
Metabolic Response to Tick-Borne Encephalitis Virus Infection and Bacterial Co-Infections

 ,
, 
Abstract
:1. Introduction
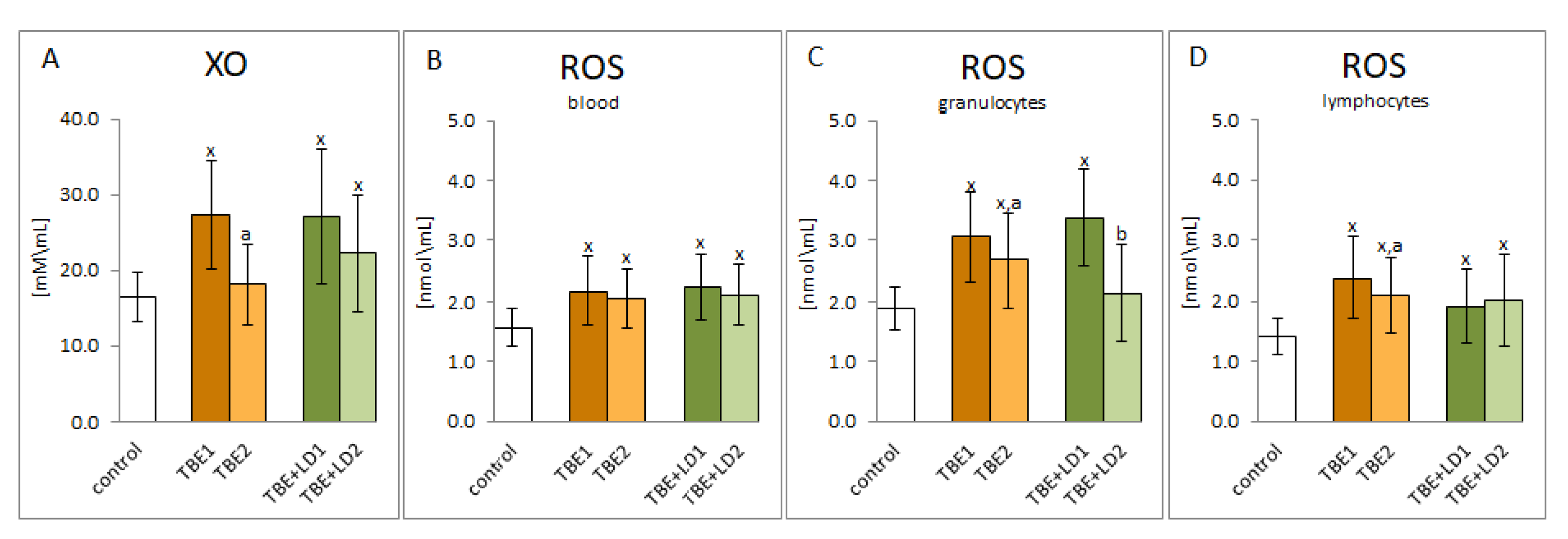
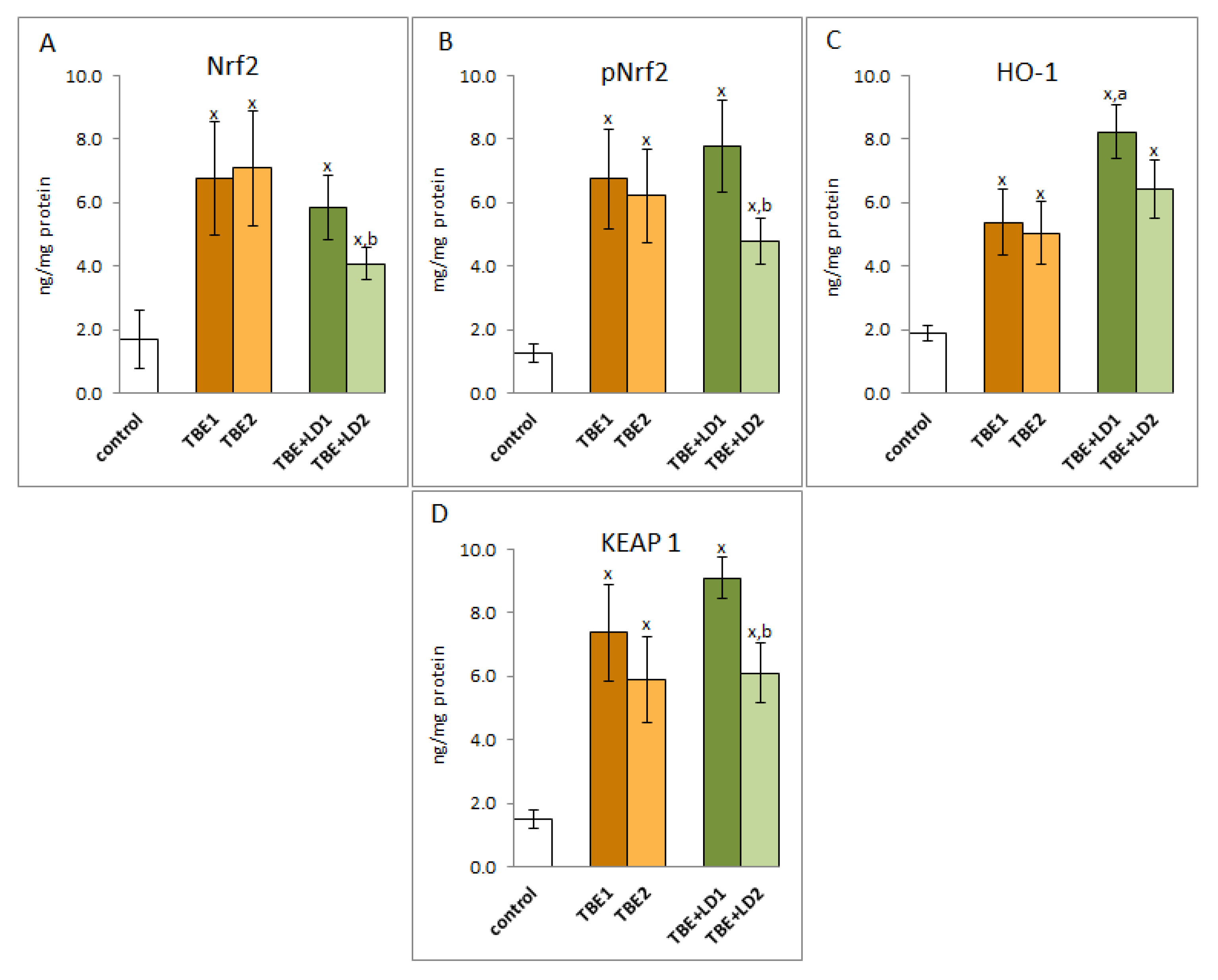
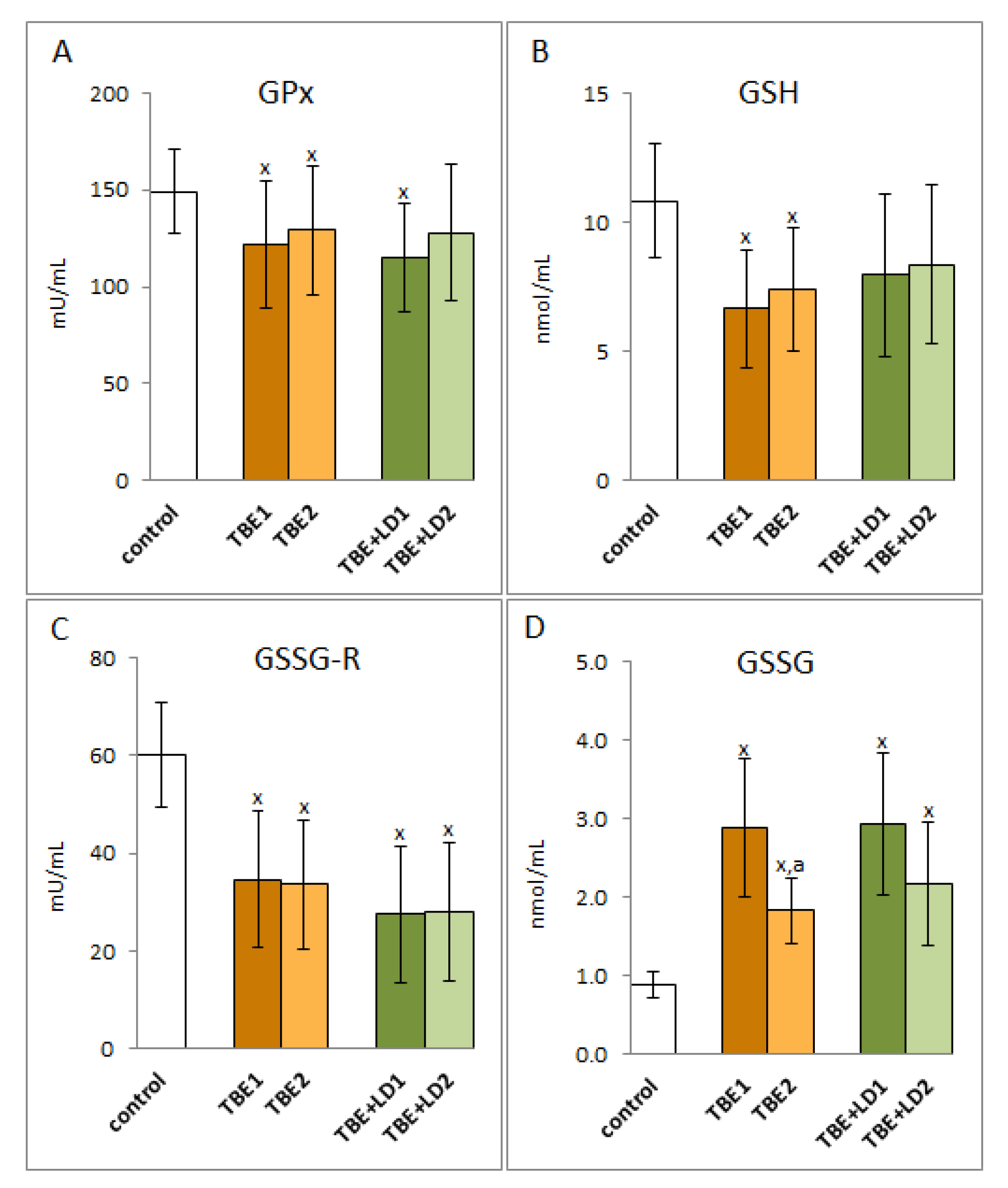
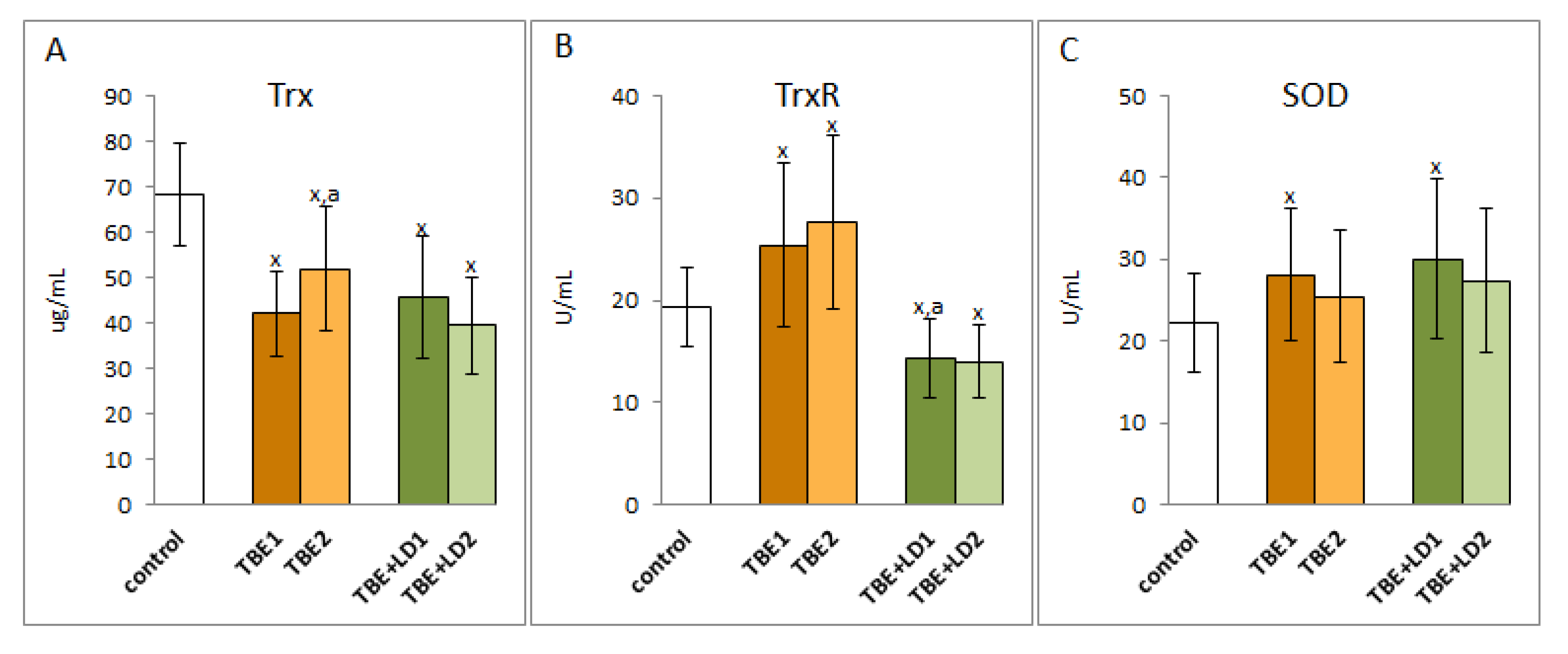
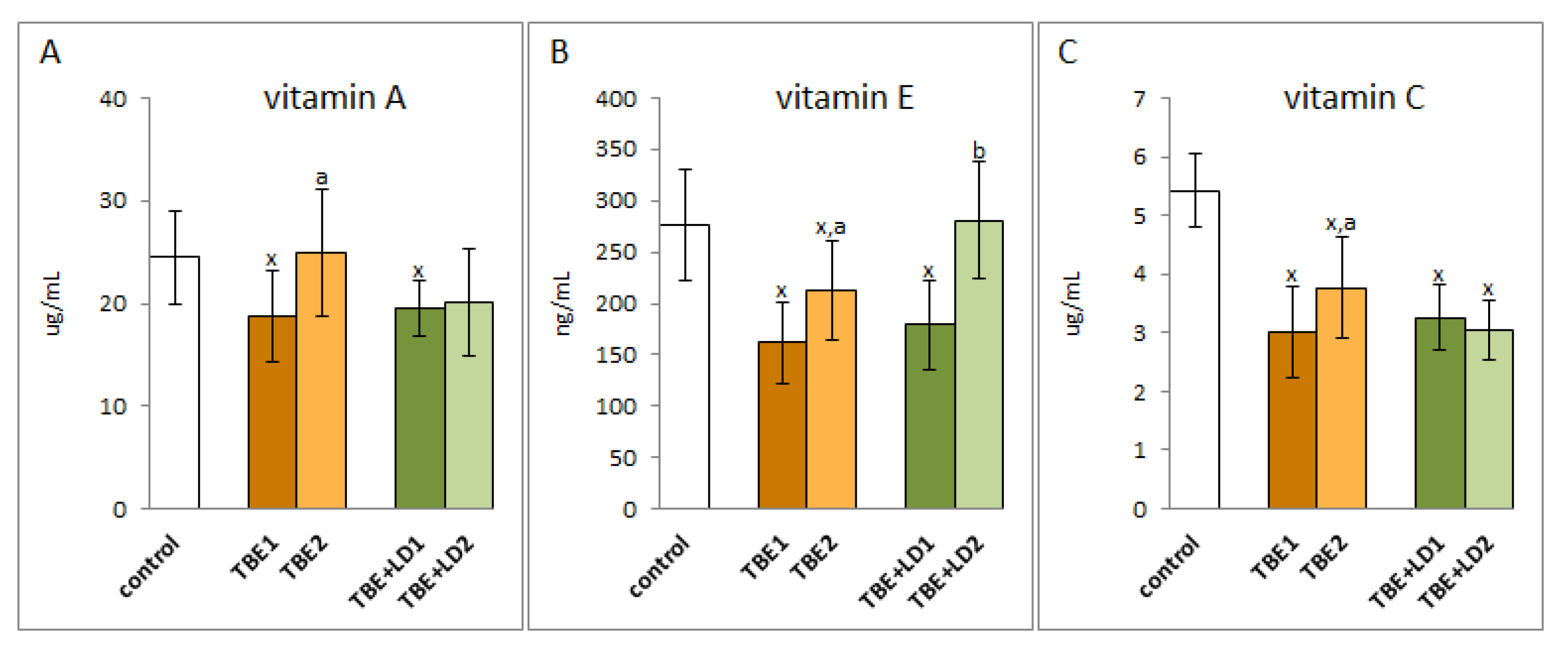
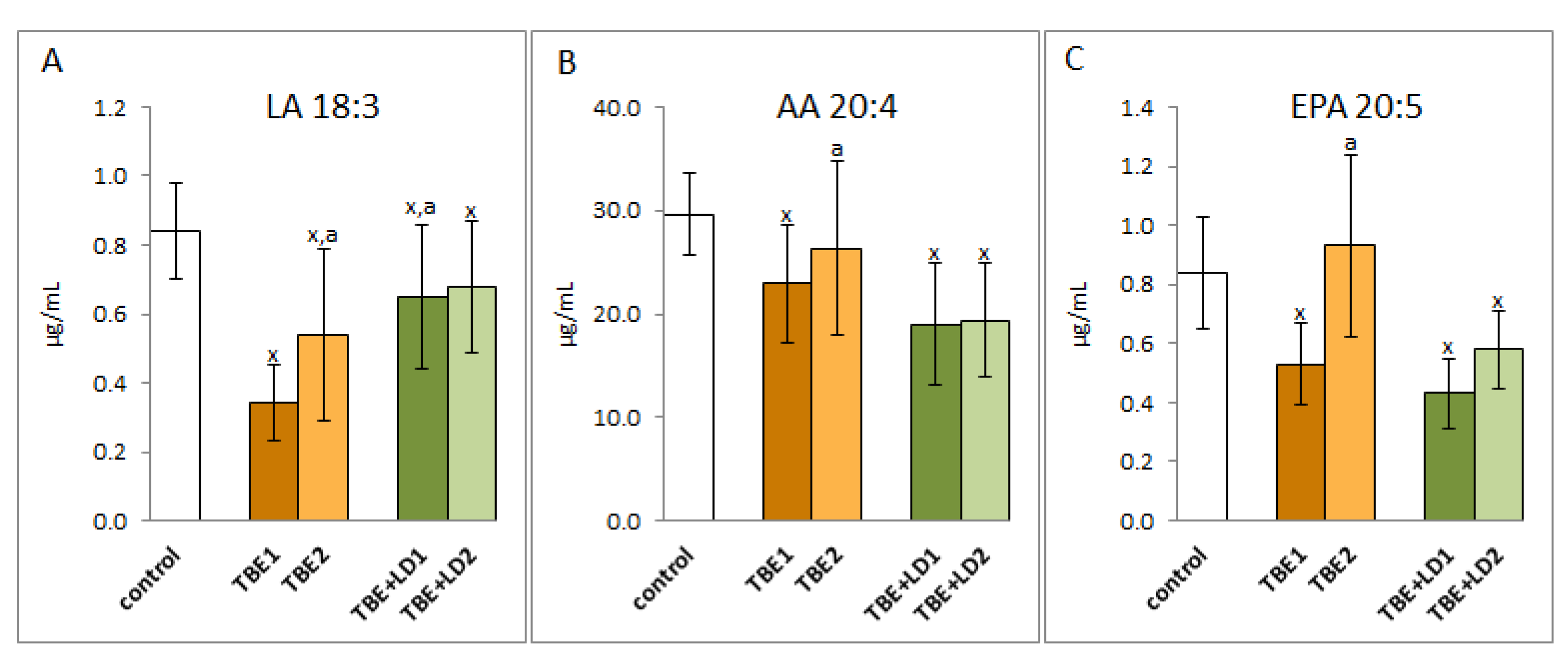
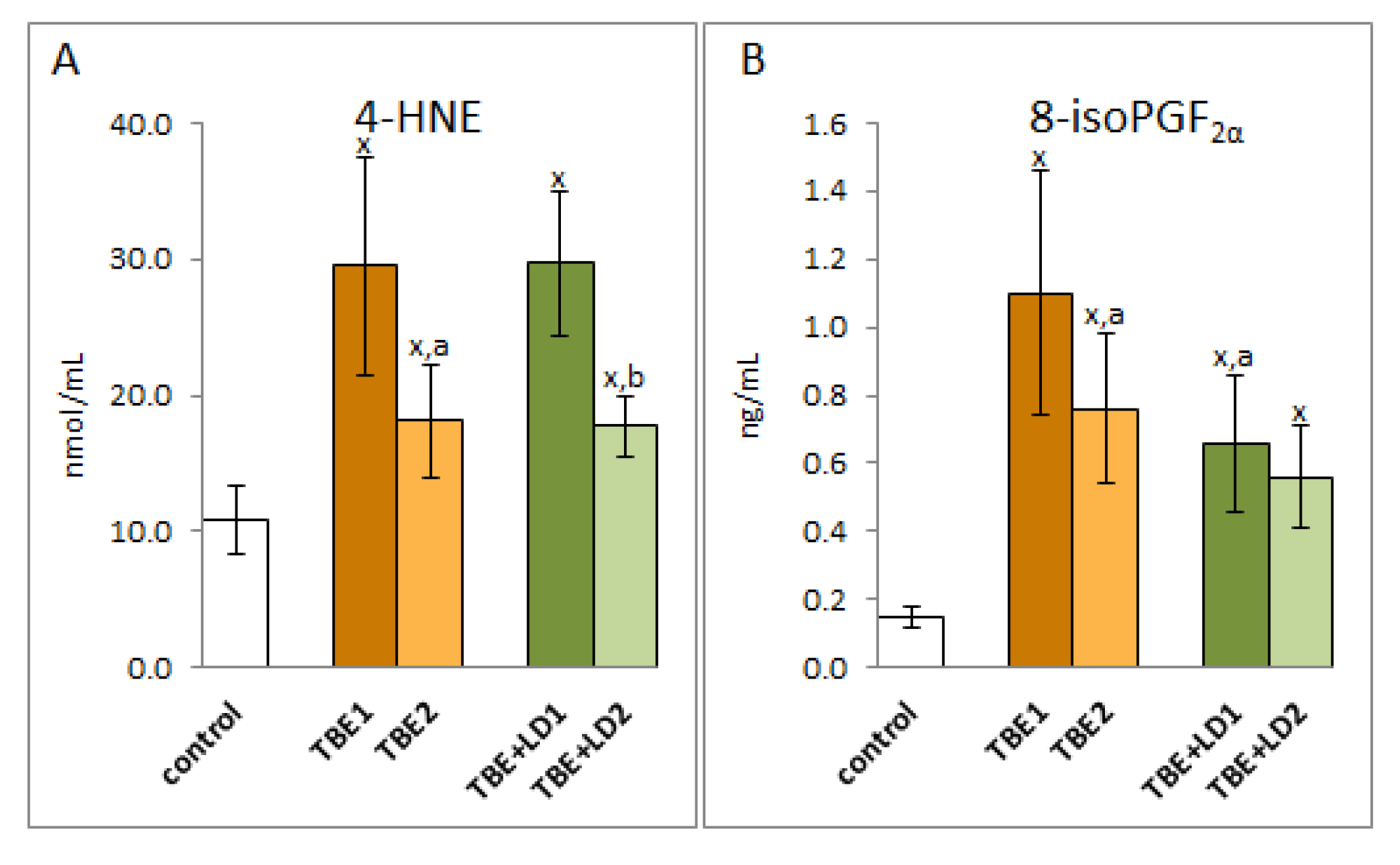
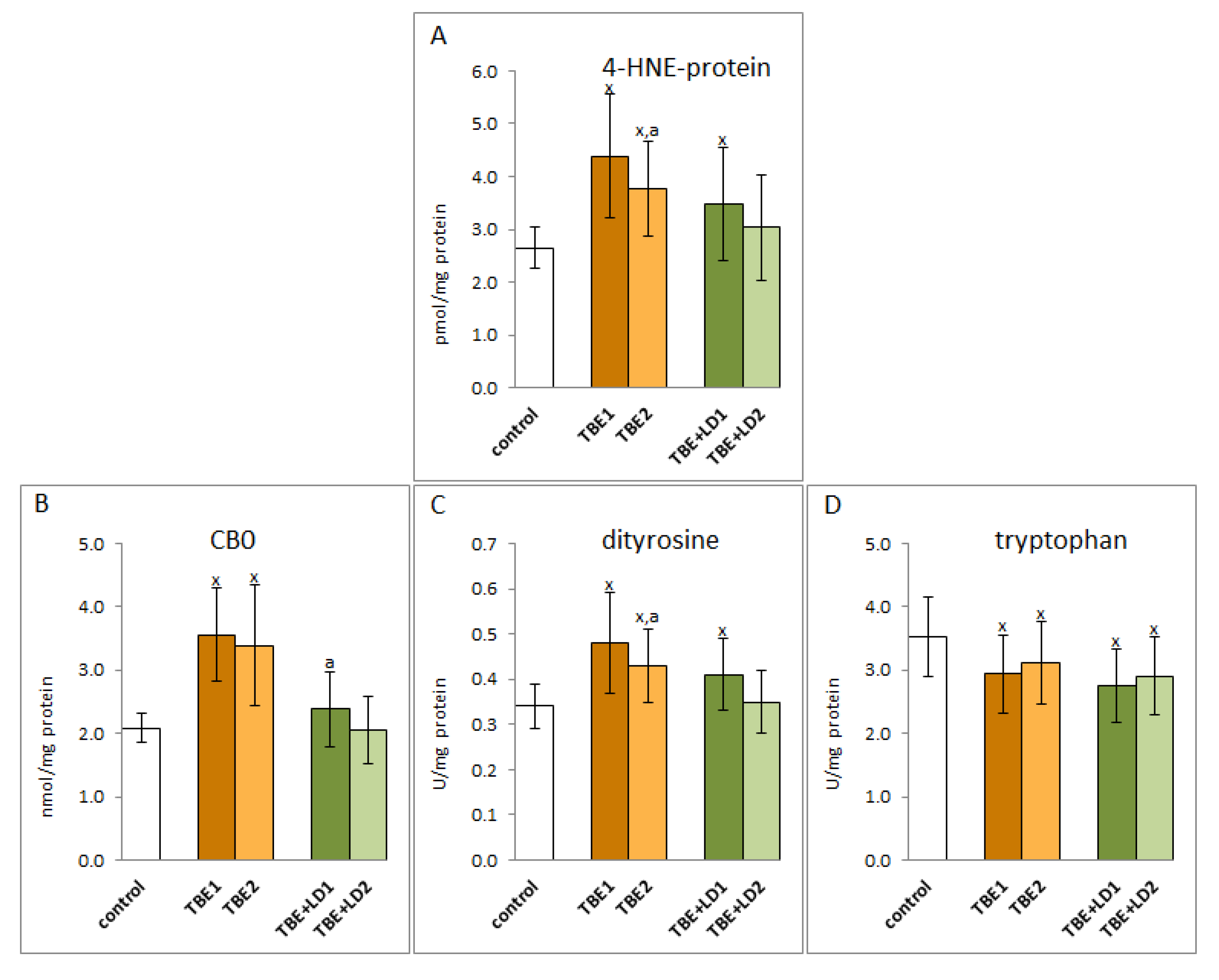
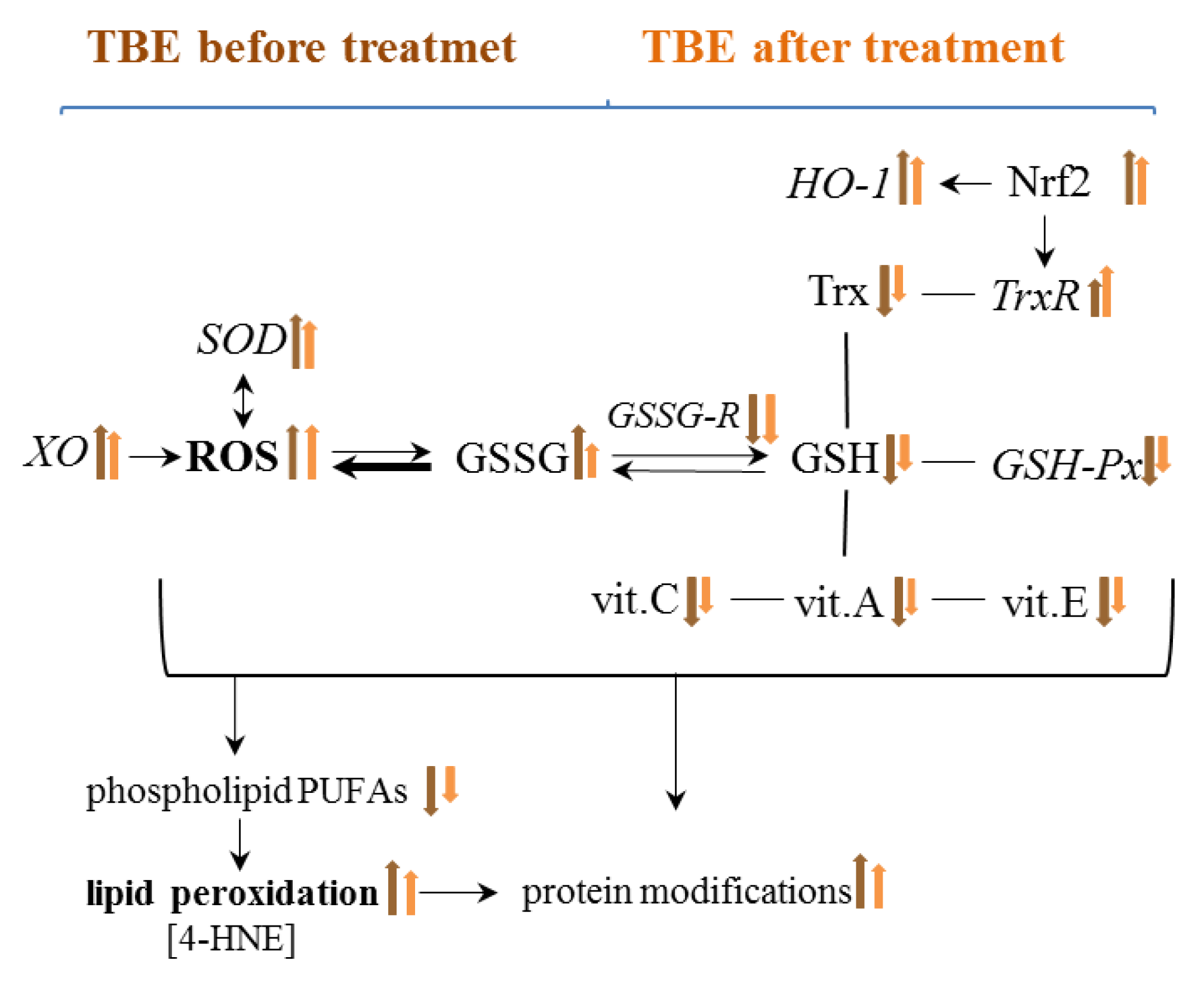
2. Results
3. Discussion
4. Materials and Methods
4.1. Samples Collection
4.2. Methods
4.2.1. Pro-Oxidant Parameters
Pro-Oxidant Enzyme Activity
ROS Level Determination
4.2.2. Antioxidant Parameters
Determination of Protein Antioxidants
Determination of Low Molecular Antioxidants
4.2.3. Phospholipid Metabolism
Phospholipid Fatty Acids Profile
Lipid Peroxidation Products
4.2.4. Determination of Protein Expression
4.2.5. Determination of Protein Oxidative Modifications
4.2.6. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zhang, Z.; Rong, L.; Li, Y.-P. Flaviviridae Viruses and Oxidative Stress: Implications for Viral Pathogenesis. Oxid. Med. Cell. Longev. 2019, 2019, 1409582. [Google Scholar] [CrossRef] [Green Version]
- Boulanger, N.; Boyer, P.; Talagrand-Reboul, E.; Hansmann, Y. Ticks and Tick-Borne Diseases. Med. Mal. Infect. 2019, 49, 87–97. [Google Scholar] [CrossRef] [PubMed]
- Kuzmenko, Y.V.; Smirnova, O.A.; Ivanov, A.V.; Starodubova, E.S.; Karpov, V.L. Nonstructural Protein 1 of Tick-Borne Encephalitis Virus Induces Oxidative Stress and Activates Antioxidant Defense by the Nrf2/ARE Pathway. Intervirology 2016, 59, 111–117. [Google Scholar] [CrossRef] [PubMed]
- Chung, K.M.; Liszewski, M.K.; Nybakken, G.; Davis, A.E.; Townsend, R.R.; Fremont, D.H.; Atkinson, J.P.; Diamond, M.S. West Nile Virus Nonstructural Protein NS1 Inhibits Complement Activation by Binding the Regulatory Protein Factor H. Proc. Natl. Acad. Sci. USA 2006, 103, 19111–19116. [Google Scholar] [CrossRef] [Green Version]
