
Lack of Detection of Toxoplasma gondii in Pipistrellus spp. Bats from Densely Cat-Populated Areas of NE Spain
 ,
,  , ,
, ,  and
and 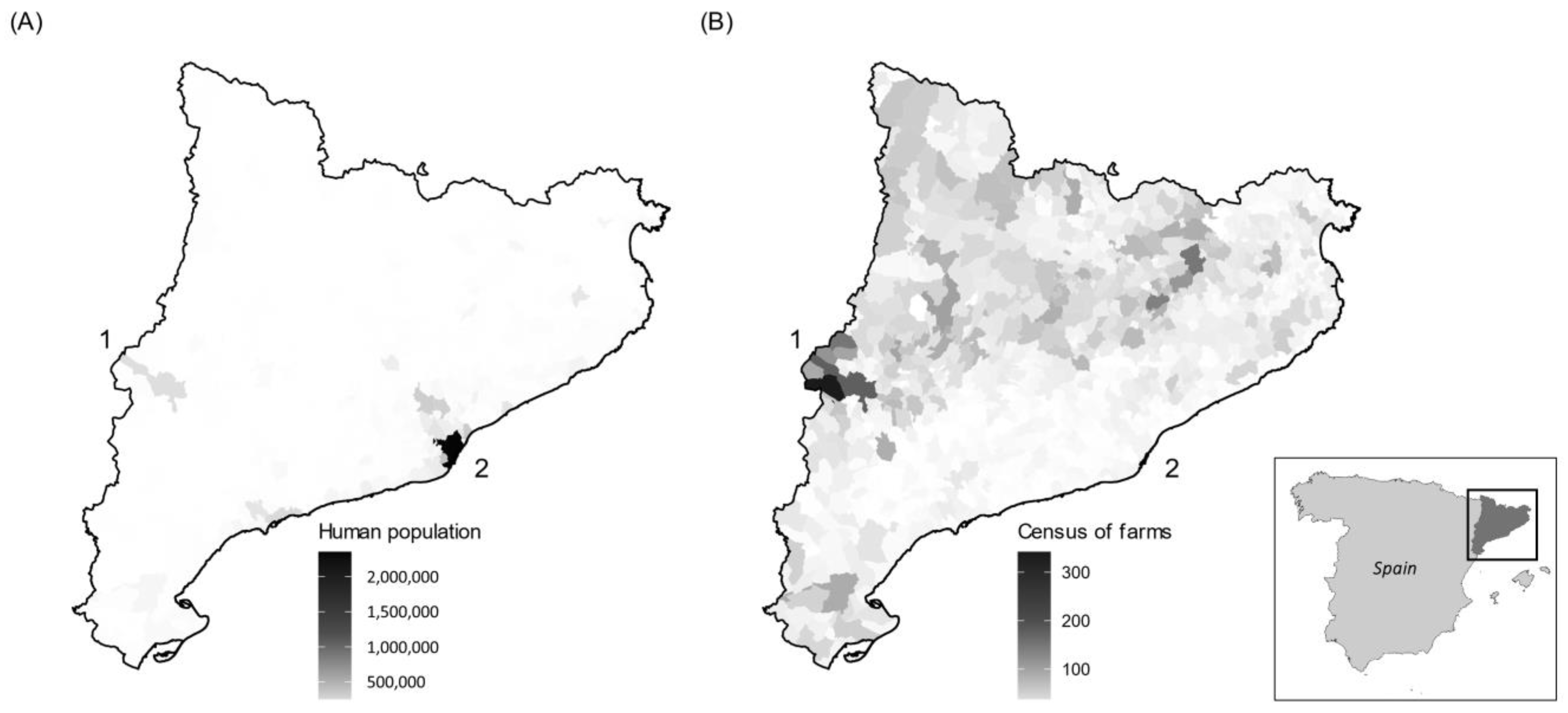{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Samples
2.2. Molecular Analysis
2.3. Statistical Analyses
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sun, H.; Wang, Y.; Zhang, Y.; Ge, W.; Zhang, F.; He, B.; Li, Z.; Fan, Q.; Wang, W.; Tu, C.; et al. Prevalence and Genetic Characterization of Toxoplasma Gondii in Bats in Myanmar. Appl. Environ. Microbiol. 2013, 79, 3526–3528. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calisher, C.H.; Childs, J.E.; Field, H.E.; Holmes, K.V.; Schountz, T. Bats: Important Reservoir Hosts of Emerging Viruses. Clin. Microbiol. Rev. 2006, 19, 531–545. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hayman, D.T.S.; Bowen, R.A.; Cryan, P.M.; McCracken, G.F.; O’Shea, T.J.; Peel, A.J.; Gilbert, A.; Webb, C.T.; Wood, J.L.N. Ecology of Zoonotic Infectious Diseases in Bats: Current Knowledge and Future Directions. Zoonoses Public Health 2013, 60, 2–21. [Google Scholar] [CrossRef] [Green Version]
- Hutson, A.; Mickleburgh, S.P.; Racey, P.A. Microchiropteran Bats: Global Status Survey and Conservation Action Plan; IUCN/SSC Chiroptera Specialist Group; IUCN: Gland, Switzerland; Cambridge, UK, 2001; p. 258. [Google Scholar]
- Frick, W.F.; Kingston, T.; Flanders, J. A Review of the Major Threats and Challenges to Global Bat Conservation. Ann. N. Y. Acad. Sci. 2019, 1469, 5–25. [Google Scholar] [CrossRef] [PubMed]
- Voigt, C.C.; Kingston, T. Bats in the Anthropocene: Conservation of Bats in a Changing World; Springer International Publishing: Berlin/Heidelberg, Germany, 2015; ISBN 9783319252209. [Google Scholar]
- Dubey, J.P. Toxoplasmosis of Animals and Humans, 3rd ed.; CRC Press: Boca Ratón, FL, USA, 2021. [Google Scholar]
- Dubey, J.P. Toxoplasmosis—A Waterborne Zoonosis. Vet. Parasitol 2004, 126, 57–72. [Google Scholar] [CrossRef]
- Sangster, C.; Gordon, A.; Hayes, D. Systemic Toxoplasmosis in Captive Flying-Foxes. Aust. Vet. J. 2012, 90, 140–142. [Google Scholar] [CrossRef]
- Tenter, A.; Heckeroth, A.; Weiss, L. Toxoplasma Gondii: From Animals to Humans. Int. J. Parasitol. 2000, 30, 1217–1258. [Google Scholar] [CrossRef] [Green Version]
