
An Entomological Investigation during a Recent Rift Valley Fever Epizootic/Epidemic Reveals New Aspects of the Vectorial Transmission of the Virus in Madagascar
 , , ,
, , ,  , , , , and
, , , , and
Abstract
:1. Introduction
2. Materials and Methods
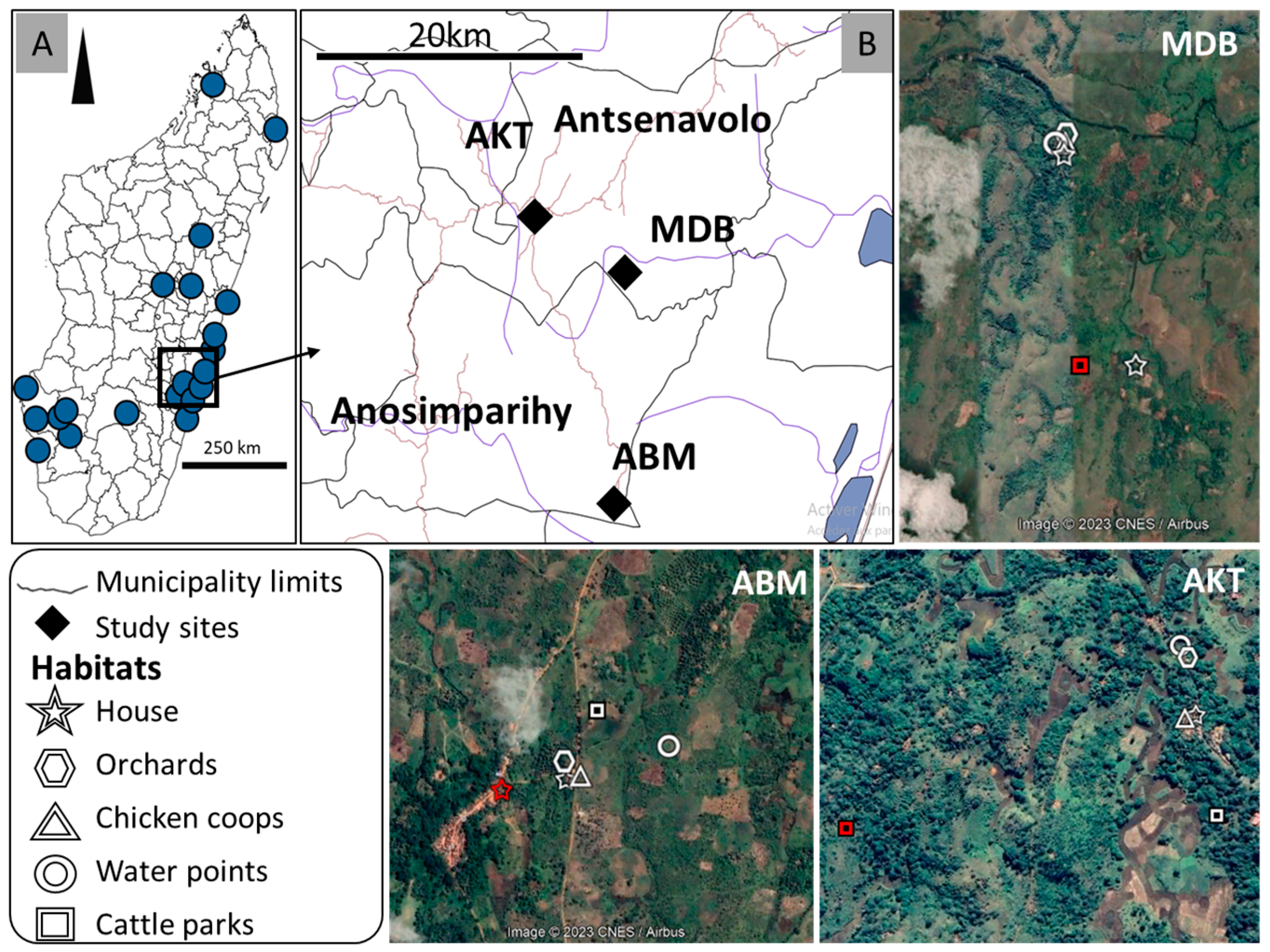
2.1. Study Sites
2.2. Mosquito Sampling
2.3. Mosquito Identification and Storing
2.4. Blood Meal Identification
2.5. RVFV Detection in Mosquitoes
2.6. Statistical Analysis
3. Results
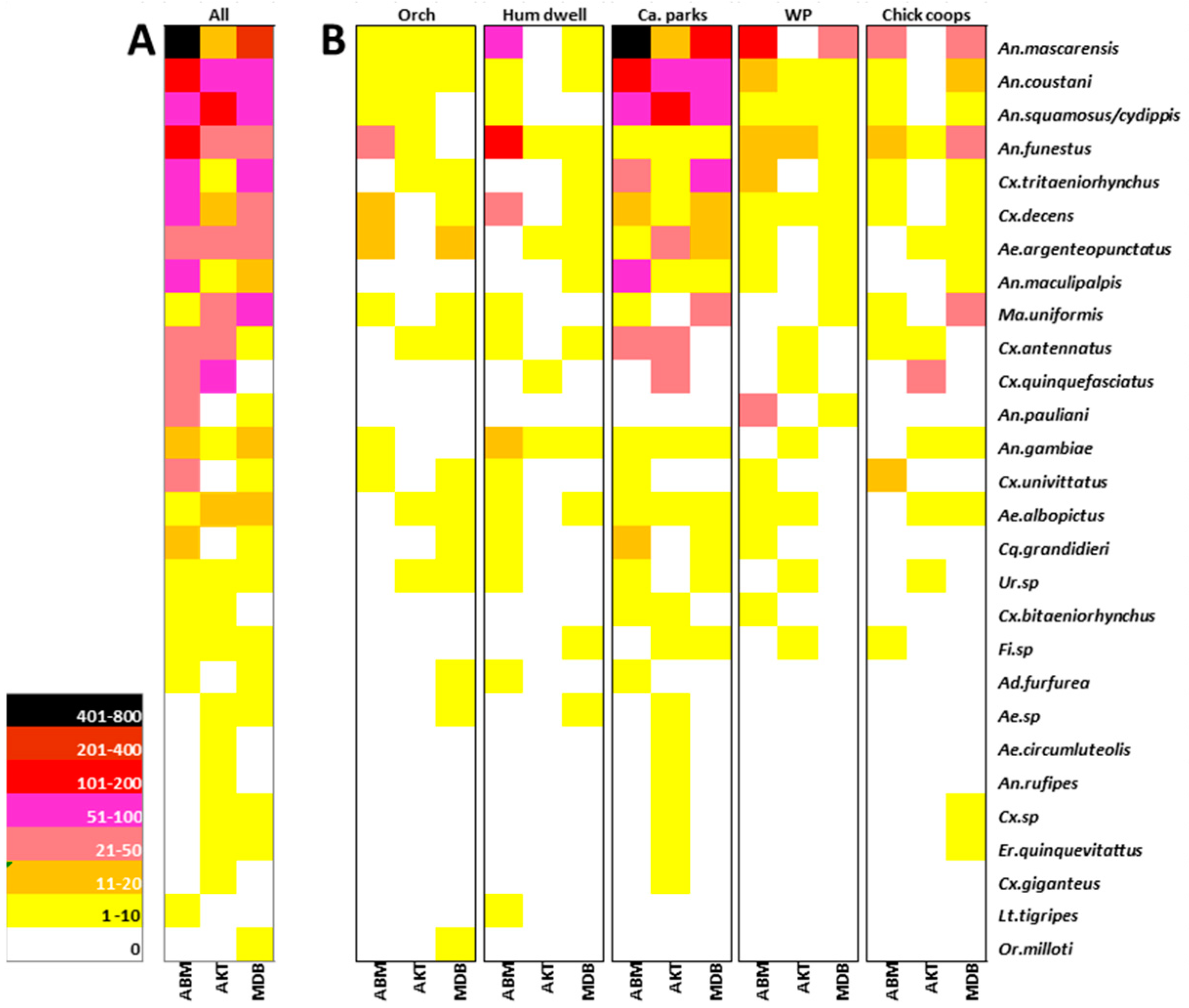
3.1. Mosquito Species Distribution and Abundance
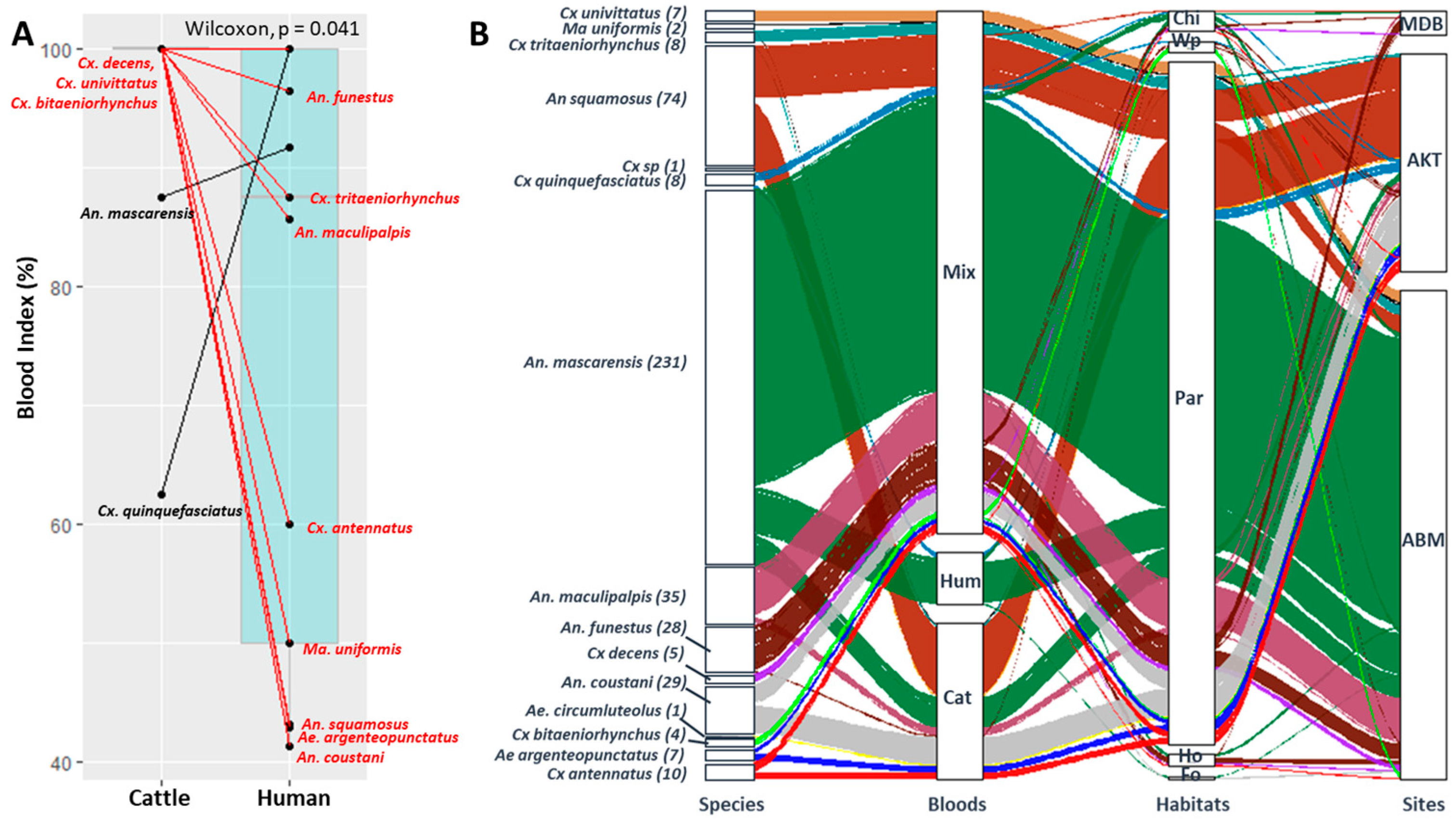
3.2. Blood Meal Analysis
3.3. Detection of RVFV in Mosquitoes
3.4. RVFV Detection and Blood Meal Sources
4. Discussion
4.1. Species Diversity and Abundance
4.2. Trophic Behavior
4.3. Virus Detection
4.4. Implications for RVFV Transmission to Humans and Animals and Recommendations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Daubney, R.; Hudson, J.R.; Garnham, P.C. Enzootic hepatitis or Rift Valley fever, an undescribed virus disease of sheep, cattle and man from East Africa. J. Pathol. Bacteriol. 1931, 34, 545–579. [Google Scholar] [CrossRef]
- Flick, R.; Bouloy, M. Rift Valley Fever Virus. Curr. Mol. Med. 2005, 5, 827–834. [Google Scholar] [CrossRef] [PubMed]
- Swanepoel, R.; Coetzer, J.A.W. Rift Valley fever. Synonyms, Enzootic hepatitis, Slenkdalkoors (Afrik). In Infectious Diseases of Livestock with Special Reference to Southern Africa; Coetzer, J.A.W., Thomson, G.R., Tustin, R.C., Eds.; Oxford University Press: Cape Town, South Africa, 1994. [Google Scholar]
- Turell, M.J.; Presley, S.M.; Gad, A.M.; Cope, S.E.; Dohm, D.J.; Morrill, J.C.; Arthur, R.R. Vector competence of egyptian mosquitoes for Rift Valley fever virus. Am. J. Trop. Med. Hyg. 1996, 54, 136–139. [Google Scholar] [CrossRef] [PubMed]
- Linthicum, K.J.; Davies, F.G.; Kairo, A. Rift Valley fever virus (family Bunyviridae, genus Phlebovirus). Isolations from diptera collected during an inter-epizootic period in Kenya. J. Hyg. 1985, 95, 197–209. [Google Scholar] [CrossRef] [PubMed]
- Lefèvre, P.C.; Blancou, J.; Chermette, R. Principales maladies infectieuses et parasitaires du bétail, Europe et régions chaudes, Tec and Doc, Paris. Bull. De L’académie Vétérinaire De Fr. 2003, 1824, 643–657. [Google Scholar]
- Evans, A.; Gakuya, F.; Paweska, J.T.; Rostal, M.; Akoolo, L.; Van Vuren, P.J.; Manyibe, T.; Macharia, J.M.; Ksiazek, T.G.; Feikin, D.R.; et al. Prevalence of antibodies against Rift Valley fever virus in Kenyan wildlife. Epidemiol. Infect. 2008, 136, 261–269. [Google Scholar] [CrossRef] [PubMed]
- Easterday, B.C.; Murphy, L.C.; Bennett, D.G. Experimental Rift Valley fever in calves, goats, and pigs. Am. J. Vet. Res. 1962, 23, 1224–1230. [Google Scholar]
- Swanepoel, R.; Coetzer, J.A.W. Rift Valley fever. In Infectious Diseases of Livestock, 2nd ed.; Coetzer, J.A., Tustin, R.C., Eds.; Oxford university Press: Cape Town, Southern Africa, 2004; pp. 1037–1070. [Google Scholar]
- Rissmann, M.; Kley, N.; Ulrich, R.; Stoek, F.; Balkema-Buschmann, A.; Eiden, M.; Groschup, M.H. Competency of Amphibians and Reptiles and Their Potential Role as Reservoir Hosts for Rift Valley Fever Virus. Viruses 2020, 12, 1206. [Google Scholar] [CrossRef]
