
Protein Concentrations in Stored Pooled Platelet Concentrates Treated with Pathogen Inactivation by Amotosalen Plus Ultraviolet a Illumination

 , , , and
, , , and 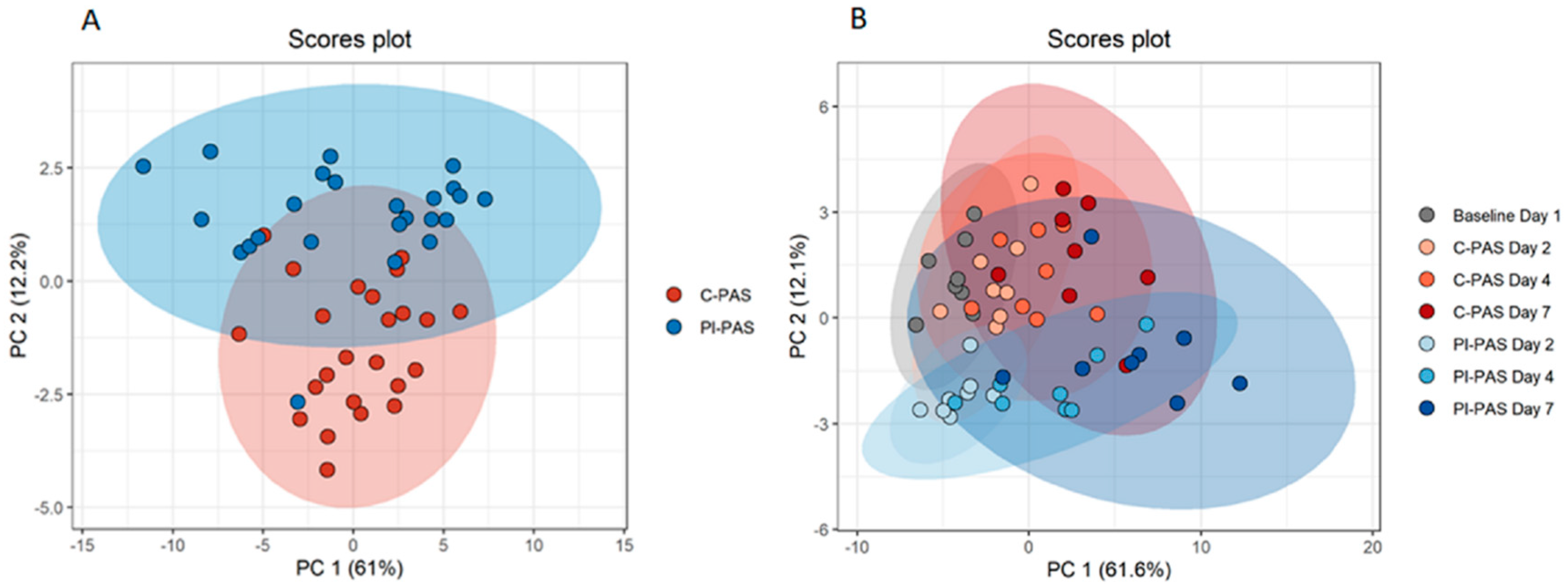{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
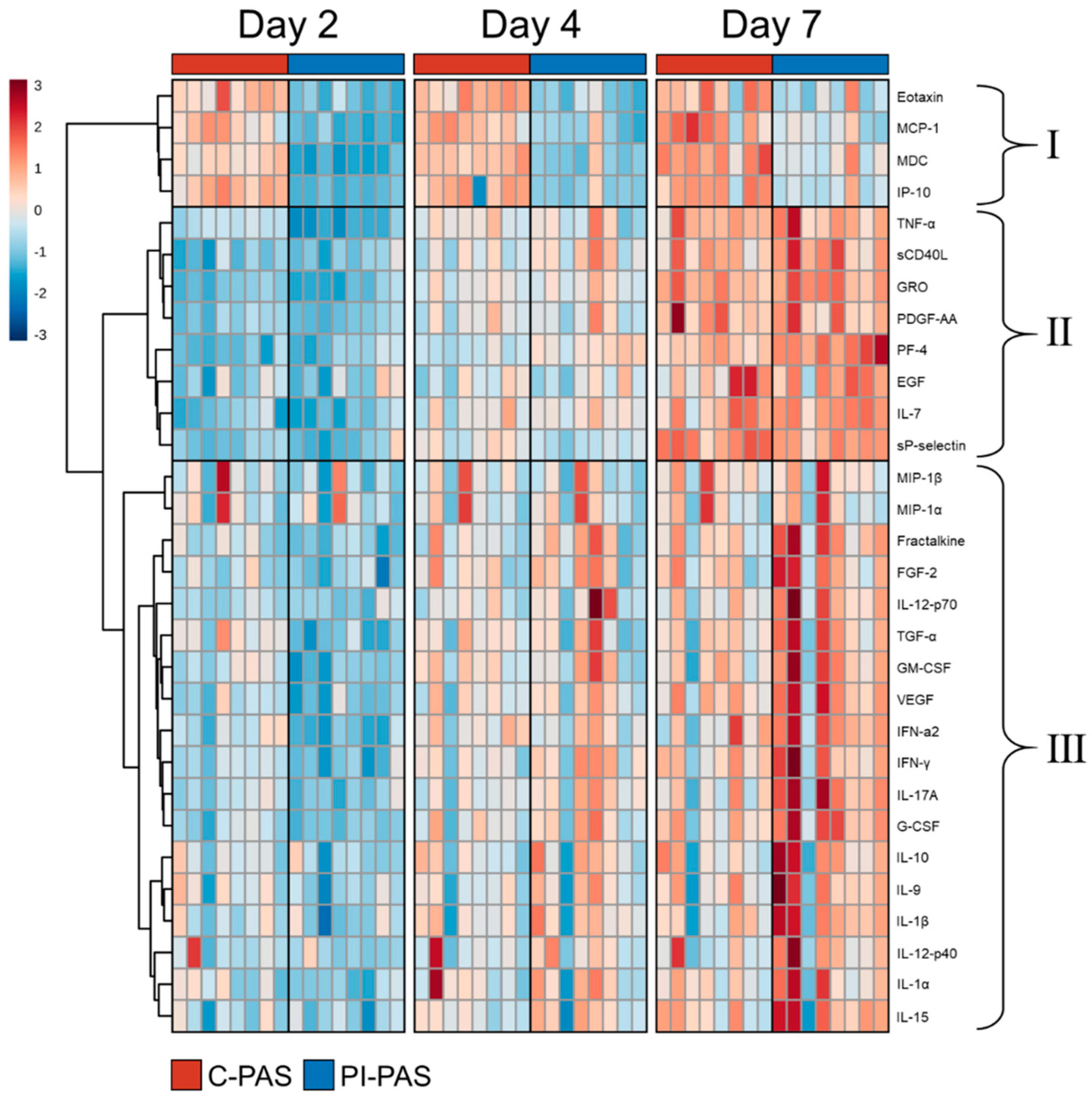
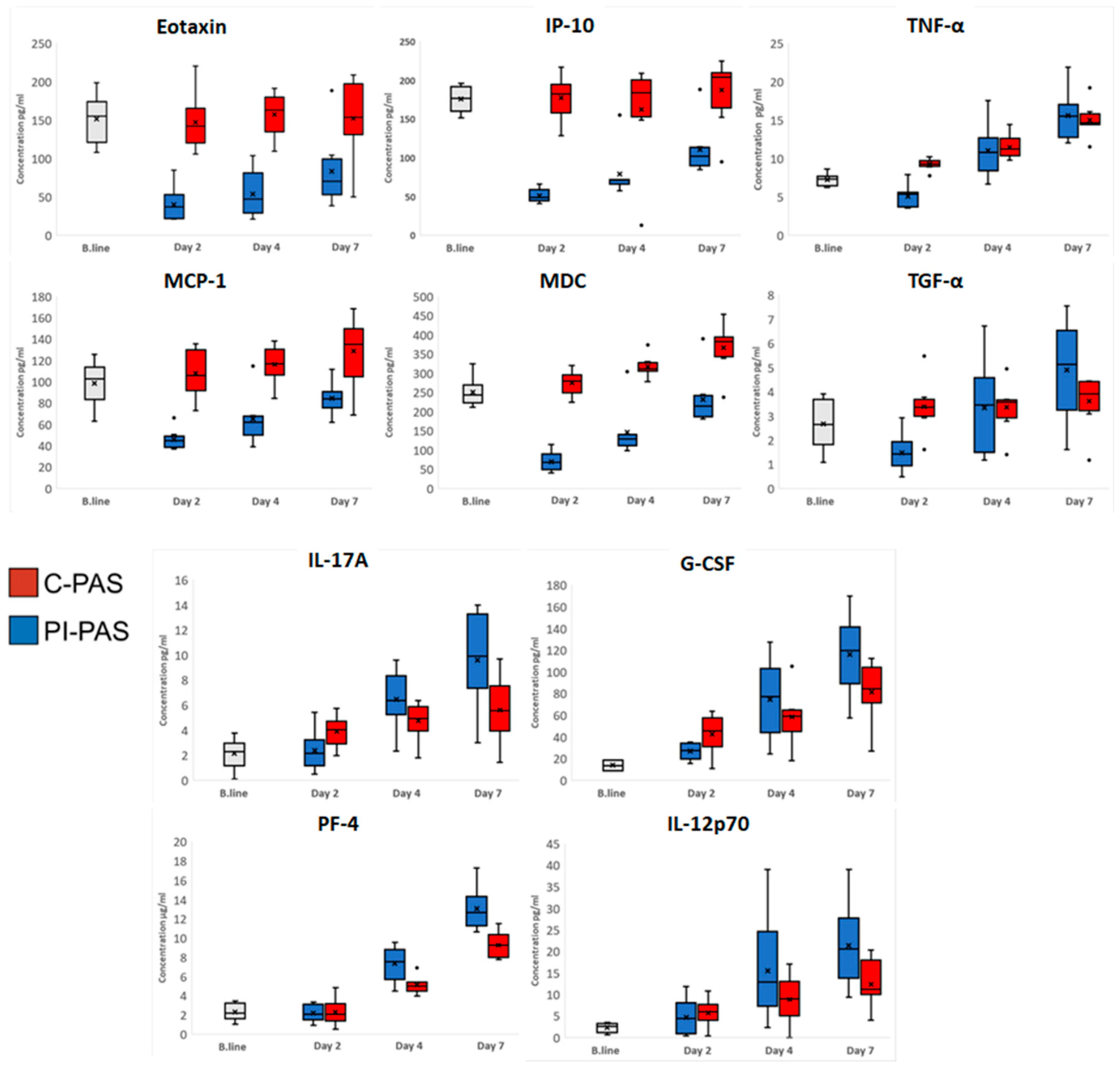
2. Results
Protein Accumulation in PCs during Storage
3. Discussion
4. Materials and Methods
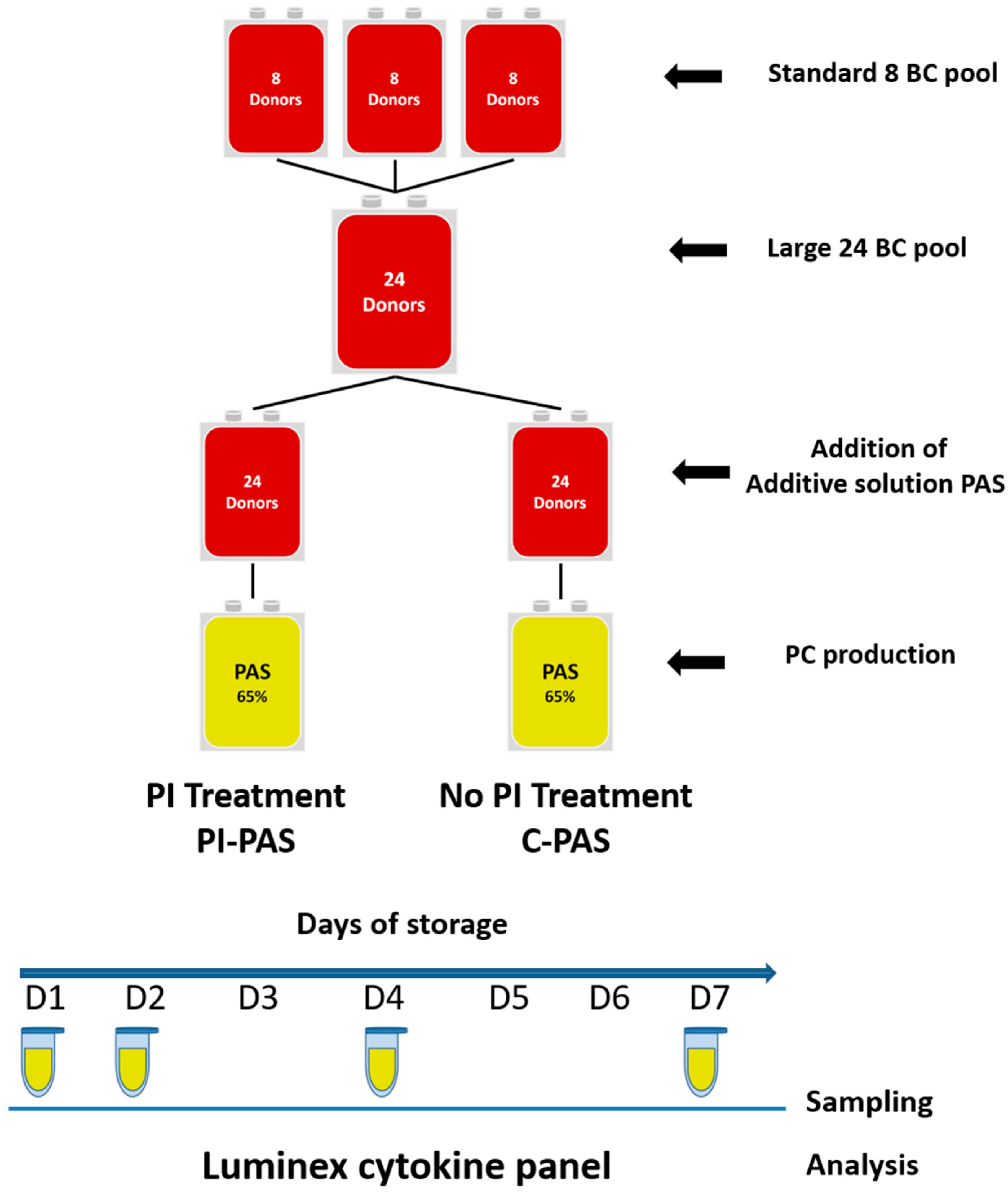
4.1. Collection, Processing, Storage, and Sampling
4.2. ELISA
4.3. Human Cytokine/Chemokine Magnetic Bead Panel Assay
4.4. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Li, Z.; Smith, S.S. Interactions between platelets, leukocytes, and the endothelium. In Platelets, 4th ed.; Michelson, A.D., Ed.; Academic Press: Cambridge, MA, USA, 2019; pp. 295–310. [Google Scholar]
- Garraud, O.; Cognasse, F. Are platelets cells? And if yes, are they immune cells? Front. Immunol. 2015, 6, 70. [Google Scholar] [CrossRef] [Green Version]
- Sut, C.; Tariket, S.; Aubron, C.; Aloui, C.; Hamzeh-Cognasse, H.; Berthelot, P.; Laradi, S.; Greinacher, A.; Garraud, O.; Cognasse, F. The non-hemostatic aspects of transfused platelets. Front. Med. 2018, 5, 42. [Google Scholar] [CrossRef] [PubMed]
