The Genetic Polymorphisms of 24 Base Pair Duplication and Point G102S of Human Chitotriosidase to Bancroftian Filariasis at the Thai–Myanmar Border

Abstract
:1. Introduction
2. Results
2.1. Demographic and Pathology Information of Study Participants
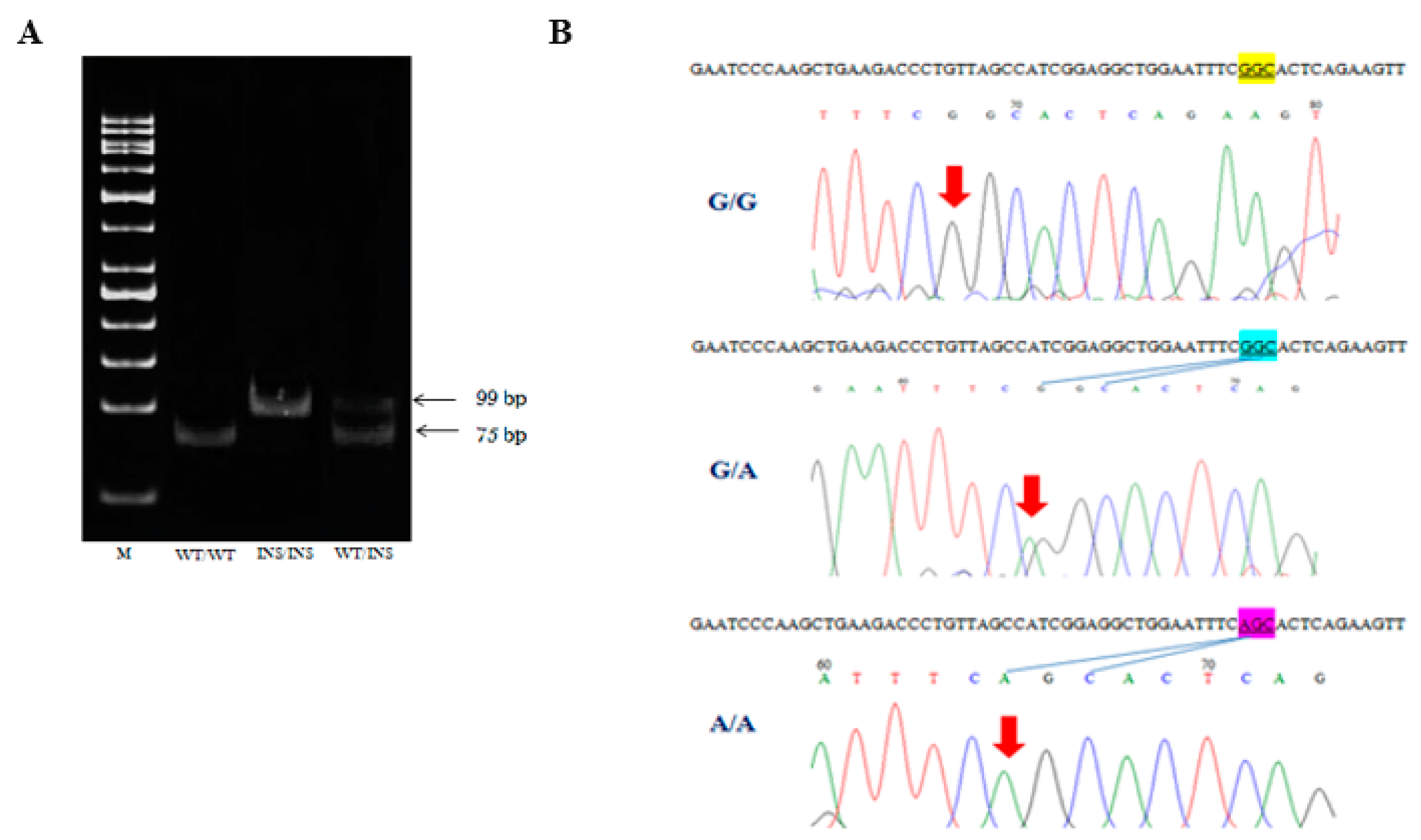
2.2. Genotyping of CHIT1 Polymorphisms: 24 bp Duplication and p. G102S
2.3. Association between CHIT1 Polymorphisms and Susceptibility to Bancroftian Filariasis in Thai and Myanmar Populations
2.3.1. Genotype Frequencies of CHIT1 Polymorphisms in BF Patients and EN
2.3.2. Allele Frequencies of CHIT1 Polymorphisms in BF Patients and EN
3. Discussion
4. Materials and Methods
4.1. Study Areas and Populations
4.2. DNA Extraction from Blood Samples
4.3. Genotyping for CHIT1 Polymorphisms
4.4. Statistical Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Nuchprayoon, S. DNA-based diagnosis of lymphatic filariasis. Southeast Asian J. Trop. Med. Public Health 2009, 40, 904–913. [Google Scholar]
- Das, P.K.; Manoharan, A.; Srividya, A.; Grenfell, B.T.; Bundy, D.A.; Vanamail, P. Frequency distribution of Wuchereria bancrofti microfilariae in human populations and its relationships with age and sex. Parasitology 1990, 101 Pt 3, 429–434. [Google Scholar] [CrossRef]
- Subrahmanyam, D.; Mehta, K.; Nelson, D.S.; Rao, Y.V.; Rao, C.K. Immune reactions in human filariasis. J. Clin. Microbiol. 1978, 8, 228–232. [Google Scholar]
- Tisch, D.J.; Hazlett, F.E.; Kastens, W.; Alpers, M.P.; Bockarie, M.J.; Kazura, J.W. Ecologic and biologic determinants of filarial antigenemia in bancroftian filariasis in Papua New Guinea. J. Infect. Dis. 2001, 184, 898–904. [Google Scholar] [CrossRef]
- Chan, S.H.; Dissanayake, S.; Mak, J.W.; Ismail, M.M.; Wee, G.B.; Srinivasan, N.; Soo, B.H.; Zaman, V. HLA and filariasis in Sri Lankans and Indians. Southeast Asian J. Trop. Med. Public Health 1984, 15, 281–286. [Google Scholar]
- Choi, E.H.; Zimmerman, P.A.; Foster, C.B.; Zhu, S.; Kumaraswami, V.; Nutman, T.B.; Chanock, S.J. Genetic polymorphisms in molecules of innate immunity and susceptibility to infection with Wuchereria bancrofti in South India. Genes Immun. 2001, 2, 248–253. [Google Scholar] [CrossRef] [PubMed]
- Junpee, A.; Tencomnao, T.; Sanprasert, V.; Nuchprayoon, S. Association between Toll-like receptor 2 (TLR2) polymorphisms and asymptomatic bancroftian filariasis. Parasitol. Res. 2010, 107, 807–816. [Google Scholar] [CrossRef] [PubMed]
- Meyrowitsch, D.W.; Simonsen, P.E.; Garred, P.; Dalgaard, M.; Magesa, S.M.; Alifrangis, M. Association between mannose-binding lectin polymorphisms and Wuchereria bancrofti infection in two communities in North-Eastern Tanzania. Am. J. Trop. Med. Hyg. 2010, 82, 115–120. [Google Scholar] [CrossRef] [PubMed]