- Smithburn, K.C.; Haddow, A.J.; Gillett, J.D. Rift Valley fever, isolation of the virus from wild mosquitoes. Br. J. Exp. Pathol. 1948, 29, 107–121. [Google Scholar]
- AFSSA. Risque de Propagation de la Fièvre de la Vallée du Rift FVR dans l’Océan Indien La Réunion et Mayotte; AFSSA: Maisons-Alfort, France, 2008. [Google Scholar]
- CDC. 2000 Update, Outbreak of Rift Valley Fever. Saudi Arabia, August–November 2000. MMWR Morb. Mortal. Wkly Rep. 2000, 49, 982–985. [Google Scholar]
- Morvan, J.; Fontenille, D.; Sulazzo, J.-F.; Coulanges, P. Possible Rift Valley Fever outbreak in man and cattle in Madagascar. Trans. R. Soc. Trop. Med. Hyg. 1991, 85, 108. [Google Scholar] [CrossRef]
- Roger, M.; Girard, S.; Faharoudine, A.; Halifa, M.; Bouloy, M.; Cetre-Sossah, C.; Cardinale, E. Rift Valley fever in ruminants, republic of Comoros, 2009. Emerg. Infect. Dis. 2011, 17, 1319–1320. [Google Scholar] [CrossRef] [PubMed]
- Sissoko, D.; Giry, C.; Gabrie, P.; Tarantola, A.; Pettinelli, F.; Collet, L.; d’Ortenzio, E.; Renault, P.; Pierre, V. Rift Valley Fever, Mayotte, 2007–2008. Emerg. Infect. Dis. 2009, 15, 568–570. [Google Scholar] [CrossRef] [PubMed]
- Tantely, M.L.; Boyer, S.; Fontenille, D. A review of mosquitoes associated with Rift Valley fever virus in Madagascar. Am. J. Trop. Med. Hyg. 2015, 92, 722–729. [Google Scholar] [CrossRef] [PubMed]
- Davies, F.G.; Linthicum, K.J.; James, A.D. Rainfall and epizootic Rift valley Fever. Bull. World Health Organ. 1985, 63, 941–943. [Google Scholar] [PubMed]
- Zeller, H.G.; Fontenille, D.; Traore-Lamizana, M.; Thiongane, Y.; Digoutte, J.P. Enzootic activity of Rift Valley fever virus in Senegal. Am. J. Trop. Med. Hyg. 1997, 56, 265–272. [Google Scholar] [CrossRef]
- Chevalier, V.; De La Rocque, S.; Baldet, T.; Vial, L.; Roger, F. Epidemiological processes involved in the emergence of vector borne diseases West Nile Fever, Rift Valley Fever, Japanese Encephalitis and Crimean-Congo Fever. Rev. Sci. Tech. Off. Int. Epiz. 2004, 23, 535–555. [Google Scholar] [CrossRef] [PubMed]
- Davies, F.G. Rift Valley Fever and Livestock Trade from the Horn of Africa (Ethiopia, Somalia, Kenya); FAO: Rome, Italy, 1998; pp. 1–28. [Google Scholar]
- Clerc, Y.; Coulanges, P.; Rodhain, F.; Ricklin, B.; Ranaivosata, J.; Albignac, R. Le programme arbovirus de l’Institut Pasteur de Madagascar, Bilan actuel. Arch. Inst. Pasteur Mad. 1981, 48, 247–257. [Google Scholar]
- Fontenille, D. Arbovirus transmission cycles in Madagascar. Arch. Inst. Pasteur Madag. 1989, 55, 1–317. [Google Scholar]
- Morvan, J.; Saluzzo, J.F.; Fontenille, D.; Rollin, P.E.; Coulanges, P. Rift Valley fever on the east coast of Madagascar. Res. Virol. 1991, 142, 475–482. [Google Scholar] [CrossRef]
- Morvan, J.; Rollin, P.E.; Laventure, S.; Rakotoarivony, I.; Roux, J. Rift Valley fever epizootic in Central Highlands of Madagascar. Res. Virol. 1992, 143, 407–415. [Google Scholar] [CrossRef]
- Zeller, H.G.; Roux, J. Surveillance de la Fièvre de la Vallée du Rift à Madagascar; Rapport Final du Projet Janvier 1996 mars 1998; Institut Pasteur de Madagascar: Antananarivo, Madagascar, 1998; p. 18. [Google Scholar]