- Ruzek, D.; Avšič Županc, T.; Borde, J.; Chrdle, A.; Eyer, L.; Karganova, G.; Kholodilov, I.; Knap, N.; Kozlovskaya, L.; Matveev, A.; et al. Tick-Borne Encephalitis in Europe and Russia: Review of Pathogenesis, Clinical Features, Therapy, and Vaccines. Antivir. Res. 2019, 164, 23–51. [Google Scholar] [CrossRef] [PubMed]
- Kubes, M.; Fuchsberger, N.; Labuda, M.; Zuffová, E.; Nuttall, P.A. Salivary Gland Extracts of Partially Fed Dermacentor Reticulatus Ticks Decrease Natural Killer Cell Activity In Vitro. Immunology 1994, 82, 113–116. [Google Scholar]
- Bogovič, P.; Lusa, L.; Korva, M.; Pavletič, M.; Resman Rus, K.; Lotrič-Furlan, S.; Avšič-Županc, T.; Strle, K.; Strle, F. Inflammatory Immune Responses in the Pathogenesis of Tick-Borne Encephalitis. J. Clin. Med. 2019, 8, 731. [Google Scholar] [CrossRef] [Green Version]
- Palus, M.; Bílý, T.; Elsterová, J.; Langhansová, H.; Salát, J.; Vancová, M.; Růžek, D. Infection and Injury of Human Astrocytes by Tick-Borne Encephalitis Virus. J. Gen. Virol. 2014, 95, 2411–2426. [Google Scholar] [CrossRef]
- Griffiths, H.R.; Gao, D.; Pararasa, C. Redox Regulation in Metabolic Programming and Inflammation. Redox Biol. 2017, 12, 50–57. [Google Scholar] [CrossRef]
- Ye, Z.-W.; Zhang, J.; Townsend, D.M.; Tew, K.D. Oxidative Stress, Redox Regulation and Diseases of Cellular Differentiation. Biochim. Biophys. Acta 2015, 1850, 1607–1621. [Google Scholar] [CrossRef] [Green Version]
- Komaravelli, N.; Casola, A. Respiratory Viral Infections and Subversion of Cellular Antioxidant Defenses. J. Pharmacogenom. Pharmacoproteom. 2014, 5, 1000141. [Google Scholar] [CrossRef]
- Shastri, M.D.; Shukla, S.D.; Chong, W.C.; Dua, K.; Peterson, G.M.; Patel, R.P.; Hansbro, P.M.; Eri, R.; O’Toole, R.F. Role of Oxidative Stress in the Pathology and Management of Human Tuberculosis. Oxid. Med. Cell. Longev. 