
Mosquito Communities Vary across Landscape and Vertical Strata in Indian River County, Florida

 , ,
, ,
Abstract
:1. Introduction
2. Results
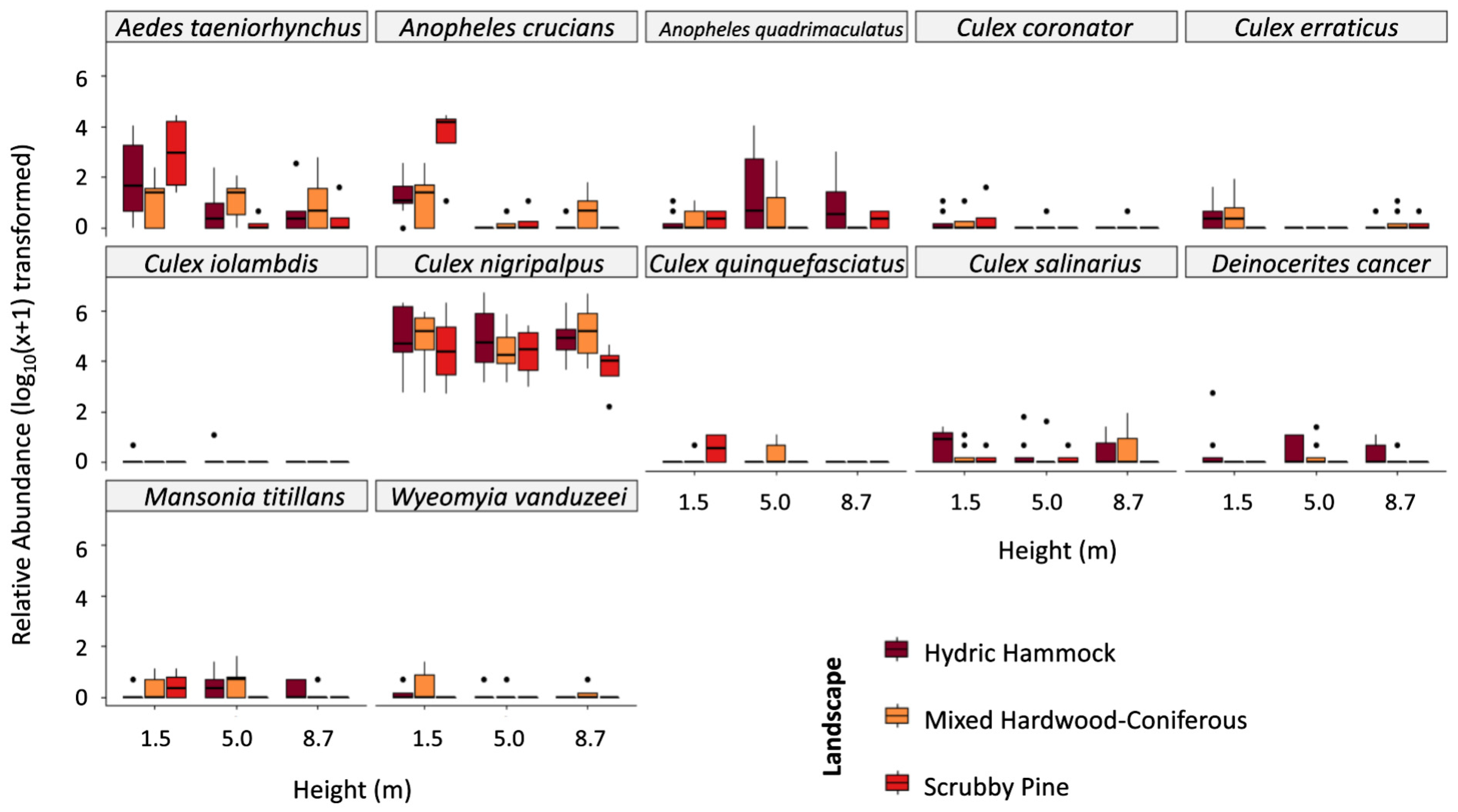
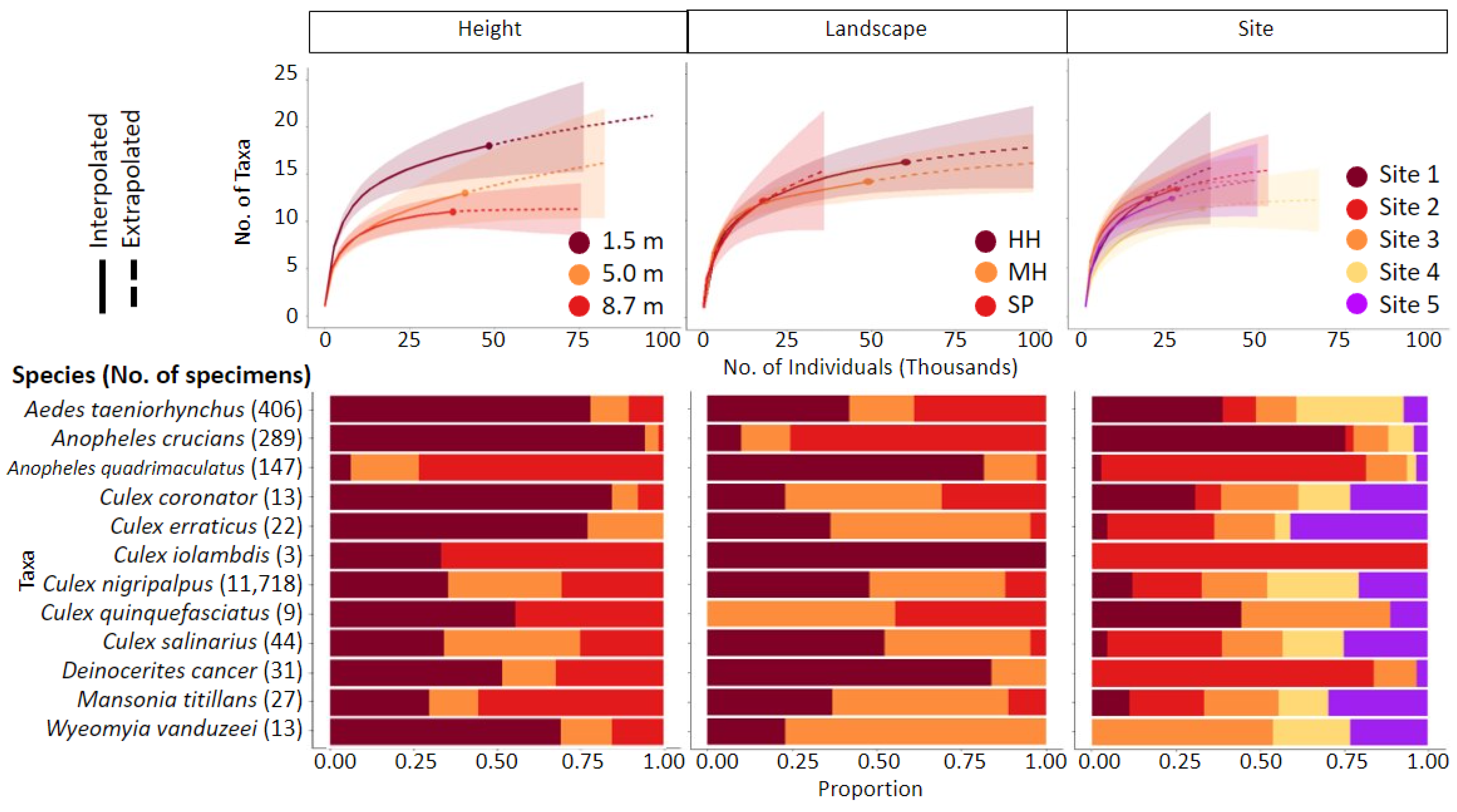
2.1. Mosquito Collections
2.2. Trapping Effort across Strata, Landscape, and Site Location
2.3. Permutational MANOVA
2.4. Generalized Linear Mixed-Effects Model
3. Discussion
4. Materials and Methods
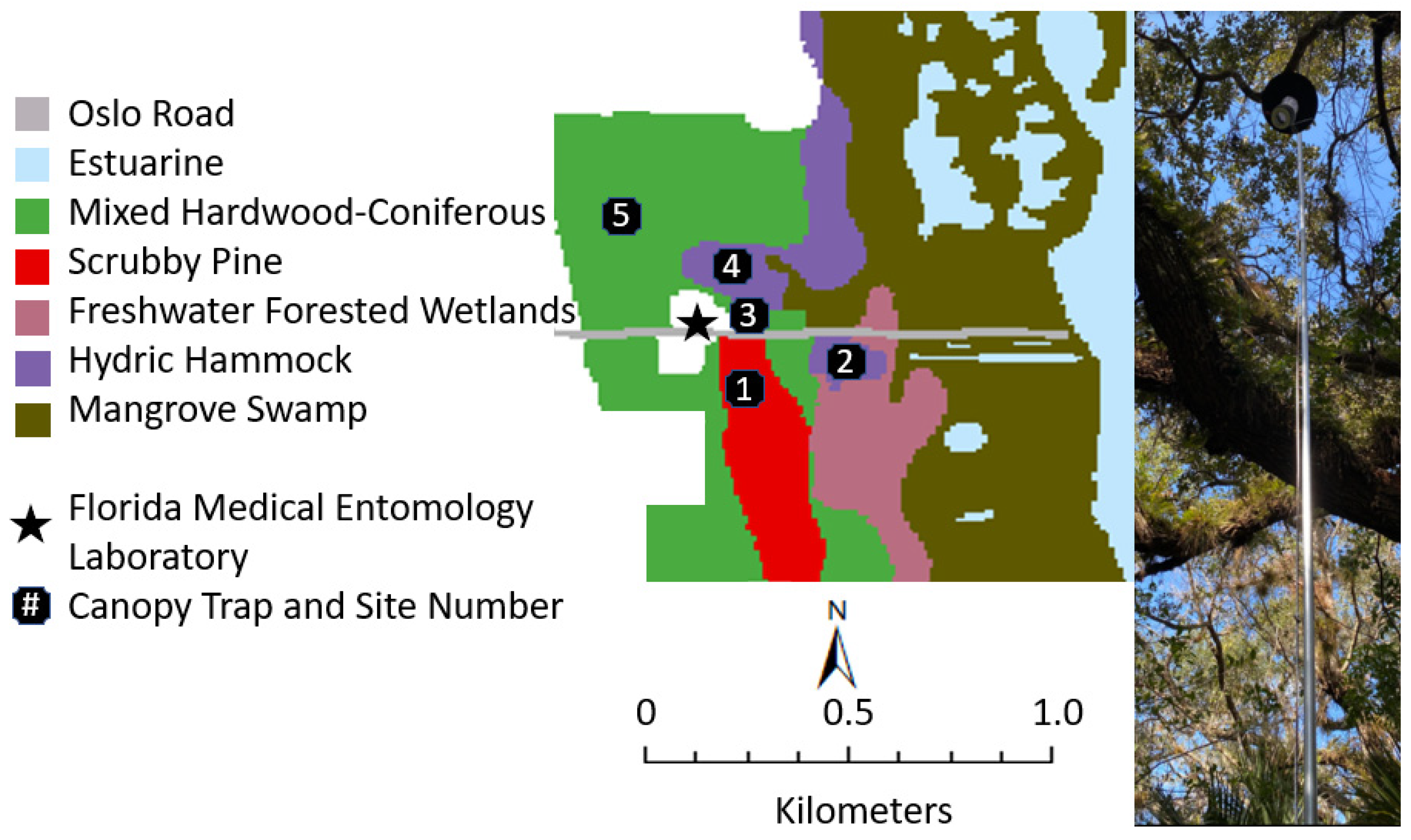
4.1. Sampling Locations and Canopy Trap Construction
4.2. Mosquito Collection and Identification
4.3. RNA Extraction and RT-PCR Assays
4.4. Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- McDermott, E.G.; Mullens, B.A. The Dark Side of Light Traps. J. Med. Entomol. 2018, 55, 251–261. [Google Scholar] [CrossRef]
- Komar, N.; Panella, N.A.; Burkhalter, K.L. Focal Amplification and Suppression of West Nile Virus Transmission Associated with Communal Bird Roosts in Northern Colorado. J. Vector Ecol. 2018, 43, 220–234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Russell, C.; Hunter, F.F. Influence of Elevation and Avian or Mammalian Hosts on Attraction of Culex pipiens (Diptera: Culicidae) in Southern Ontario. Can. Entomol. 2010, 142, 250–255. [Google Scholar] [CrossRef]
- Anderson, J.F.; Andreadis, T.G.; Main, A.J.; Kline, D.L. Prevalence of West Nile Virus in Tree Canopy-Inhabitng Culex pipiens and Associated Mosquitoes. Am. J. Trop. Med. Hyg. 2004, 71, 112–119. [Google Scholar] [CrossRef] [PubMed]