- WOAH. 25th Conference of WOAH. Analysis of the Animal Health Situation in Members on the Region during 2021 and 2022; Regional Commission for Africa: Gaborone, Botswana, 2023. [Google Scholar]
- Corso, B.; Pinto, J.; Beltràn-Alcrudo, D.; De Simone, L.; Lubroth, J. Rift Valley Fever Outbreaks in Madagascar and Potential Risks to Neighbouring Countries; FAO Empress Watch: Rome, Italy, 2008; p. 8. [Google Scholar]
- WHO. Rift Valley fever, Madagascar. Wkly. Epidemiol. Rec. 2008, 83, 157–168. [Google Scholar]
- Andriamandimby, S.F.; Randrianarivo-Solofoniaina, A.E.; Jeanmaire, E.M.; Ravololomanana, L.; Razafimanantsoa, L.T.; Rakotojoelinandrasana, T.; Razainirina, J.; Hoffman, J.; Ravalohery, J.P.; Razafisandratantsoa, J.T.; et al. Rift Valley Fever during rainy seasons, Madagascar, 2008 and 2009. Em. Inf. Dis. 2010, 16, 963–970. [Google Scholar] [CrossRef]
- Ratovonjato, J.; Olive, M.M.; Tantely, M.L.; Andrianaivolambo, L.; Tata, E.; Razainirina, J.; Jeanmaire, E.M.; Reynes, J.M.; Elissa, N. Detection, isolation, and genetic characterisation of Rift Valley Fever virus from Anopheles (Anopheles) coustani, Anopheles (Anopheles) squamosus, Culex (Culex) antennatus of the Haute Matsiatra region, Madagascar. Vector-Borne Zoo. Dis. 2010, 11, 753–759. [Google Scholar] [CrossRef]
- Jeanmaire, E.; Rabenarivahiny, R.; Biarmann, M.; Rabibisoa, L.; Ravaomanana, F.; Randriamparany, T.; Andriamandimby, S.F.; Squarzoni, D.C.; Fenozara, P.; de La Rocque, S.; et al. Prevalence of Rift Valley Fever infection in ruminants in Madagascar after the 2008 outbreak. Vector-Borne Zoo. Dis. 2011, 11, 395–402. [Google Scholar] [CrossRef] [PubMed]
- Jeffries, C.; Tantely, M.L.; Raharimalala, F.; Hurn, E.; Boyer, S.; Walker, T. Diverse novel resident Wolbachia strains in Culicine mosquitoes from Madagascar. Sci. Rep. 2018, 8, 17456. [Google Scholar] [CrossRef] [PubMed]
- Nepomichene, T.N.J.J.; Raharimalala, F.N.; Andriamandimby, S.F.; Ravalohery, J.-P.; Failloux, A.-B.; Heraud, J.-M.; Boyer, S. Vector competence of Culex antennatus and Anopheles coustani mosquitoes for Rift Valley fever virus in Madagascar. Med. Vet. Entomol. 2018, 32, 259–262. [Google Scholar] [CrossRef] [PubMed]
- Ravaonjanahary, C. Les Aedes de Madagascar (Diptera-Culicidae). Étude monographique du genre. 2. Biologie d’Aedes (Diceromyia) tiptoni. Trav. Doc. L’orstom 1978, 87, 1–210. [Google Scholar]
- Grjébine, A. Insectes Diptères Culicidae Anophelinae; Faune de Madagascar; ORSTOM: Paris, France, 1966; pp. 1–500. [Google Scholar]
- Doucet, J. Étude des Culicidae de la région de Vangaindrano (Diptera). Mém. Inst. Sci. Mad. 1951, 6, 83–113. [Google Scholar]
- Edwards, F.W. Mosquitoes of the Ethiopian Region. III. Culicine Adults and Pupae; British Museum (Natural History): London, UK, 1941. [Google Scholar]
- Brunhes, J.; Hervy, J. Insectes Diptères Culicidae Culicinae Genre Orthopodomyia de la sous-région malgache et de la région afrotropicale. Mus. Natl. Hist. Nat. 1995, 85, 1–120. [Google Scholar]
