
Genetic Variants of Glucose-6-Phosphate Dehydrogenase and Their Associated Enzyme Activity: A Systematic Review and Meta-Analysis
 , , , , , ,
, , , , , ,  , , , , add
Show full author list
, , , , add
Show full author list
Abstract
:1. Introduction
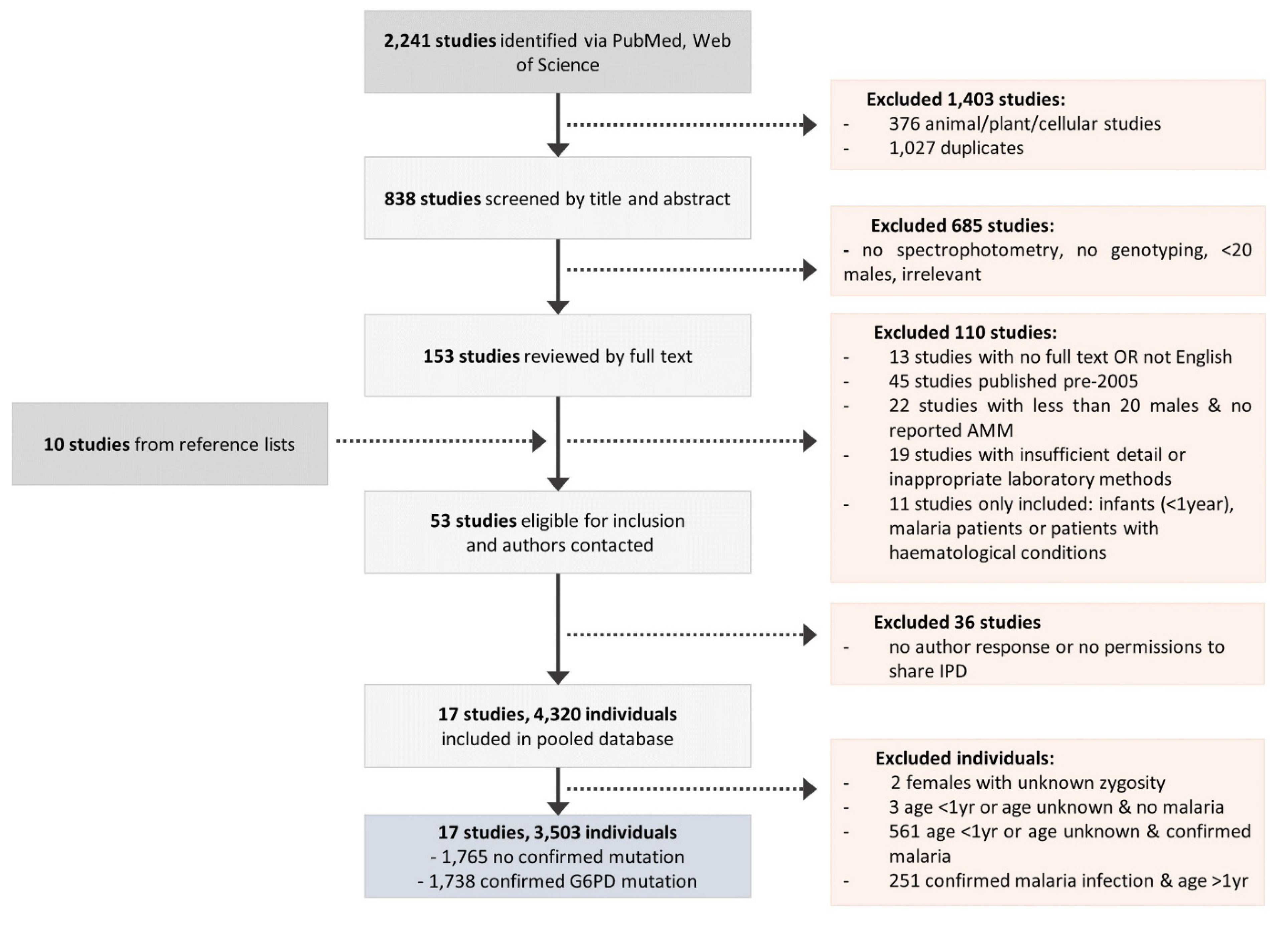
2. Methods
3. Results
3.1. Characteristics of the Pooled Database
3.2. Data-Rich Variants
3.3. Data-Poor Variants
3.4. Diagnostic Implications of Observed Variability
3.5. Assessment of Study Quality and Risk of Bias
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- World Health Organization. World Malaria Report 2020: 20 Years of Global Progress and Challenges; World Health Organization: Geneva, Switzerland, 2020. [Google Scholar]
- Battle, K.E.; Lucas, T.C.D.; Nguyen, M.; Howes, R.E.; Nandi, A.K.; Twohig, K.A.; Pfeffer, D.A.; Cameron, E.; Rao, P.C.; Casey, D.; et al. Mapping the global endemicity and clinical burden of Plasmodium vivax, 2000–2017: A spatial and temporal modelling study. Lancet 2019. [Google Scholar] [CrossRef]
- Baird, J.K.; Valecha, N.; Duparc, S.; White, N.J.; Price, R.N. Diagnosis and treatment of Plasmodium vivax malaria. Am. J. Trop. Med. Hyg. 2016, 95, 35–51. [Google Scholar] [CrossRef] [PubMed]
- Battle, K.E.; Baird, J.K. The global burden of Plasmodium vivax malaria is obscure and insidious. PLoS Med. 2021, 18, e1003799. [Google Scholar] [CrossRef] [PubMed]
- Thriemer, K.; Ley, B.; von Seidlein, L. Towards the elimination of Plasmodium vivax malaria: Implementing the radical cure. PLoS Med. 2021, 18, e1003494. [Google Scholar] [CrossRef] [PubMed]
- Howes, R.E.; Piel, F.B.; Patil, A.P.; Nyangiri, O.A.; Gething, P.W.; Dewi, M.; Hogg, M.M.; Battle, K.E.; Padilla, C.D.; Baird, J.K. G6PD deficiency prevalence and estimates of affected populations in malaria endemic countries: A geostatistical model-based map. PLoS Med. 2012, 9, e1001339. [Google Scholar] [CrossRef]
- Luzzatto, L.; Ally, M.; Notaro, R. Glucose-6-Phosphate Dehydrogenase Deficiency. Blood 2020, 136, 1225–1240. [Google Scholar] [CrossRef]
- World Health Organization. Guidelines for the Treatment of Malaria, 3rd ed.; World Health Organization: Geneva, Switzerland, 2015. [Google Scholar]
- Domingo, G.J.; Satyagraha, A.W.; Anvikar, A.; Baird, K.; Bancone, G.; Bansil, P.; Carter, N.; Cheng, Q.; Culpepper, J.; Eziefula, C.; et al. G6PD testing in support of treatment and elimination of malaria: Recommendations for evaluation of G6PD tests. Malar. J. 2013, 12, 391. [Google Scholar] [CrossRef]
- Alam, M.; Kibria, M.; Jahan, N.; Price, R.; Ley, B. Spectrophotometry assays to determine G6PD activity from Trinity Biotech and Pointe Scientific G6PD show good correlation. BMC Res. Notes 2018, 11, 855. [Google Scholar] [CrossRef]
- Pfeffer, D.; Ley, B.; Howes, R.; Adu, P.; Alam, M.; Bansil, P.; Boum, Y.; Brito, M.; Charoenkwan, P.; Clements, A.; et al. Quantification of glucose-6-phosphate dehydrogenase activity by spectrophotometry: A systematic review and meta-analysis. PLoS Med. 2020, 17, 1003081–1003084. [Google Scholar] [CrossRef]