- Andreadis, T.G.; Armstrong, P.M. A Two-Year Evaluation of Elevated Canopy Trapping for Culex Mosquitoes and West Nile Virus in an Operational Surveillance Program in the Northeastern United States. J. Am. Mosq. Control Assoc. 2007, 23, 137–148. [Google Scholar] [CrossRef]
- Drummond, C.L.; Drobnack, J.; Backenson, P.B.; Ebel, G.D.; Kramer, L.D. Impact of Trap Elevation on Estimates of Abundance, Parity Rates, and Body Size of Culex pipiens and Culex restuans (Diptera: Culicidae). J. Med. Entomol. 2006, 43, 8. [Google Scholar] [CrossRef]
- Swanson, D.A.; Adler, P.H. Vertical Distribution of Haematophagous Diptera in Temperate Forests of the Southeastern USA. Med. Vet. Entomol. 2010, 24, 182–188. [Google Scholar] [CrossRef]
- Anderson, J.F.; Andreadis, T.G.; Main, A.J.; Ferrandino, F.J.; Vossbrinck, C.R. West Nile Virus from Female and Male Mosquitoes (Diptera: Culicidae) in Subterranean, Ground, and Canopy Habitats in Connecticut. J. Med. Entomol. 2006, 43, 10. [Google Scholar] [CrossRef]
- Russell, C.B.; Hunter, F.F. Attraction of Culex Pipiens/Restuans (Diptera: Culicidae) Mosquitoes to Bird Uropygial Gland Odors at Two Elevations in the Niagara Region of Ontario. J. Med. Entomol. 2005, 42, 5. [Google Scholar] [CrossRef]
- Deegan, C.S.; Burns, J.E.; Huguenin, M.; Steinhaus, E.Y.; Panella, N.A.; Beckett, S.; Komar, N. Sentinel Pigeon Surveillance for West Nile Virus by Using Lard-Can Traps at Differing Elevations and Canopy Cover Classes. J. Med. Entomol. 2005, 42, 6. [Google Scholar] [CrossRef]