- Brass, L.F.; Tomaiuolo, M.; Welsh, J.; Poventud-Fuentes, I.; Zhu, L.; Diamond, S.L.; Stalker, T.J. Hemostatic thrombus formation in flowing blood. In Platelets; Academic Press: Cambridge, MA, USA, 2019. [Google Scholar] [CrossRef]
- Smith, S.A.; Morrissey, J.H. Interactions between platelets and the coagulation system. In Platelets; Academic Press: Cambridge, MA, USA, 2019. [Google Scholar] [CrossRef]
- Estcourt, L.J. Why has demand for platelet components increased? A review. Transfus. Med. 2014, 24, 260–268. [Google Scholar] [CrossRef]
- Charlton, A.; Wallis, J.; Robertson, J.; Watson, D.; Iqbal, A.; Tinegate, H. Where did platelets go in 2012? A survey of platelet transfusion practice in the North of England. Transfus. Med. 2014, 24, 213–218. [Google Scholar] [CrossRef] [PubMed]
- Newland, A.; Bentley, R.; Jakubowska, A.; Liebman, H.; Lorens, J.; Peck-Radosavljevic, M.; Taieb, V.; Takami, A.; Tateishi, R.; Younossi, Z.M. A systematic literature review on the use of platelet transfusions in patients with thrombocytopenia. Hematology 2019, 24, 679–719. [Google Scholar] [CrossRef] [Green Version]
- Roh, J.; Choi, S.J.; Kim, S.; Min, H.; Kim, H.O. Blood supply and demand in Korea: What is in store for the future? Yonsei Med. J. 2020, 61, 400. [Google Scholar] [CrossRef] [PubMed]