- Ley, B.; Luter, N.; Espino, F.E.; Devine, A.; Kalnoky, M.; Lubell, Y.; Thriemer, K.; Baird, J.K.; Poirot, E.; Conan, N.; et al. The challenges of introducing routine G6PD testing into radical cure: A workshop report. Malar. J. 2015, 14, 377. [Google Scholar] [CrossRef] [Green Version]
- Ley, B.; Thriemer, K.; Jaswal, J.; Poirot, E.; Alam, M.S.; Phru, C.S.; Khan, W.A.; Dysoley, L.; Qi, G.; Kheong, C.C.; et al. Barriers to routine G6PD testing prior to treatment with primaquine. Malar. J. 2017, 16, 329. [Google Scholar] [CrossRef] [PubMed]
- White, N.J. Tafenoquine—A radical improvement? N. Engl. J. Med. 2019, 380, 285–286. [Google Scholar] [CrossRef] [PubMed]
- Alam, M.S.; Kibria, M.G.; Jahan, N.; Thriemer, K.; Hossain, M.S.; Douglas, N.M.; Phru, C.S.; Khan, W.A.; Price, R.N.; Ley, B. Field evaluation of quantitative point of care diagnostics to measure glucose-6-phosphate dehydrogenase activity. PLoS ONE 2018, 13, e0206331. [Google Scholar] [CrossRef] [PubMed]
- Ley, B.; Alam, M.S.; O’Donnell, J.J.; Hossain, M.S.; Kibria, M.G.; Jahan, N.; Khan, W.A.; Thriemer, K.; Chatfield, M.D.; Price, R.N.; et al. A Comparison of Three Quantitative Methods to Estimate G6PD Activity in the Chittagong Hill Tracts, Bangladesh. PLoS ONE 2017, 12, e0169930. [Google Scholar] [CrossRef]
- Martini, G.; Toniolo, D.; Vulliamy, T.; Luzzatto, L.; Dono, R.; Viglietto, G.; Paonessa, G.; d’Urso, M.; Persico, M. Structural analysis of the X-linked gene encoding human glucose 6-phosphate dehydrogenase. EMBO J. 1986, 5, 1849–1855. [Google Scholar] [CrossRef]
- Minucci, A.; Moradkhani, K.; Hwang, M.J.; Zuppi, C.; Giardina, B.; Capoluongo, E. Glucose-6-phosphate dehydrogenase (G6PD) mutations database: Review of the “old” and update of the new mutations. Blood Cells. Mol. Dis. 2012, 48, 154–165. [Google Scholar] [CrossRef]
- Gómez-Manzo, S.; Marcial-Quino, J.; Vanoye-Carlo, A.; Serrano-Posada, H.; Ortega-Cuellar, D.; González-Valdez, A.; Castillo-Rodríguez, R.A.; Hernández-Ochoa, B.; Sierra-Palacios, E.; Rodríguez-Bustamante, E.; et al. Glucose-6-phosphate dehydrogenase: Update and analysis of new mutations around the world. Int. J. Mol. Sci. 2016, 17, 2069. [Google Scholar] [CrossRef]
- Yoshida, A.; Beutler, E.; Motulsky, A.G. Human glucose-6-phosphate dehydrogenase variants. Bull. World Health Organ. 1971, 45, 243–253. [Google Scholar]
- WHO Working Group. Glucose-6-phosphate dehydrogenase deficiency. Bull. World Health Organ. 1989, 67, 601–611. [Google Scholar]
- WHO Global Malaria Programme Malaria Policy Advisory Group. Meeting Report of the Technical Consultation to Review the Classification of Glucose-6-Phosphate Dehydrogenase (G6PD); WHO/UCN/GMP/MPAG/2022.01; WHO Global Malaria Programme Malaria Policy Advisory Group: Geneva, Switzerland, 2022. [Google Scholar]
- Kwak, S.G.; Kim, J.H. Central limit theorem: The cornerstone of modern statistics. Korean J. Anesthesiol. 2017, 70, 144–156. [Google Scholar] [CrossRef]
- Tukey, J.W. Exploratory Data Analysis; Addison-Wesley: Boston, MA, USA, 1977. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing. Available online: https://www.R-project.org (accessed on 13 September 2022).
