
Molecular Detection and Genotyping of Coxiella-Like Endosymbionts in Ticks Collected from Animals and Vegetation in Zambia

 , ,
, , 
 ,
,  ,
,
Abstract
:1. Introduction
2. Results
2.1. Morphological Tick Identification
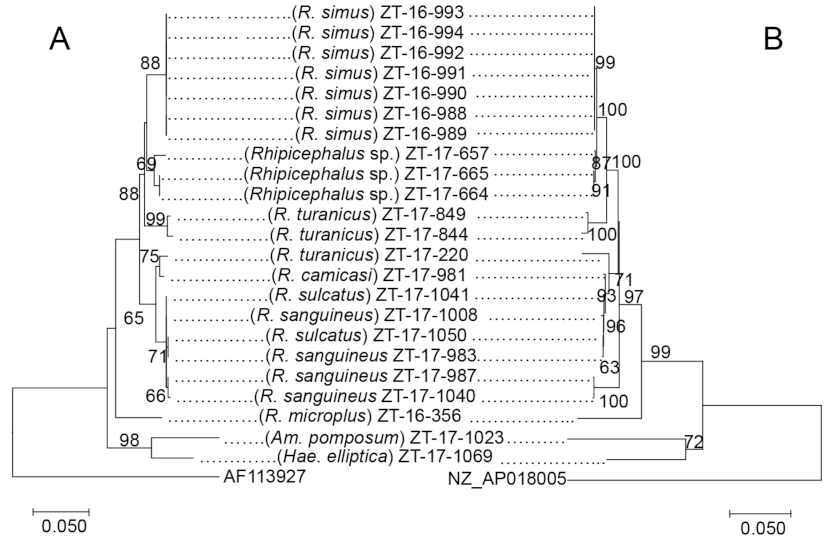
2.2. Molecular Identification and Phylogenetic Analysis of Ticks Based on the Mitochondrial 16S rRNA Gene Sequences
2.3. Screening of CLE
2.4. Multi-Locus Sequence Typing of groEL, dnaK, rpoB, 16S rRNA, and 23S rRNA Genes of CLE
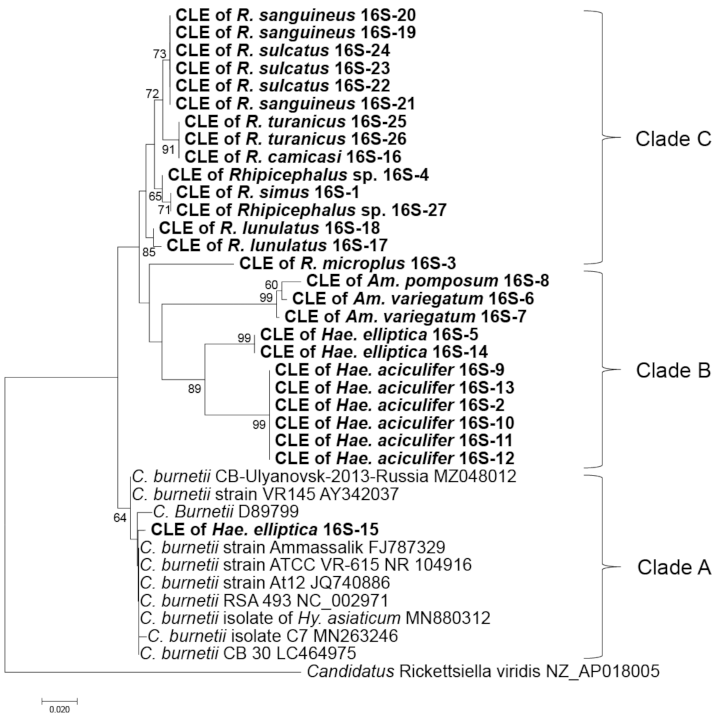
2.5. Comparison of Alleles of CLE and Their Phylogenetic Relationship with the Host Ticks
3. Discussion
4. Materials and Methods
4.1. Ethical Consideration
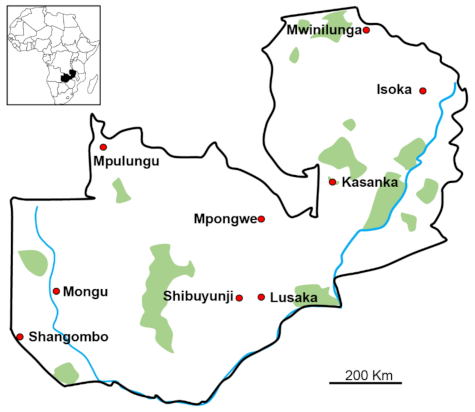
4.2. Tick Collection, Morphological Identification, and DNA Extraction
4.3. Molecular Tick Identification
4.4. Detection of CLE and Typing
4.5. Sequencing
4.6. Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Nicholson, W.L.; Sonenshine, D.E.; Noden, B.H.; Brown, R.N. Ticks (Ixodida). Med. Vet. Entomol. 2019, 603–672. [Google Scholar] [CrossRef]
- Nakao, R.; Abe, T.; Nijhof, A.M.; Yamamoto, S.; Jongejan, F.; Ikemura, T.; Sugimoto, C. A novel approach, based on BLSOMs (Batch Learning Self-Organizing Maps), to the microbiome analysis of ticks. ISME J. 2013, 7, 1003–1015. [Google Scholar] [CrossRef] [PubMed]
- Qiu, Y.; Nakao, R.; Ohnuma, A.; Kawamori, F.; Sugimoto, C. Microbial population analysis of the salivary glands of ticks; a possible strategy for the surveillance of bacterial pathogens. PLoS ONE 2014, 9, e0103961. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palomar, A.M.; Portillo, A.; Santibáñez, P.; Mazuelas, D.; Arizaga, J.; Crespo, A.; Gutiérrez, Ó.; Cuadrado, J.F.; Oteo, J.A. Crimean-Congo hemorrhagic fever virus in ticks from migratory birds, Morocco. Emerg. Infect. Dis. 2013, 19, 260–263. [Google Scholar] [CrossRef] [PubMed]
- Arraga-Alvarado, C.M.; Qurollo, B.A.; Parra, O.C.; Berrueta, M.A.; Hegarty, B.C.; Breitschwerdt, E.B. Case report: Molecular evidence of Anaplasma platys infection in two women from Venezuela. Am. J. Trop. Med. Hyg. 2014, 91, 1161–1165. [Google Scholar] [CrossRef]
- Maggi, R.G.; Mascarelli, P.E.; Havenga, L.N.; Naidoo, V.; Breitschwerdt, E.B. Coinfection with Anaplasma platys, Bartonella henselae and Candidatus Mycoplasma haematoparvum in a veterinarian. Parasite Vectors 2013, 6, 103. [Google Scholar] [CrossRef] [Green Version]