2018, 2018, e7695364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Georgescu, S.R.; Mitran, C.I.; Mitran, M.I.; Caruntu, C.; Sarbu, M.I.; Matei, C.; Nicolae, I.; Tocut, S.M.; Popa, M.I.; Tampa, M. New Insights in the Pathogenesis of HPV Infection and the Associated Carcinogenic Processes: The Role of Chronic Inflammation and Oxidative Stress. J. Immunol. Res. 2018, 2018, 5315816. [Google Scholar] [CrossRef] [Green Version]
- Ivanov, A.V.; Bartosch, B.; Isaguliants, M.G. Oxidative Stress in Infection and Consequent Disease. Oxid. Med. Cell. Longev. 2017, 2017, e3496043. [Google Scholar] [CrossRef] [PubMed]
- Pohanka, M. Role of Oxidative Stress in Infectious Diseases. A Review. Folia Microbiol. 2013, 58, 503–513. [Google Scholar] [CrossRef]
- Novaes, R.D.; Teixeira, A.L.; de Miranda, A.S. Oxidative Stress in Microbial Diseases: Pathogen, Host, and Therapeutics. Oxid. Med. Cell. Longev. 2019, 2019, e8159562. [Google Scholar] [CrossRef]
- Nathan, C.; Cunningham-Bussel, A. Beyond Oxidative Stress: An Immunologist’s Guide to Reactive Oxygen Species. Nat. Rev. Immunol. 2013, 13, 349–361. [Google Scholar] [CrossRef] [Green Version]
- Mullen, L.; Mengozzi, M.; Hanschmann, E.-M.; Alberts, B.; Ghezzi, P. How the Redox State Regulates Immunity. Free Radic. Biol. Med. 2020, 157, 3–14. [Google Scholar] [CrossRef]
- Du, Y.; Mi, Z.; Xie, Y.; Lu, D.; Zheng, H.; Sun, H.; Zhang, M.; Niu, Y. Insights into the Molecular Basis of Tick-Borne Encephalitis from Multiplatform Metabolomics. PLoS Negl. Trop. Dis. 2021, 15, e0009172. [Google Scholar] [CrossRef]
- Lim, J.K.M.; Leprivier, G. The Impact of Oncogenic RAS on Redox Balance and Implications for Cancer Development. Cell Death Dis. 2019, 10, 955. [Google Scholar] [CrossRef]
- Trachootham, D.; Lu, W.; Ogasawara, M.A.; Nilsa, R.-D.V.; Huang, P. Redox Regulation of Cell Survival. Antioxid. Redox Signal. 2008, 10, 1343–1374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, B.W.L.; Ghode, P.; Ong, D.S.T. Redox Regulation of Cell State and Fate. Redox Biol. 2019, 25, 101056. [Google Scholar] [CrossRef] [PubMed]