- Whiting, P.F.; Rutjes, A.W.; Westwood, M.E.; Mallett, S.; Deeks, J.J.; Reitsma, J.B.; Leeflang, M.M.; Sterne, J.A.; Bossuyt, P.M. QUADAS-2: A revised tool for the quality assessment of diagnostic accuracy studies. Ann. Intern. Med. 2011, 155, 529–536. [Google Scholar] [CrossRef] [PubMed]
- Bancone, G.; Chu, C.S.; Chowwiwat, N.; Somsakchaicharoen, R.; Wilaisrisak, P.; Charunwatthana, P.; Bansil, P.; McGray, S.; Domingo, G.J.; Nosten, F.H. Suitability of capillary blood for quantitative assessment of G6PD activity and performances of G6PD point-of-care tests. Am. J. Trop. Med. Hyg. 2015, 92, 818–824. [Google Scholar] [CrossRef] [PubMed]
- Bancone, G.; Chowwiwat, N.; Somsakchaicharoen, R.; Poodpanya, L.; Moo, P.K.; Gornsawun, G.; Kajeechiwa, L.; Thwin, M.M.; Rakthinthong, S.; Nosten, S.; et al. Single low dose Primaquine (0.25 mg/kg) does not cause clinically significant haemolysis in G6PD deficient subjects. PLoS ONE 2016, 11, e0151898. [Google Scholar] [CrossRef] [PubMed]
- Bancone, G.; Gilder, M.E.; Chowwiwat, N.; Gornsawun, G.; Win, E.; Cho, W.W.; Moo, E.; Min, A.M.; Charunwatthana, P.; Carrara, V.I.; et al. Prevalences of inherited red blood cell disorders in pregnant women of different ethnicities living along the Thailand-Myanmar border. Wellcome Open Res. 2017, 2, 72. [Google Scholar] [CrossRef]
- Brito, M.A.M.; Peixoto, H.M.; Almeida, A.C.G.d.; Oliveira, M.R.F.d.; Romero, G.A.S.; Moura-Neto, J.P.; Singh, N.; Monteiro, W.M.; Lacerda, M.V.G.d. Validation of the rapid test Carestart G6PD among malaria vivax-infected subjects in the Brazilian Amazon. Rev. Soc. Bras. Med. Trop. 2016, 49, 446–455. [Google Scholar] [CrossRef]
- Deng, Z.; Yang, F.; Bai, Y.; He, L.; Li, Q.; Wu, Y.; Luo, L.; Li, H.; Ma, L.; Yang, Z.; et al. Co-inheritance of glucose-6-phosphate dehydrogenase deficiency mutations and hemoglobin E in a Kachin population in a malaria-endemic region of Southeast Asia. PLoS ONE 2017, 12, e0177917. [Google Scholar] [CrossRef]
- Johnson, M.K.; Clark, T.D.; Njama-Meya, D.; Rosenthal, P.J.; Parikh, S. Impact of the method of G6PD deficiency assessment on genetic association studies of malaria susceptibility. PLoS ONE 2009, 4, e7246. [Google Scholar] [CrossRef]
- Kim, S.; Nguon, C.; Guillard, B.; Duong, S.; Chy, S.; Sum, S.; Nhem, S.; Bouchier, C.; Tichit, M.; Christophel, E.; et al. Performance of the CareStart™ G6PD deficiency screening test, a point-of-care diagnostic for primaquine therapy screening. PLoS ONE 2011, 6, e28357. [Google Scholar] [CrossRef]
- LaRue, N.; Kahn, M.; Murray, M.; Leader, B.T.; Bansil, P.; McGray, S.; Kalnoky, M.; Zhang, H.; Huang, H.; Jiang, H.; et al. Comparison of quantitative and qualitative tests for glucose-6-phosphate dehydrogenase deficiency. Am. J. Trop. Med. Hyg. 2014, 91, 854–861. [Google Scholar] [CrossRef]
