
Genetic Diversity among Four Populations of Aedes aegypti (Diptera: Culicidae) from Honduras as Revealed by Mitochondrial DNA Cytochrome Oxidase I




Abstract
:1. Introduction
2. Results
2.1. Sequence Diversity of COI and ITS2
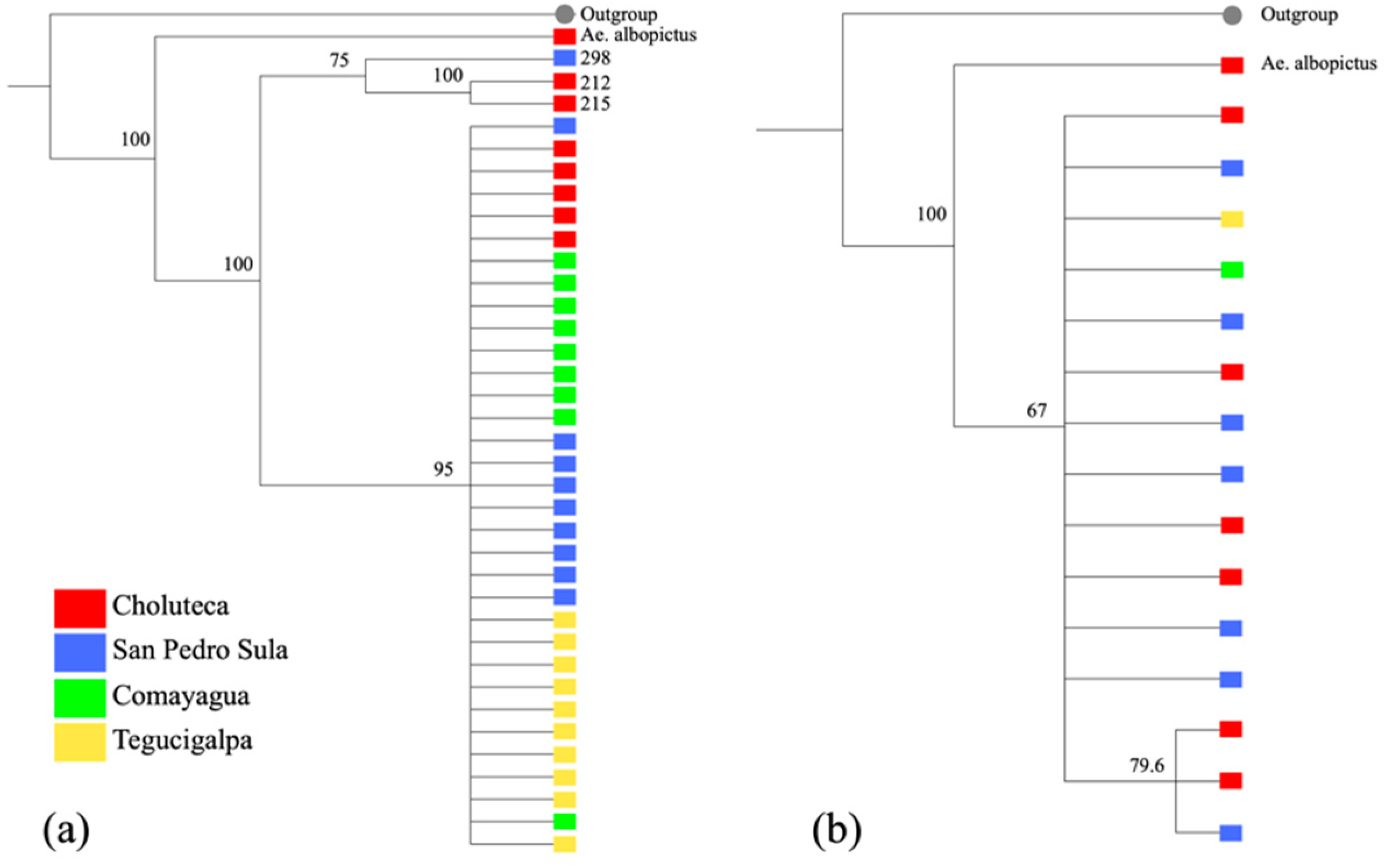
2.2. Phylogenetic Analysis Based on COI
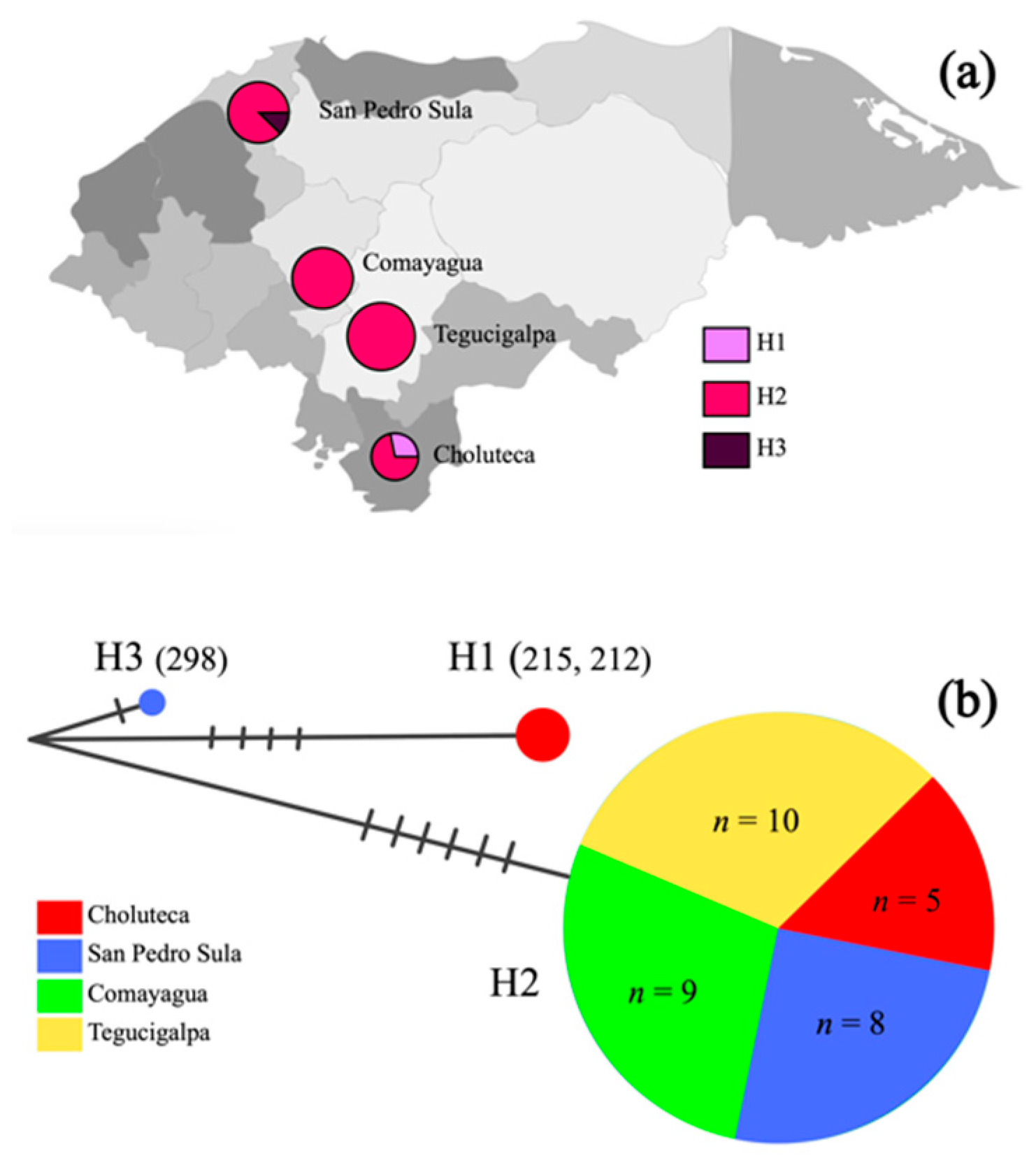
2.3. Mitochondrial COI Haplotype Analysis
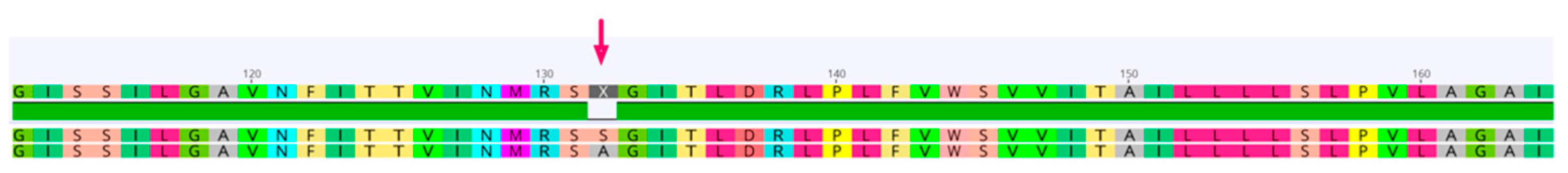
2.4. Amino Acid Sequence Analysis
3. Discussion
4. Materials and Methods
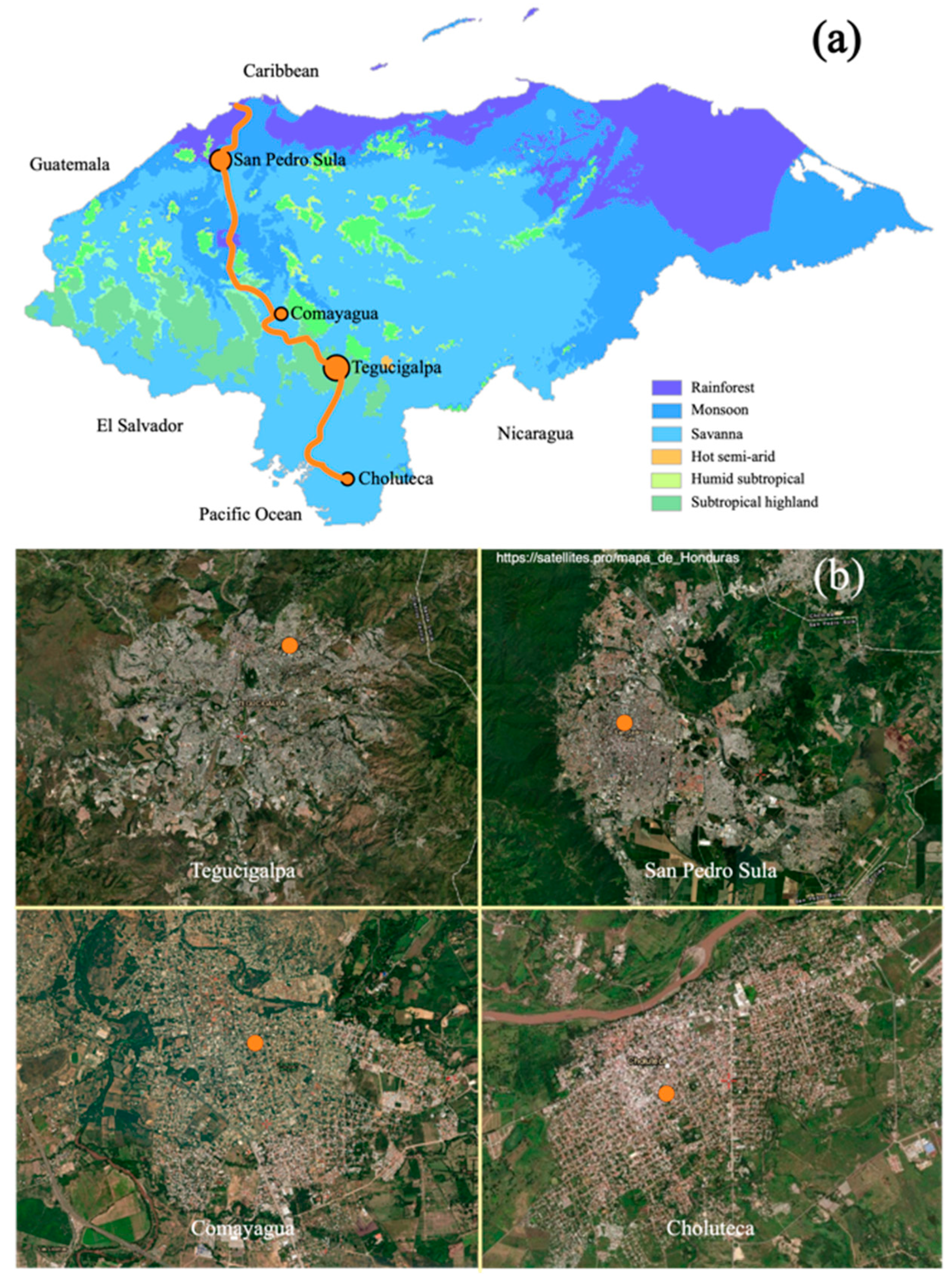
4.1. Study Sites, Mosquito Collection, and Morphological Identification
4.2. DNA Extraction, Gene Amplification, and Sequencing
4.3. Data Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Crawford, J.E.; Alves, J.M.; Palmer, W.J.; Day, J.P.; Sylla, M.; Ramasamy, R.; Surendran, S.N.; Black, W.C.T.; Pain, A.; Jiggins, F.M. Population genomics reveals that an anthropophilic population of Aedes aegypti mosquitoes in West Africa recently gave rise to American and Asian populations of this major disease vector. BMC Biol. 2017, 15, 16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brathwaite Dick, O.; San Martin, J.L.; Montoya, R.H.; del Diego, J.; Zambrano, B.; Dayan, G.H. The history of dengue outbreaks in the Americas. Am. J. Trop. Med. Hyg. 2012, 87, 584–593. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Espinal, M.A.; Andrus, J.K.; Jauregui, B.; Waterman, S.H.; Morens, D.M.; Santos, J.I.; Horstick, O.; Francis, L.A.; Olson, D. Emerging and Reemerging Aedes-Transmitted Arbovirus Infections in the Region of the Americas: Implications for Health Policy. Am. J. Public Health 2019, 109, 387–392. [Google Scholar] [CrossRef]
- Zambrano, L.I.; Rodriguez, E.; Espinoza-Salvado, I.A.; Rodriguez-Morales, A.J. Dengue in Honduras and the Americas: The epidemics are back! Travel Med. Infect Dis. 2019, 31, 101456. [Google Scholar] [CrossRef] [PubMed]
- Guzman, M.G.; Kouri, G. Dengue and dengue hemorrhagic fever in the Americas: Lessons and challenges. J. Clin. Virol. 2003, 27, 1–13. [Google Scholar] [CrossRef]
