
Was It Chikungunya? Laboratorial and Clinical Investigations of Cases Occurred during a Triple Arboviruses’ Outbreak in Rio de Janeiro, Brazil

 , ,
, , 
 ,
,  ,
, 
Abstract
:1. Introduction
2. Results
2.1. Clinical Diagnosis of Chikungunya
2.2. Baseline Characteristics of Chikungunya Cases
2.3. Clinical Manifestations of Chikungunya Cases
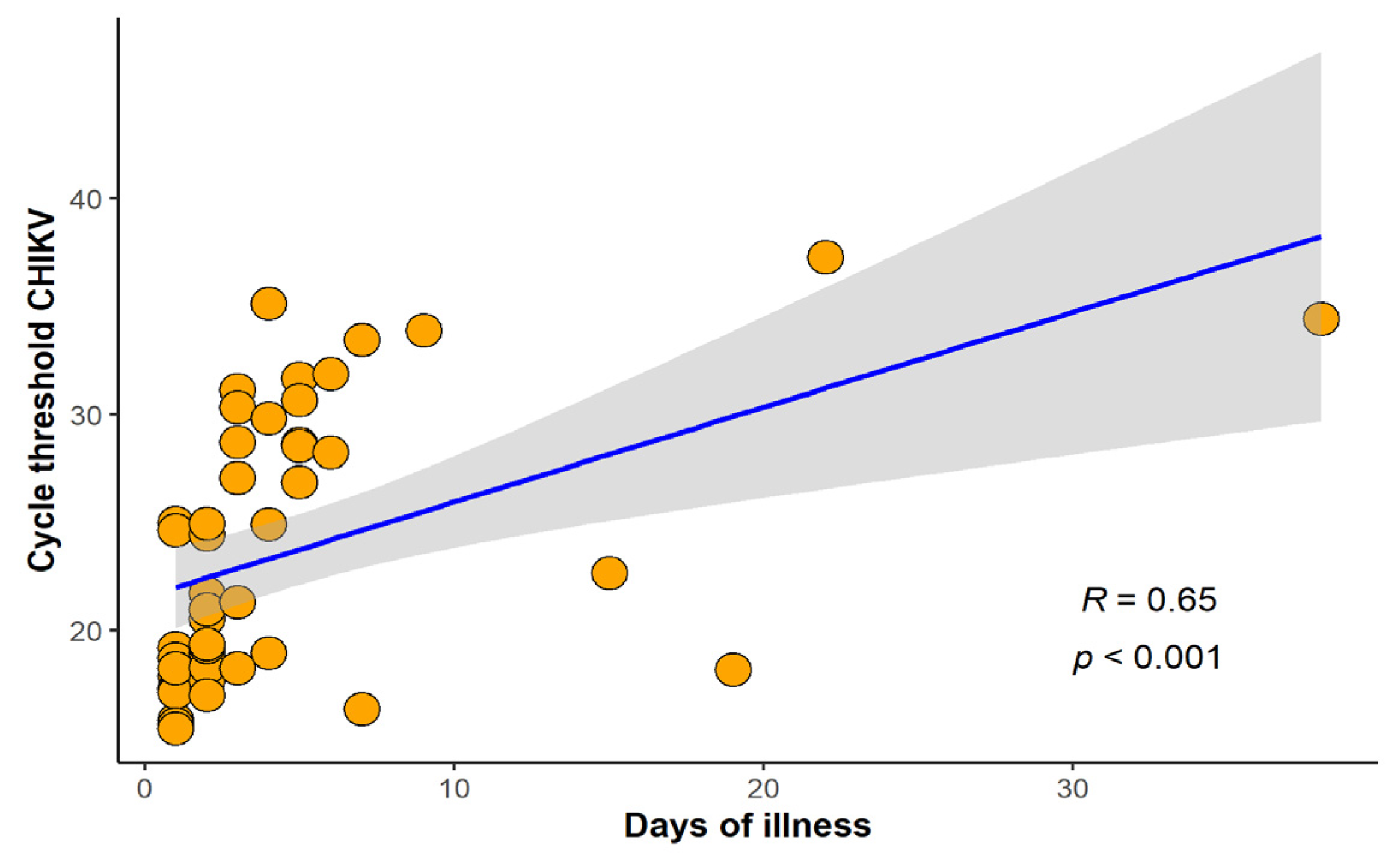
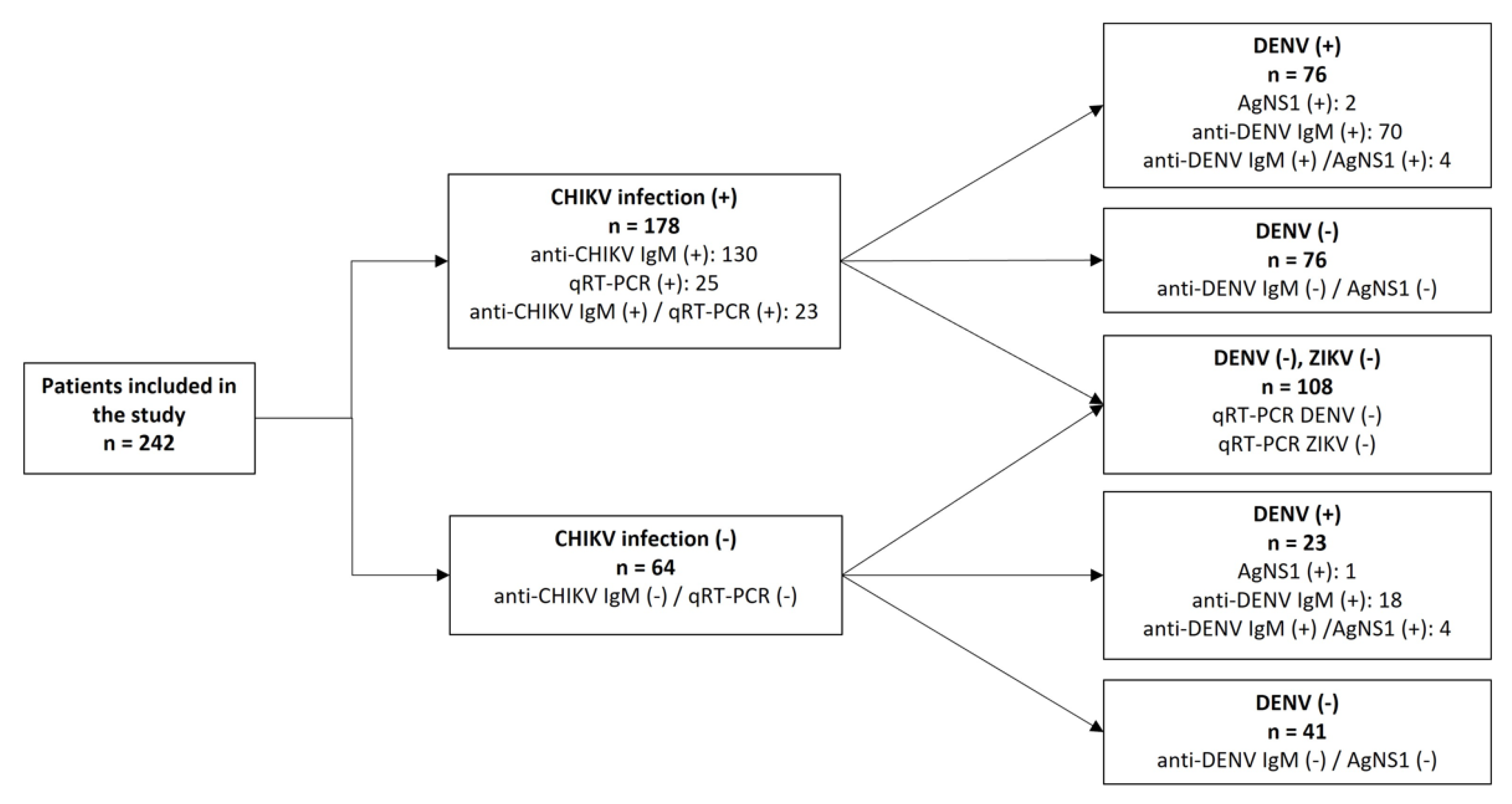
2.4. Flaviviruses Differential Diagnosis
2.5. Genotype Characterization of Representative CHIKV Strains Circulating in Rio de Janeiro in 2018
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Study Sites and Sample Collection
5.2. Serological Diagnosis of Chikungunya and Dengue
5.3. Molecular Diagnosis
5.4. Chikungunya Virus Genotyping
5.5. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Strauss, J.H.; Strauss, E.G. The alphaviruses: Gene expression, replication, and evolution. Microbiol. Rev. 1994, 58, 491–562. [Google Scholar] [CrossRef] [PubMed]
- Lum, F.M.; Ng, L.F. Cellular and molecular mechanisms of chikungunya pathogenesis. Antivir. Res. 2015, 120, 165–174. [Google Scholar] [CrossRef] [PubMed]
- Simizu, B.; Yamamoto, K.; Hashimoto, K.; Ogata, T. Structural proteins of Chikungunya virus. J. Virol. 1984, 51, 254–258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Solignat, M.; Gay, B.; Higgs, S.; Briant, L.; Devaux, C. Replication cycle of Chikungunya: A re-emerging arbovirus. Virology 2009, 393, 183–197. [Google Scholar] [CrossRef] [Green Version]
- Powers, A.M.; Brault, A.C.; Tesh, R.B.; Weaver, S.C. Re-emergence of chikungunya and o’nyong-nyong viruses: Evidence for distinct geographical lineages and distant evolutionary relationships. J. Gen. Virol. 2000, 81, 471–479. [Google Scholar] [CrossRef]
- Volk, S.M.; Chen, R.; Tsetsarkin, K.A.; Adams, A.P.; Garcia, T.I.; Sall, A.A.; Nasar, F.; Schuh, A.J.; Holmes, E.C.; Higgs, S.; et al. Genome-scale phylogenetic analyses of chikungunya virus reveal independent emergences of recent epidemics and various evolutionary rates. J. Virol. 2010, 84, 6497–6504. [Google Scholar] [CrossRef] [Green Version]
- Azeredo, E.L.; Dos Santos, F.B.; Barbosa, L.S.; Souza, T.M.A.; Badolato-Corrêa, J.; Sánchez-Arcila, J.C.; Nunes, P.C.G.; de-Oliveira-Pinto, L.M.; de Filippis, A.M.; Dal Fabbro, M.; et al. Clinical and Laboratory Profile of Zika and Dengue Infected Patients: Lessons Learned From the Co-circulation of Dengue, Zika and Chikungunya in Brazil. PLoS Curr. 2018, 15, 10. [Google Scholar] [CrossRef]
- Carey, D.E. Chikungunya and dengue: A case of mistaken identity? J. Hist. Med. Allied Sci. 1971, 26, 243–262. [Google Scholar] [CrossRef]
- Cunha, R.V.; Trinta, K.S.; Montalbano, C.A.; Sucupira, M.V.; de Lima, M.M.; Marques, E.; Romanholi, I.H.; Croda, J. Seroprevalence of chikungunya virus in a rural community in Brazil. PLoS Negl. Trop. Dis. 2017, 11, e0005319. [Google Scholar] [CrossRef]
- Lumsden, W.H. An epidemic of virus disease in Southern Province, Tanganyika Territory, in 1952–1953. II. General description and epidemiology. Trans. R. Soc. Trop. Med. Hyg. 1955, 49, 33–57. [Google Scholar] [CrossRef]
- Robinson, M.C. An epidemic of virus disease in Southern Province, Tanganyika Territory, in 1952–1953. I. Clinical features. Trans. R. Soc. Trop. Med. Hyg. 1955, 49, 28–32. [Google Scholar] [CrossRef]
- Lo Presti, A.; Cella, E.; Angeletti, S.; Ciccozzi, M. Molecular epidemiology, evolution and phylogeny of Chikungunya virus: An updating review. Infect. Genet. Evol. 2016, 41, 270–278. [Google Scholar] [CrossRef] [PubMed]