- da Cunha Ramos, H.; Brunhes, J. Insecta, Diptera, Culicidae, Uranotaenia. Faune de Madagascar 91, 3rd ed.; CIRAD: Montpellier, Paris, 2004; p. 461. [Google Scholar]
- Pitzer, J.B.; Kaufman, P.E.; Tenbroeck, S.H.; Maruniak, J.E. Host blood meal identification by multiplex polymerase chain reaction for dispersal evidence of stable flies (Diptera, Muscidae) between livestock facilities. J. Med. Entomol. 2011, 48, 53–60. [Google Scholar] [CrossRef] [PubMed]
- Weidmann, M.; Sanchez-Seco, M.P.; Sall, A.A.; Ly, P.O.; Thiongan, Y.; Lô, M.M.; Schley, H.; Hufert, F.T. Rapid detection of important human pathogenic Phleboviruses. J. Clin. Virol. 2008, 41, 138–142. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. A Language and Environment for Statistical Computing. Foundation for Statistical Computing; R Core Team: Vienna, Austria, 2021; Available online: https://www.R–project.org/ (accessed on 1 February 2024).
- de La Rocque, S.; Formenty, P. Rift Valley fever, Disease ecology and early warning. Sustainable Management of Animal Production and Health; Odongo, N., Garcia, M., Viljoen, G., Eds.; FAO: Rome, Italy, 2010; pp. 327–333. [Google Scholar]
- Tantely, M.L.; Rakotoniaina, J.C.; Andrianaivolamb, O.L.; Tata, E.; Razafindrasata, F.; Fontenille, D.; Elissa, N. Biology of mosquitoes that are potential vectors of Rift Valley fever virus in different biotopes of the Central Highlands of Madagascar. J. Med. Entomol. 2013, 50, 603–610. [Google Scholar] [CrossRef] [PubMed]
- Nepomichene, T.; Tata, E.; Boyer, S. Malaria case in Madagascar.; probable implication of a new vector, Anopheles coustani. Malaria J. 2015, 14, 475. [Google Scholar] [CrossRef]
- Tantely, M.L.; Guis, H.; Randriananjantenaina, I.; Raharinirina, M.R.; Velonirina, H.J.; Cardinale, E.; Raveloarijaona, N.; Cêtre-Sossah, C.; Garros, C.; Girod, R. Mosquito species associated with horses in Madagascar, A review of their vector status with regard to the epidemiology of West Nile fever. Med. Vet. Entomol. 2021, 36, 13. [Google Scholar] [CrossRef] [PubMed]
- Andrianaivolambo, L.; Domarle, O.; Randrianarivelojosia, M.; Ratovonjato, J.; Le Goff, G.; Talman, A.; Robert, V. Anthropophilic mosquitoes and malaria transmission in the eastern foothills of the central highlands of Madagascar. Acta. Trop. 2010, 116, 240–245. [Google Scholar] [CrossRef] [PubMed]
- Tantely, M.L.; Cêtre–Sossah, C.; Rakotondranaivo, T.; Cardinale, E.; Boyer, S. Population dynamics of mosquito species in a West Nile Virus endemic area in Madagascar. Parasite 2017, 24, 3. [Google Scholar] [CrossRef] [PubMed]
- Fontenille, D.; Powell, J.R. From anonymous to public enemy, how does a mosquito become a feared Arbovirus vector? Pathogens 2020, 9, 265. [Google Scholar] [CrossRef]
- Tantely, M.L.; Le Goff, G.; Boyer, S.; Fontenille, D. An updated checklist of mosquito species (Diptera, Culicidae) from Madagascar. Parasite 2016, 23, 42. [Google Scholar] [CrossRef]
- Fontenille, D.; Campbell, G.H. Is Anopheles mascarensis a new malaria vector in Madagascar? Am. J. Trop. Med. Hyg. 1992, 46, 28–30. [Google Scholar] [CrossRef]