- Shah, A.; Stanworth, S.J.; McKechnie, S. Evidence and triggers for the transfusion of blood and blood products. Anaesthesia 2015, 70, 10-e3. [Google Scholar] [CrossRef]
- Mookerjee, S.; Foster, H.R.; Waller, A.K.; Ghevaert, C.J. In vitro-derived platelets: The challenges we will have to face to assess quality and safety. Platelets 2020, 31, 724–730. [Google Scholar] [CrossRef]
- Högman, C.F. Storage of blood components. Curr. Opin. Hematol. 1999, 6, 427. [Google Scholar] [CrossRef] [PubMed]
- Vasconcelos, E.; Figueiredo, A.C.; Seghatchian, J. Quality of platelet concentrates derived by platelet rich plasma, buffy coat and Apheresis. Transfus. Apher. Sci. 2003, 29, 13–16. [Google Scholar] [CrossRef]
- Schubert, P.; Devine, D.V. Towards targeting platelet storage lesion-related signaling pathways. Blood Transfus. 2010, 8, s69. [Google Scholar] [CrossRef] [PubMed]
- Schrezenmeier, H.; Walther-Wenke, G.; Müller, T.H.; Weinauer, F.; Younis, A.; Holland-Letz, T.; Geis, G.; Asmus, J.; Bauerfeind, U.; Burkhart, J.; et al. Bacterial contamination of platelet concentrates: Results of a prospective multicenter study comparing pooled whole blood-derived platelets and apheresis platelets. Transfusion 2007, 47, 644–652. [Google Scholar] [CrossRef]