- Darbro, J.M.; Harrington, L.C. Bird-Baited Traps for Surveillance of West Nile Mosquito Vectors: Effect of Bird Species, Trap Height, and Mosquito Escape Rates. J. Med. Entomol. 2006, 43, 10. [Google Scholar] [CrossRef]
- Russell, C.; Hunter, F.F. Culex Pipiens (Culicidae) Is Attracted to Humans in Southern Ontario, but Will It Serve as a Bridge Vector of West Nile Virus? Can. Entomol. 2012, 144, 667–671. [Google Scholar] [CrossRef]
- Savage, H.M.; Anderson, M.; Gordon, E.; Mcmillen, L.; Colton, L.; Delorey, M.; Sutherland, G.; Aspen, S.; Charnetzky, D.; Burkhalter, K.; et al. Host-Seeking Heights, Host-Seeking Activity Patterns, and West Nile Virus Infection Rates for Members of the Culex pipiens Complex at Different Habitat Types within the Hybrid Zone, Shelby County, TN, 2002 (Diptera: Culicidae). J. Med. Entomol. 2008, 45, 13. [Google Scholar] [CrossRef] [Green Version]
- Mackay, A.J.; Roy, A.; Yates, M.M.; Foil, L.D. West Nile Virus Detection in Mosquitoes in East Baton Rouge Parish, Louisiana, from November 2002 to October 2004. J. Am. Mosq. Control Assoc. 2008, 24, 28–35. [Google Scholar] [CrossRef] [PubMed]
- Love, G.J.; Smith, W.W. The Stratification of Mosquitoes. Mosq. News 1958, 18, 279–283. [Google Scholar]
- Love, G.J.; Platt, R.B.; Goddwin, M.H., Jr. Observations on the Spatial Distribution of Mosquitoes in Southwestern Georgia. Mosq. News 1963, 23, 13–22. [Google Scholar]