- Diallo, M.; Thonnon, J.; Traore-Lamizana, M.; Fontenille, D. Vectors of chikungunya virus in Senegal: Current data and transmission cycles. Am. J. Trop. Med. Hyg. 1999, 60, 281–286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gilotra, S.K.; Shah, K.V. Laboratory studies on transmission of chikungunya virus by mosquitoes. Am. J. Epidemiol. 1967, 86, 379–385. [Google Scholar] [CrossRef]
- Rougeron, V.; Sam, I.C.; Caron, M.; Nkoghe, D.; Leroy, E.; Roques, P. Chikungunya, a paradigm of neglected tropical disease that emerged to be a new health global risk. J. Clin. Virol. Off. Publ. Pan Am. Soc. Clin. Virol. 2015, 64, 144–152. [Google Scholar] [CrossRef]
- Jupp, P.G.; McIntosh, B.M.; Dos Santos, I.; DeMoor, P. Laboratory vector studies on six mosquito and one tick species with Chikungunya virus. Trans. R. Soc. Trop. Med. Hyg. 1981, 75, 15–19. [Google Scholar] [CrossRef]
- Powers, A.M.; Logue, C.H. Changing patterns of Chikungunya virus: Re-emergence of a zoonotic arbovirus. J. Gen. Virol. 2007, 88, 2363–2377. [Google Scholar] [CrossRef]
- Rao, T.R. Vectors of Dengue and Chikungunya viruses: A brief review. Indian J. Med. Res. 1964, 52, 719–726. [Google Scholar]
- Josseran, L.; Paquet, C.; Zehgnoun, A.; Caillere, N.; Le Tertre, A.; Solet, J.L.; Ledrans, M. Chikungunya disease outbreak, Reunion Island. Emerg. Infect. Dis. 2006, 12, 1994–1995. [Google Scholar] [CrossRef]
- Schuffenecker, I.; Iteman, I.; Michault, A.; Murri, S.; Frangeul, L.; Vaney, M.C.; Lavenir, R.; Pardigon, N.; Reynes, J.M.; Pettinelli, F.; et al. Genome microevolution of chikungunya viruses causing the Indian Ocean outbreak. PLoS Med. 2006, 3, e263. [Google Scholar] [CrossRef] [Green Version]
- Pialoux, G.; Gaüzère, B.-A.; Jauréguiberry, S.; Strobel, M. Chikungunya, an epidemic arbovirosis. Lancet Infect. Dis. 2007, 7, 319–327. [Google Scholar] [CrossRef]
- Simon, F.; Savini, H.; Parola, P. Chikungunya: A paradigm of emergence and globalization of vector-borne diseases. Med. Clin. N. Am. 2008, 92, 1323–1343. [Google Scholar] [CrossRef] [PubMed]
- Mohan, A.; Kiran, D.H.; Manohar, I.C.; Kumar, D.P. Epidemiology, clinical manifestations, and diagnosis of Chikungunya fever: Lessons learned from the re-emerging epidemic. Indian J. Dermatol. 2010, 55, 54–63. [Google Scholar] [CrossRef] [PubMed]
- Silva, J.V.J., Jr.; Ludwig-Begall, L.F.; Oliveira-Filho, E.F.; Oliveira, R.A.S.; Durães-Carvalho, R.; Lopes, T.R.R.; Silva, D.E.A.; Gil, L. A scoping review of Chikungunya virus infection: Epidemiology, clinical characteristics, viral co-circulation complications, and control. Acta Trop. 2018, 188, 213–224. [Google Scholar] [CrossRef]
- Nunes, M.R.; Faria, N.R.; de Vasconcelos, J.M.; Golding, N.; Kraemer, M.U.; de Oliveira, L.F.; Azevedo Rdo, S.; da Silva, D.E.; da Silva, E.V.; da Silva, S.P.; et al. Emergence and potential for spread of Chikungunya virus in Brazil. BMC Med. 2015, 13, 102. [Google Scholar] [CrossRef] [Green Version]
- SVS/MS. Monitoramento dos Casos de Arboviroses Urbanas Causados por Vírus Transmitidos Pelo Mosquito Aedes (Dengue, Chikungunya e Zika), Semanas Epidemiológicas 1 a 52 de 2018; Brazilian-Ministry-of-Health: Brasília, Brazil, 2019; Volume 50, p. 14. [Google Scholar]
- Brasil, P.; Calvet, G.A.; Siqueira, A.M.; Wakimoto, M.; de Sequeira, P.C.; Nobre, A.; Quintana Mde, S.; Mendonça, M.C.; Lupi, O.; de Souza, R.V.; et al. Zika virus outbreak in Rio de Janeiro, Brazil: Clinical characterization, epidemiological and virological aspects. PLoS Negl. Trop. Dis. 2016, 10, e0004636. [Google Scholar] [CrossRef]
- Figueiredo, M.L.; Figueiredo, L.T. Emerging alphaviruses in the Americas: Chikungunya and Mayaro. Rev. Soc. Bras. Med. Trop. 2014, 47, 677–683. [Google Scholar] [CrossRef] [Green Version]
- Benelli, G.; Mehlhorn, H. Declining malaria, rising of dengue and Zika virus: Insights for mosquito vector control. Parasitol. Res. 2016, 115, 1747–1754. [Google Scholar] [CrossRef] [Green Version]
