
Association of Fungi and Archaea of the Gut Microbiota with Crohn’s Disease in Pediatric Patients—Pilot Study

 , , and
, , and 
Abstract
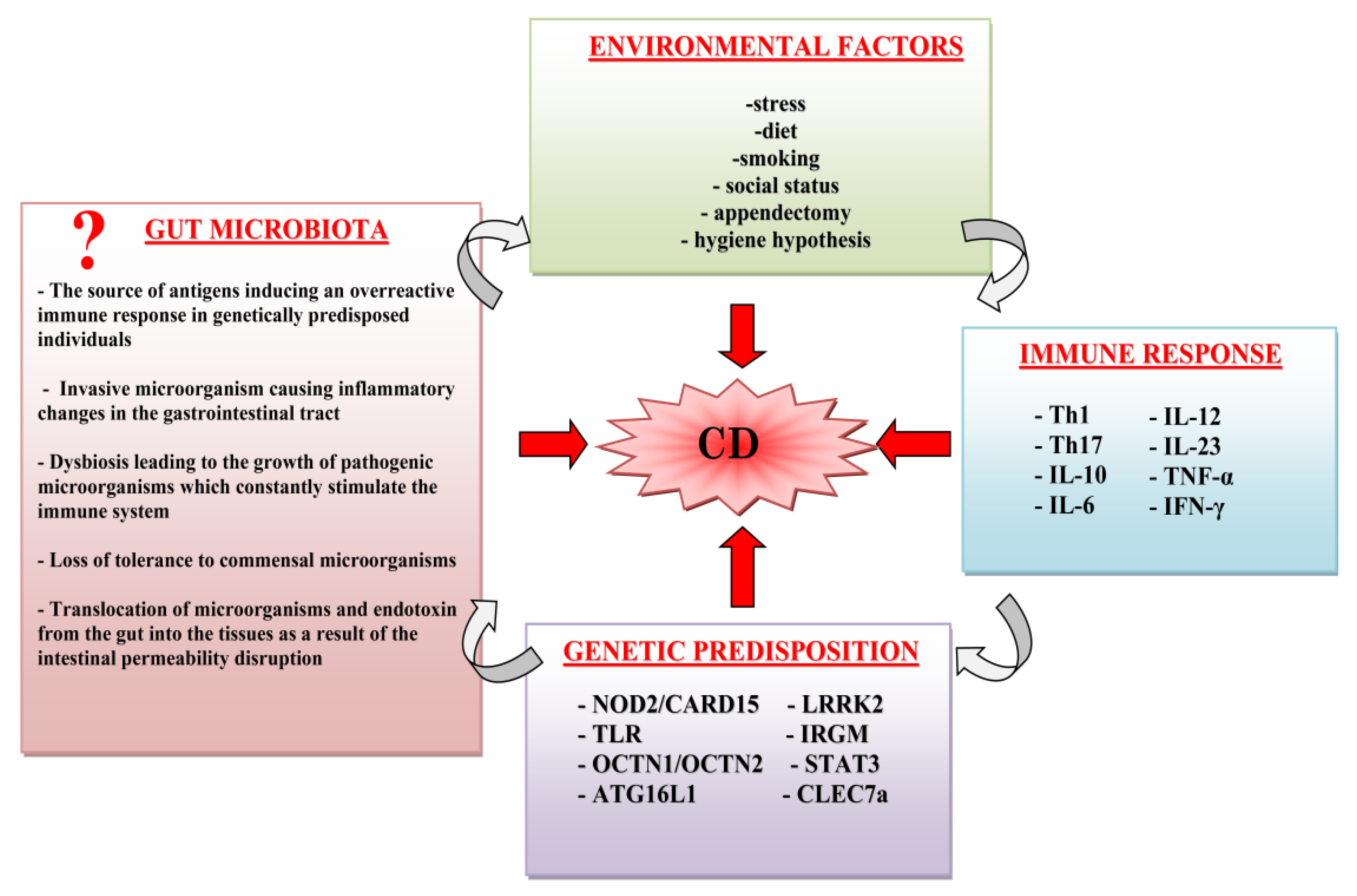
:1. Introduction
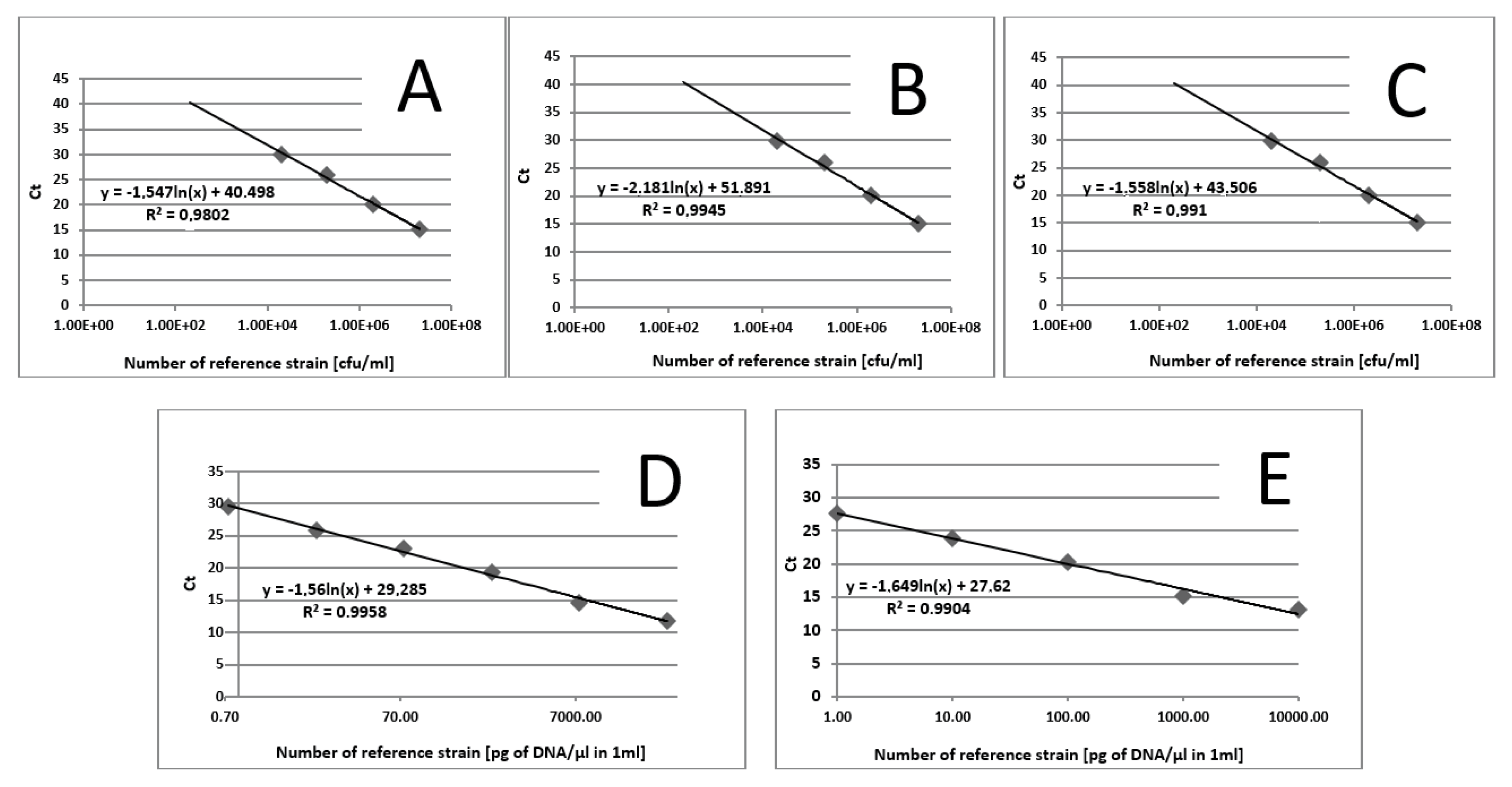
2. Results
2.1. Characteristics of the Subjects
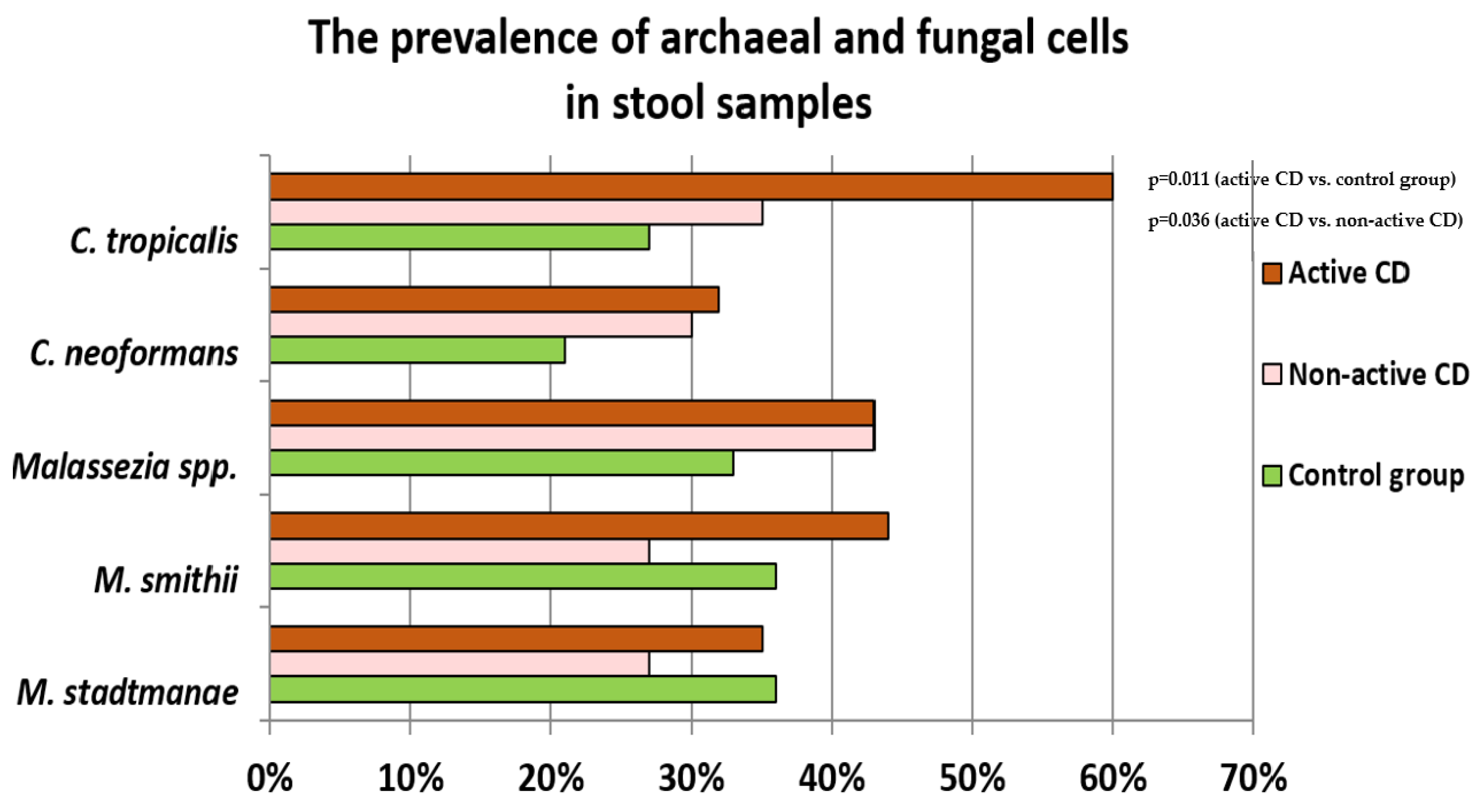
2.2. Prevalence of Fungi and Archaea in Stool Samples of Controls and Patients with CD
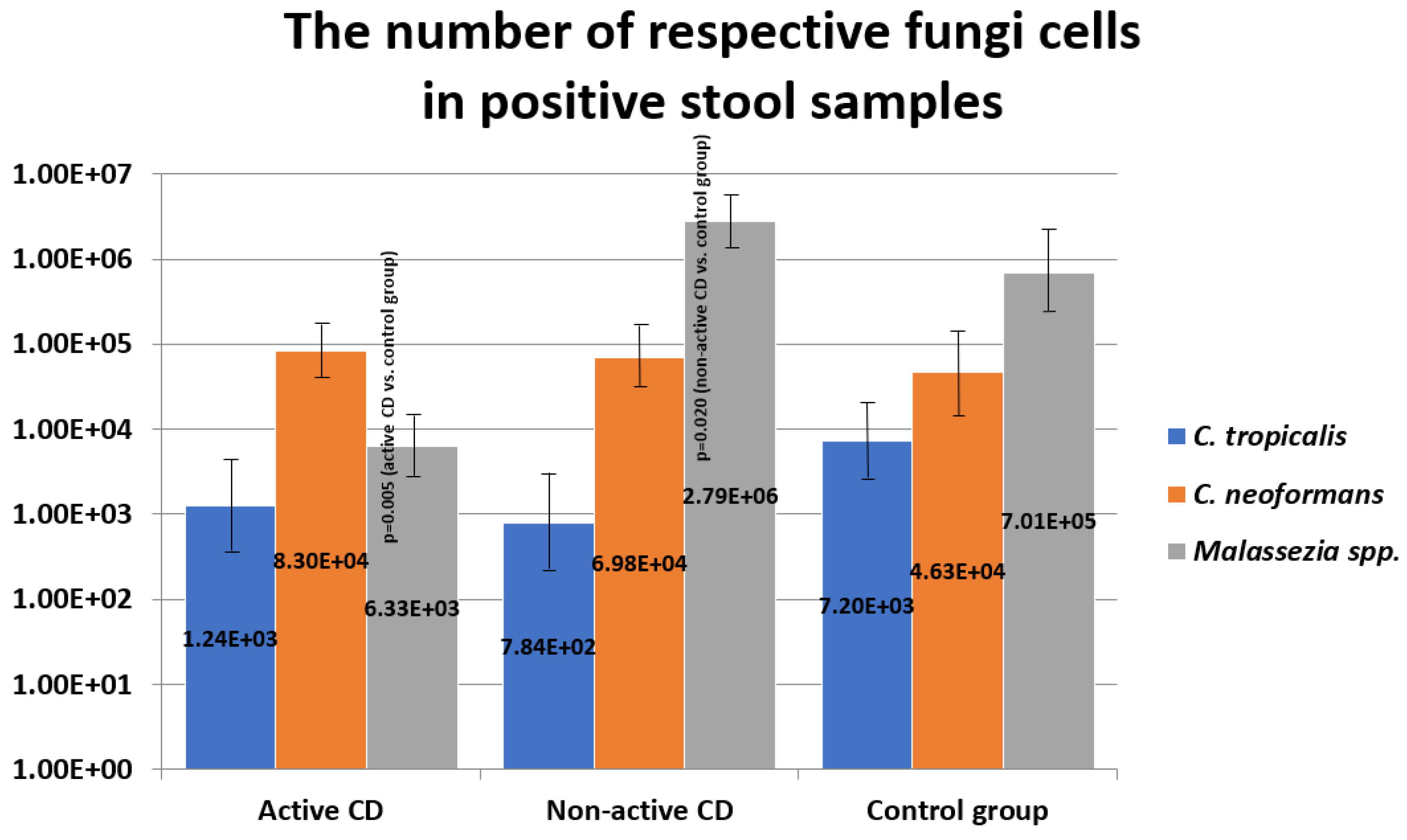
2.3. The Number of Fungi Cells in Stool Samples
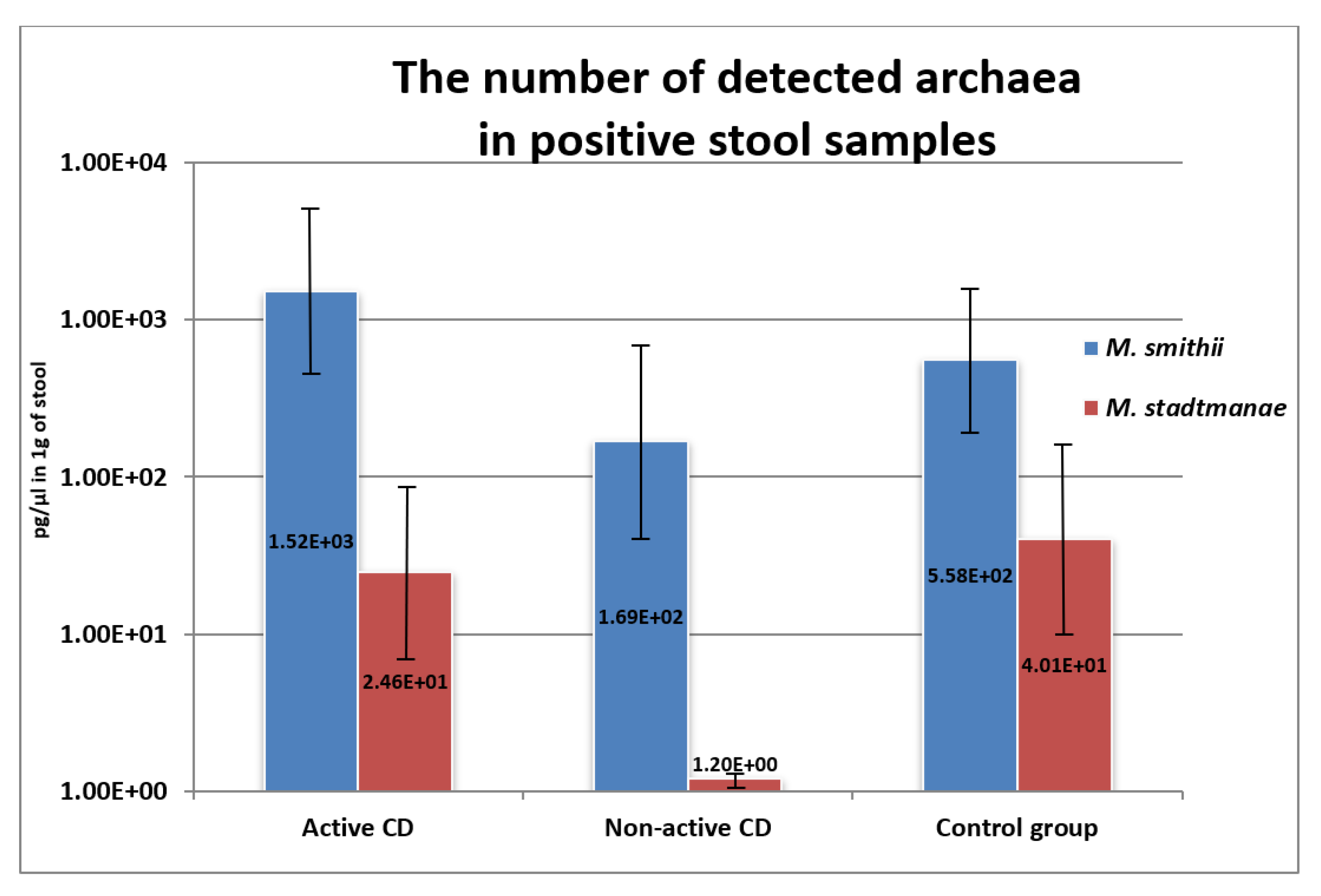
2.4. Methanogen DNA Concentration in Stool Samples
3. Discussion
4. Materials and Methods
4.1. Patients
4.2. DNA Extraction from the Stool Samples
4.3. Quantitative Real-Time PCR (qPCR):
4.4. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Khanna, S.; Raffals, L.E. The Microbiome in Crohn’s Disease: Role in Pathogenesis and Role of Microbiome Replacement Therapies. Gastroenterol. Clin. N. Am. 2017, 46, 481–492. [Google Scholar] [CrossRef] [PubMed]
- Kowalska-Duplaga, K.; Gosiewski, T.; Kapusta, P.; Sroka-Oleksiak, A.; Wędrychowicz, A.; Pieczarkowski, S.; Ludwig-Słomczyńska, A.H.; Wołkow, P.P.; Fyderek, K. Differences in the intestinal microbiome of healthy children and patients with newly diagnosed Crohn’s disease. Sci. Rep. 2019, 9, 18880. [Google Scholar] [CrossRef] [Green Version]
- Salamon, D.; Gosiewski, T.; Krawczyk, A.; Sroka-Oleksiak, A.; Duplaga, M.; Fyderek, K.; Kowalska-Duplaga, K. Quantitative changes in selected bacteria in the stool during the treatment of Crohn’s disease. Adv. Med. Sci. 2020, 65, 348–353. [Google Scholar] [CrossRef]