- Ley, B.; Alam, M.S.; Kibria, M.G.; Marfurt, J.; Phru, C.S.; Ami, J.Q.; Thriemer, K.; Auburn, S.; Jahan, N.; Johora, F.T.; et al. Glucose-6-phosphate dehydrogenase activity in individuals with and without malaria: Analysis of clinical trial, cross-sectional and case-control data from Bangladesh. PLoS Med. 2021, 18, e1003576. [Google Scholar] [CrossRef]
- Pal, S.; Myburgh, J.; Bansil, P.; Hann, A.; Robertson, L.; Gerth-Guyette, E.; Ambler, G.; Bizilj, G.; Kahn, M.; Zobrist, S.; et al. Reference and point-of-care testing for G6PD deficiency: Blood disorder interference, contrived specimens, and fingerstick equivalence and precision. PLoS ONE 2021, 16, e0257560. [Google Scholar] [CrossRef] [PubMed]
- Reading, N.S.; Sirdah, M.M.; Shubair, M.E.; Nelson, B.E.; Al-Kahlout, M.S.; Al-Tayeb, J.M.; Aboud, L.N.; Shaban, M.A.; Luzzatto, L.; Prchal, J.T. Favism, the commonest form of severe hemolytic anemia in Palestinian children, varies in severity with three different variants of G6PD deficiency within the same community. Blood Cells Mol. Dis. 2016, 60, 58–64. [Google Scholar] [CrossRef] [PubMed]
- Roca-Feltrer, A.; Khim, N.; Kim, S.; Chy, S.; Canier, L.; Kerleguer, A.; Tor, P.; Chuor, C.M.; Kheng, S.; Siv, S.; et al. Field trial evaluation of the performances of point-of-care tests for screening G6PD deficiency in Cambodia. PLoS ONE 2014, 9, e116143. [Google Scholar] [CrossRef]
- Roh, M.E.; Oyet, C.; Orikiriza, P.; Wade, M.; Mwanga-Amumpaire, J.; Boum, Y., 2nd; Kiwanuka, G.N.; Parikh, S. Screening for Glucose-6-Phosphate Dehydrogenase Deficiency Using Three Detection Methods: A Cross-Sectional Survey in Southwestern Uganda. Am. J. Trop. Med. Hyg. 2016, 95, 1094–1099. [Google Scholar] [CrossRef] [PubMed]
- Satyagraha, A.W.; Sadhewa, A.; Baramuli, V.; Elvira, R.; Ridenour, C.; Elyazar, I.; Noviyanti, R.; Coutrier, F.N.; Harahap, A.R.; Baird, J.K. G6PD Deficiency at Sumba in Eastern Indonesia Is Prevalent, Diverse and Severe: Implications for Primaquine Therapy against Relapsing Vivax Malaria. PLoS Negl. Trop. Dis. 2015, 9, e0003602. [Google Scholar] [CrossRef]
- Satyagraha, A.W.; Sadhewa, A.; Elvira, R.; Elyazar, I.; Feriandika, D.; Antonjaya, U.; Oyong, D.; Subekti, D.; Rozi, I.E.; Domingo, G.J.; et al. Assessment of point-of-care diagnostics for G6PD deficiency in malaria endemic rural Eastern Indonesia. PLoS Negl. Trop. Dis. 2016, 10, e0004457. [Google Scholar] [CrossRef]
- Satyagraha, A.W.; Sadhewa, A.; Panggalo, L.V.; Subekti, D.; Elyazar, I.; Soebianto, S.; Mahpud, N.; Harahap, A.R.; Baird, J.K. Genotypes and phenotypes of G6PD deficiency among Indonesian females across diagnostic thresholds of G6PD activity guiding safe primaquine therapy of latent malaria. PLoS Negl. Trop. Dis. 2021, 15, e0009610. [Google Scholar] [CrossRef]
- Xia, Z.; Chen, P.; Tang, N.; Yan, T.; Zhou, Y.; Xiao, Q.; Huang, Q.; Li, Q. Rapid detection of G6PD mutations by multicolor melting curve analysis. Mol. Genet. Metab. 2016, 119, 168–173. [Google Scholar] [CrossRef]