- Erik, M.F.; Vizzoni, V.F.; Balsemão-Pires, E.; Moerbeck, L.; Gazeta, G.S.; Piesman, J.; Voloch, C.M.; Soares, C.A.G. Coxiella symbionts are widespread into hard ticks. Parasitol. Res. 2016, 115, 4691–4699. [Google Scholar] [CrossRef]
- Bonnet, S.I.; Binetruy, F.; Hernández-Jarguín, A.M.; Duron, O. The tick microbiome: Why non-pathogenic microorganisms matter in tick biology and pathogen transmission. Front. Cell. Infect. Microbiol. 2017, 7. [Google Scholar] [CrossRef]
- Duron, O.; Binetruy, F.; Noël, V.; Cremaschi, J.; McCoy, K.D.; Arnathau, C.; Plantard, O.; Goolsby, J.; Pérez de León, A.A.; Heylen, D.J.A.; et al. Evolutionary changes in symbiont community structure in ticks. Mol. Ecol. 2017, 26, 2905–2921. [Google Scholar] [CrossRef] [Green Version]
- Rio, R.; Attardo, G.M.; Weiss, B.L. Grandeur Alliances: Symbiont Metabolic Integration and Obligate Arthropod Hematophagy. Trends Parasitol. 2016, 32, 739–749. [Google Scholar] [CrossRef] [Green Version]
- Hosokawa, T.; Koga, R.; Kikuchi, Y.; Meng, X.Y.; Fukatsu, T. Wolbachia as a bacteriocyte-associated nutritional mutualist. Proc. Natl. Acad. Sci. USA 2010, 107, 769–774. [Google Scholar] [CrossRef] [Green Version]
- Cirimotich, C.; Dong, Y.; Clayton, A.; Sandiford, S.; Souza-Neto, J.; Mulenga, M.; Dimopoulos, G. Natural Microbe-Mediated Refractoriness to Plasmodium Infection in Anopheles gambiae. Science 2011, 332, 855–858. [Google Scholar] [CrossRef] [Green Version]
- Moreira, L.A.; Iturbe-Ormaetxe, I.; Jeffery, J.A.; Lu, G.; Pyke, A.T.; Hedges, L.M.; Rocha, B.C.; Hall-Mendelin, S.; Day, A.; Riegler, M.; et al. A Wolbachia Symbiont in Aedes aegypti Limits Infection with Dengue, Chikungunya, and Plasmodium. Cell 2009, 139, 1268–1278. [Google Scholar] [CrossRef] [Green Version]
- Klyachko, O.; Stein, B.D.; Grindle, N.; Clay, K.; Fuqua, C. Localization and visualization of a Coxiella-type symbiont within the lone star tick, Amblyomma americanum. Appl. Environ. Microbiol. 2007, 73, 6584–6594. [Google Scholar] [CrossRef] [Green Version]
- Machado-Ferreira, E.; Dietrich, G.; Hojgaard, A.; Levin, M.; Piesman, J.; Zeidner, N.S.; Soares, C.A.G. Coxiella Symbionts in the Cayenne Tick Amblyomma cajennense. Microb. Ecol. 2011, 62, 134–142. [Google Scholar] [CrossRef]
- Díaz-S´anchez, S.; Estrada-Pena, A.; Cabezas-Cruz, A.; de la Fuente, J. Evolutionary insights into the tick hologenome. Trends Parasitol. 2019, 35, 725–737. [Google Scholar] [CrossRef]
- Narasimhan, S.; Fikrig, E. Tick microbiome: The force within. Trends Parasitol. 2015, 31, 315–323. [Google Scholar] [CrossRef] [Green Version]