- Matricon, J.; Barnich, N.; Ardid, D. Immunopathogenesis of Inflammatory Bowel Disease. Self Nonself 2010, 1, 299–309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sartor, R.B. Therapeutic manipulation of the enteric microflora in inflammatory bowel disease: Antibiotics, probiotics and prebiotics. Gastroenterology 2004, 126, 1620–1633. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Radwan, P.; Radwan-Kwiatek, K.; Skrzydło-Radomańska, B. The role of enteric microflora in inflammatory bowel disease. Gastroenterol. Rev. 2009, 4, 1–6. [Google Scholar]
- Wang, A.Y.; Popov, J.; Pai, N. Fecal microbial transplant for the treatment of pediatric inflammatory bowel disease. Word J. Gastroenterol. 2016, 22, 10304–10315. [Google Scholar] [CrossRef]
- Rabizadeh, S.; Rhee, K.J.; Wu, S.; Huso, D.; Gan, C.M.; Golub, J.E.; Wu, X.; Zhang, M.; Sears, C.L. Enterotoxigenic Bacteroides fragilis: A potential instigator of colitis. Inflamm. Bowel Dis. 2007, 13, 1475–1483. [Google Scholar] [CrossRef] [Green Version]
- McNees, A.L.; Markesich, D.; Zayyani, N.R.; Graham, D.Y. Mycobacterium paratuberculosis as a cause of Crohn’s disease. Expert. Rev. Gastroenterol. Hepatol. 2015, 9, 1523–1534. [Google Scholar] [CrossRef]
- Jeyanathan, M.; Boutros-Tadros, O.; Radhi, J.; Semret, M.; Bitton, A.; Behr, M.A. Visualization of Mycobacterium avium in Crohn’s tissue by oil-immersion microscopy. Microbes Infect. 2007, 9, 1567–1573. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Man, S.M.; Day, A.S.; Leach, S.T.; Lemberg, D.A.; Dutt, S.; Stormon, M.; Otley, A.; O’Loughlin, E.V.; Magoffin, A.; et al. Detection and isolation of Campylobacter species other than C. jejuni from children with Crohn’s disease. J. Clin. Microbiol. 2009, 47, 453–455. [Google Scholar] [CrossRef] [Green Version]
- Kaakoush, N.O.; Mitchell, H.M. Campylobacter concisus—A new player in intestinal disease. Front. Cell. Infect. Microbiol. 2012, 2, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miquel, S.; Peyretaillade, E.; Claret, L.; de Vallée, A.; Dossat, C.; Vacherie, B.; Zineb, E.; Segurens, B.; Barbe, V.; Sauvanet, P.; et al. Complete genome sequence of Crohn’s disease-associated adherent-invasive E. coli strain LF82. PLoS ONE 2010, 5, e12714. [Google Scholar] [CrossRef]
- Martinez-Medina, M.; Aldeguer, X.; Lopez-Siles, M.; González-Huix, F.; López-Oliu, C.; Dahbi, G.; Blanco, J.E.; Blanco, J.; Garcia-Gil, L.J.; Darfeuille-Michaud, A. Molecular diversity of Escherichia coli in the human gut: New ecological evidence supporting the role of adherent-invasive E. coli (AIEC) in Crohn’s disease. Inflamm. Bowel Dis. 2009, 15, 872–882. [Google Scholar] [CrossRef]
- Underhill, D.M.; Iliev, I.D. The mycobiota: Interactions between commensal fungi and the host immune system. Nat. Rev. Immunol. 2014, 14, 405–416. [Google Scholar] [CrossRef]
- Joossens, S.; Reinisch, W.; Vermeire, S.; Sendid, B.; Poulain, D.; Peeters, M.; Geboes, K.; Bossuyt, X.; Vandewalle, P.; Oberhuber, G.; et al. The value of serologic markers in indeterminate colitis: A prospective follow-up study. Gastroenterology 2002, 122, 1242–1247. [Google Scholar] [CrossRef] [Green Version]