- Schwarz, M.; Byrd, B.D.; Marayati, B.F.; Blum, P.W.; Wells, M.B.; Greene, A.D.; Taylor, M.; Wasserberg, G. Horizontal Distribution Affects the Vertical Distribution of Native and Invasive Container-inhabiting Aedes Mosquitoes within an Urban Landscape. J. Vector Ecol. 2020, 45, 16–24. [Google Scholar] [CrossRef]
- Godsey, M.S.; King, R.J.; Burkhalter, K.; Delorey, M.; Colton, L.; Charnetzky, D.; Sutherland, G.; Ezenwa, V.O.; Wilson, L.A.; Coffey, M.; et al. Ecology of Potential West Nile Virus Vectors in Southeastern Louisiana: Enzootic Transmission in the Relative Absence of Culex quinquefasciatus. Am. J. Trop. Med. Hyg. 2013, 88, 986–996. [Google Scholar] [CrossRef] [Green Version]
- Obenauer, P.J.; Kaufman, P.E.; Allan, S.A.; Kline, D.L. Infusion-Baited Ovitraps to Survey Ovipositional Height Preferences of Container-Inhabiting Mosquitoes in Two Florida Habitats. J. Med. Entomol. 2009, 46, 1507–1513. [Google Scholar] [CrossRef] [Green Version]
- Obenauer, P.J.; Kaufman, P.E.; Allan, S.A.; Kline, D.L. Host-Seeking Height Preferences of Aedes Albopictus (Diptera: Culicidae) in North Central Florida Suburban and Sylvatic Locales. J. Med. Entomol. 2009, 46, 900–908. [Google Scholar] [CrossRef] [Green Version]
- Smith, J.P.; Walsh, J.D.; Cope, E.H.; Tennant, R.A.; Kozak, J.A.; Darsie, R.F. Culex coronator Dyar and Knab: A New Florida Species Record. J. Am. Mosq. Control Assoc. 2006, 22, 330–332. [Google Scholar] [CrossRef]