- Noda, H.; Munderloh, U.G.; Kurtti, T.J. Endosymbionts of ticks and their relationship to Wolbachia spp. and tick-borne pathogens of humans and animals. Appl. Environ. Microbiol. 1997, 63, 3926–3932. [Google Scholar] [CrossRef] [Green Version]
- Duron, O.; Jourdain, E.; McCoy, K.D. Diversity and global distribution of the Coxiella intracellular bacterium in seabird ticks. Ticks Tick Borne Dis. 2014, 5, 557–563. [Google Scholar] [CrossRef]
- Duron, O.; Noël, V.; McCoy, K.D.; Bonazzi, M.; Sidi-Boumedine, K.; Morel, O.; Vavre, F.; Zenner, L.; Jourdain, E.; Durand, P.; et al. The Recent Evolution of a Maternally-Inherited Endosymbiont of Ticks Led to the Emergence of the Q Fever Pathogen, Coxiella burnetii. PLoS Pathog. 2015, 11, e1004892. [Google Scholar] [CrossRef] [Green Version]
- Smith, T.A.; Driscoll, T.; Gillespie, J.J.; Raghavan, R. A Coxiella-like endosymbiont is a potential vitamin source for the lone star tick. Genome Biol. Evol. 2015, 7, 831–838. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Almeida, A.P.; Marcili, A.; Leite, R.C.; Nieri-Bastos, F.A.; Domingues, L.N.; Martins, J.R.; Labruna, M.B. Coxiella symbiont in the tick Ornithodoros rostratus (Acari: Argasidae). Ticks Tick Borne Dis. 2012, 3, 203–206. [Google Scholar] [CrossRef] [PubMed]
- Duron, O.; Schneppat, U.E.; Berthomieu, A.; Goodman, S.M.; Droz, B.; Paupy, C.; Obame Nkoghe, J.; Rahola, N.; Tortosa, P. Origin, acquisition and diversification of heritable bacterial endosymbionts in louse flies and bat flies. Mol. Ecol. 2014, 23, 2105–2117. [Google Scholar] [CrossRef] [PubMed]
- Lalzar, I.; Harrus, S.; Mumcuoglu, K.Y.; Gottlieb, Y. Composition and seasonal variation of Rhipicephalus turanicus and Rhip-icephalus sanguineus bacterial communities. Appl. Environ. Microbiol. 2012, 78, 4110–4116. [Google Scholar] [CrossRef] [Green Version]
- Jasinskas, A.; Zhong, J.; Barbour, A.G. Highly prevalent Coxiella sp. bacterium in the tick vector Amblyomma americanum. Appl. Environ. Microbiol. 2007, 73, 334–336. [Google Scholar] [CrossRef] [Green Version]
- Clay, K.; Klyachko, O.; Grindle, N.; Civitello, D.; Oleske, D.; Fuqua, C. Microbial communities and interactions in the lone star tick, Amblyomma americanum. Mol. Ecol. 2008, 17, 4371–4381. [Google Scholar] [CrossRef]
- Walker, A., R.; Bouattor, J.L.; Camicas, A.; Estrada-Pena, I.G.; Horak, A.A.; Latiff, R.G.; Pegram, P.M. Ticks of Domestic Animals in Africa: A Guide to Identification of Species; Bioscience Reports: Edinburgh Scotland, UK, 2003; pp. 1–221. ISBN 0-9545173-0-X. [Google Scholar]