- Reese, G.E.; Constantinides, V.A.; Simillis, C.; Darzi, A.W.; Orchard, T.R.; Fazio, V.W.; Tekkis, P.P. Diagnostic precision of anti-Saccharomyces cerevisiae antibodies and perinuclear antineutrophil cytoplasmic antibodies in inflammatory bowel disease. Am. J. Gastroenterol. 2006, 101, 2410–2422. [Google Scholar] [CrossRef]
- Hoarau, G.; Mukherjee, K.; Gower-Rousseau, C.; Hager, C.; Chandra, J.; Retuerto, M.A.; Neut, C.; Vermeire, S.; Clemente, J.; Colombel, J.F.; et al. Bacteriome and Mycobiome Interactions Underscore Microbial Dysbiosis in Familial Crohn’s Disease. Mbio 2016, 7, e01250-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iliev, I.D.; Funari, V.A.; Taylor, K.D.; Nguyen, Q.; Reyes, C.N.; Strom, S.P.; Brown, J.; Becker, C.A.; Fleshner, P.R.; Dubinsky, M.; et al. Interactions between commensal fungi and the C-type lectin receptor Dectin-1 influence colitis. Science 2012, 336, 1314–1317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zwolinska-Wcislo, M.; Brzozowski, T.; Budak, A.; Kwiecień, S.; Sliwowski, Z.; Drozdowicz, D.; Trojanowska, D.; Rudnicka-Sosin, L.; Mach, T.; Konturek, S.J.; et al. Effect of Candida colonization on human ulcerative colitis and the healing of inflammatory changes of the colon in the experimental model of colitis ulcerosa. J. Physiol. Pharmacol. 2009, 60, 107–118. [Google Scholar]
- Krawczyk, A.; Sroka-Oleksiak, A.; Kowalska-Duplaga, K.; Fyderek, K.; Gosiewski, T.; Salamon, D. Impact of biological treatment on intestinal microbiome in children with Crohn’s disease. World Sci. News 2018, 104, 252–263. [Google Scholar]