- Pietersz, R.N.I.; Reesink, H.W.; Panzer, S.; Oknaian, S.; Kuperman, S.; Gabriel, C.; Rapaille, A.; Lambermont, M.; Deneys, V.; Sondag, D.; et al. Bacterial contamination in platelet concentrates. Vox Sang. 2014, 106, 256–283. [Google Scholar] [CrossRef] [PubMed]
- Kleinman, S.; Reed, W.; Stassinopoulos, A. A patient-oriented risk-benefit analysis of pathogen-inactivated blood components: Application to apheresis platelets in the United States. Transfusion 2013, 53, 1603–1618. [Google Scholar] [CrossRef] [PubMed]
- Spindler-Raffel, E.; Benjamin, R.J.; McDonald, C.P.; Ramirez-Arcos, S.; Aplin, K.; Bekeredjian-Ding, I.; de Korte, D.; Gabriel, C.; Gathof, B.; Hanschmann, K.M.; et al. Enlargement of the WHO international repository for platelet transfusion-relevant bacteria reference strains. Vox Sang. 2017, 112, 713–722. [Google Scholar] [CrossRef]
- Bacterial Risk Control Strategies for Blood Collection Establishments and Transfusion Services to Enhance the Safety and Availability of Platelets for Transfusion|FDA n.d. Available online: https://www.fda.gov/regulatory-information/search-fda-guidance-documents/bacterial-risk-control-strategies-blood-collection-establishments-and-transfusion-services-enhance (accessed on 28 December 2021).