- Shin, D.; O’Meara, G.F.; Civana, A.; Shroyer, D.A.; Miqueli, E. Culex interrogator (Diptera: Culicidae), a Mosquito Species New to Florida. J. Vector Ecol. 2016, 41, 316–319. [Google Scholar] [CrossRef]
- Burkett-Cadena, N.D.; Blosser, E.M. Aedeomyia Squamipennis (Diptera: Culicidae) in Florida, USA, a New State and Country Record. J. Med. Entomol. 2017, 54, 788–792. [Google Scholar] [CrossRef] [PubMed]
- Darsie, R.F., Jr.; Shroyer, D.A. Culex (Culex) declarator, a Mosquito Speces New to Florida. J. Am. Mosq. Control Assoc. 2004, 20, 224–227. [Google Scholar] [PubMed]
- Reeves, L.E.; Medina, J.; Miqueli, E.; Sloyer, K.E.; Petrie, W.; Vasquez, C.; Burkett-Cadena, N.D. Establishment of Aedes (Ochlerotatus) Scapularis (Diptera: Culicidae) in Mainland Florida, With Notes on the Ochlerotatus Group in the United States. J. Med. Entomol. 2021, 58, 717–729. [Google Scholar] [CrossRef] [PubMed]
- Kondapaneni, R.; Malcolm, A.N.; Vazquez, B.M.; Zeng, E.; Chen, T.-Y.; Kosinski, K.J.; Romero-Weaver, A.L.; Giordano, B.V.; Allen, B.; Riles, M.T.; et al. Mosquito Control Priorities in Florida—Survey Results from Florida Mosquito Control Districts. Pathogens 2021, 10, 947. [Google Scholar] [CrossRef] [PubMed]
- Florida Department of Health Mosquito-Borne Disease Surveillance. Weekly Florida Arbovirus Reports. 2021. Available online: http://www.floridahealth.gov/diseases-and-conditions/mosquito-borne-diseases/surveillance.html (accessed on 2 December 2021).