- Silva, N.M.d.; Teixeira, R.A.G.; Cardoso, C.G.; Junior, J.B.S.; Coelho, G.E.; Oliveira, E.S.F.d. Chikungunya surveillance in Brazil: Challenges in the context of Public Health. Epidemiol. Serv. Saúde 2018, 27. [Google Scholar] [CrossRef] [Green Version]
- SVS/MS. Febre de Chikungunya: Manejo Clínico; Brazilian-Ministry-of-Health: Brasília, Brazil, 2015; p. 28. [Google Scholar]
- Schatzmayr, H.G.; Nogueira, R.M.R.; Rosa, A.P.A.T. An outbreak of dengue virus at Rio de Janeiro—1986. Mem. Inst. Oswaldo Cruz 1986, 81, 245–246. [Google Scholar] [CrossRef] [Green Version]
- Zanluca, C.; Melo, V.C.; Mosimann, A.L.; Santos, G.I.; Santos, C.N.; Luz, K. First report of autochthonous transmission of Zika virus in Brazil. Mem. Inst. Oswaldo Cruz 2015, 110, 569–572. [Google Scholar] [CrossRef] [PubMed]
- de Souza, T.M.A.; Ribeiro, E.D.; Corrêa, V.C.E.; Damasco, P.V.; Santos, C.C.; de Bruycker-Nogueira, F.; Chouin-Carneiro, T.; Faria, N.; Nunes, P.C.G.; Heringer, M.; et al. Following in the Footsteps of the Chikungunya Virus in Brazil: The First Autochthonous Cases in Amapá in 2014 and Its Emergence in Rio de Janeiro during 2016. Viruses 2018, 10, 623. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silva, M.M.O.; Tauro, L.B.; Kikuti, M.; Anjos, R.O.; Santos, V.C.; Gonçalves, T.S.F.; Paploski, I.A.D.; Moreira, P.S.S.; Nascimento, L.C.J.; Campos, G.S.; et al. Concomitant Transmission of Dengue, Chikungunya, and Zika Viruses in Brazil: Clinical and Epidemiological Findings From Surveillance for Acute Febrile Illness. Clin. Infect. Dis. 2019, 69, 1353–1359. [Google Scholar] [CrossRef] [PubMed]
- Gautret, P.; Simon, F. Dengue, chikungunya and Zika and mass gatherings: What happened in Brazil, 2014. Travel Med. Infect. Dis. 2016, 14, 7–8. [Google Scholar] [CrossRef]
- Alcon, S.; Talarmin, A.; Debruyne, M.; Falconar, A.; Deubel, V.; Flamand, M. Enzyme-linked immunosorbent assay specific to Dengue virus type 1 nonstructural protein NS1 reveals circulation of the antigen in the blood during the acute phase of disease in patients experiencing primary or secondary infections. J. Clin. Microbiol. 2002, 40, 376–381. [Google Scholar] [CrossRef] [Green Version]
- Lima Mda, R.; Nogueira, R.M.; Schatzmayr, H.G.; dos Santos, F.B. Comparison of three commercially available dengue NS1 antigen capture assays for acute diagnosis of dengue in Brazil. PLoS Negl. Trop. Dis. 2010, 4, e738. [Google Scholar] [CrossRef]
- Lima, M.d.R.Q.; Chouin-Carneiro, T.; Azeredo, E.; Barbosa, L.S.; Souza, T.M.A.; Silva, J.B.C.d.; Nunes, P.C.G.; Dal Fabbro, M.; Facco, I.H.R.; Venâncio-da-Cunha, R.; et al. The inability of a dengue NS1 ELISA to detect Zika infections. Sci. Rep. 2019, 9, 18596. [Google Scholar] [CrossRef] [Green Version]
- Muller, D.A.; Depelsenaire, A.C.; Young, P.R. Clinical and Laboratory Diagnosis of Dengue Virus Infection. J. Infect. Dis. 2017, 215, S89–S95. [Google Scholar] [CrossRef]
- Shu, P.Y.; Chen, L.K.; Chang, S.F.; Su, C.L.; Chien, L.J.; Chin, C.; Lin, T.H.; Huang, J.H. Dengue virus serotyping based on envelope and membrane and nonstructural protein NS1 serotype-specific capture immunoglobulin M enzyme-linked immunosorbent assays. J. Clin. Microbiol. 2004, 42, 2489–2494. [Google Scholar] [CrossRef] [Green Version]
- Salam, N.; Mustafa, S.; Hafiz, A.; Chaudhary, A.A.; Deeba, F.; Parveen, S. Global prevalence and distribution of coinfection of malaria, dengue and chikungunya: A systematic review. BMC Public Health 2018, 18, 710. [Google Scholar] [CrossRef]
- Dos Santos, S.M.R.; Sanz Duro, R.L.; Santos, G.L.; Hunter, J.; da Aparecida Rodrigues Teles, M.; Brustulin, R.; de Padua Milagres, F.A.; Sabino, E.C.; Diaz, R.S.; Komninakis, S.V. Detection of coinfection with Chikungunya virus and Dengue virus serotype 2 in serum samples of patients in State of Tocantins, Brazil. J. Infect. Public Health 2020, 13, 724–729. [Google Scholar] [CrossRef] [PubMed]