- Hong, H.; Xiao, W.; Lazarus, H.M.; Good, C.E.; Maitta, R.W.; Jacobs, M.R. Detection of septic transfusion reactions to platelet transfusions by active and passive surveillance. Blood 2016, 127, 496–502. [Google Scholar] [CrossRef] [Green Version]
- Estcourt, L.J.; Malouf, R.; Hopewell, S.; Trivella, M.; Doree, C.; Stanworth, S.J.; Murphy, M.F. Pathogen-reduced platelets for the prevention of bleeding. Cochrane Database Syst. Rev. 2017, 2017. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, L.; Cook, D.N.; Wiesehahn, G.P.; Alfonso, R.; Behrman, B.; Cimino, G.D.; Corten, L.; Damonte, P.B.; Dikeman, R.; Dupuis, K.; et al. Photochemical inactivation of viruses and bacteria in platelet concentrates by use of a novel psoralen and long-wavelength ultraviolet light. Transfusion 1997, 37, 423–435. [Google Scholar] [CrossRef] [PubMed]
- Wollowitz, S. Fundamentals of the psoralen-based Helinx™ technology for inactivation of infectious pathogens and leukocytes in platelets and plasma. Semin. Hematol. 2001, 38, 4–11. [Google Scholar] [CrossRef]
- Cid, J. Prevention of transfusion-associated graft-versus-host disease with pathogen-reduced platelets with amotosalen and ultraviolet A light: A review. Vox Sang. 2017, 112, 607–613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mohr, H.; Steil, L.; Gravemann, U.; Thiele, T.; Hammer, E.; Greinacher, A.; Müller, T.H.; Völker, U. A novel approach to pathogen reduction in platelet concentrates using short-wave ultraviolet light. Transfusion 2009, 49, 2612–2624. [Google Scholar] [CrossRef] [PubMed]
- Pelletier, J.P.R.; Transue, S.; Snyder, E.L. Pathogen inactivation techniques. Best Pract. Res. Clin. Haematol. 2006, 19, 205–242. [Google Scholar] [CrossRef]
- Rosskopf, K.; Helmberg, W.; Schlenke, P. Pathogen reduction of double-dose platelet concentrates from pools of eight buffy coats: Product quality, safety, and economic aspects. Transfusion 2020, 60, 2058–2066. [Google Scholar] [CrossRef]
- Sigle, J.P.; Infanti, L.; Studt, J.D.; Martinez, M.; Stern, M.; Gratwohl, A.; Passweg, J.; Tichelli, A.; Buser, A.S. Comparison of transfusion efficacy of amotosalen-based pathogen-reduced platelet components and gamma-irradiated platelet components. Transfusion 2013, 53, 1788–1797. [Google Scholar] [CrossRef] [PubMed]
- Infanti, L.; Holbro, A.; Passweg, J.; Bolliger, D.; Tsakiris, D.A.; Merki, R.; Plattner, A.; Tappe, D.; Irsch, J.; Lin, J.S.; et al. Clinical impact of amotosalen-ultraviolet A pathogen-inactivated platelets stored for up to 7 days. Transfusion 2019, 59, 3350–3361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garban, F.; Guyard, A.; Labussière, H.; Bulabois, C.E.; Marchand, T.; Mounier, C.; Caillot, D.; Bay, J.O.; Coiteux, V.; Schmidt-Tanguy, A.; et al. Comparison of the hemostatic efficacy of pathogen-reduced platelets vs. Untreated platelets in patients with thrombocytopenia and malignant hematologic diseases: A randomized clinical trial. JAMA Oncol. 2018, 4, 468–475. [Google Scholar] [CrossRef] [PubMed]
- Levy, J.H.; Neal, M.D.; Herman, J.H. Bacterial contamination of platelets for transfusion: Strategies for prevention. Crit. Care 2018, 22, 271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kocazeybek, B.; Arabaci, Ü.; Akdur, H.; Sezgiç, M.; Erentürk, S. Prospective evaluation of platelets prepared by single and random methods during 5 days of storage: Aspects related to quality and quantity. Transfus. Apher. Sci. 2002, 26, 29–34. [Google Scholar] [CrossRef]
- Akay, O.M.; Gündüz, E.; Başyiǧit, H.; Gulbas, Z. Platelet function testing during 5-day storage of single and random donor plateletpheresis. Transfus. Apher. Sci. 2007, 36, 285–289. [Google Scholar] [CrossRef] [PubMed]
- Gulliksson, H. Defining the optimal storage conditions for the long-term storage of platelets. Transfus. Med. Rev. 2003, 17, 209–215. [Google Scholar] [CrossRef]
- Sut, C.; Tariket, S.; Aloui, C.; Arthaud, C.A.; Eyraud, M.A.; Fagan, J.; Chavarin, P.; Hamzeh-Cognasse, H.; Laradi, S.; Garraud, O.; et al. Soluble CD40L and CD62P levels differ in single-donor apheresis platelet concentrates and buffy coat–derived pooled platelet concentrates. Transfusion 2019, 59, 16–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Devine, D.V.; Serrano, K. The Platelet Storage Lesion. Clin. Lab. Med. 2010, 30. [Google Scholar] [CrossRef] [PubMed]
- Shrivastava, M. The platelet storage lesion. Transfus. Apher. Sci. 2009, 41, 105–113. [Google Scholar] [CrossRef]
- Singh, S.; Shams Hakimi, C.; Jeppsson, A.; Hesse, C. Platelet storage lesion in interim platelet unit concentrates: A comparison with buffy-coat and apheresis concentrates. Transfus. Apher. Sci. 2017, 56, 870–874. [Google Scholar] [CrossRef] [PubMed]