- Manrique-Saide, P.; Escobedo-Ortegón, J.; Bolio-González, M.; Sauri-Arceo, C.; Dzib-Florez, S.; Guillermo-May, G.; Ceh-Pavía, E.; Lenhart, A. Incrimination of the Mosquito, Aedes taeniorhynchus, as the Primary Vector of Heartworm, Dirofilaria immitis, in Coastal Yucatan, Mexico. Med. Vet. Entomol. 2010, 24, 456–460. [Google Scholar] [CrossRef] [PubMed]
- King, W.V. Experiments on the Development of Malaria Parasites in Theee American Species of Anopheles. J. Exp. Med. 1916, 23, 703–716. [Google Scholar] [CrossRef] [Green Version]
- Levine, R.S.; Peterson, A.T.; Benedict, M.Q. Distribution of Members of Anopheles quadrimaculatus Say s.l. (Diptera: Culicidae) and Implications for Their Roles in Malaria Transmission in the United States. J. Med. Entomol. 2004, 41, 607–613. [Google Scholar] [CrossRef] [Green Version]
- Alto, B.W.; Connelly, C.R.; O’Meara, G.F.; Hickman, D.; Karr, N. Reproductive Biology and Susceptibility of Florida Culex coronator to Infection with West Nile Virus. Vector-Borne Zoonotic Dis. 2014, 14, 606–614. [Google Scholar] [CrossRef] [Green Version]
- Connelly, C.R.; Alto, B.W.; O’Meara, G.F. The Spread of Culex coronator (Diptera: Culicidae) throughout Florida. J. Vector Ecol. 2016, 41, 195–199. [Google Scholar] [CrossRef] [Green Version]
- Rutledge, C.R.; Day, J.F.; Lord, C.C.; Stark, L.M.; Tabachnick, W.J. West Nile Virus Infection Rates in Culex nigripalpus (Diptera: Culicidae) Do Not Reflect Transmission Rates in Florida. J. Med. Entomol. 2003, 40, 253–258. [Google Scholar] [CrossRef] [PubMed]
- Day, J.F.; Curtis, G.A. When It Rains, They Soar—and That Makes Culex nigripalpus a Dangerous Mosquito. Am. Entomol. 1994, 40, 162–167. [Google Scholar] [CrossRef]
- Sardelis, M.; Turell, M.J.; Dohm, D.J.; O’Guinn, M.L. Vector Competence of Selected North American Culex and Coquillettidia Mosquitoes for West Nile Virus. Emerg. Infect. Dis. 2001, 7, 1018–1022. [Google Scholar] [CrossRef] [PubMed]
- Bingham, A.M.; Burkett-Cadena, N.D.; Hassan, H.K.; Unnasch, T.R. Vector Competence and Capacity of Culex erraticus (Diptera: Culicidae) for Eastern Equine Encephalitis Virus in the Southeastern United States. J. Med. Entomol. 2016, 53, 473–476. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giordano, B.V.; Allen, B.T.; Wishard, R.; Xue, R.-D.; Campbell, L.P. Light Trap Collections of Mosquitoes (Diptera: Culicidae) Using Dry Ice and Octenol Attractants in Adjacent Mosquito Control Programs. Fla. Entomol. 2021, 103, 499–504. [Google Scholar] [CrossRef]
- Pratt, H.D.; Seabrook, E.L. The Occurrence of Culex iolambdis Dyar in Florida and Puerto Rico, with a Description of the Larva (Diptera, Culicidae). Proc. Entomol. Soc. Wash. 1952, 54, 3–27. [Google Scholar]
- Adames, A.J. Mosquito Studies (Diptera, Culicidae). XXIV. A Revision of the Crabhole Mosquitoes of the Genus Deinocerites. Contrib. Am. Entomol. Inst. 1971, 7, 1–154. [Google Scholar]
- O’Meara, G.F.; Cutwa, M.M.; Evans, L.F. Bromeliad-Inhabiting Mosquitoes in South Florida: Native and Exotic Plants Differ in Species Composition. J. Vector Ecol. 2003, 28, 37–46. [Google Scholar]
- Day, J.F.; Tabachnick, W.J.; Smartt, C.T. Factors That Influence the Transmission of West Nile Virus in Florida. J. Med. Entomol. 2015, 52, 743–754. [Google Scholar] [CrossRef]
- Florida Fish and Wildlife. Conservation Commission Florida Land Cover Classification System. 2018. Available online: https://myfwc.com/research/gis/applications/articles/fl-land-cover-classification/ (accessed on 5 October 2021).