- Pan American Health Organization. PLISA Health Information Platform for the Americas. Available online: https://www3.paho.org/data/index.php/en/mnu-topics/indicadores-dengue-en/dengue-nacional-en/252-dengue-pais-ano-en.html (accessed on 21 April 2022).
- Pialoux, G.; Gauzere, B.A.; Jaureguiberry, S.; Strobel, M. Chikungunya, an epidemic arbovirosis. Lancet Infect Dis. 2007, 7, 319–327. [Google Scholar] [CrossRef]
- Gubler, D.J.; Vasilakis, N.; Musso, D. History and Emergence of Zika Virus. J. Infect Dis. 2017, 216, S860–S867. [Google Scholar] [CrossRef] [Green Version]
- Sherpa, S.; Rioux, D.; Goindin, D.; Fouque, F.; Francois, O.; Despres, L. At the Origin of a Worldwide Invasion: Unraveling the Genetic Makeup of the Caribbean Bridgehead Populations of the Dengue Vector Aedes aegypti. Genome Biol. Evol. 2018, 10, 56–71. [Google Scholar] [CrossRef] [Green Version]
- Steffler, L.M.; Dolabella, S.S.; Ribolla, P.E.; Dreyer, C.S.; Araujo, E.D.; Oliveira, R.G.; Martins, W.F.; La Corte, R. Genetic variability and spatial distribution in small geographic scale of Aedes aegypti (Diptera: Culicidae) under different climatic conditions in Northeastern Brazil. Parasit. Vectors 2016, 9, 530. [Google Scholar] [CrossRef] [Green Version]
- Jaimes-Duenez, J.; Arboleda, S.; Triana-Chavez, O.; Gomez-Palacio, A. Spatio-temporal distribution of Aedes aegypti (Diptera: Culicidae) mitochondrial lineages in cities with distinct dengue incidence rates suggests complex population dynamics of the dengue vector in Colombia. PLoS Negl. Trop Dis. 2015, 9, e0003553. [Google Scholar] [CrossRef]
- Monteiro, F.A.; Schama, R.; Martins, A.J.; Gloria-Soria, A.; Brown, J.E.; Powell, J.R. Genetic diversity of Brazilian Aedes aegypti: Patterns following an eradication program. PLoS Negl. Trop Dis. 2014, 8, e3167. [Google Scholar] [CrossRef] [Green Version]
- Maitra, A.; Cunha-Machado, A.S.; Souza Leandro, A.; Costa, F.M.D.; Scarpassa, V.M. Exploring deeper genetic structures: Aedes aegypti in Brazil. Acta Trop. 2019, 195, 68–77. [Google Scholar] [CrossRef] [PubMed]
- Souza, K.R.; Ribeiro, G., Jr.; Silva dos Santos, C.G.; de Lima, E.C.; Melo, P.R.; Reis, M.G.; Blanton, R.E.; Silva, L.K. Vector control measures failed to affect genetic structure of Aedes aegypti in a sentinel metropolitan area of Brazil. Acta Trop. 2013, 128, 598–605. [Google Scholar] [CrossRef]