- Ley, B.; Alam, M.S.; Satyagraha, A.W.; Phru, C.S.; Thriemer, K.; Tadesse, D.; Shibiru, T.; Hailu, A.; Kibria, M.G.; Hossain, M.S.; et al. Variation in Glucose-6-Phosphate Dehydrogenase activity following acute malaria. PLoS Negl. Trop. Dis. 2022, 16, e0010406. [Google Scholar] [CrossRef]
- Pal, S.; Bansil, P.; Bancone, G.; Hrutkay, S.; Kahn, M.; Gornsawun, G.; Penpitchaporn, P.; Chu, C.S.; Nosten, F.; Domingo, G.J. Evaluation of a Novel Quantitative Test for Glucose-6-Phosphate Dehydrogenase Deficiency: Bringing Quantitative Testing for Glucose-6-Phosphate Dehydrogenase Deficiency Closer to the Patient. Am. J. Trop. Med. Hyg. 2019, 100, 213–221. [Google Scholar] [CrossRef]
- Beutler, E.; Blume, K.G.; Kaplan, J.C.; Löhr, G.W.; Ramot, B.; Valentine, W.N. International Committee for Standardization in Haematology: Recommended Methods for Red-Cell Enzyme Analysis. Br. J. Haematol. 1977, 35, 331–340. [Google Scholar] [CrossRef] [PubMed]
- Thielemans, L.; Gornsawun, G.; Hanboonkunupakarn, B.; Paw, M.K.; Porn, P.; Moo, P.K.; Van Overmeire, B.; Proux, S.; Nosten, F.; McGready, R. Diagnostic performances of the fluorescent spot test for G6PD deficiency in newborns along the Thailand-Myanmar border: A cohort study. Wellcome Open Res. 2018, 3, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Travis, S.F.; Kumar, S.P.; Paez, P.C.; Delivoria-Papadopoulos, M. Red cell metabolic alterations in postnatal life in term infants. Glycolytic enzymes and glucose-6-phosphate dehydrogenase. Pediatric Res. 1980, 14, 1349. [Google Scholar] [CrossRef]
- Yang, W.-C.; Tai, S.; Hsu, C.-L.; Fu, C.-M.; Chou, A.-K.; Shao, P.-L.; Li, M.-J. Reference levels for glucose-6-phosphate dehydrogenase enzyme activity in infants 7–90 days old in Taiwan. J. Formos. Med. Assoc. 2020, 119, 69–74. [Google Scholar] [CrossRef]
- Los, E.; Ford, G.A. Klinefelter Syndrome. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2022. [Google Scholar]
- Minucci, A.; Giardina, B.; Zuppi, C.; Capoluongo, E. Glucose-6-phosphate dehydrogenase laboratory assay: How, when, and why? IUBMB Life 2008, 61, 27–34. [Google Scholar] [CrossRef]
- Brok-Simoni, F.; Ashkenazi, Y.E.; Ramot, B.; Holtzman, F. The diurnal rhythm of enzymes in human red cells. Br. J. Haematol. 1976, 32, 601–607. [Google Scholar] [CrossRef]
- Wolach, B.; Ashkenazi, M.; Grossmann, R.; Gavrieli, R.; Friedman, Z.; Bashan, N.; Roos, D. Diurnal fluctuation of leukocyte G6PD activity. A possible explanation for the normal neutrophil bactericidal activity and the low incidence of pyogenic infections in patients with severe G6PD deficiency in Israel. Pediatr. Res. 2004, 55, 807–813. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Hemi-/Homozygous * | Compound Heterozygous | Heterozygous * | |
|---|---|---|---|
| Sex | |||
| Female | 150 (8.6) | 20 (1.2) | 1012 (58.2) |
| Male | 556 (31.9) | - | - |
| Age | |||
| Mean (range) | 23.8 (1.0–78.0) | 23.2 (1.0–40.0) | 26.4 (1.0–75.0) |
| Country | |||
| Bangladesh | 94 (5.4) | - | 50 (2.9) |
| Brazil | 2 (0.1) | - | 0 (0.0) |
| Cambodia | 125 (7.2) | - | 66 (3.8) |