- Almería, S.; Cabezón, O.; Paniagua, J.; Cano-Terriza, D.; Jiménez-Ruiz, S.; Arenas-Montes, A.; Dubey, J.P.; García-Bocanegra, I. Toxoplasma Gondii in Sympatric Domestic and Wild Ungulates in the Mediterranean Ecosystem. Parasitol. Res. 2018, 117, 665–671. [Google Scholar] [CrossRef] [Green Version]
- Cabezón, O.; García-Bocanegra, I.; Molina-López, R.; Marco, I.; Blanco, J.M.; Höfle, U.; Margalida, A.; Bach-Raich, E.; Darwich, L.; Echeverría, I.; et al. Seropositivity and Risk Factors Associated with Toxoplasma Gondii Infection in Wild Birds from Spain. PLoS ONE 2011, 6, e29549. [Google Scholar] [CrossRef] [Green Version]
- Ribas, M.P.; Almería, S.; Fernández-Aguilar, X.; de Pedro, G.; Lizarraga, P.; Alarcia-Alejos, O.; Molina-López, R.; Obón, E.; Gholipour, H.; Temiño, C.; et al. Tracking Toxoplasma Gondii in Freshwater Ecosystems: Interaction with the Invasive American Mink (Neovison Vison) in Spain. Parasitol. Res. 2018, 117, 2275–2281. [Google Scholar] [CrossRef]
- Dodd, N.S.; Lord, J.S.; Jehle, R.; Parker, S.; Parker, F.; Brooks, D.R.; Hide, G. Toxoplasma Gondii: Prevalence in Species and Genotypes of British Bats (Pipistrellus Pipistrellus and P. Pygmaeus). Exp. Parasitol. 2014, 139, 6–11. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Xin, S.; Murata, F.H.A.; Cerqueira-Cézar, C.K.; Kwok, O.C.H.; Su, C.; Dubey, J.P. Recent Epidemiologic, Clinical, Subclinical and Genetic Diversity of Toxoplasma Gondii Infections in Bats. Res. Vet. Sci. 2021, 140, 193–197. [Google Scholar] [CrossRef]
- Jiang, H.H.; Qin, S.Y.; Wang, W.; He, B.; Hu, T.S.; Wu, J.M.; Fan, Q.S.; Tu, C.C.; Liu, Q.; Zhu, X.Q. Prevalence and Genetic Characterization of Toxoplasma Gondii Infection in Bats in Southern China. Vet. Parasitol. 2014, 203, 318–321. [Google Scholar] [CrossRef]
- National Air and Space Administration (NASA) Socioeconomic Data and Applications Center (SEDAC): Gridded Population of the World. Available online: https://sedac.ciesin.columbia.edu/data/collection/gpw-v4 (accessed on 27 October 2022).
- Aegerter, J.; Fouracre, D.; Smith, G.C. A First Estimate of the Structure and Density of the Populations of Pet Cats and Dogs across Great Britain. PLoS ONE 2017, 12, e0174709. [Google Scholar] [CrossRef] [Green Version]