- Qiu, Y.; Hokkaido University, Sapporo, Hokkaido, Japan. Personal communication, 2018.
- Simuunza, M.; Weir, W.; Courcier, E.; Tait, A.; Shiels, B. Epidemiological analysis of tick-borne diseases in Zambia. Vet. Parasitol. 2011, 175, 331–342. [Google Scholar] [CrossRef]
- Ahmadu, B.; Lovelace, C.E.; Samui, K.L.; Mahan, S. Some observations on the seroprevalence of heartwater and tick infestation in Zambian goats. Onderstepoort J. Vet. Res. 2004, 71, 161–164. [Google Scholar] [CrossRef]
- Makala, L.H.; Mangani, P.; Fujisaki, K.; Nagasawa, H. The current status of major tick borne diseases in Zambia. Vet. Res. 2003, 34, 27–45. [Google Scholar] [CrossRef] [Green Version]
- De Castro, J.J.; James, A.D.; Minjauw, B.; Di Giulio, G.U.; Permin, A.; Pegram, R.G.; Chizyuka, H.G.B.; Sinyangwe, P. Long-term studies on the economic impact of ticks on Sanga cattle in Zambia. Exp. Appl. Acarol. 1997, 21, 3–19. [Google Scholar] [CrossRef]
- Qiu, Y.; Nakao, R.; Hang’ombe, B.M.; Sato, K.; Kajihara, M.; Kanchela, S.; Changula, K.; Eto, Y.; Ndebe, J.; Sasaki, M.; et al. Human Borreliosis Caused by a New World Relapsing Fever Borrelia–like Organism in the Old World. Clin. Infect. Dis. 2019, 69, 107–112. [Google Scholar] [CrossRef]
- Qiu, Y.; Kaneko, C.; Kajihara, M.; Saasa, N.; Simulundu, E.; Muleya, W.; Thu, M.J.; Hang’ombe, M.B.; Katakura, K.; Takada, A.; et al. Tick-borne haemoparasites and Anaplasmataceae in domestic dogs in Zambia. Ticks Tick Borne Dis. 2018, 9, 988–995. [Google Scholar] [CrossRef] [Green Version]
- Vlahakis, P.A.; Chitanga, S.; Simuunza, M.C.; Simulundu, E.; Qiu, Y.; Changula, K.; Chambaro, H.M.; Kajihara, M.; Nakao, R.; Takada, A.; et al. Molecular detection and characterization of zoonotic Anaplasma species in domestic dogs in Lusaka, Zambia. Ticks Tick Borne Dis. 2018, 9, 39–43. [Google Scholar] [CrossRef]
- Beati, L.; Patel, J.; Lucas-Williams, H.; Adakal, H.; Kanduma, E.G.; Tembo Mwase, E.; Krecek, R.; Mertins, J.W.; Alfred, J.T.; Kelly, S.; et al. Phylogeography and demographic history of Amblyomma variegatum (Fabricius) (Acari: Ixodidae), the tropical bont tick. Vector Borne Zoonotic Dis. 2012, 12, 514–525. [Google Scholar] [CrossRef] [Green Version]
- Dantas-Torres, F.; Maia, C.; Latrofa, M.S.; Annoscia, G.; Cardoso, L.; Otranto, D. Genetic characterization of Rhipicephalus sanguineus (sensu lato) ticks from dogs in Portugal. Parasite Vectors 2017, 10, 133. [Google Scholar] [CrossRef] [Green Version]