- de Lourdes Munoz, M.; Mercado-Curiel, R.F.; Diaz-Badillo, A.; Perez Ramirez, G.; Black, W.C.T. Gene flow pattern among Aedes aegypti populations in Mexico. J. Am. Mosq. Control Assoc. 2013, 29, 1–18. [Google Scholar] [CrossRef]
- Paupy, C.; Le Goff, G.; Brengues, C.; Guerra, M.; Revollo, J.; Barja Simon, Z.; Herve, J.P.; Fontenille, D. Genetic structure and phylogeography of Aedes aegypti, the dengue and yellow-fever mosquito vector in Bolivia. Infect. Genet. Evol. 2012, 12, 1260–1269. [Google Scholar] [CrossRef] [PubMed]
- Duenas, J.C.; Llinas, G.A.; Panzetia-Dutari, G.M.; Gardenal, C.N. Two different routes of colonization of Aedes aegypti in Argentina from neighboring countries. J. Med. Entomol. 2009, 46, 1344–1354. [Google Scholar] [CrossRef]
- Lima, R.S., Jr.; Scarpassa, V.M. Evidence of two lineages of the dengue vector Aedes aegypti in the Brazilian Amazon, based on mitochondrial DNA ND4 gene sequences. Genet. Mol. Biol. 2009, 32, 414–422. [Google Scholar] [CrossRef] [Green Version]
- Scarpassa, V.M.; Cardoza, T.B.; Cardoso Junior, R.P. Population genetics and phylogeography of Aedes aegypti (Diptera: Culicidae) from Brazil. Am. J. Trop. Med. Hyg. 2008, 78, 895–903. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Joyce, A.L.; Torres, M.M.; Torres, R.; Moreno, M. Genetic variability of the Aedes aegypti (Diptera: Culicidae) mosquito in El Salvador, vector of dengue, yellow fever, chikungunya and Zika. Parasit. Vectors 2018, 11, 637. [Google Scholar] [CrossRef] [Green Version]
- Garcia-Franco, F.; Munoz Mde, L.; Lozano-Fuentes, S.; Fernandez-Salas, I.; Garcia-Rejon, J.; Beaty, B.J.; Black, W.C.T. Large genetic distances among Aedes aegypti populations along the South Pacific coast of Mexico. Am. J. Trop. Med. Hyg. 2002, 66, 594–598. [Google Scholar] [CrossRef] [Green Version]