- Muturi, E.J.; Muriu, S.; Shililu, J.; Mwangangi, J.M.; Jacob, B.G.; Mbogo, C.; Githure, J.; Novak, R.J. Blood-feeding patterns of Culex quinquefasciatus and other culicines and implications for disease transmission in Mwea rice scheme, Kenya. Parasitol. Res. 2008, 102, 1329–1335. [Google Scholar] [CrossRef]
- Khaklang, S.; Kittayapong, P. Species composition and blood meal analysis of mosquitoes collected from a tourist island, Koh Chang, Thailand. J. Vector Ecol. 2014, 39, 448–452. [Google Scholar] [CrossRef]
- Hammon, W.M.; Reeves, W.C. Laboratory Transmission of St. Louis Encephalitis Virus by Three Genera of Mosquitoes. J. Exp. Med. 1943, 78, 241–253. [Google Scholar] [CrossRef]
- McIntosh, B.M. Rift Valley fever. 1. Vector studies in the field. J. S. Afr. Vet. Med. Assoc. 1972, 43, 391–395. [Google Scholar] [PubMed]
- Turell, M.; Dohm, D.J.; Mores, C.N.; Terracina, L.; Wallette, D.L.; Hribar, L.J.; Pecor, J.E.; Blow, J.A. Potential for North American mosquitoes to transmit Rift Valley fever virus. J. S. Afr. Vet. Med. Assoc. 2008, 24, 502–507. [Google Scholar] [CrossRef]
- Jupp, P.G.; Kemp, A.; Grobbelaar, A.; Leman, P.; Burt, F.J.; Alahmed, A.M.; Al Mujalli, D.; Khamees, A.; Swanepoel, R. The 2000 epidemic of Rift Valley fever in Saudi Arabia, mosquito vector studies. Med. Vet. Entomol. 2002, 16, 245–252. [Google Scholar] [CrossRef] [PubMed]
- Léong Pock Tsy, J.M.; Duchemin, J.B.; Marrama, L.; Rabarison, P.; Le Goff, G.; Rajaonarivelo, V.; Robert, V. Distribution of the species of the Anopheles gambiae complex and first evidence of Anopheles merus as a malaria vector in Madagascar. Malaria J. 2003, 2, 33. [Google Scholar] [CrossRef] [PubMed]
- Seufi, A.M.; Galal, F.H. Role of Culex and Anopheles mosquito species as potential vectors of Rift Valley fever virus in Sudan outbreak, 2007. Infect. Dis. 2010, 10, 65. [Google Scholar] [CrossRef] [PubMed]
- Moutailler, S.; Krida, G.; Schaffner, F.; Vazeille, M.; Failloux, A.B. Potential vectors of Rift Valley fever virus in the Mediterranean Region. Vector Borne Zoonotic Dis. 2008, 8, 749–754. [Google Scholar] [CrossRef] [PubMed]
- Turell, M.; Bailey, C.; Beaman, J. Vector competence of a Houston, Texas strain of Aedes albopictus for Rift Valley fever virus. J. Am. Mosq. Control. Assoc. 1988, 4, 94–96. [Google Scholar]
- Jupp, P.; Cornel, A.; Turell, M.; Bailey, C.; Beaman, J. Vector competence tests with Rift Valley fever virus and five south African species of mosquito. Am. Mosq. Control. Assoc. 1988, 4, 4–8. [Google Scholar]
- Junglen, S.; Kurth, A.; Kuehl, H.; Quan, P.L.; Ellerbrok, H.; Pauli, G.; Nitsche, A.; Nunn, C.; Rich, S.M.; Lipkin, W.I.; et al. Examining landscape factors influencing relative distribution of mosquito genera and frequency of virus infection. EcoHealth 2009, 6, 239–249. [Google Scholar] [CrossRef] [PubMed]
- Chevalier, V.; Pépin, M.; Plée, L.; Lancelot, R. Rift Valley fever—A threat for Europe? Eurosurveillance 2010, 15, 19506. [Google Scholar] [CrossRef] [PubMed]
- Hamon, J.; Sales, S.; Coz, J.; Ouedraogo, C.S.; Dyemkouma, A.; Diallo, B. Observations sur les préférences alimentaires des moustiques de la République de Haute-Volta. Bull. Soc. Pathol. Exot. 1964, 57, 1133–1150. [Google Scholar]