- Casas, A.I.; Nogales, C.; Mucke, H.A.M.; Petraina, A.; Cuadrado, A.; Rojo, A.I.; Ghezzi, P.; Jaquet, V.; Augsburger, F.; Dufrasne, F.; et al. On the Clinical Pharmacology of Reactive Oxygen Species. Pharmacol. Rev. 2020, 72, 801–828. [Google Scholar] [CrossRef] [PubMed]
- Belikov, A.V.; Schraven, B.; Simeoni, L. T Cells and Reactive Oxygen Species. J. Biomed. Sci. 2015, 22, 85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dunnill, C.; Patton, T.; Brennan, J.; Barrett, J.; Dryden, M.; Cooke, J.; Leaper, D.; Georgopoulos, N.T. Reactive Oxygen Species (ROS) and Wound Healing: The Functional Role of ROS and Emerging ROS-Modulating Technologies for Augmentation of the Healing Process. Int. Wound J. 2017, 14, 89–96. [Google Scholar] [CrossRef] [PubMed]
- Filomeni, G.; Desideri, E.; Cardaci, S.; Rotilio, G.; Ciriolo, M.R. Under the ROS: Thiol Network Is the Principal Suspect for Autophagy Commitment. Autophagy 2010, 6, 999–1005. [Google Scholar] [CrossRef] [Green Version]
- Amatore, D.; Sgarbanti, R.; Aquilano, K.; Baldelli, S.; Limongi, D.; Civitelli, L.; Nencioni, L.; Garaci, E.; Ciriolo, M.R.; Palamara, A.T. Influenza Virus Replication in Lung Epithelial Cells Depends on Redox-Sensitive Pathways Activated by NOX4-Derived ROS. Cell. Microbiol. 2015, 17, 131–145. [Google Scholar] [CrossRef] [Green Version]
- Zal, F.; Mostafavi-Pour, Z.; Amini, F.; Heidari, A. Effect of Vitamin E and C Supplements on Lipid Peroxidation and GSH-Dependent Antioxidant Enzyme Status in the Blood of Women Consuming Oral Contraceptives. Contraception 2012, 86, 62–66. [Google Scholar] [CrossRef]
- Ayala, A.; Muñoz, M.F.; Argüelles, S. Lipid Peroxidation: Production, Metabolism, and Signaling Mechanisms of Malondialdehyde and 4-Hydroxy-2-Nonenal. Oxid. Med. Cell. Longev. 2014, 2014, 360438. [Google Scholar] [CrossRef]
- Miller, E.; Morel, A.; Saso, L.; Saluk, J. Isoprostanes and Neuroprostanes as Biomarkers of Oxidative Stress in Neurodegenerative Diseases. Oxid. Med. Cell. Longev. 2014, 2014, 572491. [Google Scholar] [CrossRef]