- Gloria-Soria, A.; Ayala, D.; Bheecarry, A.; Calderon-Arguedas, O.; Chadee, D.D.; Chiappero, M.; Coetzee, M.; Elahee, K.B.; Fernandez-Salas, I.; Kamal, H.A.; et al. Global genetic diversity of Aedes aegypti. Mol. Ecol. 2016, 25, 5377–5395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eskildsen, G.A.; Rovira, J.R.; Smith, O.; Miller, M.J.; Bennett, K.L.; McMillan, W.O.; Loaiza, J. Maternal invasion history of Aedes aegypti and Aedes albopictus into the Isthmus of Panama: Implications for the control of emergent viral disease agents. PLoS ONE 2018, 13, e0194874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kawada, H.; Futami, K.; Higa, Y.; Rai, G.; Suzuki, T.; Rai, S.K. Distribution and pyrethroid resistance status of Aedes aegypti and Aedes albopictus populations and possible phylogenetic reasons for the recent invasion of Aedes aegypti in Nepal. Parasit. Vectors 2020, 13, 213. [Google Scholar] [CrossRef] [PubMed]
- Khater, E.I.M.; Baig, F.; Kamal, H.A.; Powell, J.R.; Saleh, A.A. Molecular Phylogenetics and Population Genetics of the Dengue Vector Aedes aegypti From the Arabian Peninsula. J. Med. Entomol. 2021, 58, 2161–2176. [Google Scholar] [CrossRef]
- Dharmarathne, H.; Weerasena, O.; Perera, K.; Galhena, G.H. Genetic characterization of Aedes aegypti (Diptera: Culicidae) in Sri Lanka based on COI gene. J. Vector Borne Dis. 2020, 57, 153–160. [Google Scholar] [CrossRef]
- Bracco, J.E.; Capurro, M.L.; Lourenco-de-Oliveira, R.; Sallum, M.A. Genetic variability of Aedes aegypti in the Americas using a mitochondrial gene: Evidence of multiple introductions. Mem. Inst. Oswaldo Cruz 2007, 102, 573–580. [Google Scholar] [CrossRef] [Green Version]
- Moore, M.; Sylla, M.; Goss, L.; Burugu, M.W.; Sang, R.; Kamau, L.W.; Kenya, E.U.; Bosio, C.; Munoz Mde, L.; Sharakova, M.; et al. Dual African origins of global Aedes aegypti s.l. populations revealed by mitochondrial DNA. PLoS Negl. Trop. Dis. 2013, 7, e2175. [Google Scholar] [CrossRef] [Green Version]
- Bennett, K.L.; Shija, F.; Linton, Y.M.; Misinzo, G.; Kaddumukasa, M.; Djouaka, R.; Anyaele, O.; Harris, A.; Irish, S.; Hlaing, T.; et al. Historical environmental change in Africa drives divergence and admixture of Aedes aegypti mosquitoes: A precursor to successful worldwide colonization? Mol. Ecol. 2016, 25, 4337–4354. [Google Scholar] [CrossRef] [Green Version]