- Simon, J.A.; Kurdzielewicz, S.; Jeanniot, E.; Dupuis, E.; Marnef, F.; Aubert, D.; Villena, I.; Poulle, M.-L. Spatial Distribution of Soil Contaminated with Toxoplasma Gondii Oocysts in Relation to the Distribution and Use of Domestic Cat Defecation Sites on Dairy Farms. Int. J. Parasitol. 2017, 47, 357–367. [Google Scholar] [CrossRef]
- Guardiola, Á.; Fernández, P. Pipistrellus Pipistrellus (Schreber, 1774). In Ficha Libro Rojo. Atlas y Libro Rojo de los Mamíferos Terrestres de España; Dirección General para la Biodiversidad -SECEM-SECEMU: Madrid, Spain, 2007; pp. 199–202. [Google Scholar]
- Guardiola, Á.; Fernández, P. Pipistrellus Pygmaeus (Leach, 1825). In Ficha Libro Rojo. Atlas y Libro Rojo de los Mamíferos Terrestres de España; Dirección General para la Biodiversidad -SECEM-SECEMU: Madrid, Spain, 2007; pp. 203–206. [Google Scholar]
- Goiti, U.; Garin, I. Pipistrellus Kuhlii (Kuhl, 1817) Orden. In Ficha Libro Rojo. Atlas y Libro Rojo de los Mamíferos Terrestres de España; Dirección General para la Biodiversidad -SECEM-SECEMU: Madrid, Spain, 2007; pp. 215–217. [Google Scholar]
- Ajzenberg, D.; Lamaury, I.; Demar, M.; Vautrin, C.; Cabié, A.; Simon, S.; Nicolas, M.; Desbois-Nogard, N.; Boukhari, R.; Riahi, H.; et al. Performance Testing of PCR Assay in Blood Samples for the Diagnosis of Toxoplasmic Encephalitis in AIDS Patients from the French Departments of America and Genetic Diversity of Toxoplasma Gondii: A Prospective and Multicentric Study. PLoS Negl. Trop. Dis. 2016, 10, e0004790. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galal, L.; Schares, G.; Stragier, C.; Vignoles, P.; Brouat, C.; Cuny, T.; Dubois, C.; Rohart, T.; Glodas, C.; Dardé, M.-L.; et al. Diversity of Toxoplasma Gondii Strains Shaped by Commensal Communities of Small Mammals. Int. J. Parasitol. 2019, 49, 267–275. [Google Scholar] [CrossRef] [PubMed]
- Dohoo, I.R.; Martin, W.; Stryhn, H.E. Veterinary Epidemiologic Research; University of Prince Edward Island: Charlottetown, PE, Canada, 2003. [Google Scholar]
- Molina-López, R.; Cabezón, O.; Pabón, M.; Darwich, L.; Obón, E.; Lopez-Gatius, F.; Dubey, J.P.; Almería, S. High Seroprevalence of Toxoplasma Gondii and Neospora Caninum in the Common Raven (Corvus Corax) in the Northeast of Spain. Res. Vet. Sci. 2012, 93, 300–302. [Google Scholar] [CrossRef] [PubMed]