- Pegram, R.G.; Clifford, C.M.; Walker, J.B.; Keirans, J.E. Clarification of the Rhipicephalus sanguineus group (Acari, Ixodoidea, Ixodidae). I. R. sulcatus Neumann, 1908 and R. turanicus Pomerantsev, 1936. Syst. Parasitol. 1987, 10, 3–26. [Google Scholar] [CrossRef]
- Olivieri, E.; Kariuki, E.; Floriano, A.M.; Castelli, M.; Tafesse, Y.M.; Magoga, G.; Kumsa, B.; Montagna, M.; Sassera, D. Multi-country investigation of the diversity and associated microorganisms isolated from tick species from domestic animals, wildlife and vegetation in selected African countries. Exp. Appl. Acarol. 2021, 83, 427–448. [Google Scholar] [CrossRef]
- Oundo, J.W.; Villinger, J.; Jeneby, M.; Ong’amo, G.; Otiende, M.Y.; Makhulu, E.E.; Musa, A.A.; Ouso, D.O.; Wambua, L. Pathogens, endosymbionts, and blood-meal sources of host-seeking ticks in the fast-changing Maasai Mara wildlife ecosystem. PLoS ONE 2020, 5, e0228366. [Google Scholar] [CrossRef]
- Seo, M.G.X.; Lee, S.H.; Ouh, I.O.; Lee, G.H.; Goo, Y.K.; Kim, S.; Kwon, O.D.; Kwak, D. Molecular detection and genotyping of Coxiella-like endosymbionts in ticks that infest horses in South Korea. PLoS ONE 2016, 11, e0165784. [Google Scholar] [CrossRef]
- Gottlieb, Y.; Lalzar, I.; Klasson, L. Distinctive genome reduction rates revealed by genomic analyses of two Coxiella-like endosymbionts in ticks. Genome Biol. Evol. 2015, 7, 1779–1796. [Google Scholar] [CrossRef] [Green Version]
- Angelakis, E.; Thiberville, S.D.; Million, M.; Raoult, D. Sternoclavicular joint infection caused by Coxiella burnetii: A case report. J. Med. Case Rep. 2016, 10, 139. [Google Scholar] [CrossRef] [Green Version]
- Guimard, T.; Amrane, S.; Prudent, E.; El Karkouri, K.; Raoult, D.; Angelakis, E. Case Report: Scalp Eschar and Neck Lymphadenopathy Associated with Bacteremia due to Coxiella-Like Bacteria. Am. J. Trop. Med. Hyg. 2017, 97, 1319–1322. [Google Scholar] [CrossRef]
- Tsementzi, D.; Gordillo, C.J.; Mahagna, M.; Gottlieb, Y.; Konstantinidis, K.T. Comparison of closely related, uncultivated Coxiella tick endosymbiont population genomes reveals clues about the mechanisms of symbiosis. Environ. Microbiol. 2018, 20, 1751–1764. [Google Scholar] [CrossRef]
- Ben-Yosef, M.; Rot, A.; Mahagna, M.; Kapri, E.; Behar, A.; Gottlieb, Y. Coxiella-like endosymbiont of Rhipicephalus sanguineus is required for physiological processes during ontogeny. Front. Microbiol. 2020, 11, 493. [Google Scholar] [CrossRef]
- Wernegreen, J.J. Endosymbiosis. Curr. Biol. 2012, 22, 555–561. [Google Scholar] [CrossRef] [Green Version]