- Kumar, M.S.; Kalimuthu, M.; Selvam, A.; Mathivanan, A.; Paramasivan, R.; Kumar, A.; Gupta, B. Genetic structure and connectivity among Aedes aegypti populations within Madurai city in Southern India. Infect. Genet. Evol. 2021, 95, 105031. [Google Scholar] [CrossRef]
- Abuelmaali, S.A.; Jamaluddin, J.A.F.; Noaman, K.; Allam, M.; Abushama, H.M.; Elnaiem, D.E.; Ishak, I.H.; Wajidi, M.F.F.; Jaal, Z.; Abu Kassim, N.F. Distribution and Genetic Diversity of Aedes aegypti Subspecies across the Sahelian Belt in Sudan. Pathogens 2021, 10, 78. [Google Scholar] [CrossRef]
- Calvez, E.; Guillaumot, L.; Millet, L.; Marie, J.; Bossin, H.; Rama, V.; Faamoe, A.; Kilama, S.; Teurlai, M.; Mathieu-Daude, F.; et al. Genetic Diversity and Phylogeny of Aedes aegypti, the Main Arbovirus Vector in the Pacific. PLoS Negl. Trop. Dis. 2016, 10, e0004374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naim, D.M.; Kamal, N.Z.M.; Mahboob, S. Population structure and genetic diversity of Aedes aegypti and Aedes albopictus in Penang as revealed by mitochondrial DNA cytochrome oxidase I. Saudi J. Biol. Sci. 2020, 27, 953–967. [Google Scholar] [CrossRef] [PubMed]
- Carvajal, T.M.; Ogishi, K.; Yaegeshi, S.; Hernandez, L.F.T.; Viacrusis, K.M.; Ho, H.T.; Amalin, D.M.; Watanabe, K. Fine-scale population genetic structure of dengue mosquito vector, Aedes aegypti, in Metropolitan Manila, Philippines. PLoS Negl. Trop. Dis. 2020, 14, e0008279. [Google Scholar] [CrossRef] [PubMed]
- Tedjou, A.N.; Kamgang, B.; Yougang, A.P.; Njiokou, F.; Wondji, C.S. Update on the geographical distribution and prevalence of Aedes aegypti and Aedes albopictus (Diptera: Culicidae), two major arbovirus vectors in Cameroon. PLoS Negl. Trop. Dis. 2019, 13, e0007137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Powell, J.R.; Tabachnick, W.J. History of domestication and spread of Aedes aegypti—A review. Mem. Inst. Oswaldo Cruz 2013, 108 (Suppl. 1), 11–17. [Google Scholar] [CrossRef]
- Sabbatani, S.; Fiorino, S. Yellow fever. Infez. Med. 2007, 15, 129–141. [Google Scholar]
- Miller, M.J.; Loaiza, J.R. Geographic expansion of the invasive mosquito Aedes albopictus across Panama—Implications for control of dengue and Chikungunya viruses. PLoS Negl. Trop. Dis. 2015, 9, e0003383. [Google Scholar] [CrossRef] [Green Version]