- Cabral, A.D.; Su, C.; Soares, R.M.; Gennari, S.M.; Sperança, M.A.; da Rosa, A.R.; Pena, H.F.J. Occurrence and Diversity of Sarcocystidae Protozoa in Muscle and Brain Tissues of Bats from São Paulo State, Brazil. Int. J. Parasitol. Parasites Wildl. 2021, 14, 91–96. [Google Scholar] [CrossRef] [PubMed]
- Qin, S.-Y.; Cong, W.; Liu, Y.; Li, N.; Wang, Z.-D.; Zhang, F.-K.; Huang, S.-Y.; Zhu, X.-Q.; Liu, Q. Molecular Detection and Genotypic Characterization of Toxoplasma Gondii Infection in Bats in Four Provinces of China. Parasit. Vectors 2014, 7, 558. [Google Scholar] [CrossRef] [PubMed]
- Gauss, C.B.L.; Almería, S.; Ortuño, A.; Garcia, F.; Dubey, J.P. Seroprevalence of Toxoplasma Gondii Antibodies in Domestic Cats from Barcelona, Spain. J. Parasitol. 2003, 89, 1067–1068. [Google Scholar] [CrossRef]
- Stavisky, J.; Brennan, M.L.; Downes, M.; Dean, R. Demographics and Economic Burden of Un-Owned Cats and Dogs in the UK: Results of a 2010 Census. BMC Vet. Res. 2012, 8, 163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dabritz, H.A.; Conrad, P.A. Cats and Toxoplasma: Implications for Public Health. Zoonoses Public Health 2010, 57, 34–52. [Google Scholar] [CrossRef]
- Gilot-Fromont, E.; Llu, M.; Dard, M.-L.; Richomme, C.; Aubert, D.; Afonso, E.; Mercier, A.; Gotteland, C.; Ville, I. The Life Cycle of Toxoplasma Gondii in the Natural Environment. In Toxoplasmosis—Recent Advances; InTech: London, UK, 2012. [Google Scholar]
- Meerburg, B.G.; Kijlstra, A. Changing Climate—Changing Pathogens: Toxoplasma Gondii in North-Western Europe. Parasitol. Res. 2009, 105, 17–24. [Google Scholar] [CrossRef] [Green Version]
- Chadwick, E.A.; Cable, J.; Chinchen, A.; Francis, J.; Guy, E.; Kean, E.F.; Paul, S.C.; Perkins, S.E.; Sherrard-Smith, E.; Wilkinson, C.; et al. Seroprevalence of Toxoplasma Gondii in the Eurasian Otter (Lutra Lutra) in England and Wales. Parasit. Vectors 2013, 6, 75. [Google Scholar] [CrossRef] [Green Version]
- Torres-Castro, M.; Muñoz-Dueñas, D.; Hernández-Betancourt, S.; Bolio-González, M.; Noh-Pech, H.; Peláez-Sánchez, R. Infección Con Toxoplasma Gondii (Eucoccidiorida: Sarcocystidae) En Murciélagos de Campeche y Yucatán, México. Rev. Biol. Trop. 2019, 67, 633–642. [Google Scholar]