| China | 113 (6.5) | 12 (0.7) | 117 (6.7) |
| Indonesia | 78 (4.5) | 2 (0.1) | 273 (15.7) |
| Myanmar | 66 (3.8) | - | 66 (3.8) |
| Palestine | 64 (3.7) | - | 4 (0.2) |
| Thailand | 82 (4.7) | 5 (0.3) | 290 (16.7) |
| Uganda | 58 (3.3) | - | 79 (4.5) |
| USA | 24 (1.4) | 1 (0.1) | 67 (3.9) |
| Spectrophotometry Assay | |||
| Pointe Scientific | 7 (0.4) | - | 14 (0.8) |
| Randox | 194 (11.2) | - | 105 (6.0) |
| Trinity | 352 (20.3) | 4 (0.2) | 544 (31.3) |
| WHO Method | 40 (2.3) | 4 (0.2) | 232 (13.3) |
| Zhongshan Biotech | 113 (6.5) | 12 (0.7) | 117 (6.7) |
| Genotyping Assay | |||
| PCR | 374 (21.5) | 18 (1.0) | 742 (42.7) |
| Sequencing | 332 (19.1) | 2 (0.1) | 270 (15.5) |
| Total | 706 (40.6) | 20 (1.2) | 1012 (58.2) |
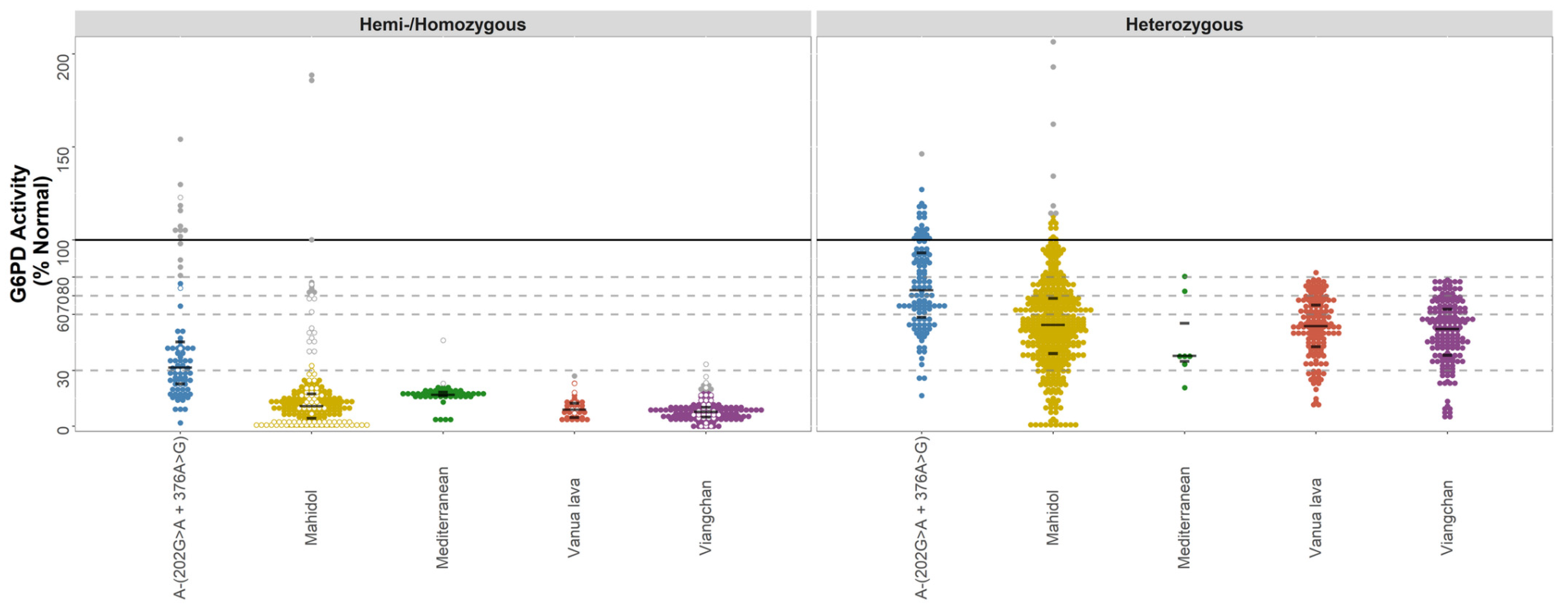
| Variant Name | n | G6PD Activity Estimate (%)* | Interquartile Range | Range, (Min–Max) | Outliers (n (Min-Max)) | |
|---|---|---|---|---|---|---|
| Data-rich Variants | A-(202A) c.376A>G + c.202G>A | |||||
| Hemi-/Homozygous | 67 | 29.0 | 20.1–38.1 | 1.7–76.6 | 14 (81.0–154.1) | |
| Heterozygous | 113 | 72.1 | 58.0–93.2 | 16.4–127.1 | 1 (146.3) | |
| Mahidol c.487G>A | ||||||
| Hemi-/Homozygous | 201 | 10.2 | 3.0–15.3 | 0.0–32.5 | 22 (39.8–188.6) | |
| Heterozygous | 381 | 54.4 | 38.8–68 | 0.0–112.8 | 7 (114.2–206.5) | |
| Mediterranean c.563C>T | ||||||
| Hemi-/Homozygous | 45 | 16.9 | 16.4–18.3 | 3.3–21.3 | 2 (23.0–46.0) | |
| Heterozygous | 7 | 37.9 | 35–55.3 | 20.7–80.5 | 0.0 | |
| Vanua Lava c.383T>C | ||||||
| Hemi-/Homozygous | 36 | 9.0 | 4.7–12.5 | 2.9–23.2 | 1 (27.0) | |
| Heterozygous | 140 | 53.8 | 42.8–65.1 | 10.9–82.5 | 0.0 | |
| Viangchan c.871G>A | ||||||
| Hemi-/Homozygous | 135 | 7.5 | 4.9–9.8 | 0–18.3 | 10 (19.2–33.3) | |
| Heterozygous * | 160 | 52.3 | 38.2–63 | 4.8–78.6 | 0.0 | |
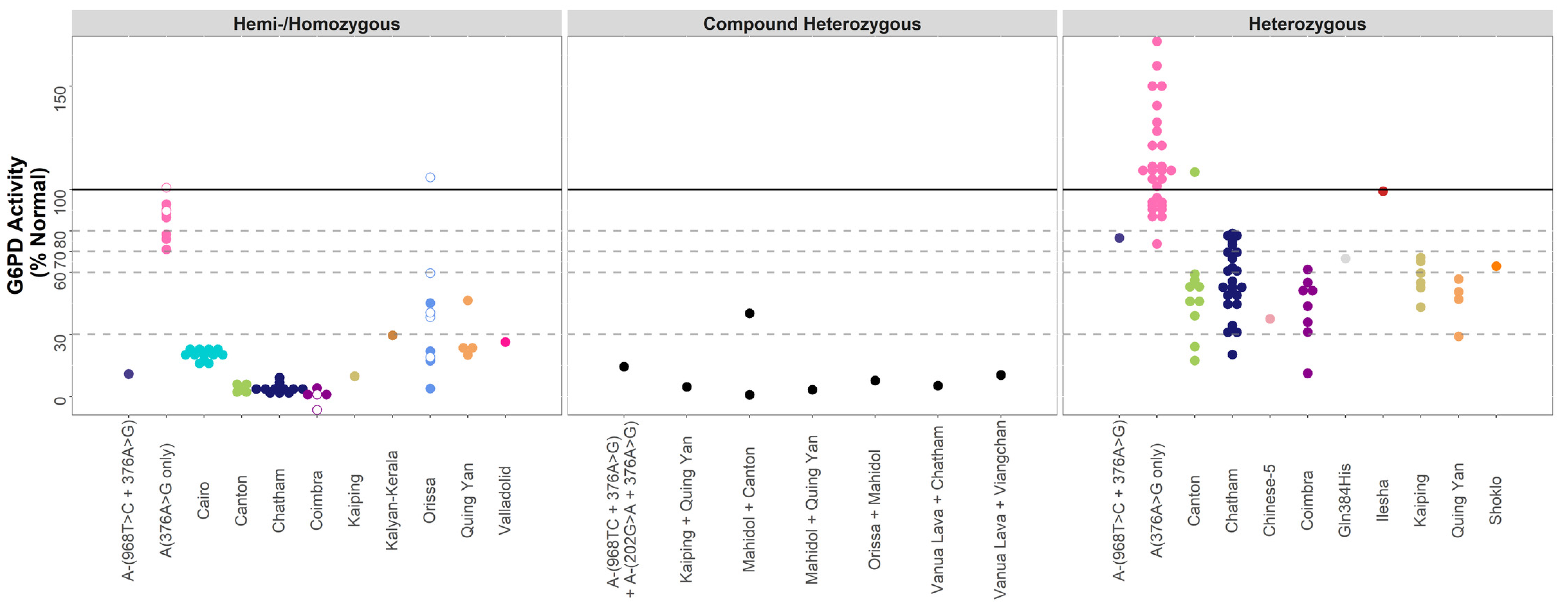
| Data-poor Variants | A-(968C) c. 376A>G + c.968T>C | |||||
| Hemi-/Homozygous | 1 | 10.9 | - | - | - | |
| Heterozygous | 1 | 76.6 | - | - | - | |
| A(376G only) c.376GA>G | ||||||
| Hemi-/Homozygous | 9 | 88.3 | 78.4–90.2 | 71.0–101 | - | |
| Heterozygous | 28 | 108.8 | 93.6–123.3 | 73.7–171.7 | - | |
| Cairo c.404A>C | ||||||
| Hemi-/Homozygous | 12 | 20.2 | 19.0–22.4 | 15.5–23.4 | - | |
| Canton c.1376G>T | ||||||
| Hemi-/Homozygous | 5 | 3.2 | 3.0–5.2 | 1.6–6.7 | - | |
| Heterozygous | 10 | 49.4 | 40.7–55.5 | 17.3–108.5 | - | |
| Chatham c.1003G>A | ||||||
| Hemi-/Homozygous | 11 | 3.4 | 2.8–3.9 | 1.2–9.1 | - | |
| Heterozygous | 25 | 53.2 | 44.6–68.9 | 20.2–78.8 | - | |
| Chinese-5 c.1024C>T | ||||||
| Heterozygous | 1 | 37.5 | - | - | - | |
| Coimbra c.592C>T | ||||||
| Hemi-/Homozygous | 6 | 0.9 | 0.5–1.6 | 0.0–4.2 | - | |
| Heterozygous | 8 | 47.3 | 34.6–52.2 | 11.2–61.3 | - | |
| Gln384Hisc.1152G>C | ||||||
| Heterozygous | 1 | 66.7 | - | - | - | |
| Ilesha c.466G>A | ||||||
| Heterozygous | 1 | 99.2 | - | - | - | |
| Kaiping c.1388G>A | ||||||
| Hemi-/Homozygous | 1 | 9.7 | - | - | - | |