- Vargas, S.L.; Céspedes, D.C.; Vergel, J.D.; Ruiz, E.P.; Luna, M.C. Coinfection of Dengue and Chikungunya viruses. Narrative review. Rev. Chil. Infectol. 2018, 35, 658–668. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edwards, T.; Signor, L.D.; Williams, C.; Donis, E.; Cuevas, L.E.; Adams, E.R. Co-infections with Chikungunya and Dengue viruses, Guatemala, 2015. Emerg. Infect. Dis. 2016, 22, 2003–2005. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castellanos, J.E.; Jaimes, N.; Coronel-Ruiz, C.; Rojas, J.P.; Mejía, L.F.; Villarreal, V.H.; Maya, L.E.; Claros, L.M.; Orjuela, C.; Calvo, E.; et al. Dengue-chikungunya coinfection outbreak in children from Cali, Colombia, in 2018–2019. Int. J. Infect. Dis. 2020, 102, 97–102. [Google Scholar] [CrossRef]
- Lima, M.; de Lima, R.C.; de Azeredo, E.L.; Dos Santos, F.B. Analysis of a routinely used commercial anti-Chikungunya IgM ELISA reveals cross-reactivities with Dengue in Brazil: A new challenge for differential diagnosis? Diagnostics 2021, 11, 819. [Google Scholar] [CrossRef]
- Abhishek, K.S.; Chakravarti, A. Simultaneous detection of IgM antibodies against dengue and chikungunya: Coinfection or cross-reactivity? J. Fam. Med. Prim. Care 2019, 8, 2420–2423. [Google Scholar] [CrossRef]
- Kaur, M.; Singh, K.; Sidhu, S.K.; Devi, P.; Kaur, M.; Soneja, S.; Singh, N. Coinfection of chikungunya and dengue viruses: A serological study from North Western region of Punjab, India. J. Lab. Physicians 2018, 10, 443–447. [Google Scholar] [CrossRef] [Green Version]
- Freitas, L.P.; Cruz, O.G.; Lowe, R.; Sá Carvalho, M. Space–time dynamics of a triple epidemic: Dengue, chikungunya and Zika clusters in the city of Rio de Janeiro. Proc. R. Soc. B Biol. Sci. 2019, 286, 20191867. [Google Scholar] [CrossRef] [Green Version]
- Queiroz, E.; Medronho, R.A. Spatial analysis of the incidence of Dengue, Zika and Chikungunya and socioeconomic determinants in the city of Rio de Janeiro, Brazil. Epidemiol. Infect. 2021, 149, e188. [Google Scholar] [CrossRef]
- Göertz, G.P.; Vogels, C.B.F.; Geertsema, C.; Koenraadt, C.J.M.; Pijlman, G.P. Mosquito co-infection with Zika and chikungunya virus allows simultaneous transmission without affecting vector competence of Aedes aegypti. PLoS Negl. Trop. Dis. 2017, 11, e0005654. [Google Scholar] [CrossRef] [Green Version]
- Hoarau, J.J.; Jaffar Bandjee, M.C.; Krejbich Trotot, P.; Das, T.; Li-Pat-Yuen, G.; Dassa, B.; Denizot, M.; Guichard, E.; Ribera, A.; Henni, T.; et al. Persistent chronic inflammation and infection by Chikungunya arthritogenic alphavirus in spite of a robust host immune response. J. Immunol. 2010, 184, 5914–5927. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poo, Y.S.; Rudd, P.A.; Gardner, J.; Wilson, J.A.; Larcher, T.; Colle, M.A.; Le, T.T.; Nakaya, H.I.; Warrilow, D.; Allcock, R.; et al. Multiple immune factors are involved in controlling acute and chronic Chikungunya virus infection. PLoS Negl. Trop. Dis. 2014, 8, e3354. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, S.; Dutta, S.K.; Sengupta, S.; Tripathi, A. Evidence of Dengue and Chikungunya virus co-infection and circulation of multiple dengue serotypes in a recent Indian outbreak. Eur. J. Clin. Microbiol. Infect. Dis. 2017, 36, 2273–2279. [Google Scholar] [CrossRef]
- WHO. Chikungunya Geneva2020. [Updated 15/09/2020]. Available online: https://www.who.int/news-room/fact-sheets/detail/chikungunya (accessed on 28 November 2021).
- Bandyopadhyay, D.; Ghosh, S.K. Mucocutaneous features of Chikungunya fever: A study from an outbreak in West Bengal, India. Int. J. Dermatol. 2008, 47, 1148–1152. [Google Scholar] [CrossRef] [PubMed]