- Shams Hakimi, C.; Hesse, C.; Wallén, H.; Boulund, F.; Grahn, A.; Jeppsson, A. In vitro assessment of platelet concentrates with multiple electrode aggregometry. Platelets 2015, 26, 132–137. [Google Scholar] [CrossRef] [PubMed]
- Waters, L.; Cameron, M.; Padula, M.P.; Marks, D.C.; Johnson, L. Refrigeration, cryopreservation and pathogen inactivation: An updated perspective on platelet storage conditions. Vox Sang. 2018, 113, 317–328. [Google Scholar] [CrossRef]
- Rijkers, M.; van der Meer, P.F.; Bontekoe, I.J.; Daal, B.B.; de Korte, D.; Leebeek, F.W.G.; Voorberg, J.; Jansen, A.J.G. Evaluation of the role of the GPIb-IX-V receptor complex in development of the platelet storage lesion. Vox Sang. 2016, 111, 247–256. [Google Scholar] [CrossRef] [PubMed]
- Cognasse, F.; Boussoulade, F.; Chavarin, P.; Acquart, S.; Fabrigli, P.; Lamy, B.; Garraud, O. Release of potential immunomodulatory factors during platelet storage. Transfusion 2006, 46, 1184–1189. [Google Scholar] [CrossRef]
- Bubel, S.; Wilhelm, D.; Entelmann, M.; Kirchner, H.; Klüter, H. Chemokines in stored platelet concentrates. Transfusion 1996, 36, 445–449. [Google Scholar] [CrossRef] [PubMed]
- Joos, M.; Wouters, E.; De Bock, R.; Peetermans, M.E. Increased tumor necrosis factor alpha (TNF alpha), interleukin 1, and interleukin 6 (IL-6) levels in the plasma of stored platelet concentrates: Relationship between TNF alpha and IL-6 levels and febrile transfusion reactions. Transfusion 1993, 33, 195–199. [Google Scholar] [CrossRef]
- Saris, A.; Peyron, I.; van der Meer, P.F.; Stuge, T.B.; Zwaginga, J.J.; van Ham, S.M.; ten Brinke, A. Storage-induced platelet apoptosis is a potential risk factor for alloimmunization upon platelet transfusion. Front. Immunol. 2018, 9, 1251. [Google Scholar] [CrossRef] [Green Version]
- Seghatchian, J.; Krailadsiri, P. Platelet storage lesion and apoptosis: Are they related? Transfus. Apher. Sci. 2001, 24, 103–105. [Google Scholar] [CrossRef]
- Leytin, V. Apoptosis in the anucleate platelet. Blood Rev. 2012, 26, 51–63. [Google Scholar] [CrossRef] [PubMed]
- Smethurst, P.A. Aging of platelets stored for transfusion. Platelets 2016, 27, 526–534. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Xie, R.; Fan, Z.; Yang, J.; Liang, W.; Wu, Q.; Wu, M.X.; Wang, Z.; Lu, Y. The contribution of oxidative stress to platelet senescence during storage. Transfusion 2019, 59, 2389–2402. [Google Scholar] [CrossRef]
- Pagel, O.; Walter, E.; Jurk, K.; Zahedi, R.P. Taking the stock of granule cargo: Platelet releasate proteomics. Platelets 2017, 28, 119–128. [Google Scholar] [CrossRef] [PubMed]
- Yeaman, M.R. Platelets: At the nexus of antimicrobial defence. Nat. Rev. Microbiol. 2014, 12, 426–437. [Google Scholar] [CrossRef] [PubMed]
- Sut, C.; Hamzeh-Cognasse, H.; Arthaud, C.A.; Eyraud, M.A.; Chettab, K.; Dumontet, C.; Laradi, S.; Burnouf, T.; Garraud, O.; Cognasse, F. Platelet concentrate supernatants alter endothelial cell mRNA and protein expression patterns as a function of storage length. Transfusion 2018, 58, 2635–2644. [Google Scholar] [CrossRef] [PubMed]
- Kostelijk, E.H.; Fijnheer, R.; Nieuwenhuis, H.K.; Gouwerok, C.W.N.; De Korte, D. Soluble P-selectin as parameter for platelet activation during storage. Thromb. Haemost. 1996, 76, 1086–1089. [Google Scholar] [CrossRef]
- Cognasse, F.; Sut, C.; Fromont, E.; Laradi, S.; Hamzeh-Cognasse, H.; Garraud, O. Platelet soluble CD40-ligand level is associated with transfusion adverse reactions in a mixed threshold-and-hit model. Blood 2017, 130, 1380–1383. [Google Scholar] [CrossRef] [Green Version]
- Khan, S.Y.; Kelher, M.R.; Heal, J.M.; Blumberg, N.; Boshkov, L.K.; Phipps, R.; Gettings, K.F.; McLaughlin, N.J.; Silliman, C.C. Soluble CD40 ligand accumulates in stored blood components, primes neutrophils through CD40, and is a potential cofactor in the development of transfusion-related acute lung injury. Blood 2006, 108, 2455–2462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Phipps, R.P.; Kaufman, J.; Blumberg, N. Platelet derived CD154 (CD40 ligand) and febrile responses to transfusion. Lancet 2001, 357, 2023–2024. [Google Scholar] [CrossRef]
- Manne, B.K.; Xiang, S.C.; Rondina, M.T. Platelet secretion in inflammatory and infectious diseases. Platelets 2017, 28, 155–164. [Google Scholar] [CrossRef] [PubMed]
- Klüter, H.; Bubel, S.; Kirchner, H.; Wilhelm, D. Febrile and allergic transfusion reactions after the transfusion of white cell-poor platelet preparations. Transfusion 1999, 39, 1179–1184. [Google Scholar] [CrossRef] [PubMed]