- Limon, J.J.; Tang, J.; Li, D.; Wolf, A.J.; Michelsen, K.S.; Funari, V.; Gargus, M.; Nguyen, C.; Sharma, P.; Maymi, V.I.; et al. Malassezia Is Associated with Crohn’s Disease and Exacerbates Colitis in Mouse Models. Cell Host Microbe 2019, 25, 377–388. [Google Scholar] [CrossRef] [Green Version]
- Liguori, G.; Lamas, B.; Richard, M.L.; Brandi, G.; da Costa, G.; Hoffmann, T.W.; Di Simone, M.P.; Calabrese, C.; Poggioli, G.; Langella, P.; et al. Fungal Dysbiosis in Mucosa-associated Microbiota of Crohn’s Disease Patients. J. Crohn’s Colitis 2016, 10, 296–305. [Google Scholar] [CrossRef]
- Li, Q.; Wang, C.; Tang, C.; He, Q.; Li, N.; Li, J. Dysbiosis of gut fungal microbiota is associated with mucosal inflammation in Crohn’s disease. J. Clin. Gastroenterol. 2014, 48, 513–523. [Google Scholar] [CrossRef] [Green Version]
- Kowalska-Duplaga, K.; Krawczyk, A.; Sroka-Oleksiak, A.; Salamon, D.; Wędrychowicz, A.; Fyderek, K.; Gosiewski, T. Dependence of Colonization of the Large Intestine by Candida on the Treatment of Crohn’s Disease. Pol. J. Microbiol. 2019, 68, 121–126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lewis, J.D.; Chen, E.Z.; Baldassano, R.N.; Otley, A.R.; Griffiths, A.M.; Lee, D.; Bittinger, K.; Bailey, A.; Friedman, E.S.; Hoffmann, C.; et al. Inflammation, Antibiotics, and Diet as Environmental Stressors of the Gut Microbiome in Pediatric Crohn’s Disease. Cell Host Microbe 2015, 18, 489–500. [Google Scholar] [CrossRef] [Green Version]
