 59, 487–491. [Google Scholar] [CrossRef] [Green Version]
- Yamaoka, N.; Toyoda, K.; Kobayashi, I.; Kunoh, H. Induced accessibility and enhanced inaccessibility at the cellular level in barley coleoptiles. XIII. Significance of haustorium formation by the pathogen Erysiphe graminis for induced accessibility to the non-pathogen E. pisi as assessed by nutritional manipulations. Physiol. Mol. Plant Pathol. 1994, 44, 217–225. [Google Scholar]
- Yamaoka, N.; Takeuchi, Y. Morphogenesis of the powdery mildew fungus in water (4) The significance of conidium adhesion to the substratum for normal appressorium development in water. Physiol. Mol. Plant Pathol. 1999, 54, 145–154. [Google Scholar] [CrossRef]
- Kobayashi, I.; Tanaka, C.; Yamaoka, N.; Kunoh, H. Morphogenesis of Erysiphe graminis conidia on artificial membranes. Trans. Mycol. Soc. Jpn. 1991, 32, 187–198. [Google Scholar]
- Suzuki, S.; Komiya, Y.; Mitsui, T.; Tsuyumu, S.; Kunoh, H.; Carver, T.L.W.; Nicholson, R.L. Release of cell wall degrading enzymes from conidia of Blumeria graminis on artificial substrata. Ann. Phytopathol. Soc. Jpn. 1998, 64, 160–167. [Google Scholar] [CrossRef] [Green Version]
- Spanu, P.D.; Abbott, J.C.; Amselem, J.; Burgis, T.A.; Soanes, D.M.; Stüber, K.; Ver Loren Van Themaat, E.; Brown, J.K.; Butcher, S.A.; Gurr, S.J.; et al. Genome expansion and gene loss in powdery mildew fungi reveal tradeoffs in extreme parasitism. Science 2010, 330, 1543–1546. [Google Scholar] [CrossRef] [PubMed]
- Frantzeskakis, L.; Kracher, B.; Kusch, S.; Yoshikawa-Maekawa, M.; Bauer, S.; Pedersen, C.; Spanu, P.D.; Maekawa, T.; Schulze-Lefert, P.; Panstruga, R. Signatures of host specialization and a recent transposable element burst in the dynamic one-speed genome of the fungal barley powdery mildew pathogen. BMC Genom. 2018, 19, 381. [Google Scholar] [CrossRef] [Green Version]
- Bindschedler, L.V.; Burgis, T.A.; Mills, D.J.S.; Ho, J.T.C.; Cramer, R.; Spanu, P.D. In planta proteomics and proteogenomics of the biotrophic barley fungal pathogen Blumeria graminis f. sp. hordei. Mol. Cell Proteom. 2009, 8, 2368–2381. [Google Scholar] [CrossRef] [Green Version]
- Godfrey, D.; Zhang, Z.; Saalbach, G.; Thordal-Christensen, H.A. A proteomics study of barley powdery mildew haustoria. Proteomics 2009, 9, 3222–3232. [Google Scholar] [CrossRef]
- Pedersen, C.; Van Themaat, E.V.L.; McGuffin, L.J.; Abbott, J.C.; Burgis, T.A.; Barton, G.; Bindschedler, L.V.; Lu, X.; Maekawa, T.; Weßling, R.; et al. Structure and evolution of barley powdery mildew effector candidates. BMC Genom. 2012, 13, 694. [Google Scholar] [CrossRef] [Green Version]
- Wahara, M.; Inoue, C.; Kohguchi, T.; Sugai, K.; Kobayashi, K.; Nishiguchi, M.; Yamaoka, N.; Yaeno, T. Improved method for in situ biolistic transformation to analyze barley-powdery mildew interactions. J. Gen. Plant Pathol. 2017, 83, 140–146. [Google Scholar] [CrossRef]
- Yamaoka, N.; Yoshida, S.; Motoyama, E.; Takeuchi, Y.; Takada, Y.; Fukunaga, N. Resistance induction in barley coleoptile cells by intracellular pH decline. Plant Cell Physiol. 2000, 41, 1321–1326. [Google Scholar] [CrossRef] [Green Version]
- Sugai, K.; Masaoka, H.; Penjore, K.; Hanboonsong, Y.; Nishiguchi, M.; Yamaoka, N. The time and spatial strategy of Blumeria graminis f. sp. hordei for surviving after failure of first infection. Physiol. Mol. Plant Pathol. 2010, 74, 346–350. [Google Scholar] [CrossRef]
- Yaeno, T.; Matsuda, O.; Iba, K. Role of chloroplast trienoic fatty acids in plant disease responses. Plant J. 2004, 40, 931–941. [Google Scholar] [CrossRef] [PubMed]




© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sugai, K.; Inoue, H.; Inoue, C.; Sato, M.; Wakazaki, M.; Kobayashi, K.; Nishiguchi, M.; Toyooka, K.; Yamaoka, N.; Yaeno, T. High Humidity Causes Abnormalities in the Process of Appressorial Formation of Blumeria graminis f. sp. hordei. Pathogens 2020, 9, 45. https://doi.org/10.3390/pathogens9010045
Sugai K, Inoue H, Inoue C, Sato M, Wakazaki M, Kobayashi K, Nishiguchi M, Toyooka K, Yamaoka N, Yaeno T. High Humidity Causes Abnormalities in the Process of Appressorial Formation of Blumeria graminis f. sp. hordei. Pathogens. 2020; 9(1):45. https://doi.org/10.3390/pathogens9010045
Chicago/Turabian StyleSugai, Koreyuki, Hiroshi Inoue, Chie Inoue, Mayuko Sato, Mayumi Wakazaki, Kappei Kobayashi, Masamichi Nishiguchi, Kiminori Toyooka, Naoto Yamaoka, and Takashi Yaeno. 2020. "High Humidity Causes Abnormalities in the Process of Appressorial Formation of Blumeria graminis f. sp. hordei" Pathogens 9, no. 1: 45. https://doi.org/10.3390/pathogens9010045