- Hamzeh-Cognasse, H.; Damien, P.; Nguyen, K.A.; Arthaud, C.A.; Eyraud, M.A.; Chavarin, P.; Absi, L.; Osselaer, J.C.; Pozzetto, B.; Cognasse, F.; et al. Immune-reactive soluble OX40 ligand, soluble CD40 ligand, and interleukin-27 are simultaneously oversecreted in platelet components associated with acute transfusion reactions. Transfusion 2014, 54, 613–625. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, K.A.; Hamzeh-Cognasse, H.; Sebban, M.; Fromont, E.; Chavarin, P.; Absi, L.; Pozzetto, B.; Cognasse, F.; Garraud, O. A computerized prediction model of hazardous inflammatory platelet transfusion outcomes. PLoS ONE 2014, 9, e97082. [Google Scholar] [CrossRef]
- Howell, W.M.; Ali, S.; Rose-Zerilli, M.J.; Ye, S. VEGF polymorphisms and severity of atherosclerosis. J. Med. Genet. 2005, 42, 485–490. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kanter, J.; Khan, S.Y.; Kelher, M.; Gore, L.; Silliman, C.C. Oncogenic and angiogenic growth factors accumulate during routine storage of apheresis platelet concentrates. Clin. Cancer Res. 2008, 14, 3942–3947. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nevzorova, T.A.; Mordakhanova, E.R.; Daminova, A.G.; Ponomareva, A.A.; Andrianova, I.A.; Le Minh, G.; Rauova, L.; Litvinov, R.I.; Weisel, J.W. Platelet factor 4-containing immune complexes induce platelet activation followed by calpain-dependent platelet death. Cell Death Discov. 2019, 5, 106. [Google Scholar] [CrossRef] [Green Version]
- Von Hundelshausen, P.; Koenen, R.R.; Sack, M.; Mause, S.F.; Adriaens, W.; Proudfoot, A.E.I.; Hackeng, T.M.; Weber, C. Heterophilic interactions of platelet factor 4 and RANTES promote monocyte arrest on endothelium. Blood 2005, 105, 924–930. [Google Scholar] [CrossRef] [Green Version]
- Cognasse, F.; Laradi, S.; Berthelot, P.; Bourlet, T.; Marotte, H.; Mismetti, P.; Garraud, O.; Hamzeh-Cognasse, H. Platelet inflammatory response to stress. Front. Immunol. 2019, 10, 1478. [Google Scholar] [CrossRef]
- Murphy, S.; Sayar, S.N.; Gardner, F.H. Storage of platelet concentrates at 22 degrees C. Blood 1970, 35, 549. [Google Scholar] [CrossRef] [Green Version]
- Snyder, E.L. The cellular and molecular basis of the platelet storage lesion: A symposium summary. Transfusion 1992, 32, 386–390. [Google Scholar] [CrossRef]
- Schöneich, C. Photo-Degradation of Therapeutic Proteins: Mechanistic Aspects. Pharm. Res. 2020, 37, 45. [Google Scholar] [CrossRef] [PubMed]
- Jóhannsson, F.; Árnason, N.; Landrö, R.; Guðmundsson, S.; Sigurjonsson, Ó.E.; Rolfsson, Ó. Metabolomics study of platelet concentrates photochemically treated with amotosalen and UVA light for pathogen inactivation. Transfusion 2020, 60, 367–377. [Google Scholar] [CrossRef] [PubMed]
- Shukla, R.; Patel, T.; Gupte, S. Release of cytokines in stored whole blood and red cell concentrate: Effect of leukoreduction. Asian J. Transfus. Sci. 2015, 9, 145. [Google Scholar] [CrossRef] [PubMed]
- Flaumenhaft, R.; Sharda, A. Platelet Secretion. In Platelets; Academic Press: Cambridge, MA, USA, 2019; pp. 349–370. [Google Scholar] [CrossRef]
- Boyanova, D.; Nilla, S.; Birschmann, I.; Dandekar, T.; Dittrich, M. PlateletWeb: A systems biologic analysis of signaling networks in human platelets. Blood 2012, 119, e22–e34. [Google Scholar] [CrossRef] [Green Version]
- Apelseth, T.O.; Bruserud, Ø.; Wentzel-Larsen, T.; Bakken, A.M.; Bjørsvik, S.; Hervig, T. In vitro evaluation of metabolic changes and residual platelet responsiveness in photochemical treated and gamma-irradiated single-donor platelet concentrates during long-term storage. Transfusion 2007, 47, 653–665. [Google Scholar] [CrossRef] [PubMed]
- Johnson, L.; Loh, Y.S.; Kwok, M.; Marks, D.C. In vitro assessment of buffy-coat derived platelet components suspended in SSP+ treated with the INTERCEPT Blood system. Transfus. Med. 2013, 23, 121–129. [Google Scholar] [CrossRef]
- Spiel, A.O.; Bartko, J.; Schwameis, M.; Firbas, C.; Siller-Matula, J.; Schuetz, M.; Weigl, M.; Jilma, B. Increased platelet aggregation and in vivo platelet activation after granulocyte colony-stimulating factor administration A randomised controlled trial. Thromb. Haemost. 2011, 105, 655–662. [Google Scholar] [CrossRef] [PubMed]
- Freson, K. The platelet proteome. In Platelets; Academic Press: Cambridge, MA, USA, 2019; pp. 155–167. [Google Scholar] [CrossRef]