- Fonzi, E.; Higa, Y.; Bertuso, A.G.; Futami, K.; Minakawa, N. Human-Mediated Marine Dispersal Influences the Population Structure of Aedes aegypti in the Philippine Archipelago. PLoS Negl. Trop. Dis. 2015, 9, e0003829. [Google Scholar] [CrossRef] [Green Version]
- Ponce, P.; Munoz-Tobar, S.; Carrazco-Montalvo, A.; Villota, S.D.; Coloma, J.; Wang, C.; Holechek, S.; Cevallos, V. Two Haplotypes of Aedes aegypti Detected by ND4 Mitochondrial Marker in Three Regions of Ecuador. Insects 2021, 12, 200. [Google Scholar] [CrossRef]
- Lv, R.C.; Zhu, C.; Wang, C.H.; Ai, L.L.; Lv, H.; Zhang, B.; Li, C.M.; An, J.; Wang, P.G.; Hu, D.; et al. Genetic diversity and population structure of Aedes aegypti after massive vector control for dengue fever prevention in Yunnan border areas. Sci. Rep. 2020, 10, 12731. [Google Scholar] [CrossRef]
- Shi, Q.M.; Zhang, H.D.; Wang, G.; Guo, X.X.; Xing, D.; Dong, Y.D.; Xiao, L.; Gao, J.; Liu, Q.M.; Sun, A.J.; et al. The genetic diversity and population structure of domestic Aedes aegypti (Diptera: Culicidae) in Yunnan Province, southwestern China. Parasit. Vectors 2017, 10, 292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yugavathy, N.; Kim-Sung, L.; Joanne, S.; Vythilingam, I. Genetic variation of the mitochondrial genes, CO1 and ND5, in Aedes aegypti from various regions of peninsular Malaysia. Trop. Biomed. 2016, 33, 543–560. [Google Scholar] [PubMed]
- Duong, C.V.; Kang, J.H.; Nguyen, V.V.; Bae, Y.J. Genetic Diversity and Population Structure of the Asian Tiger Mosquito (Aedes albopictus) in Vietnam: Evidence for Genetic Differentiation by Climate Region. Genes 2021, 12, 1579. [Google Scholar] [CrossRef]
- Campos, M.; Ward, D.; Morales, R.F.; Gomes, A.R.; Silva, K.; Sepulveda, N.; Gomez, L.F.; Clark, T.G.; Campino, S. Surveillance of Aedes aegypti populations in the city of Praia, Cape Verde: Zika virus infection, insecticide resistance and genetic diversity. Parasit. Vectors 2020, 13, 481. [Google Scholar] [CrossRef]