- Stamatiades, G.A.; Ioannou, P.; Petrikkos, G.; Tsioutis, C. Fungal infections in patients with inflammatory bowel disease: A systematic review. Mycoses 2018, 61, 366–376. [Google Scholar] [CrossRef]
- Lam, S.; Zuo, T.; Ho, M.; Chan, F.K.L.; Chan, P.K.S.; Ng, S.C. Review article: Fungal alterations in inflammatory bowel diseases. Aliment Pharmacol. Ther. 2019, 50, 1159–1171. [Google Scholar] [CrossRef] [PubMed]
- Gevers, D.; Kugathasan, S.; Denson, L.A.; Vázquez-Baeza, Y.; Van Treuren, W.; Ren, B.; Schwager, E.; Knights, D.; Song, S.J.; Yassour, M.; et al. The treatment-naive microbiome in new-onset Crohn’s disease. Cell Host Microbe 2014, 15, 382–392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scanlan, P.D.; Shanahan, F.; Marchesi, J.R. Human methanogen diversity and incidence in healthy and diseased colonic groups using mcrA gene analysis. BMC Microbiol. 2008, 8, 79. [Google Scholar] [CrossRef] [Green Version]
- Blais-Lecours, P.; Marsolais, D.; Berberi, M.; Hache, C.; Bourdages, R.; Duchaine, C. Increased Prevalence of Methanosphaera stadtmanae in Inflammatory Bowel Diseases. PLoS ONE 2014, 9, e87734. [Google Scholar] [CrossRef] [PubMed]
- Peled, Y.; Gilat, T.; Liberman, E.; Bujanover, Y.J. The development of methane production in childhood and adolescence. Pediatr. Gastroenterol. Nutr. 1985, 4, 575–579. [Google Scholar] [CrossRef] [PubMed]
- Chehoud, C.; Albenberg, L.G.; Judge, C.; Hoffmann, C.; Grunberg, S.; Bittinger, K.; Baldassano, R.N.; Lewis, J.D.; Bushman, F.D.; Wu, G.D. Fungal Signature in the Gut Microbiota of Pediatric Patients With Inflammatory Bowel Disease. Inflamm. Bowel Dis. 2015, 21, 1948–1956. [Google Scholar] [CrossRef]