- Ali Ou Alla, S.; Combe, B. Arthritis after infection with Chikungunya virus. Best Pract. Res. Clin. Rheumatol. 2011, 25, 337–346. [Google Scholar] [CrossRef] [PubMed]
- Kucharz, E.J.; Cebula-Byrska, I. Chikungunya fever. Eur. J. Intern. Med. 2012, 23, 325–329. [Google Scholar] [CrossRef]
- Javelle, E.; Tiong, T.H.; Leparc-Goffart, I.; Savini, H.; Simon, F. Inflammation of the external ear in acute chikungunya infection: Experience from the outbreak in Johor Bahru, Malaysia, 2008. J. Clin. Virol. Off. Publ. Pan Am. Soc. Clin. Virol. 2014, 59, 270–273. [Google Scholar] [CrossRef] [PubMed]
- Hyle, E.P.; Alame, D. Case records of the Massachusetts General Hospital. Case 13-2015. A 27-year-old woman with arthralgias and a rash. N. Engl. J. Med. 2015, 372, 1657–1664. [Google Scholar] [CrossRef]
- Taraphdar, D.; Sarkar, A.; Mukhopadhyay, B.B.; Chatterjee, S. A comparative study of clinical features between monotypic and dual infection cases with Chikungunya virus and Dengue virus in West Bengal, India. Am. J. Trop. Med. Hyg. 2012, 86, 720–723. [Google Scholar] [CrossRef]
- de Andrade, G.C.; Ventura, C.V.; Mello Filho, P.A.; Maia, M.; Vianello, S.; Rodrigues, E.B. Arboviruses and the eye. Int. J. Retin. Vitr. 2017, 3, 4. [Google Scholar] [CrossRef] [Green Version]
- Inamadar, A.C.; Palit, A.; Sampagavi, V.V.; Raghunath, S.; Deshmukh, N.S. Cutaneous manifestations of chikungunya fever: Observations made during a recent outbreak in south India. Int. J. Dermatol. 2008, 47, 154–159. [Google Scholar] [CrossRef] [PubMed]
- Badawi, A.; Ryoo, S.G.; Vasileva, D.; Yaghoubi, S. Prevalence of chronic comorbidities in chikungunya: A systematic review and meta-analysis. Int. J. Infect. Dis. 2018, 67, 107–113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Almeida Barreto, F.K.; Montenegro, R.M.; Fernandes, V.O.; Oliveira, R.; de Araújo Batista, L.A.; Hussain, A.; de Góes Cavalcanti, L.P. Chikungunya and diabetes, what do we know? Diabetol. Metab. Syndr. 2018, 10, 32. [Google Scholar] [CrossRef] [PubMed]
- Fabri, A.A.; Rodrigues, C.; Santos, C.C.D.; Chalhoub, F.L.L.; Sampaio, S.A.; Faria, N.; Torres, M.C.; Fonseca, V.; Brasil, P.; Calvet, G.; et al. Co-Circulation of two independent clades and persistence of CHIKV-ECSA genotype during epidemic waves in Rio de Janeiro, southeast Brazil. Pathogens 2020, 9, 984. [Google Scholar] [CrossRef]
- Souza, T.M.; Azeredo, E.L.; Badolato-Corrêa, J.; Damasco, P.V.; Santos, C.; Petitinga-Paiva, F.; Nunes, P.C.; Barbosa, L.S.; Cipitelli, M.C.; Chouin-Carneiro, T.; et al. First report of the East-Central South African genotype of Chikungunya virus in Rio de Janeiro, Brazil. PLoS Curr. 2017, 9. [Google Scholar] [CrossRef]
- Harsha, P.K.; Reddy, V.; Rao, D.; Pattabiraman, C.; Mani, R.S. Continual circulation of ECSA genotype and identification of a novel mutation I317V in the E1 gene of Chikungunya viral strains in southern India during 2015–2016. J. Med. Virol. 2020, 92, 1007–1012. [Google Scholar] [CrossRef]
- Kaur, N.; Jain, J.; Kumar, A.; Narang, M.; Zakaria, M.K.; Marcello, A.; Kumar, D.; Gaind, R.; Sunil, S. Chikungunya outbreak in Delhi, India, 2016: Report on coinfection status and comorbid conditions in patients. New Microbes New Infect. 2017, 20, 39–42. [Google Scholar] [CrossRef]
- Estofolete, C.F.; Terzian, A.C.B.; Colombo, T.E.; de Freitas Guimarães, G.; Ferraz, H.C.J.; da Silva, R.A.; Greque, G.V.; Nogueira, M.L. Co-infection between Zika and different Dengue serotypes during DENV outbreak in Brazil. J. Infect. Public Health 2019, 12, 178–181. [Google Scholar] [CrossRef]
- IBGE. Rio de Janeiro|Cidades e Estados|IBGE. Available online: https://www.ibge.gov.br/cidades-e-estados/rj/ (accessed on 20 November 2021).