- Brown, J.E.; McBride, C.S.; Johnson, P.; Ritchie, S.; Paupy, C.; Bossin, H.; Lutomiah, J.; Fernandez-Salas, I.; Ponlawat, A.; Cornel, A.J.; et al. Worldwide patterns of genetic differentiation imply multiple ‘domestications’ of Aedes aegypti, a major vector of human diseases. Proc. Biol. Sci. 2011, 278, 2446–2454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garjito, T.A.; Widiarti, W.; Hidajat, M.C.; Handayani, S.W.; Mujiyono, M.; Prihatin, M.T.; Ubaidillah, R.; Sudomo, M.; Satoto, T.B.T.; Manguin, S.; et al. Homogeneity and Possible Replacement of Populations of the Dengue Vectors Aedes aegypti and Aedes albopictus in Indonesia. Front Cell Infect. Microbiol. 2021, 11, 705129. [Google Scholar] [CrossRef]
- Garcia-Rejon, J.E.; Navarro, J.C.; Cigarroa-Toledo, N.; Baak-Baak, C.M. An Updated Review of the Invasive Aedes albopictus in the Americas; Geographical Distribution, Host Feeding Patterns, Arbovirus Infection, and the Potential for Vertical Transmission of Dengue Virus. Insects 2021, 12, 967. [Google Scholar] [CrossRef]
- Beck, H.E.; Zimmermann, N.E.; McVicar, T.R.; Vergopolan, N.; Berg, A.; Wood, E.F. Present and future Koppen-Geiger climate classification maps at 1-km resolution. Sci. Data 2018, 5, 180214. [Google Scholar] [CrossRef] [Green Version]
- Polson, A.K.; Curtis, C.M.S.O.J.; Chantha, N.; Rawlins, S. The Use of Ovitraps Baited with Hay Infusion as a Surveillance Tool for Aedes aegypti Mosquitoes in Cambodia; WHO Regional Office for South-East Asia: New Delhi, India, 2002. [Google Scholar]
- Proyecto ZIKA AIRS, Z. Manual Práctico para el Funcionamiento de un Insectario: Procedimientos de Cría de la Especie Aedes aegypti y Principios Básicos de Bioseguridad; Abt Associates: Rockville, MD, USA, 2019; p. 91. [Google Scholar]
- Rueda, L. Pictorial keys for the identification of mosquitoes (Diptera: Culicidae) associated with Dengue Virus Transmission. Zootaxa 2004, 589. [Google Scholar] [CrossRef]
- Folmer, O.; Black, M.; Hoeh, W.; Lutz, R.; Vrijenhoek, R. DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Mol. Mar. Biol. Biotechnol. 1994, 3, 294–299. [Google Scholar]