- Cabral, A.D.; Gama, A.R.; Sodré, M.M.; Savani, E.S.M.M.; Galvão-Dias, M.A.; Jordão, L.R.; Maeda, M.M.; Yai, L.E.O.; Gennari, S.M.; Pena, H.F.J. First Isolation and Genotyping of Toxoplasma Gondii from Bats (Mammalia: Chiroptera). Vet. Parasitol. 2013, 193, 100–104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Graczyk, T.K.; Knight, R.; Tamang, L. Mechanical Transmission of Human Protozoan Parasites by Insects. Clin. Microbiol. Rev. 2005, 18, 128–132. [Google Scholar] [CrossRef] [Green Version]
- Guthrie, A.; Rooker, L.; Tan, R.; Gerhold, R.; Trainor, K.; Jiang, T.; Su, C. Newly Describerd Toxoplasma Gondii Strain Causes High Mortality in Red Necked Wallabies (Macropus Rufogriseus) in a Zoo. J. Zoo Wildl. Med. 2017, 48, 694–702. [Google Scholar] [CrossRef]
- Nishimura, M.; Goyama, T.; Tomikawa, S.; Fereig, R.M.; El-Alfy, E.-S.N.; Nagamune, K.; Kobayashi, Y.; Nishikawa, Y. Outbreak of Toxoplasmosis in Four Squirrel Monkeys (Saimiri Sciureus) in Japan. Parasitol. Int. 2019, 68, 79–86. [Google Scholar] [CrossRef]
- Mühldorfer, K.; Speck, S.; Wibbelt, G. Diseases in Free-Ranging Bats from Germany. BMC Vet. Res. 2011, 7, 61. [Google Scholar] [CrossRef] [PubMed]
- Mühldorfer, K.; Speck, S.; Kurth, A.; Lesnik, R.; Freuling, C.; Müller, T.; Kramer-Schadt, S.; Wibbelt, G. Diseases and Causes of Death in European Bats: Dynamics in Disease Susceptibility and Infection Rates. PLoS ONE 2011, 6, e29773. [Google Scholar] [CrossRef] [PubMed] [Green Version]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lobato-Bailón, L.; López-Morales, A.; Quintela, R.; Ribas, M.P.; Molina-López, R.; Obon, E.; Napp, S.; Pailler-García, L.; Espunyes, J.; Cabezón, Ó. Lack of Detection of Toxoplasma gondii in Pipistrellus spp. Bats from Densely Cat-Populated Areas of NE Spain. Pathogens 2022, 11, 1451. https://doi.org/10.3390/pathogens11121451
Lobato-Bailón L, López-Morales A, Quintela R, Ribas MP, Molina-López R, Obon E, Napp S, Pailler-García L, Espunyes J, Cabezón Ó. Lack of Detection of Toxoplasma gondii in Pipistrellus spp. Bats from Densely Cat-Populated Areas of NE Spain. Pathogens. 2022; 11(12):1451. https://doi.org/10.3390/pathogens11121451
Chicago/Turabian StyleLobato-Bailón, Lourdes, Ane López-Morales, Rita Quintela, Maria Puig Ribas, Rafael Molina-López, Elena Obon, Sebastián Napp, Lola Pailler-García, Johan Espunyes, and Óscar Cabezón. 2022. "Lack of Detection of Toxoplasma gondii in Pipistrellus spp. Bats from Densely Cat-Populated Areas of NE Spain" Pathogens 11, no. 12: 1451. https://doi.org/10.3390/pathogens11121451