- Levine, A.; Koletzko, S.; Turner, D.; Escher, J.C.; Cucchiara, S.; de Ridder, L.; Kolho, K.L.; Veres, G.; Russell, R.K.; Paerregaard, A.; et al. European Society of Pediatric Gastroenterology, Hepatology, and Nutrition. J. Pediatr. Gastroenterol. Nutr. 2014, 58, 795–806. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brinkman, N.E.; Haugland, R.A.; Wymer, L.J.; Byappanahalli, M.; Whitman, R.L.; Vesper, S.J. Evaluation of a rapid, quantitative real-time PCR method for enumeration of pathogenic Candida cells in water. Appl. Environ. Microbiol. 2003, 69, 1775–1782. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Veron, V.; Simon, S.; Blanchet, D.; Aznar, C. Real-time polymerase chain reaction detection of Cryptococcus neoformans and Cryptococcus gattii in human samples. Diagn. Microbiol. Infect. Dis. 2009, 65, 69–72. [Google Scholar] [CrossRef]
- Xie, Z.; Ran, Y.; Zhang, H.; Zhang, M.; Wan, H.; Li, C. An analysis of the Malassezia species distribution in the skin of patients with pityriasis versicolor in Chengdu, China. Sci. World J. 2014, 182596. [Google Scholar]
- Dridi, B.; Henry, M.; El Khéchine, A.; Raoult, D.; Drancourt, M. High prevalence of Methanobrevibacter smithii and Methanosphaera stadtmanae detected in the human gut using an improved DNA detection protocol. PLoS ONE 2009, 4, e7063. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer and Probe | Sequence (5′ to 3′) | Thermal Cycling Programme | References |
|---|---|---|---|