- Burkhart, J.M.; Gambaryan, S.; Watson, S.P.; Jurk, K.; Walter, U.; Sickmann, A.; Heemskerk, J.W.M.; Zahedi, R.P. What can proteomics tell us about platelets? Circ. Res. 2014, 114, 1204–1219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parsons, M.E.M.; Szklanna, P.B.; Guerrero, J.A.; Wynne, K.; Dervin, F.; O’Connell, K.; Allen, S.; Egan, K.; Bennett, C.; McGuigan, C.; et al. Platelet Releasate Proteome Profiling Reveals a Core Set of Proteins with Low Variance between Healthy Adults. Proteomics 2018, 18, 1800219. [Google Scholar] [CrossRef]
- Burkhart, J.M.; Vaudel, M.; Gambaryan, S.; Radau, S.; Walter, U.; Martens, L.; Geiger, J.; Sickmann, A.; Zahedi, R.P. The first comprehensive and quantitative analysis of human platelet protein composition allows the comparative analysis of structural and functional pathways. Blood 2012, 120, e73–e82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arnason, N.A.; Johannson, F.; Landrö, R.; Hardarsson, B.; Irsch, J.; Gudmundsson, S.; Rolfsson, O.; Sigurjonsson, O.E. Pathogen inactivation with amotosalen plus UVA illumination minimally impacts microRNA expression in platelets during storage under standard blood banking conditions. Transfusion 2019, 59, 3727–3735. [Google Scholar] [CrossRef]
- Kondkar, A.A.; Bray, M.S.; Leal, S.M.; Nagalla, S.; Liu, D.J.; Jin, Y.; Dong, J.F.; Ren, Q.; Whiteheart, S.W.; Shaw, C.; et al. VAMP8/endobrevin is overexpressed in hyperreactive human platelets: Suggested role for platelet microRNA. J. Thromb. Haemost. 2010, 8, 369–378. [Google Scholar] [CrossRef] [Green Version]
- Joshi, S.; Banerjee, M.; Zhang, J.; Kesaraju, A.; Pokrovskaya, I.D.; Storrie, B.; Whiteheart, S.W. Alterations in platelet secretion differentially affect thrombosis and hemostasis. Blood Adv. 2018, 2, 2187–2198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Aelst, B.; Feys, H.B.; Devloo, R.; Vanhoorelbeke, K.; Vandekerckhove, P.; Compernolle, V. Riboflavin and amotosalen photochemical treatments of platelet concentrates reduce thrombus formation kinetics in vitro. Vox Sang. 2015, 108, 328–339. [Google Scholar] [CrossRef] [PubMed]
- Feys, H.B.; Van Aelst, B.; Compernolle, V. Biomolecular Consequences of Platelet Pathogen Inactivation Methods. Transfus. Med. Rev. 2019, 33, 29–34. [Google Scholar] [CrossRef] [PubMed]
- Van Aelst, B.; Devloo, R.; Zachée, P.; T’Kindt, R.; Sandra, K.; Vandekerckhove, P.; Compernolle, V.; Feys, H.B. Psoralen and ultraviolet a light treatment directly affects phosphatidylinositol 3-kinase signal transduction by altering plasma membrane packing. J. Biol. Chem. 2016, 291, 24364–24376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaiser-Guignard, J.; Canellini, G.; Lion, N.; Abonnenc, M.; Osselaer, J.C.; Tissot, J.D. The clinical and biological impact of new pathogen inactivation technologies on platelet concentrates. Blood Rev. 2014, 28, 235–241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Osselaer, J.C.; Cazenave, J.P.; Lambermont, M.; Garraud, O.; Hidajat, M.; Barbolla, L.; Tardivel, R.; Defoin, L.; Waller, C.; Mendel, I.; et al. An active haemovigilance programme characterizing the safety profile of 7437 platelet transfusions prepared with amotosalen photochemical treatment. Vox Sang. 2008, 94, 315–323. [Google Scholar] [CrossRef]
- Knutson, F.; Osselaer, J.; Pierelli, L.; Lozano, M.; Cid, J.; Tardivel, R.; Garraud, O.; Hervig, T.; Domanovic, D.; Cukjati, M.; et al. A prospective, active haemovigilance study with combined cohort analysis of 19 175 transfusions of platelet components prepared with amotosalen-UVA photochemical treatment. Vox Sang. 2015, 109, 343–352. [Google Scholar] [CrossRef] [PubMed] [Green Version]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Arnason, N.A.; Johannsson, F.; Landrö, R.; Hardarsson, B.; Gudmundsson, S.; Lian, A.-M.; Reseland, J.; Rolfsson, O.; Sigurjonsson, O.E. Protein Concentrations in Stored Pooled Platelet Concentrates Treated with Pathogen Inactivation by Amotosalen Plus Ultraviolet a Illumination. Pathogens 2022, 11, 350. https://doi.org/10.3390/pathogens11030350
Arnason NA, Johannsson F, Landrö R, Hardarsson B, Gudmundsson S, Lian A-M, Reseland J, Rolfsson O, Sigurjonsson OE. Protein Concentrations in Stored Pooled Platelet Concentrates Treated with Pathogen Inactivation by Amotosalen Plus Ultraviolet a Illumination. Pathogens. 2022; 11(3):350. https://doi.org/10.3390/pathogens11030350
Chicago/Turabian StyleArnason, Niels Arni, Freyr Johannsson, Ragna Landrö, Björn Hardarsson, Sveinn Gudmundsson, Aina-Mari Lian, Janne Reseland, Ottar Rolfsson, and Olafur E. Sigurjonsson. 2022. "Protein Concentrations in Stored Pooled Platelet Concentrates Treated with Pathogen Inactivation by Amotosalen Plus Ultraviolet a Illumination" Pathogens 11, no. 3: 350. https://doi.org/10.3390/pathogens11030350