- Bennett, G.M.; Moran, N.A. Heritable symbiosis: The advantages and perils of an evolutionary rabbit hole. Proc. Natl. Acad. Sci. USA 2015, 112, 10169–10176. [Google Scholar] [CrossRef] [Green Version]
- Van Schaik, E.; Chen, C.; Mertens, K.; Weber, M.M.; Samuel, J.E. Molecular pathogenesis of the obligate intracellular bacterium Coxiella burnetii. Nat. Rev. Microbiol. 2013, 11, 561–573. [Google Scholar] [CrossRef] [Green Version]
- Seo, M.G.; Lee, S.H.; VanBik, D.; Ouh, I.O.; Yun, S.H.; Choi, E.; Park, Y.S.; Lee, S.E.; Kim, J.W.; Cho, G.J.; et al. Detection and genotyping of Coxiella burnetii and Coxiella-like bacteria in horses in South Korea. PLoS ONE 2016, 11, e0156710. [Google Scholar] [CrossRef]
- Nardi, T.; Olivieri, E.; Kariuki, E.; Sassera, D.; Castelli, M. Sequence of a Coxiella endosymbiont of the tick Amblyomma nuttalli suggests a pattern of convergent genome reduction in the Coxiella genus. Genome Biol. Evol. 2021, 13, 1. [Google Scholar] [CrossRef]
- Brenner, A.E.; Muñoz-Leal, S.; Sachan, M.; Labruna, M.B.; Raghavan, R. Coxiella burnetii and related tick endosymbionts evolved from pathogenic ancestors. Genome Biol. Evol. 2021. [Google Scholar] [CrossRef]
- Cafiso, A.; Sassera, D.; Romeo, C.; Serra, V.; Hervet, C.; Bandi, C.; Plantard, O.; Bazzocchi, C. Midichloria mitochondrii, endosymbiont of Ixodes ricinus: Evidence for the transmission to the vertebrate host during the tick blood meal. Ticks Tick Borne Dis. 2018. [Google Scholar] [CrossRef] [PubMed]
- Takano, A.; Toyomane, K.; Konnai, S.; Ohashi, K.; Nakao, M.; Ito, T.; Andoh, M.; Maeda, K.; Watarai, M.; Sato, K.; et al. Tick Surveillance for Relapsing Fever Spirochete Borrelia miyamotoi in Hokkaido, Japan. PLoS ONE 2014, 9, e104532. [Google Scholar] [CrossRef] [PubMed]
- Nava, S.; Estrada-Peña, A.; Petney, T.; Beati, L.; Labruna, M.B.; Szabó, M.P.; José, M.; Venzal, J.M.; Mastropaolo, M.; Mangold, A.J.; et al. The taxonomic status of Rhipicephalus sanguineus (Latreille, 1806). Vet. Parasitol. 2015, 208, 2–8. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.H.; Chen, F.; Chen, Y.Z.; Song, H.Q.; Lin, R.Q.; Zhou, D.H.; Zhu, X.Q. Complete mitochondrial genome sequence data provides genetic evidence that the brown dog tick Rhipicephalus sanguineus (Acari: Ixodidae) represents a species complex. Int. J. Biol. Sci. 2013, 9, 361–369. [Google Scholar] [CrossRef] [Green Version]