- Muhammad, Y.; Aminu, Y.K.; Ahmad, A.E.; Iliya, S.; Muhd, N.; Yahaya, M.; Mustapha, A.S.; Tahiru, A.; Abdulkadir, S.S.; Ibrahim, J.S.; et al. An Elevated 8-Isoprostaglandin F2 Alpha (8-Iso-PGF2α) in COVID-19 Subjects Co-Infected with Malaria. Pan Afr. Med. J. 2020, 37, 78. [Google Scholar] [CrossRef]
- Zheng, L.; Fei, J.; Feng, C.-M.; Xu, Z.; Fu, L.; Zhao, H. Serum 8-Iso-PGF2α Predicts the Severity and Prognosis in Patients With Community-Acquired Pneumonia: A Retrospective Cohort Study. Front. Med. 2021, 8, 633442. [Google Scholar] [CrossRef] [PubMed]
- Barrera, G.; Pizzimenti, S.; Ciamporcero, E.S.; Daga, M.; Ullio, C.; Arcaro, A.; Cetrangolo, G.P.; Ferretti, C.; Dianzani, C.; Lepore, A.; et al. Role of 4-Hydroxynonenal-Protein Adducts in Human Diseases. Antioxid. Redox Signal. 2015, 22, 1681–1702. [Google Scholar] [CrossRef] [Green Version]
- Gęgotek, A.; Skrzydlewska, E. Biological Effect of Protein Modifications by Lipid Peroxidation Products. Chem. Phys. Lipids 2019, 221, 46–52. [Google Scholar] [CrossRef] [PubMed]
- Pohl, E.E.; Jovanovic, O. The Role of Phosphatidylethanolamine Adducts in Modification of the Activity of Membrane Proteins under Oxidative Stress. Molecules 2019, 24, 4545. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bauer, G.; Zarkovic, N. Revealing Mechanisms of Selective, Concentration-Dependent Potentials of 4-Hydroxy-2-Nonenal to Induce Apoptosis in Cancer Cells through Inactivation of Membrane-Associated Catalase. Free Radic. Biol. Med. 2015, 81, 128–144. [Google Scholar] [CrossRef] [PubMed]
- Fang, C.; Zhang, H.; Wan, J.; Wu, Y.; Li, K.; Jin, C.; Chen, W.; Wang, S.; Wang, W.; Zhang, H.; et al. Control of Leaf Senescence by an MeOH-Jasmonates Cascade That Is Epigenetically Regulated by OsSRT1 in Rice. Mol. Plant 2016, 9, 1366–1378. [Google Scholar] [CrossRef] [Green Version]