- Mondet, B.; Diaite, A.; Ndione, J.A.; Fall, A.G.; Chevalier, V.; Lancelot, R.; Ndiaye, M.; Ponçon, N. Rainfall patterns and population dynamics of Aedes (Aedimorphus) vexans arabiensis, Patton 1905 (Diptera, Culicidae).; a potential vector of Rift Valley fever virus in Senegal. J. Vector. Ecol. 2005, 30, 102–106. [Google Scholar] [PubMed]
- Brunhes, J. Épidémiologie de la filariose de Bancroft à Madagascar. II. Recherches sur Wuchereria voceli Galliard.; 1959 et étude morphologique comparée des souches malgache et comorienne de Wuchereria bancrofti Cobbold, 1877. Sér. Ent. Méd. Parasitol. 1972, 10, 193–205. [Google Scholar]
- Fontenille, D.; Lepers, J.P.; Campbell, G.H.; Coluzzi, M.; Rakotoarivony, I.; Coulanges, P. Malaria transmission and vector biology in Manarintsoa, High Plateaux of Madagascar. Am. J. Trop. Med. Hyg. 1990, 43, 107–115. [Google Scholar] [CrossRef]
- Rice, B.L.; Golden, C.D.; Randriamady, H.J.; Rakotomalala, A.A.N.A.; Vonona, M.A.; Anjaranirina, E.J.G.; Hazen, J.; Castro, M.C.; Metcalf, J.E.; Hartl, D.L. Fine-scale variation in malaria prevalence across ecological regions in Madagascar, a cross-sectional study. BMC Public Health 2021, 21, 1018. [Google Scholar] [CrossRef]
- Vaughan, J.A.; Turell, M.J. Facilitation of Rift Valley Fever virus transmission by Plasmodium berghei sporozoites in Anopheles stephensi mosquitoes. Am. J. Trop. Med. Hyg. 1996, 55, 407–409. [Google Scholar] [CrossRef]
- Miller, B.R.; Monath, T.P.; Tabachnick, W.J.; Ezike, V.I. Epidemic yellow fever caused by an incompetent mosquito vector. Trop. Med. Parasitol. 1989, 40, 396–399. [Google Scholar]
- Lancelot, R.; Béral, M.; Rakotoharinome, V.M.; Andriamandimby, S.F.; Heraud, J.M.; Costea, C.; Apollonia, A.; Squarzoni-Diawg, C.; de La Rocquea, S.; Formenty, P.B.H.; et al. Drivers of Rift Valley fever epidemics in Madagascar. Proc. Natl. Acad. Sci. USA 2017, 114, 938–943. [Google Scholar] [CrossRef]
- Bird, B.H.; Ksiazek, T.G.; Nichol, S.T.; McLachlan, J. Rift Valley fever virus. J. Am. Vet. Med. Assoc. 2009, 234, 883–893. [Google Scholar] [CrossRef] [PubMed]
- Linthicum, K.J.; Britch, S.C.; Anyamba, A. Rift Valley fever, an emerging mosquito-borne disease. Annu. Rev. Entomol. 2016, 61, 395–415. [Google Scholar] [CrossRef] [PubMed]
- Lumley, S.; Hernández-Triana, L.M.; Horton, D.L.; de Marco, M.D.M.F.; Medlock, J.M.; Hewson, R.; Fooks, A.R.; Johnson, N. Competence of mosquitoes native to the United Kingdom to support replication and transmission of Rift Valley fever virus. Par. Vect. 2018, 11, 308. [Google Scholar] [CrossRef] [PubMed]
- Peters, C.J.; Linthicum, K.J. Rift Valley fever. In Handbook of Zoonoses, 2nd ed.; Beran, G.W., Ed.; CRC Press: Boca Raton, FL, USA, 1994; pp. 125–138. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Species | Ambalamainty | Ambodiakatra | Madiolamba | Total | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| BGs | LTs | MTPTs | IPSCs | BGs | LTs | MTPTs | IPSCs | BGs | LTs | MTPTs | IPSCs | ||
| Ad. furfurea | 0 | 2 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 3 |
| Ae. albopictus * | 7 | 0 | 0 | 0 | 8 | 0 | 1 | 2 | 6 | 5 | 0 | 0 | 29 |
| Ae. argenteopunctatus * | 0 | 23 | 0 | 0 | 0 | 31 | 1 | 0 | 0 | 36 | 0 | 0 | 91 |