- McCann, K.M.; Grant, W.N.; Spratt, D.M.; Shannon, M.; Hedtke, S.M. Cryptic species diversity in ticks that transmit disease in Australia. Int. J. Parasitol. Parasite Wildl. 2019, 10, 125–131. [Google Scholar] [CrossRef]
- Qiu, Y.; Simuunza, M.; Kajihara, M.; Chambaro, M.; Harima, H.; Eto, Y.; Simulundu, E.; Squarre, D.; Torii, S.; Takada, A.; et al. Screening of tick-borne pathogens in argasid ticks in Zambia: Expansion of the geographic distribution of Rickettsia lusitaniae and Rickettsia hoogstraalii and detection of putative novel Anaplasma species. Ticks Tick Borne Dis. 2021, 12. [Google Scholar] [CrossRef]
- Chitanga, S.; Simulundu, E.; Simuunza, M.C.; Changula, K.; Qiu, Y.; Kajihara, M.; Nakao, R.; Syakalima, M.; Takada, A.; Mweene, A.S.; et al. First molecular detection and genetic characterization of Coxiella burnetii in Zambian dogs and rodents. Parasite Vectors 2018, 11, 40. [Google Scholar] [CrossRef]
- Duron, O.; Morel, O.; Nöel, V.; Buysse, M.; Binetruy, F.; Lancelot, R.; Loire, E.; M.´enard, C.; Bouchez, O.; Vavre, F.; et al. Tick-bacteria mutualism depends on B vitamin synthesis pathways. Curr. Biol. 2018, 28, 1896–1902. [Google Scholar] [CrossRef] [Green Version]
- Ushijima, Y.; Oliver, J.H.; Keirans, J.E.; Tsurumi, M.; Kawabata, H.; Watanabe, H.; Fukunaga, M. Mitochodrial Sequence Variation in Carios capensis (Neumann), a Parasite of Seabirds, Collected on Torishima Island in Japan. J. Parasitol. 2003, 89, 196–198. [Google Scholar] [CrossRef]
- Nakao, R.; Qiu, Y.; Igarashi, M.; Magona, J.W.; Zhoua, L.; Ito, K.; Sugimoto, C. High prevalence of spotted fever group rick-ettsiae in Amblyomma variegatum from Uganda and their identification using sizes of intergenic spacers. Ticks Tick Borne Dis. 2013, 4, 506–512. [Google Scholar] [CrossRef]
- Rozas, J.; Ferrer-Mata, A.; Sánchez-DelBarrio, J.C.; Guirao-Rico, S.; Librado, P.; Ramos-Onsins, S.; Sanchez-Gracia, A. DnaSP 6: DNA Sequence Polymorphism Analysis of Large Data Sets. Mol. Boil. Evol. 2017, 34, 3299–3302. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA 7: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Tick Species | Sampling Site | Host or Habitat | Number Tested (M; F) | Number Positive (M; F) | Positive Rate (%) | Year |
|---|---|---|---|---|---|---|
| Amblyomma pomposum | Mpulungu, Shangombo | dog, cattle | 2 (1; 1) | 2 (1; 1) | 100 | 2016, 2017 |
| Amblyomma variegatum | Mpulungu, Shangombo | cattle | 13 (7; 6) | 7 (3; 4) | 54 | 2016, 2017 |
| Argas walkarae | Isoka | vegetation | 10 (NA) | 0 | 0 | 2017 |
| Haemaphysalis aciculifer | Kasanka | unknown | 9 (5; 4) | 9 (5; 4) | 100 | 2017 |