- Ma, J.-Q.; Tuersun, H.; Jiao, S.-J.; Zheng, J.-H.; Xiao, J.-B.; Hasim, A. Functional Role of NRF2 in Cervical Carcinogenesis. PLoS ONE 2015, 10, e0133876. [Google Scholar] [CrossRef]
- Itoh, K.; Wakabayashi, N.; Katoh, Y.; Ishii, T.; O’Connor, T.; Yamamoto, M. Keap1 Regulates Both Cytoplasmic-Nuclear Shuttling and Degradation of Nrf2 in Response to Electrophiles. Genes Cells 2003, 8, 379–391. [Google Scholar] [CrossRef]
- Cuadrado, A.; Rojo, A.I.; Wells, G.; Hayes, J.D.; Cousin, S.P.; Rumsey, W.L.; Attucks, O.C.; Franklin, S.; Levonen, A.-L.; Kensler, T.W.; et al. Therapeutic Targeting of the NRF2 and KEAP1 Partnership in Chronic Diseases. Nat. Rev. Drug Discov. 2019, 18, 295–317. [Google Scholar] [CrossRef] [Green Version]
- Gęgotek, A.; Skrzydlewska, E. The Role of Transcription Factor Nrf2 in Skin Cells Metabolism. Arch. Dermatol. Res. 2015, 307, 385–396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parola, M.; Robino, G.; Marra, F.; Pinzani, M.; Bellomo, G.; Leonarduzzi, G.; Chiarugi, P.; Camandola, S.; Poli, G.; Waeg, G.; et al. HNE Interacts Directly with JNK Isoforms in Human Hepatic Stellate Cells. J. Clin. Investig. 1998, 102, 1942–1950. [Google Scholar] [CrossRef] [PubMed]
- Wyler, E.; Franke, V.; Menegatti, J.; Kocks, C.; Boltengagen, A.; Praktiknjo, S.; Walch-Rückheim, B.; Bosse, J.; Rajewsky, N.; Grässer, F.; et al. Single-Cell RNA-Sequencing of Herpes Simplex Virus 1-Infected Cells Connects NRF2 Activation to an Antiviral Program. Nat. Commun. 2019, 10, 4878. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, H.; Falgout, B.; Takeda, K.; Yamada, K.M.; Dhawan, S. Nrf2-Dependent Induction of Innate Host Defense via Heme Oxygenase-1 Inhibits Zika Virus Replication. Virology 2017, 503, 1–5. [Google Scholar] [CrossRef]
- Ingram, S.L. Regulation of Oxidoreductase Enzymes during Inflammation. Available online: https://research.brighton.ac.uk/en/studentTheses/regulation-of-oxidoreductase-enzymes-during-inflammation (accessed on 26 February 2022).