| Ae. circumluteolus £, V | 0 | 0 | 0 | 0 | 0 | 2 | 0 | 0 | 0 | 0 | 0 | 0 | 2 |
| Ae. sp. | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 2 | 0 | 0 | 3 |
| An. coustani **, V | 0 | 143 | 0 | 0 | 0 | 69 | 0 | 0 | 0 | 87 | 0 | 0 | 299 |
| An. funestus | 0 | 190 | 6 | 4 | 1 | 17 | 2 | 6 | 0 | 33 | 1 | 9 | 269 |
| An. gambiae sl £, V | 1 | 13 | 0 | 0 | 0 | 8 | 0 | 1 | 0 | 11 | 0 | 0 | 34 |
| An. maculipalpis | 0 | 64 | 0 | 0 | 0 | 7 | 0 | 0 | 2 | 12 | 0 | 0 | 85 |
| An. mascarensis | 5 | 765 | 18 | 0 | 0 | 18 | 0 | 0 | 0 | 241 | 2 | 0 | 1049 |
| An. pauliani | 0 | 50 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 51 |
| An. rufipes | 0 | 0 | 0 | 0 | 0 | 2 | 0 | 0 | 0 | 0 | 0 | 0 | 2 |
| An. squamosus/cydippis ** | 0 | 91 | 0 | 0 | 0 | 123 | 0 | 0 | 0 | 83 | 0 | 0 | 297 |
| Cq. grandidieri | 0 | 14 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 5 | 0 | 0 | 19 |
| Cx. antennatus *** | 0 | 44 | 0 | 0 | 0 | 26 | 4 | 1 | 1 | 2 | 0 | 0 | 78 |
| Cx. bitaeniorhynchus £ | 0 | 5 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 6 |
| Cx. decens ** | 0 | 64 | 0 | 0 | 0 | 12 | 0 | 0 | 0 | 27 | 1 | 0 | 104 |
| Cx. giganteus | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 1 |
| Cx. quinquefasciatus **, £ | 0 | 0 | 0 | 0 | 17 | 4 | 1 | 53 | 0 | 0 | 0 | 0 | 75 |
| Cx. tritaeniorhynchus *** | 2 | 63 | 0 | 0 | 0 | 10 | 0 | 0 | 0 | 93 | 0 | 0 | 168 |
| Cx. univittatus **, £, V | 0 | 32 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 33 |
| Cx. sp. | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 1 | 0 | 0 | 2 |
| Er. quinquevitattus **, £ | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 2 |
| Ficalbia sp. | 0 | 1 | 0 | 0 | 0 | 3 | 0 | 0 | 0 | 2 | 0 | 0 | 6 |
| Ma. uniformis ** | 0 | 9 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 73 | 0 | 0 | 82 |
| Or. milloti | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 1 |
| Ur. sp. | 0 | 4 | 0 | 0 | 1 | 3 | 0 | 0 | 0 | 1 | 1 | 0 | 10 |
| Lt tigripes | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 |
| Unidentified genus | 1 | 2 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 4 |
| Total | 16 | 1580 | 24 | 4 | 28 | 339 | 10 | 63 | 9 | 719 | 5 | 9 | 2806 |
| Ambalamainty | Ambodiakatra | Madiolamba | Total | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Females | Males | Females | Males | Females | Males | |||||
| Species | Go | Non-Go | Go | Non-Go | Go | Non-Go | ||||
| Ad. furfurea | - | 2 | - | - | - | - | - | 1 | - | 3 |
| Ae. albopictus *** , µ | - | 5 | 1 | - | 6 (1) | 1 | - | 7 | 1 | 21 (1) |
| Ae. argenteopunctatus *** , µ | - | 4 | 2 | 2 (2) | 6 (1) | - | - | 8 | - | 22 (3) |
| Ae. circumluteolus £, V | - | - | - | 1 | 1 | - | - | - | - | 2 |
| Ae. sp. | - | - | - | - | 1 | - | - | 2 | - | 3 |
| An. coustani **, V | 1 (1) | 17 (1) | 3 | 4 (1) | 6 | 1 | - | 12 | 1 | 45 (3) |
| An. funestus | 4 | 23 | 2 | 3 | 6 | 2 | 3 | 7 | 3 | 53 |
| An. gambiae sl **, £, V, µ | - | 4 | 2 | - | 4 | 2 | - | 4 (1) | 1 | 17 (1) |
| An. maculipalpis | 3 | 6 | - | 1 | 1 | - | 1 | 4 | - | 16 |
| An. mascarensis **, µ | 26 (3) | 60 | 4 | 2 | 3 (1) | - | 1 | 29 (5) | 2 | 127 (9) |
| An. pauliani | - | 5 | - | - | - | - | - | 1 | - | 6 |