- Panda, A.K.; Sahoo, P.K.; Kerketta, A.S.; Kar, S.K.; Ravindran, B.; Satapathy, A.K. Human lymphatic filariasis: Genetic polymorphism of endothelin-1 and tumor necrosis factor receptor II correlates with development of chronic disease. J. Infect. Dis. 2011, 204, 315–322. [Google Scholar] [CrossRef] [PubMed]
- Venugopal, G.; O’Regan, N.L.; Babu, S.; Schumann, R.R.; Srikantam, A.; Merle, R.; Hartmann, S.; Steinfelder, S. Association of a PD-L2 Gene Polymorphism with Chronic Lymphatic Filariasis in a South Indian Cohort. Am. J. Trop. Med. Hyg. 2018. [Google Scholar] [CrossRef] [PubMed]
- Harris, M.T.; Lai, K.; Arnold, K.; Martinez, H.F.; Specht, C.A.; Fuhrman, J.A. Chitin synthase in the filarial parasite, Brugia malayi. Mol. Biochem. Parasitol. 2000, 111, 351–362. [Google Scholar] [CrossRef]
- Fuhrman, J.A.; Piessens, W.F. Chitin synthesis and sheath morphogenesis in Brugia malayi microfilariae. Mol. Biochem. Parasitol. 1985, 17, 93–104. [Google Scholar] [CrossRef]
- Di Rosa, M.; Malaguarnera, G.; De Gregorio, C.; Drago, F.; Malaguarnera, L. Evaluation of CHI3L-1 and CHIT-1 expression in differentiated and polarized macrophages. Inflammation 2013, 36, 482–492. [Google Scholar] [CrossRef]
- Kanneganti, M.; Kamba, A.; Mizoguchi, E. Role of chitotriosidase (chitinase 1) under normal and disease conditions. J. Epithel. Biol. Pharmacol. 2012, 5, 1–9. [Google Scholar] [CrossRef]
- Labadaridis, I.; Dimitriou, E.; Theodorakis, M.; Kafalidis, G.; Velegraki, A.; Michelakakis, H. Chitotriosidase in neonates with fungal and bacterial infections. Arch. Dis. Child. Fetal Neonatal Ed. 2005, 90, F531–F532. [Google Scholar] [CrossRef]
- Lehrnbecher, T.; Bernig, T.; Hanisch, M.; Koehl, U.; Behl, M.; Reinhardt, D.; Creutzig, U.; Klingebiel, T.; Chanock, S.J.; Schwabe, D. Common genetic variants in the interleukin-6 and chitotriosidase genes are associated with the risk for serious infection in children undergoing therapy for acute myeloid leukemia. Leukemia 2005, 19, 1745–1750. [Google Scholar] [CrossRef]
- Verwer, P.E.; Notenboom, C.C.; Eadie, K.; Fahal, A.H.; Verbrugh, H.A.; van de Sande, W.W. A Polymorphism in the Chitotriosidase Gene Associated with Risk of Mycetoma Due to Madurella mycetomatis Mycetoma--A Retrospective Study. PLoS Negl. Trop. Dis. 2015, 9, e0004061. [Google Scholar] [CrossRef] [PubMed]
- Lee, P.; Waalen, J.; Crain, K.; Smargon, A.; Beutler, E. Human chitotriosidase polymorphisms G354R and A442V associated with reduced enzyme activity. Blood Cells Mol. Dis. 2007, 39, 353–360. [Google Scholar] [CrossRef] [PubMed]