- IBGE. Rio de Janeiro (RJ)|Cidades e Estados|IBGE. Available online: https://www.ibge.gov.br/cidades-e-estados/rj/rio-de-janeiro.html (accessed on 20 November 2021).
- IBGE. Campos dos Goytacazes (RJ)|Cidades e Estados|IBGE. Available online: https://www.ibge.gov.br/cidades-e-estados/rj/campos-dos-goytacazes.html (accessed on 20 November 2021).
- Lanciotti, R.S.; Kosoy, O.L.; Laven, J.J.; Panella, A.J.; Velez, J.O.; Lambert, A.J.; Campbell, G.L. Chikungunya virus in US travelers returning from India, 2006. Emerg. Infect. Dis. 2007, 13, 764–767. [Google Scholar] [CrossRef]
- Lanciotti, R.S.; Calisher, C.H.; Gubler, D.J.; Chang, G.J.; Vorndam, A.V. Rapid detection and typing of Dengue viruses from clinical samples by using reverse transcriptase-polymerase chain reaction. J. Clin. Microbiol. 1992, 30, 545–551. [Google Scholar] [CrossRef] [Green Version]
- Johnson, B.W.; Russell, B.J.; Lanciotti, R.S. Serotype-specific detection of dengue viruses in a fourplex real-time reverse transcriptase PCR assay. J. Clin. Microbiol. 2005, 43, 4977–4983. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lanciotti, R.S.; Kosoy, O.L.; Laven, J.J.; Velez, J.O.; Lambert, A.J.; Johnson, A.J.; Stanfield, S.M.; Duffy, M.R. Genetic and serologic properties of Zika virus associated with an epidemic, Yap State, Micronesia, 2007. Emerg. Infect. Dis. 2008, 14, 1232–1239. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT Multiple Sequence Alignment Software Version 7: Improvements in Performance and Usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [Green Version]
- Myung, I.J. Tutorial on maximum likelihood estimation. J. Math. Psychol. 2003, 47, 90–100. [Google Scholar] [CrossRef]
- Nguyen, L.T.; Schmidt, H.A.; von Haeseler, A.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum-likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef]
- Felsenstein, J. Confidence limits on phylogenies: An approach using the bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Total n = 178 | Acute n = 147 | Subacute n = 31 | p | ||||
|---|---|---|---|---|---|---|---|
| Age (years old) | |||||||
| median, IQR | 44 | (28–56) | 42 | (26–56) | 51 | (33–62) | 0.063 |
| Age groups | |||||||
| ≤15 years | 12 | 7% | 11 | 7% | 1 | 3% | |
| 16–30 years | 42 | 24% | 38 | 26% | 4 | 13% | |
| 31–45 years | 42 | 24% | 34 | 23% | 8 | 26% | |
| 46–59 years | 48 | 27% | 39 | 27% | 9 | 29% | |
| ≥60 years | 34 | 19% | 25 | 17% | 9 | 29% | |
| Sex | |||||||
| Male | 64 | 36% | 56 | 38% | 8 | 26% | 0.195 |
| Female | 114 | 64% | 91 | 62% | 23 | 74% | |
| Comorbidity | |||||||
| Overall | 42 | 24% | 32 | 22% | 10 | 32% | 0.211 |
| Stratified | |||||||
| Hypertension | 25 | 14% | 18 | 12% | 7 | 23% | 0.155 |
| Diabetes | 8 | 4% | 5 | 3% | 3 | 10% | 0.145 |
| Sinusitis | 7 | 4% | 4 | 3% | 3 | 10% | 0.103 |
| Rhinitis | 4 | 2% | 3 | 2% | 1 | 3% | 0.538 |
| Asthma | 2 | 1% | 1 | 1% | 1 | 3% | 0.319 |
| Heart disease | 2 | 1% | 2 | 1% | 0 | 0% | 1.000 |
| Others | 5 | 3% | 4 | 3% | 1 | 3% | 1.000 |
| Signs and symptoms | |||||||
| Fever | 178 | 100% | 147 | 100% | 31 | 100% | 1.000 |
| Polyarthralgia | 158 | 89% | 131 | 89% | 27 | 87% | 0.746 |
| Location not reported | 52 | 33% | 43 | 33% | 9 | 33% | |
| Location reported * | 106 | 67% | 88 | 67% | 18 | 67% | |
| Hand/wrist | 35 | 33% | 29 | 33% | 6 | 33% | |
| Foot/ankle | 28 | 26% | 22 | 25% | 6 | 33% | |
| knee | 21 | 20% | 19 | 22% | 2 | 11% | |
| shoulder | 14 | 13% | 12 | 14% | 2 | 11% | |
| elbow | 8 | 8% | 6 | 7% | 2 | 11% | |
| Myalgia | 123 | 69% | 101 | 69% | 22 | 71% | 0.805 |
| Headache | 111 | 62% | 93 | 63% | 18 | 58% | 0.587 |
| Exanthema | 95 | 53% | 78 | 53% | 17 | 55% | 0.857 |