- Darsie, R.F.; Morris, C.D. Keys to the Adult Females and Fourth Instar Larvae of the Mosquitoes of Florida (Diptera, Culicidae). Tech. Bull. Fla. Mosq. Control. Assoc. 2003, 1, 1–159. [Google Scholar]
- Darsie, R.F.; Ward, R.A. Identification and Geographical Distribution of the Mosquitoes of North America, North of Mexico, 2nd ed.; University Press of Florida: Gainesville, FL, USA, 2005. [Google Scholar]
- Burkett-Cadena, N.D. Mosquitoes of the Southeastern United States; University of Alabama Press: Tuscaloosa, AL, USA, 2013. [Google Scholar]
- Condotta, S.A.; Hunter, F.F.; Bidochka, M.J. West Nile Virus Infection Rates in Pooled and Individual Mosquito Samples. Vector-Borne Zoonotic Dis. 2004, 4, 198–203. [Google Scholar] [CrossRef] [PubMed]
- Lanciotti, R.S.; Kerst, A.J.; Nasci, R.S.; Godsey, M.S.; Mitchell, C.J.; Savage, H.M.; Komar, N.; Panella, N.A.; Allen, B.C.; Volpe, K.E.; et al. Rapid Detection of West Nile Virus from Human Clinical Specimens, Field-Collected Mosquitoes, and Avian Samples by a TaqMan Reverse Transcriptase-PCR Assay. J. Clin. Microbiol. 2000, 38, 4066–4071. [Google Scholar] [CrossRef] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020; Available online: https://www.R-project.org/ (accessed on 2 December 2021).
- Chao, A.; Gotelli, N.J.; Sander, E.L.; Ma, K.H.; Colwell, R.K.; Elllison, A.M. Rarefaction and Extrapolation with Hill Numbers: A Framework for Sampling and Estimation in Species Diversity Studies. Ecol. Monogr. 2014, 84, 45–67. [Google Scholar] [CrossRef] [Green Version]
- Hsieh, T.C.; Ma, K.H.; Chao, A. INEXT: INterpolation and EXTrapolation for Species Diversity. 2020. Available online: http://chao.stat.nthu.edu.tw/wordpress/software-download/ (accessed on 10 August 2021).
- Oksanen, J.; Blanchet, G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; et al. Vegan: Community Ecology Package. R Package Version 2.5-7. 2020. Available online: https://CRAN.R-project.org/package=vegan (accessed on 5 October 2021).