| Heterozygous | 6 | 57.3 | 53.1–63.8 | 43.2–67.2 | - | |
| Kalyan-Kerala c.949G>A | ||||||
| Hemi-/Homozygous | 1 | 29.4 | - | - | - | |
| Orissa c.131C>G | ||||||
| Hemi-/Homozygous | 9 | 38.3 | 19–45.1 | 3.8–105.9 | - | |
| Quing Yan or Chinese-4 c.392G>T | ||||||
| Hemi-/Homozygous | 4 | 23.4 | 22.5–29.2 | 20.0–46.4 | - | |
| Heterozygous | 4 | 48.8 | 42.5–52.1 | 29.0–56.7 | - | |
| Shoklo c.701T>C | ||||||
| Heterozygous | 1 | 63 | - | - | - | |
| Valladolid c.406C>T | ||||||
| Hemi-/Homozygous | 1 | 26.2 | - | - | - | |
| A-(968C) + A-(202A) c. 376A>G + c.968T>C + c.202G>A | ||||||
| Compound Heterozygous | 1 | 14.3 | - | - | - | |
| Kaipingc.1388G>A + Quing Yan or Chinese-4 c.392G>T | ||||||
| Compound Heterozygous | 1 | 4.7 | - | - | - | |
| Mahidol c.487G>A + Cantonc.1376G>T | ||||||
| Compound Heterozygous | 2 | 20.5 | - | 0.9–40.1 | - | |
| Data-poor variants, Compound heterozygous | Mahidolc.487G>A + Quing Yanc.392G>T | |||||
| Compound Heterozygous | 1 | 3.3 | - | - | - | |
| Orissac.131C>G + Mahidolc.487G>A | ||||||
| Compound Heterozygous | 1 | 7.7 | - | - | - | |
| Vanua Lava c.383T>C + Chatham c.1003G>A | ||||||
| Compound Heterozygous | 1 | 5.3 | - | - | - | |
| Vanua Lava c.383T>C + Viangchan c.871G>A | ||||||
| Compound Heterozygous | 1 | 10.4 | - | - | - |
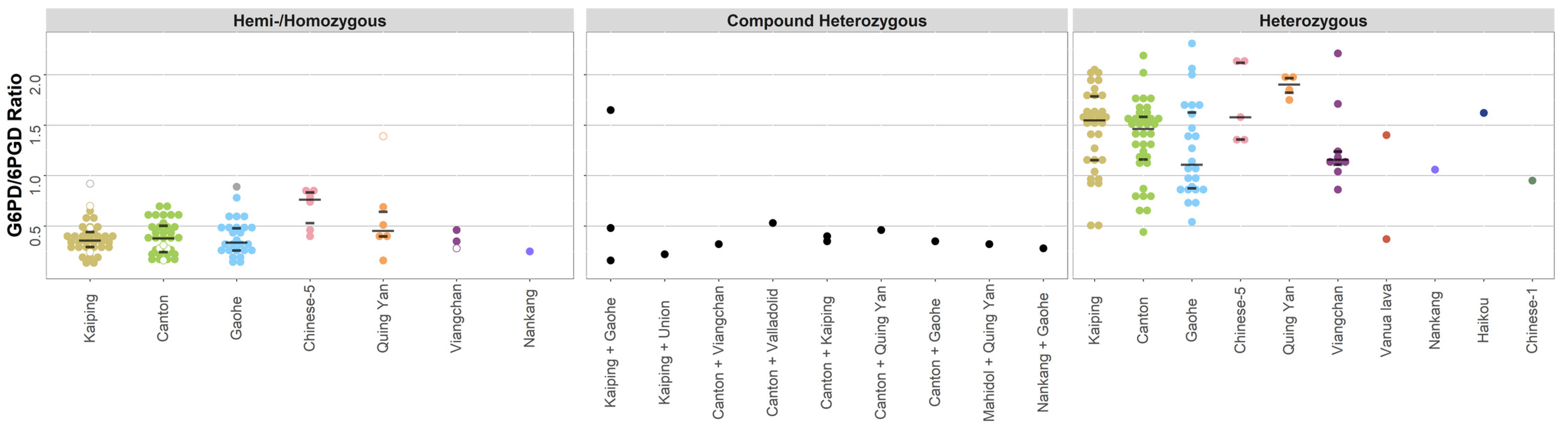
| Variant Name | n | G6PD Activity (G6PD/6PGD) * | Interquartile Range | Range, (Min–Max) | Outliers (n (Min–Max)) |
|---|---|---|---|---|---|
| Canton c.1376G>T | |||||
| Hemi-/Homozygous | 35 | 0.4 | 0.2–0.5 | 0.1–0.7 | - |
| Heterozygous | 36 | 1.5 | 1.2–1.6 | 0.4–2.2 | 2 (9.4–9.5) |
| Kaiping c.1388G>A | |||||
| Hemi-/Homozygous | 35 | 0.4 | 0.3–0.4 | 0.1–0.7 | 1 (0.9) |
| Heterozygous | 32 | 1.6 | 1.2–1.8 | 0.5–2 | - |
| Gaohe c.95A>G | |||||
| Hemi-/Homozygous | 25 | 0.3 | 0.3–0.5 | 0.1–0.8 | 1 (0.9) |
| Heterozygous | 24 | 1.1 | 0.9–1.6 | 0.5–2.3 | - |
| Chinese-1 c.835A>T | |||||
| Heterozygous | 1 | 0.9 | - | - | - |
| Chinese-5 c.1024C>T | |||||
| Hemi-/Homozygous | 6 | 0.8 | 0.5–0.8 | 0.4–0.8 | - |
| Heterozygous | 5 | 1.6 | 1.4–2.1 | 1.4–2.1 | - |
| Haikou c.835A>G | |||||
| Heterozygous | 1 | 1.6 | - | - | - |
| Nankang c.517T>C | |||||
| Hemi-/Homozygous | 1 | 0.2 | - | - | - |
| Heterozygous | 1 | 1.1 | - | - | - |
| Quing Yan or Chinese-4 c.392G>T | |||||
| Hemi-/Homozygous | 6 | 0.5 | 0.4–0.6 | 0.2–1.4 | - |
| Heterozygous | 4 | 1.9 | 1.8–2.0 | 1.8–2.0 | - |
| Vanua Lava c.383T>C | |||||
| Heterozygous | 2 | 0.9 | 0.6–1.1 | 0.4–1.4 | - |
| Viangchan c.871G>A | |||||
| Hemi-/Homozygous | 3 | 0.3 | 0.3–0.4 | 0.3–0.5 | - |
| Heterozygous | 9 | 1.2 | 1.1–1.2 | 0.9–2.2 | - |
| Canton c.1376G>T + Gaohe c.95A>G | |||||
| Compound Heterozygous | 1 | 0.3 | - | - | - |
| Canton c.1376G>T + Kaiping c.1388G>A | |||||
| Compound Heterozygous | 2 | 0.4 | 0.4–0.4 | 0.3–0.4 | - |
| Canton c.1376G>T + Quing Yan or Chinese-4 c.392G>T | |||||
| Compound Heterozygous | 1 | 0.5 | - | - | - |