- Djadid, N.D.; Gholizadeh, S.; Aghajari, M.; Zehi, A.H.; Raeisi, A.; Zakeri, S. Genetic analysis of rDNA-ITS2 and RAPD loci in field populations of the malaria vector, Anopheles stephensi (Diptera: Culicidae): Implications for the control program in Iran. Acta Trop. 2006, 97, 65–74. [Google Scholar] [CrossRef] [PubMed]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Behura, S.K.; Lobo, N.F.; Haas, B.; de Bruyn, B.; Lovin, D.D.; Shumway, M.F.; Puiu, D.; Romero-Severson, J.; Nene, V.; Severson, D.W. Complete sequences of mitochondria genomes of Aedes aegypti and Culex quinquefasciatus and comparative analysis of mitochondrial DNA fragments inserted in the nuclear genomes. Insect Biochem. Mol. Biol. 2011, 41, 770–777. [Google Scholar] [CrossRef] [Green Version]
- Stecher, G.; Tamura, K.; Kumar, S. Molecular Evolutionary Genetics Analysis (MEGA) for macOS. Mol. Biol. Evol. 2020, 37, 1237–1239. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef] [PubMed]
- Rozas, J.; Ferrer-Mata, A.; Sanchez-Del Barrio, J.C.; Guirao-Rico, S.; Librado, P.; Ramos-Onsins, S.E.; Sanchez-Gracia, A. DnaSP 6: DNA Sequence Polymorphism Analysis of Large Data Sets. Mol. Biol. Evol. 2017, 34, 3299–3302. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Statistics | Honduras | Region 1 | World 2 |
|---|---|---|---|
| Length (bp) | 505 | 352 | 450 |
| Number of sequences | 35 | 217 | 670 |
| Identical sites (%) | 493 (97.6%) | 288 (81.8%) | 323 (71.8%) |
| Pairwise % Identity | 99.7% | 98.9% | 99.3% |
| Overall mean distance | 0.00 | 0.01 | 0.01 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Escobar, D.; Ortiz, B.; Urrutia, O.; Fontecha, G. Genetic Diversity among Four Populations of Aedes aegypti (Diptera: Culicidae) from Honduras as Revealed by Mitochondrial DNA Cytochrome Oxidase I. Pathogens 2022, 11, 620. https://doi.org/10.3390/pathogens11060620
Escobar D, Ortiz B, Urrutia O, Fontecha G. Genetic Diversity among Four Populations of Aedes aegypti (Diptera: Culicidae) from Honduras as Revealed by Mitochondrial DNA Cytochrome Oxidase I. Pathogens. 2022; 11(6):620. https://doi.org/10.3390/pathogens11060620
Chicago/Turabian StyleEscobar, Denis, Bryan Ortiz, Oscar Urrutia, and Gustavo Fontecha. 2022. "Genetic Diversity among Four Populations of Aedes aegypti (Diptera: Culicidae) from Honduras as Revealed by Mitochondrial DNA Cytochrome Oxidase I" Pathogens 11, no. 6: 620. https://doi.org/10.3390/pathogens11060620