- Lee, C. Therapeutic Modulation of Virus-Induced Oxidative Stress via the Nrf2-Dependent Antioxidative Pathway. Oxid. Med. Cell. Longev. 2018, 2018, 6208067. [Google Scholar] [CrossRef]
- Liu, T.; Zhang, L.; Joo, D.; Sun, S.-C. NF-ΚB Signaling in Inflammation. Signal Transduct. Target. Ther. 2017, 2, 17023. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Z.; Chen, X.; Sun, J.; Li, Q.; Lian, X.; Li, S.; Wang, Y.; Tian, L. Inhibition of Nuclear Thioredoxin Aggregation Attenuates PM2.5-Induced NF-ΚB Activation and pro-Inflammatory Responses. Free Radic. Biol. Med. 2019, 130, 206–214. [Google Scholar] [CrossRef]
- Pastore, A.; Piemonte, F. S-Glutathionylation Signaling in Cell Biology: Progress and Prospects. Eur. J. Pharm. Sci. 2012, 46, 279–292. [Google Scholar] [CrossRef]
- Kerstholt, M.; Vrijmoeth, H.; Lachmandas, E.; Oosting, M.; Lupse, M.; Flonta, M.; Dinarello, C.A.; Netea, M.G.; Joosten, L.A.B. Role of Glutathione Metabolism in Host Defense against Borrelia Burgdorferi Infection. Proc. Natl. Acad. Sci. USA 2018, 115, E2320–E2328. [Google Scholar] [CrossRef] [Green Version]
- Ghezzi, P. Role of Glutathione in Immunity and Inflammation in the Lung. Int. J. Gen. Med. 2011, 4, 105–113. [Google Scholar] [CrossRef] [Green Version]
- Ji, C.; Kozak, K.R.; Marnett, L.J. IκB Kinase, a Molecular Target for Inhibition by 4-Hydroxy-2-Nonenal. J. Biol. Chem. 2001, 276, 18223–18228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, Y.S.; Park, Z.Y.; Kim, S.Y.; Jeong, E.; Lee, J.Y. Alteration of Toll-like Receptor 4 Activation by 4-Hydroxy-2-Nonenal Mediated by the Suppression of Receptor Homodimerization. Chem.-Biol. Interact. 2009, 182, 59–66. [Google Scholar] [CrossRef] [PubMed]
- EUR-Lex-32012D0340-EN-EUR-Lex. Available online: https://eur-lex.europa.eu/eli/dec_impl/2012/340/oj (accessed on 7 February 2022).
- Stanek, G.; Fingerle, V.; Hunfeld, K.-P.; Jaulhac, B.; Kaiser, R.; Krause, A.; Kristoferitsch, W.; O’Connell, S.; Ornstein, K.; Strle, F.; et al. Lyme Borreliosis: Clinical Case Definitions for Diagnosis and Management in Europe. Clin. Microbiol. Infect. 2011, 17, 69–79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mygland, A.; Ljøstad, U.; Fingerle, V.; Rupprecht, T.; Schmutzhard, E.; Steiner, I. European Federation of Neurological Societies EFNS Guidelines on the Diagnosis and Management of European Lyme Neuroborreliosis. Eur. J. Neurol. 2010, 17, 8-e4. [Google Scholar] [CrossRef]
- Prajda, N.; Weber, G. Malignant Transformation-Linked Imbalance: Decreased Xanthine Oxidase Activity in Hepatomas. FEBS Lett. 1975, 59, 245–249. [Google Scholar] [CrossRef] [Green Version]
- Kuzkaya, N.; Weissmann, N.; Harrison, D.G.; Dikalov, S. Interactions of Peroxynitrite, Tetrahydrobiopterin, Ascorbic Acid, and Thiols: Implications for Uncoupling Endothelial Nitric-Oxide Synthase. J. Biol. Chem. 2003, 278, 22546–22554. [Google Scholar] [CrossRef] [Green Version]
- Paglia, D.E.; Valentine, W.N. Studies on the Quantitative and Qualitative Characterization of Erythrocyte Glutathione Peroxidase. J. Lab. Clin. Med. 1967, 70, 158–169. [Google Scholar]
- Mize, C.E.; Langdon, R.G. Hepatic Glutathione Reductase. I. Purification and General Kinetic Properties. J. Biol. Chem. 1962, 237, 1589–1595. [Google Scholar] [CrossRef]
- Misra, H.P.; Fridovich, I. The Role of Superoxide Anion in the Autoxidation of Epinephrine and a Simple Assay for Superoxide Dismutase. J. Biol. Chem. 1972, 247, 3170–3175. [Google Scholar] [CrossRef]
- Sykes, J.A.; McCormack, F.X.; O’Brien, T.J. A Preliminary Study of the Superoxide Dismutase Content of Some Human Tumors. Cancer Res. 1978, 38, 2759–2762. [Google Scholar]
- Holmgren, A.; Björnstedt, M. Thioredoxin and Thioredoxin Reductase. Methods Enzymol. 1995, 252, 199–208. [Google Scholar] [CrossRef] [PubMed]
- Lovell, M.A.; Xie, C.; Gabbita, S.P.; Markesbery, W.R. Decreased Thioredoxin and Increased Thioredoxin Reductase Levels in Alzheimer’s Disease Brain. Free Radic. Biol. Med. 2000, 28, 418–427. [Google Scholar] [CrossRef]
- Maeso, N.; García-Martínez, D.; Rupérez, F.J.; Cifuentes, A.; Barbas, C. Capillary Electrophoresis of Glutathione to Monitor Oxidative Stress and Response to Antioxidant Treatments in an Animal Model. J. Chromatogr. B 2005, 822, 61–69. [Google Scholar] [CrossRef] [PubMed]
- De Leenheer, A.P.; De Bevere, V.O.R.C.; De Ruyter, M.G.M.; Claeys, A.E. Simultaneous Determination of Retinol and α-Tocopherol in Human Serum by High-Performance Liquid Chromatography. J. Chromatogr. B Biomed. Sci. Appl. 1979, 162, 408–413. [Google Scholar] [CrossRef]
- Ivanović, D.; Popović, A.; Radulović, D.; Medenica, M. Reversed-Phase Ion-Pair HPLC Determination of Some Water-Soluble Vitamins in Pharmaceuticals. J. Pharm. Biomed. Anal. 1999, 18, 999–1004. [Google Scholar] [CrossRef]