- Brooks, M.E.; Kristensen, K.; van Benthem, K.J.; Magnusson, A.; Berg, C.W.; Nielsen, A.; Skaug, H.J.; Maechler, M.; Bolker, B.M. GlmmTMB Balances Speed and Flexibility among Packages for Zero-Inflated Generalized Linear Mixed Modeling. R J. 2017, 9, 378–400. [Google Scholar] [CrossRef] [Green Version]
- Hardin, J.W.; Hilbe, J.M. Generalized Linear Models and Extensions, 3rd ed.; StatCorp LP: College Station, TX, USA, 2012. [Google Scholar]
- Wickham, H. Ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016. [Google Scholar]
- Rund, S.S.C.; Braak, K.; Cator, L.; Copas, K.; Emrich, S.J.; Giraldo-Calderón, G.I.; Johansson, M.A.; Heydari, N.; Hobern, D.; Kelly, S.A.; et al. MIReAD, a minimum information standard for reporting arthropod abundance data. Sci. Data 2019, 6, 40. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Height (m) | Genus | |||||
|---|---|---|---|---|---|---|
| Aedes | Anopheles | Culex | Deinocerites | Mansonia | Wyeomyia | |
| 1.5 | 324 | 4193 | 282 | 16 | 8 | 10 |
| 5.0 | 47 | 4027 | 42 | 5 | 4 | 2 |
| 8.2 | 42 | 3595 | 112 | 10 | 15 | 2 |
| Variable | 1.5 m | 5.0 m | 8.7 m | SP | MH | HH | Site 1 (SP) | Site 2 (HH) | Site 3 (MH) | Site 4 (HH) | Site 5 (MH) |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Abundance | 4833 | 4127 | 3776 | 1803 | 4913 | 6020 | 1803 | 2643 | 2428 | 3377 | 2485 |
| Richness | 18 | 13 | 11 | 12 | 14 | 16 | 12 | 13 | 13 | 11 | 12 |
| No. Singletons | 4 | 4 | 1 | 4 | 3 | 3 | 4 | 3 | 2 | 3 | 3 |
| Species Density | 25.998± 11.660 | 20.998 ± 11.659 | 11.250 ± 0.729 | 19.996 ± 11.656 | 16.999 ± 4.533 | 20.499 ± 7.193 | 19.996 ± 11.656 | 15.999 ± 4.524 | 14.000 ± 2.283 | 12.000 ± 1.870 | 14.999 ± 4.515 |
| Shannon Diversity | 1.835 ± 0.032 | 1.215 ± 0.017 | 1.328 ± 0.024 | 2.103 ± 0.050 | 1.311 ± 0.018 | 1.429 ± 0.022 | 2.103 ± 0.050 | 1.537 ± 0.034 | 1.382 ± 0.030 | 1.298 ± 0.020 | 1.239 ± 0.023 |
| Simpson Diversity | 1.348 ± 0.015 | 1.064 ± 0.006 | 1.113 ± 0.009 | 1.587 ± 0.033 | 1.095 ± 0.007 | 1.147 ± 0.007 | 1.587 ± 0.033 | 1.193 ± 0.013 | 1.120 ± 0.011 | 1.112 ± 0.009 | 1.070 ± 0.008 |
| Variable | R2 | F (df) | p-Value | Post Hoc 1 |
|---|---|---|---|---|
| Height | 0.142 | 11.89 (2) | <0.001 | s:l padj = 0.003 s:m padj = 0.004 |
| Landscape | 0.038 | 3.13 (2) | 0.007 | ns |
| Site | 0.078 | 6.46 (4) | <0.001 | 2:5 padj = 0.020 2:4 padj = 0.027 |
| Sampling Date | 0.416 | 6.29 (11) | <0.001 | ns |
| Height:Landscape | 0.082 | 3.42 (4) | <0.001 | |
| Height:Site | 0.028 | 2.37 (8) | 0.032 | |
| Residual | 0.216 | (36) | ||
| Total | 1.000 | (59) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Giordano, B.V.; Cruz, A.; Pérez-Ramos, D.W.; Ramos, M.M.; Tavares, Y.; Caragata, E.P. Mosquito Communities Vary across Landscape and Vertical Strata in Indian River County, Florida. Pathogens 2021, 10, 1575. https://doi.org/10.3390/pathogens10121575
Giordano BV, Cruz A, Pérez-Ramos DW, Ramos MM, Tavares Y, Caragata EP. Mosquito Communities Vary across Landscape and Vertical Strata in Indian River County, Florida. Pathogens. 2021; 10(12):1575. https://doi.org/10.3390/pathogens10121575
Chicago/Turabian StyleGiordano, Bryan V., Anthony Cruz, Daniel W. Pérez-Ramos, Martina M. Ramos, Yasmin Tavares, and Eric P. Caragata. 2021. "Mosquito Communities Vary across Landscape and Vertical Strata in Indian River County, Florida" Pathogens 10, no. 12: 1575. https://doi.org/10.3390/pathogens10121575