- Pant, S.; Agarwal, J.; Goel, A.; Gangwar, P.K.; Waseem, M.; Gupta, P.; Sankhwar, S.N.; Purkait, B. Does 24bp Duplication of Human CHIT1 Gene (Chitotriosidase1) Predispose to Filarial Chyluria? A Case-Control Study. J. Clin. Diagn. Res. 2016, 10, PC01–PC04. [Google Scholar] [CrossRef] [PubMed]
- Hise, A.G.; Hazlett, F.E.; Bockarie, M.J.; Zimmerman, P.A.; Tisch, D.J.; Kazura, J.W. Polymorphisms of innate immunity genes and susceptibility to lymphatic filariasis. Genes Immun. 2003, 4, 524–527. [Google Scholar] [CrossRef]
- Boot, R.G.; Renkema, G.H.; Verhoek, M.; Strijland, A.; Bliek, J.; de Meulemeester, T.M.; Mannens, M.M.; Aerts, J.M. The human chitotriosidase gene. Nature of inherited enzyme deficiency. J. Biol. Chem. 1998, 273, 25680–25685. [Google Scholar] [CrossRef]
- Woo, K.H.; Lee, B.H.; Heo, S.H.; Kim, J.M.; Kim, G.H.; Kim, Y.M.; Kim, J.H.; Choi, I.H.; Yang, S.H.; Yoo, H.W. Allele frequency of a 24 bp duplication in exon 10 of the CHIT1 gene in the general Korean population and in Korean patients with Gaucher disease. J. Hum. Genet. 2014, 59, 276–279. [Google Scholar] [CrossRef] [PubMed]
- Arndt, S.; Hobbs, A.; Sinclaire, I.; Lane, A.B. Chitotriosidase deficiency: A mutation update in an african population. JIMD Rep. 2013, 10, 11–16. [Google Scholar]
- Hall, A.J.; Quinnell, R.J.; Raiko, A.; Lagog, M.; Siba, P.; Morroll, S.; Falcone, F.H. Chitotriosidase deficiency is not associated with human hookworm infection in a Papua New Guinean population. Infect. Genet. Evol. 2007, 7, 743–747. [Google Scholar] [CrossRef] [PubMed]
- Sinha, S.; Singh, J.; Jindal, S.K.; Birbian, N.; Singla, N. Association of 24 bp duplication of human CHIT1 gene with asthma in a heterozygous population of north India: A case-control study. Lung 2014, 192, 685–691. [Google Scholar] [CrossRef] [PubMed]
- Bierbaum, S.; Superti-Furga, A.; Heinzmann, A. Genetic polymorphisms of chitotriosidase in Caucasian children with bronchial Asthma. Int. J. Immunogenet. 2006, 33, 201–204. [Google Scholar] [CrossRef] [PubMed]
- Malaguarnera, L. Chitotriosidase: The yin and yang. Cell Mol. Life Sci. 2006, 63, 3018–3029. [Google Scholar] [CrossRef]
- Bierbaum, S.; Nickel, R.; Koch, A.; Lau, S.; Deichmann, K.A.; Wahn, U.; Superti-Furga, A.; Heinzmann, A. Polymorphisms and haplotypes of acid mammalian chitinase are associated with bronchial asthma. Am. J. Respir. Crit. Care Med. 2005, 172, 1505–1509. [Google Scholar] [CrossRef]


{kind=link}
| Polymorphism | SNP ID | SNP Influence | Primer Sequences |
|---|---|---|---|