- Christie, W.W. Advances in Lipid Methodology II; Oily Press Lipid Library; Oily Press: Bridgewater, NJ, USA, 1993. [Google Scholar]
- Tsikas, D.; Rothmann, S.; Schneider, J.Y.; Gutzki, F.-M.; Beckmann, B.; Frölich, J.C. Simultaneous GC-MS/MS Measurement of Malondialdehyde and 4-Hydroxy-2-Nonenal in Human Plasma: Effects of Long-Term L-Arginine Administration. Anal. Biochem. 2017, 524, 31–44. [Google Scholar] [CrossRef]
- Coolen, S.A.J.; van Buuren, B.; Duchateau, G.; Upritchard, J.; Verhagen, H. Kinetics of Biomarkers: Biological and Technical Validity of Isoprostanes in Plasma. Amino Acids 2005, 29, 429–436. [Google Scholar] [CrossRef]
- Hnasko, R.; Lin, A.; McGarvey, J.A.; Stanker, L.H. A Rapid Method to Improve Protein Detection by Indirect ELISA. Biochem. Biophys. Res. Commun. 2011, 410, 726–731. [Google Scholar] [CrossRef]
- Hawkins, C.L.; Morgan, P.E.; Davies, M.J. Quantification of Protein Modification by Oxidants. Free Radic. Biol. Med. 2009, 46, 965–988. [Google Scholar] [CrossRef]
- Levine, R.L.; Garland, D.; Oliver, C.N.; Amici, A.; Climent, I.; Lenz, A.G.; Ahn, B.W.; Shaltiel, S.; Stadtman, E.R. Determination of Carbonyl Content in Oxidatively Modified Proteins. Methods Enzymol. 1990, 186, 464–478. [Google Scholar] [CrossRef]
- Weber, D.; Milkovic, L.; Bennett, S.J.; Griffiths, H.R.; Zarkovic, N.; Grune, T. Measurement of HNE-Protein Adducts in Human Plasma and Serum by ELISA-Comparison of Two Primary Antibodies. Redox Biol. 2013, 1, 226–233. [Google Scholar] [CrossRef] [PubMed] [Green Version]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| CTR | TBE | TBE+LD/HGA | |
|---|---|---|---|
| Age (Years) | 41 (28–55) | 40 (23–58) | 42 (22–63) |
| Sex | 5/20 female (25%) 15/20 male (75%) | 14/40 female (35%) 26/40 male (65%) | 4/6 female (67%) 2/6 male (33%) |
| Confirmed tick bite | 0/20 (0%) | 21/40 (52%) | 5/6 (83%) |
| Time since tick bite (days) | - | 26.8 ± 16.6 | 17.58 ± 4.95 |
| Duration of hospitalization (days) | - | 12.85 ± 2.6 | 12.34 ± 1.97 |
| Duration of symptoms (days) | - | 7.5 ± 6.9 | 4.26 ± 2.63 |
| Clinical form | |||
| Meningitis | 0/20(0%) | 25/40 (62.5%) | 5/6 (83%) |
| Meningoencephalitis | 0/20 (0%) | 15/40 (37.5%) | 1/6 (17%) |
| Meningoencephalomyelitis | 0/20 (0%) | 0/40 (0%) | 0/6 (0%) |
| Normal Range | TBE | TBEV+Bb/Ap | |||
|---|---|---|---|---|---|
| At Admission | After Recovery | At Admission | After Recovery | ||
| Complete blood count | |||||
| WBC [103/μL] | 4.00–10.00 | 10.16 ± 2.37 | 6.11 ± 1.48 | 7.27 ± 1.36 | 5.37 ± 1.46 |
| Neutrophils [%] | 40.0–72.0 | 73.77 ± 9.52 | 51.18 ± 7.93 | 60.33 ± 7.07 | 42.58 ± 11.58 |
| Lymphocytes [%] | 18.00–48.00 | 16.2 ± 7.76 | 34.83 ± 6.35 | 27.37 ± 6.11 | 42.72 ± 11.24 |
| Monocytes [%] | 2.50–10.00 | 8.99 ± 2.53 | 9.79 ± 2.15 | 10.15 ± 2.61 | 9.56 ± 3.18 |
| RBC [106/μL] | 4.00–5.50 | 4.40 ± 0.43 | 4.50 ± 0.32 | 4.28 ± 0.56 | 4.14 ± 0.62 |
| HGB [g/dL] | 12.00–16.00 | 13.32 ± 1.28 | 13.68 ± 1.09 | 12.67 ± 1.26 | 12.5 ± 1.39 |
| PLT [103/μL] | 130–350 | 251.44 ± 87.43 | 270.59 ± 134.4 | 262.67 ± 66.98 | 218.4 ± 19.42 |
| CRP [mg/L] | 0.00–5.00 | 11.52 ± 15.61 | 0.93 ± 0.54 | 2.35 ± 2.08 | 0.81 ± 0.26 |
| Glucose [mg/dL] | 70–110 | 96.77 ± 10.27 | 89.62 ± 12.1 | 92.67 ± 8.5 | 91.75 ± 5.97 |
| Creatinine [mg/dL] | 0.50–0.90 | 0.88 ± 0.16 | 0.82 ± 0.12 | 0.79 ± 0.08 | 0.76 ± 0.12 |
| ALT [U/I] | 0–31 | 21.10 ± 19.17 | 24.17 ± 17.20 | 17.75 ± 11.27 | 12 ± 5.24 |
| AST [U/I] | 0–32 | 15.59 ± 5.26 | 22.42 ± 9.23 | 17 ± 3.67 | 16.33 ± 3.21 |
| CSF analysis | |||||
| Cytosis [cells/µL] | 0–5 | 169.7 ± 115.64 | 26.90 ± 25.59 | 99 ± 120.57 | 14.8 ± 12.15 |
| Protein [mg/dL] | 15–45 | 75.95 ± 20.56 | 54.46 ± 26.32 | 69 ± 58.49 | 48.2 ± 27.74 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dobrzyńska, M.; Moniuszko-Malinowska, A.; Jarocka-Karpowicz, I.; Czupryna, P.; Groth, M.; Skrzydlewska, E. Metabolic Response to Tick-Borne Encephalitis Virus Infection and Bacterial Co-Infections. Pathogens 2022, 11, 384. https://doi.org/10.3390/pathogens11040384
Dobrzyńska M, Moniuszko-Malinowska A, Jarocka-Karpowicz I, Czupryna P, Groth M, Skrzydlewska E. Metabolic Response to Tick-Borne Encephalitis Virus Infection and Bacterial Co-Infections. Pathogens. 2022; 11(4):384. https://doi.org/10.3390/pathogens11040384
Chicago/Turabian StyleDobrzyńska, Marta, Anna Moniuszko-Malinowska, Iwona Jarocka-Karpowicz, Piotr Czupryna, Monika Groth, and Elżbieta Skrzydlewska. 2022. "Metabolic Response to Tick-Borne Encephalitis Virus Infection and Bacterial Co-Infections" Pathogens 11, no. 4: 384. https://doi.org/10.3390/pathogens11040384