| C. tropicalis Forward primer Reverse primerProbe | GCGGTAGGAGAATTGCGTT TCATTATGCCAACATCCTAGGTTTA 6-FAM-CGCAGTCCTCAGTCTAGGCTGGCAG-BHQ-1 |  | [35] |
| C. neoformans Forward primer Reverse primerProbe | GCCGCGACCTGCAAAG GGTAATCACCTTCCCACTAACACAT 6-FAM-ACGTCGGCTCGCC-BHQ-1 |  | [36] |
| Malassezia spp. Forward primer Reverse primerProbe | GTAGACTCCATCTAAAGCTAAAT CTTTTAACTCTCTTTCCAAAGT 6-FAM-CCCTCACGGTACTTGTTCGCT-BHQ-1 |  | [37] |
| M. smithii Forward primer Reverse primerProbe | CCGGGTATCTAATCCGGTTC CTCCCAGGGTAGAGGTGAAA 6-FAM-CCGTCAGAATCGTTCCAGTCAG-BHQ-1 |  | [38] |
| M. stadtmanae Forward primer Reverse primerProbe | AGGAGCGACAGCAGAATGAT CAGGACGCTTCACAGTACGA 6-FAM-TGAGAGGAGGTGCATGGCCG-BHQ-1 |  | [38] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Krawczyk, A.; Salamon, D.; Kowalska-Duplaga, K.; Bogiel, T.; Gosiewski, T. Association of Fungi and Archaea of the Gut Microbiota with Crohn’s Disease in Pediatric Patients—Pilot Study. Pathogens 2021, 10, 1119. https://doi.org/10.3390/pathogens10091119
Krawczyk A, Salamon D, Kowalska-Duplaga K, Bogiel T, Gosiewski T. Association of Fungi and Archaea of the Gut Microbiota with Crohn’s Disease in Pediatric Patients—Pilot Study. Pathogens. 2021; 10(9):1119. https://doi.org/10.3390/pathogens10091119
Chicago/Turabian StyleKrawczyk, Agnieszka, Dominika Salamon, Kinga Kowalska-Duplaga, Tomasz Bogiel, and Tomasz Gosiewski. 2021. "Association of Fungi and Archaea of the Gut Microbiota with Crohn’s Disease in Pediatric Patients—Pilot Study" Pathogens 10, no. 9: 1119. https://doi.org/10.3390/pathogens10091119