| Canton c.1376G>T + Valladolid c.406C>T | |||||
| Compound Heterozygous | 1 | 0.5 | - | - | - |
| Canton c.1376G>T + Viangchan c.871G>A | |||||
| Hemi-/Homozygous | 1 | 0.3 | - | - | - |
| Kaiping c.1388G>A + Gaohe c.95A>G | |||||
| Compound Heterozygous | 3 | 0.5 | 0.3–1.1 | 0.2–1.6 | - |
| Kaiping c.1388G>A + Union c.1360C>T | |||||
| Compound Heterozygous | 1 | 0.2 | - | - | - |
| Mahidol c.487G>A + Quing Yan or Chinese-4 c.392G>T | |||||
| Compound Heterozygous | 1 | 0.3 | - | - | - |
| Nankang c.517T>C + Gaohe c.95A>G | |||||
| Compound Heterozygous | 1 | 0.3 | - | - | - |
| Variant | n * | Studies (n) | Number (%) Included Using Diagnostic Thresholds * | ||||
|---|---|---|---|---|---|---|---|
| <30% | <60% | <70% | <80% | ≥80% | |||
| A-(202A) c.376A>G + c.202G>A | |||||||
| Hemi-/Homozygous | 67 | 6 | 37 (55.2) | 64 (95.5) | 65 (97.0) | 67 (100.0) | 0 (0.0) |
| Heterozygous | 113 | 4 | 3 (2.7) | 29 (25.7) | 52 (46.0) | 67 (59.3) | 46 (40.7) |
| Mahidol c.487G>A | |||||||
| Hemi-/Homozygous | 201 | 5 | 200 (99.5) | 201 (100.0) | 201 (100.0) | 201 (100.0) | 0 (0.0) |
| Heterozygous | 381 | 7 | 56 (14.7) | 235 (61.7) | 296 (77.7) | 337 (88.5) | 44 (11.5) |
| Mediterranean c.563C>T | |||||||
| Hemi-/Homozygous | 45 | 3 | 45 (100.0) | 45 (100.0) | 45 (100.0) | 45 (100.0) | 0 (0.0) |
| Heterozygous | 7 | 4 | 1 (14.3) | 5 (71.4) | 5 (71.4) | 6 (85.7) | 1 (14.3) |
| Vanua Lava c.383T>C | |||||||
| Hemi-/Homozygous | 36 | 3 | 36 (100.0) | 36 (100.0) | 36 (100.0) | 36 (100.0) | 0 (0.0) |
| Heterozygous | 140 | 3 | 18 (12.9) | 91 (65.0) | 119 (85.0) | 139 (99.3) | 1 (0.7) |
| Viangchan c.871G>A | |||||||
| Hemi-/Homozygous | 135 | 6 | 135 (100.0) | 135 (100.0) | 135 (100.0) | 135 (100.0) | 0 (0.0) |
| Heterozygous | 160 | 6 | 19 (11.9) | 109 (68.1) | 141 (88.1) | 160 (100.0) | 0 (0.0) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pfeffer, D.A.; Satyagraha, A.W.; Sadhewa, A.; Alam, M.S.; Bancone, G.; Boum, Y., II; Brito, M.; Cui, L.; Deng, Z.; Domingo, G.J.; et al. Genetic Variants of Glucose-6-Phosphate Dehydrogenase and Their Associated Enzyme Activity: A Systematic Review and Meta-Analysis. Pathogens 2022, 11, 1045. https://doi.org/10.3390/pathogens11091045
Pfeffer DA, Satyagraha AW, Sadhewa A, Alam MS, Bancone G, Boum Y II, Brito M, Cui L, Deng Z, Domingo GJ, et al. Genetic Variants of Glucose-6-Phosphate Dehydrogenase and Their Associated Enzyme Activity: A Systematic Review and Meta-Analysis. Pathogens. 2022; 11(9):1045. https://doi.org/10.3390/pathogens11091045
Chicago/Turabian StylePfeffer, Daniel A., Ari Winasti Satyagraha, Arkasha Sadhewa, Mohammad Shafiul Alam, Germana Bancone, Yap Boum, II, Marcelo Brito, Liwang Cui, Zeshuai Deng, Gonzalo J. Domingo, and et al. 2022. "Genetic Variants of Glucose-6-Phosphate Dehydrogenase and Their Associated Enzyme Activity: A Systematic Review and Meta-Analysis" Pathogens 11, no. 9: 1045. https://doi.org/10.3390/pathogens11091045