| Prostration | 81 | 46% | 67 | 46% | 14 | 45% | 0.966 |
| Pruritus | 82 | 46% | 63 | 43% | 19 | 61% | 0.061 |
| Nausea | 67 | 38% | 57 | 39% | 10 | 32% | 0.496 |
| Retro-orbital pain | 66 | 37% | 57 | 39% | 9 | 29% | 0.307 |
| Hyporexia | 58 | 33% | 50 | 34% | 8 | 26% | 0.376 |
| Lower limb swelling | 57 | 32% | 46 | 31% | 11 | 35% | 0.649 |
| Lower back pain | 49 | 28% | 42 | 29% | 7 | 23% | 0.497 |
| Vomiting | 34 | 19% | 24 | 16% | 10 | 32% | 0.040 |
| Diarrhea | 31 | 17% | 23 | 16% | 8 | 26% | 0.175 |
| Asthenia | 27 | 15% | 23 | 16% | 4 | 13% | 1.000 |
| Conjunctival hyperemia | 26 | 15% | 23 | 16% | 3 | 10% | 0.577 |
| Dizziness | 27 | 15% | 23 | 16% | 4 | 13% | 1.000 |
| Abdominal pain | 24 | 13% | 21 | 14% | 3 | 10% | 0.772 |
| Arthritis | 14 | 8% | 11 | 7% | 3 | 10% | 0.714 |
| Cough | 14 | 8% | 14 | 10% | 0 | 0% | 0.134 |
| Paresthesia | 12 | 7% | 10 | 7% | 2 | 6% | 1.000 |
| Chills | 11 | 6% | 11 | 7% | 0 | 0% | 0.216 |
| Co-detection | |||||||
| Chikungunya/dengue | 76 | 43% | 62 | 42% | 14 | 45% | 0.760 |
| Chikungunya Cases n = 102 | Recent Dengue Cases n = 23 | Chikungunya/Dengue Cases n = 76 | p | ||||
|---|---|---|---|---|---|---|---|
| Age (years) | |||||||
| median, IQR | 43 | (28–59) | 44 | (38–56) | 44 | (28–56) | 0.833 |
| Sex | |||||||
| Male | 36 | 35% | 14 | 61% | 28 | 37% | 0.068 |
| Female | 66 | 65% | 9 | 39% | 48 | 63% | |
| Comorbidity | |||||||
| Overall | 32 | 31% | 2 | 9% | 10 | 13% | 0.004 |
| Stratified | |||||||
| Hypertension | 20 | 20% | 1 | 4% | 5 | 7% | 0.013 |
| Diabetes | 5 | 5% | 1 | 4% | 3 | 4% | 1.000 |
| Sinusitis | 7 | 7% | 0 | 0% | 0 | 0% | 0.026 |
| Rhinitis | 4 | 4% | 0 | 0% | 0 | 0% | 0.233 |
| Asthma | 1 | 1% | 0 | 0% | 1 | 1% | 1.000 |
| Heart disease | 2 | 2% | 0 | 0% | 0 | 0% | 0.614 |
| Others | 4 | 4% | 0 | 0% | 1 | 1% | 0.543 |
| Signs and symptoms | |||||||
| Fever | 102 | 100% | 23 | 100% | 76 | 100% | 1.000 |
| Polyarthralgia | 92 | 90% | 17 | 74% | 66 | 87% | 0.109 |
| Location not reported | 24 | 26% | 12 | 71% | 28 | 42% | |
| Location reported * | 68 | 74% | 5 | 29% | 38 | 58% | |
| Hand/wrist | 23 | 34% | 2 | 40% | 12 | 32% | |
| Foot/ankle | 17 | 25% | 1 | 20% | 11 | 29% | |
| Knee | 13 | 19% | 0 | 0% | 8 | 21% | |
| Shoulder | 8 | 12% | 0 | 0% | 6 | 16% | |
| Elbow | 7 | 10% | 2 | 40% | 1 | 3% | |
| Myalgia | 72 | 71% | 15 | 65% | 51 | 67% | 0.824 |
| Exanthema | 60 | 59% | 12 | 52% | 35 | 46% | 0.239 |
| Headache | 66 | 65% | 13 | 57% | 45 | 46% | 0.654 |
| Prostration | 51 | 50% | 7 | 30% | 30 | 39% | 0.147 |
| Pruritus | 54 | 53% | 9 | 39% | 28 | 37% | 0.084 |
| Nausea | 41 | 40% | 7 | 30% | 26 | 34% | 0.570 |
| Retro-orbital pain | 45 | 44% | 8 | 35% | 21 | 28% | 0.077 |
| Hyporexia | 37 | 36% | 3 | 13% | 21 | 28% | 0.074 |
| Lower limb swelling | 40 | 39% | 0 | 0% | 17 | 22% | <0.001 |
| Lower back pain | 33 | 32% | 6 | 26% | 16 | 21% | 0.244 |
| Vomiting | 22 | 22% | 6 | 26% | 12 | 16% | 0.464 |
| Diarrhea | 15 | 15% | 7 | 30% | 16 | 21% | 0.183 |
| Asthenia | 17 | 17% | 2 | 9% | 10 | 13% | 0.570 |
| Conjunctival hyperemia | 15 | 15% | 1 | 4% | 11 | 14% | 0.397 |
| Dizziness | 20 | 20% | 3 | 13% | 7 | 9% | 0.151 |
| Abdominal pain | 17 | 17% | 0 | 0% | 7 | 9% | 0.054 |
| Arthritis | 12 | 12% | 0 | 0% | 2 | 3% | 0.023 |
| Cough | 11 | 11% | 3 | 13% | 3 | 4% | 0.189 |
| Paresthesia | 9 | 9% | 0 | 0% | 3 | 4% | 0.238 |
| Chills | 7 | 7% | 0 | 0% | 4 | 5% | 0.559 |
| Primer Identification | Primer Sequence (5′-3′) | * Position in the Genome |
|---|---|---|
| CHIK 1A | ACT GCT CTA CTC TGC AAA GC | 39_F |
| CHIK 1B | CTC CGG CGT GAC TTC TGT A | 1136_R |
| CHIK 2A | CCG TGT GCT GTT CTC AGT AG | _788_F |