| 24 bp duplication | Rs 772902435 | Associated with an abnormal mRNA splicing of CHIT1 and results in the production of inactive truncated CHIT1 [21] | F 5′-AGC TAT CTG AAG CAG AAG-3′ R 5′-GGA GGA GCC GGC AAA GTC-3′ [6] |
| p. G102S | Rs 750476665 | Associated with the substitution of glycine (Gly) to serine (Ser) at the amino acid position 102 of CHIT1, resulting in decreasing of CHIT1 enzymatic activity and specificity [21] | F 5′-GGC AGC TGG CAG AGT AAA TCC-3′ R 5′-CCC AGA AGG AAA TTC AGC CC-3′ |
| Characteristics | Bancroftian Filariasis Patients | Endemic Normal |
|---|---|---|
| Gender (M/F) | 27/24 | 19/18 |
| Median age (range) | 30.6 (16–61) | 31.8 (18–68) |
| Mean age (±SD) | 29.9 ± 12.5 | 30.8 ± 12.9 |
| Treatment | No | No |
| Bed net usage | Yes | Yes |
| Pathology | None | None |
| Ag Test (ICT card test and Og4C3 ELISA) | All positive | All negative |
| W. bancrofti circulating antigen levels (U/mL, range) | 29,986 (133–71,339) | <32 (<32) |
| Genotype | Number of Endemic Normal (%) | Number of Bancroftian Filariasis Patients (%) | p Value | |
|---|---|---|---|---|
| 24 bp duplication | WT/WT | 5 (11.1%) | 6 (17.2%) | 0.623 |
| WT/INS | 22 (48.9%) | 18 (51.4%) | ||
| INS/INS | 18 (40.0%) | 11 (31.4%) | ||
| Total | 45 (100.0%) | 35 (100.0%) | ||
| p. G102S | G/G | 15 (40.5%) | 22 (43.1%) | 0.876 |
| G/A | 15 (40.5%) | 18 (35.3%) | ||
| A/A | 7 (19.0%) | 11 (21.6%) | ||
| Total | 37 (100.0%) | 51 (100.0%) | ||
| Genotype | Number of Endemic Normal (%) | Number of Bancroftian Filariasis Patients (%) | p Value | |
|---|---|---|---|---|
| 24 bp duplication | WT | 32 (35.6%) | 30 (42.9%) | 0.347 |
| INS | 58 (64.4%) | 40 (57.1%) | ||
| Total | 90 (100.0%) | 70 (100.0%) | ||
| p. G102S | G | 45 (60.8%) | 62 (60.8%) | 0.997 |
| A | 29 (39.2%) | 40 (39.2%) | ||
| Total | 74 (100.0%) | 102 (100.0%) | ||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sanprasert, V.; Charuchaibovorn, S.; Nuchprayoon, S. The Genetic Polymorphisms of 24 Base Pair Duplication and Point G102S of Human Chitotriosidase to Bancroftian Filariasis at the Thai–Myanmar Border. Pathogens 2019, 8, 41. https://doi.org/10.3390/pathogens8010041
Sanprasert V, Charuchaibovorn S, Nuchprayoon S. The Genetic Polymorphisms of 24 Base Pair Duplication and Point G102S of Human Chitotriosidase to Bancroftian Filariasis at the Thai–Myanmar Border. Pathogens. 2019; 8(1):41. https://doi.org/10.3390/pathogens8010041
Chicago/Turabian StyleSanprasert, Vivornpun, Sarit Charuchaibovorn, and Surang Nuchprayoon. 2019. "The Genetic Polymorphisms of 24 Base Pair Duplication and Point G102S of Human Chitotriosidase to Bancroftian Filariasis at the Thai–Myanmar Border" Pathogens 8, no. 1: 41. https://doi.org/10.3390/pathogens8010041