| An. rufipes | - | - | - | - | 2 | - | - | - | - | 2 |
| An. squamosus/cydippis ** | 2 | 12 (3) | 1 | 8 (2) | 9 (2) | - | 1 | 9(1) | - | 42 (8) |
| Coquillettidia grandidieri | - | 4 | - | - | - | - | - | 2 | 1 | 7 |
| Cx. antennatus *** | 1 (1) | 7 (1) | - | 4 (1) | 6 (1) | - | - | 2 | - | 20 (4) |
| Cx. bitaeniorhynchus £ | 1 | 1 | - | 1 | - | - | - | - | - | 3 |
| Cx. decens ** | 2 (1) | 9 (1) | 4 | - | 3 | - | 1 | 5 | 2 | 26 (2) |
| Cx. giganteus | - | - | - | - | 1 | - | - | - | - | 1 |
| Cx. quinquefasciatus **, £ | - | - | - | 3 | 6 | 10 | - | - | - | 19 |
| Cx. tritaeniorhynchus *** | 1 | 9 | - | 2 | 3 | - | - | 12 (5) | 1 | 28 (5) |
| Cx. univittatus **, £, V | 1 | 6 | 2 | - | - | - | - | 1 | - | 10 |
| Cx. sp. | - | - | - | 1 | - | - | - | 1 | - | 2 |
| Er. quinquevitattus **, £ | - | - | - | - | - | 1 | - | 1 | - | 2 |
| Ficalbia sp. | - | 1 | - | - | 2 | 1 | - | 2 | - | 6 |
| Ma. uniformis ** | 1 | 5 | - | - | - | - | - | 10 | 1 | 17 |
| Or. milloti | - | - | - | - | - | - | - | - | 1 | 1 |
| Ur. sp. *, µ | - | 2 (1) | - | - | 3 | - | - | 2 | - | 7 (1) |
| Lt. tigripes | - | 1 | - | - | - | - | - | - | - | 1 |
| Genre sp. | - | 2 | - | - | 1 | - | - | - | - | 3 |
| Total | 43 (6) | 185 (7) | 21 | 32 (6) | 70 (6) | 18 | 7 | 122 (12) | 14 | 512 (37) |
| Species | Engorged Positives | Blood Meals (Mixed and Single) | |||
|---|---|---|---|---|---|
| RVFV + Pools | Ind Go/Pool+ | Mixed | Human | Cattle | |
| Ae. argenteopunctatus | 2 | 4 * + 5 | 3 | 0 | 4 |
| An. coustani | 2 | 1 + 5 | 6 | 0 | 0 |
| An. mascarensis | 3 | 10 + 10 + 10 | 20 | 10 | 0 |
| An. squamosus/cydippis | 2 | 2 + 10 | 12 | 0 | 0 |
| Cx. antennatus | 2 | 1 + 2 | 2 | 0 | 1 |
| Cx. decens | 1 | 1 | 1 | 0 | 0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tantely, L.M.; Andriamandimby, S.F.; Ambinintsoa, M.F.; Raharinirina, M.R.; Rafisandratantsoa, J.T.; Ravalohery, J.-P.; Harimanana, A.; Ranoelison, N.N.; Irinantenaina, J.; Ankasitrahana, M.F.; et al. An Entomological Investigation during a Recent Rift Valley Fever Epizootic/Epidemic Reveals New Aspects of the Vectorial Transmission of the Virus in Madagascar. Pathogens 2024, 13, 258. https://doi.org/10.3390/pathogens13030258
Tantely LM, Andriamandimby SF, Ambinintsoa MF, Raharinirina MR, Rafisandratantsoa JT, Ravalohery J-P, Harimanana A, Ranoelison NN, Irinantenaina J, Ankasitrahana MF, et al. An Entomological Investigation during a Recent Rift Valley Fever Epizootic/Epidemic Reveals New Aspects of the Vectorial Transmission of the Virus in Madagascar. Pathogens. 2024; 13(3):258. https://doi.org/10.3390/pathogens13030258
Chicago/Turabian StyleTantely, Luciano Michaël, Soa Fy Andriamandimby, Maminirina Fidelis Ambinintsoa, Manou Rominah Raharinirina, Jean Théophile Rafisandratantsoa, Jean-Pierre Ravalohery, Aina Harimanana, Nirina Nantenaina Ranoelison, Judickaelle Irinantenaina, Miamina Fidy Ankasitrahana, and et al. 2024. "An Entomological Investigation during a Recent Rift Valley Fever Epizootic/Epidemic Reveals New Aspects of the Vectorial Transmission of the Virus in Madagascar" Pathogens 13, no. 3: 258. https://doi.org/10.3390/pathogens13030258