| CHIK 2B | GTT CTG CTT CTC GTT CTT CC | 1590_R |
| CHIK 3A | AGG AGT GCC GGA AAG ACA TG | 1264_F |
| CHIK 3B | CCT GCA GCT TCT TCC TTC | 2128_R |
| CHIK 4A | TGG TAC TTT CCC CGC AGA C | 1756_F |
| CHIK 4B | TCA CAG GCA GTG TAC ACC | 2634_R |
| CHIK 5A | GGC AAG ACC TGG TGA CTA GC | 2287_F |
| CHIK 5B | ATA GGG ACC AAG CTC TTA GC | _3139_R |
| CHIK 6A | GTG CTT CAG AGG GTG GGT TA | 2756_F |
| CHIK 6B | GTG ACT CTC TTA GTA GGC AG | 3637_R |
| CHIK 7A | CCT GAA TGA AAT ATG CAC GCG C | 3233_F |
| CHIK 7B | TTC TTC GCG ATG TCC ATG C | 4117_R |
| CHIK 8A | ACG CAA TGA AAC TGC AAA TG | 3784_F |
| CHIK 8B | CGT GGT GCT GTA TCC TTT TC | 4655_R |
| CHIK 9A | CCT ATC GAG AAG TCG CAA AG | 4339_F |
| CHIK 9B | ATT ACC CAG TCA GAC ACG G | 5257_R |
| CHIK 10A | GAG CAA GTC TGC CTA TAT GC | 4758_F |
| CHIK 10B | ACG TGG ACC AGT CGC TAT C | 5622_R |
| CHIK 11A | ACT GGG TAA TGA GCA CCG TAC | 5248_F |
| CHIK 11B | TGA CGG ATT GAA TGT CGC TC | 6167_R |
| CHIK 12A | ACG AGG AGA AGT GTT ACC CAC | 5758_F |
| CHIK 12B | GCC TGT ATA ACC TGC ACC | 6607_R |
| CHIK 13A | GCA ACG TCA CAC AGA TGA GG | 6286_F |
| CHIK 13B | CCA TCA ATT CAT CGG AGA CG | 7104_R |
| CHIK 14A | AGC CGC ACA CTT TAA GCC AG | 6737_F |
| CHIK 14B | AGG CTG GTA CCT CCT ATT G | 7615_R |
| CHIK 15A | ATC AGA TGG CAA CGA ACA GG | 7329_F |
| CHIK 15B | GTG GTG CCA GTT GTA GTA C | 8140_R |
| CHIK 16A | CGG AAG AAT AAG AAG CAA AAG C | 7760_F |
| CHIK 16B | AGT GCC CTT CTC CAC AGT C | 8621_R |
| CHIK 17A | AAT GAA GGA GCC CGT ACA GC | 8264_F |
| CHIK 17B | TTG CCG GAC TGT TGT GAC | 9102_R |
| CHIK 18A | ACC GTG CAC GAT TAC TGG AAC | 8809_F |
| CHIK 18B | CAG AAT TAT CTC ATG CGG GTG G | 9613_R |
| CHIK 19A | TGC AGG GTG CCT AAA GCA AG | 9338_F |
| CHIK 19B | GTA ATC AAG CGA TAG CGT TGG | 10132_R |
| CHIK 20A | TAC CGT CCC TTT CCT GCT TA | 9760_F |
| CHIK 20B | AAT TGT CCT GGT CTT CCT GC | 10593_R |
| CHIK 21A | GAA GTC CGA ATC ATG CAA AAC | 10321_F |
| CHIK 21B | GTG TAC TTG TGT AGA ACA GAC | 11119_R |
| CHIK 22A | AGC AAC AAA CCC GGT AAG AG | 10777_F |
| CHIK 22B | TAG TTG TCA AGT TAG TGC CTG C | 11325_R |
| CHIK 23A | ATG GGT GCA GAA GAT CAC G | 11218_F |
| CHIK 23B ECSA | GTA TAG CCC TTT GAA CTA CTT C | 11613_R |
| CHIK 23B ASIAN | GCT ATA TAT GGT GTG TCT CTT AGG | 11522_R |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
de Souza, T.M.A.; de Lima, R.C.; Solórzano, V.E.F.; Damasco, P.V.; de Souza, L.J.; Sanchez-Arcila, J.C.; Guimarães, G.M.C.; Paiva, I.A.; da Rocha Queiroz Lima, M.; de Bruycker-Nogueira, F.; et al. Was It Chikungunya? Laboratorial and Clinical Investigations of Cases Occurred during a Triple Arboviruses’ Outbreak in Rio de Janeiro, Brazil. Pathogens 2022, 11, 245. https://doi.org/10.3390/pathogens11020245
de Souza TMA, de Lima RC, Solórzano VEF, Damasco PV, de Souza LJ, Sanchez-Arcila JC, Guimarães GMC, Paiva IA, da Rocha Queiroz Lima M, de Bruycker-Nogueira F, et al. Was It Chikungunya? Laboratorial and Clinical Investigations of Cases Occurred during a Triple Arboviruses’ Outbreak in Rio de Janeiro, Brazil. Pathogens. 2022; 11(2):245. https://doi.org/10.3390/pathogens11020245
Chicago/Turabian Stylede Souza, Thiara Manuele Alves, Raquel Curtinhas de Lima, Victor Edgar Fiestas Solórzano, Paulo Vieira Damasco, Luiz José de Souza, Juan Camilo Sanchez-Arcila, Gabriel Macedo Costa Guimarães, Iury Amâncio Paiva, Monique da Rocha Queiroz Lima, Fernanda de Bruycker-Nogueira, and et al. 2022. "Was It Chikungunya? Laboratorial and Clinical Investigations of Cases Occurred during a Triple Arboviruses’ Outbreak in Rio de Janeiro, Brazil" Pathogens 11, no. 2: 245. https://doi.org/10.3390/pathogens11020245