| Haemaphysalis elliptica | Mpulungu, Shibuyunji | vegetation, unknown | 10 (6; 4) | 4 (2; 2) | 40 | 2017 |
| Hyalomma marginatum | Mongu, Shangombo | cattle | 6 (1; 5) | 0 | 0 | 2017 |
| Hyalomma marginatum rufipes | Shibuyunji | unknown | 1 (1; 0) | 0 | 0 | 2017 |
| Hyalomma truncatum | Shangombo, Shibuyunji | cattle, vegetation | 11 (4; 7) | 1 (0; 1) | 9 | 2016, 2017 |
| Ornithodoros faini | Lusaka | cave | 10 (4; 6) | 2 (1; 1) | 20 | 2017 |
| Rhipicephalus appendiculatus | Mpongwe, Mpulungu, Shangombo | cattle, vegetation | 10 (6; 4) | 4 (2; 2) | 40 | 2016, 2017 |
| Rhipicephalus camicasi | Mpongwe | goat | 1 (0; 1) | 1 (0; 1) | 100 | 2017 |
| Rhipicephalus decoloratus | Mpongwe | cattle | 10 (5; 5) | 4 (2; 2) | 40 | 2017 |
| Rhipicephalus evertsi evertsi | Mpongwe | cattle | 1(0; 1) | 1 (0; 1) | 100 | 2017 |
| Rhipicephalus geigyi | Mongu | cattle | 5 (0; 5) | 1 (0; 1) | 20 | 2017 |
| Rhipicephalus guilhoni | Mpulungu | vegetation | 2 (2; 0) | 2 (2; 0) | 100 | 2017 |
| Rhipicephalus lunulatus | Isoka, Shangombo | dog, goat | 12 (5; 7) | 6 (2; 4) | 50 | 2016, 2017 |
| Rhipicephalus microplus | Mpongwe | cattle | 9 (3; 6) | 3 (1; 2) | 33 | 2017 |
| Rhipicephalus muhsamae | Mpulungu | vegetation | 2 (2; 0) | 2 (2; 0) | 100 | 2017 |
| Rhipicephalus sanguineus | Mpulungu, Shangombo | dog | 13 (6; 7) | 7 (2; 5) | 54 | 2016, 2017 |
| Rhipicephalus simus | Mwinilunga | unknown | 8 (5; 3) | 8 (5; 3) | 100 | 2017 |
| Rhipicephalus sulcatus | Mpulungu | cattle | 6 (4; 2) | 4 (2; 2) | 67 | 2017 |
| Rhipicephalus turanicus | Mpongwe | vegetation | 10 (7; 3) | 4 (2; 2) | 40 | 2017 |
| Rhipicephalus spp. | Isoka | cattle | 14 (8; 6) | 8 (3; 5) | 57 | 2017 |
| Totals | 175 (82; 83) | 80 (38; 42) | 45.7 |
| Tick Species | CLE Target Gene | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| groEL | dnaK | rpoB | 16S rRNA | 23S rRNA | |||||||||||
| No. Tested | No. +ve | Allele Type | No. Tested | No. +ve | Allele Type | No. Tested | No. +ve | Allele Type | No. Tested | No. +ve | Allele Type | No. Tested | No. +ve | Allele Type | |
| Amblyomma pomposum | 2 | 2 | G2, G6 | 2 | 1 | D20 | 2 | 2 | R11, R12 | 2 | 1 | 16S-6 | 2 | 2 | 23S-3 |
| Amblyomma variegatum | 13 | 7 | G2 | 7 | 1 | D20 | 7 | 7 | R8, R11, R12, R19 | 7 | 2 | 16S-7, 16S-8 | 7 | 7 | 23S-3, 23S-14 |
| Haemaphysalis aciculifer | 9 | 9 | G5, G6 | 9 | 4 | D20, D21, D22 | 9 | 9 | R2, R18 | 9 | 9 | 16S-2, 16S-9, 16S-10, 16S-11, 16S-12 | 9 | 9 | 23S-5, 23S-6, 23S-15 |
| Haemaphysalis elliptica | 10 | 4 | G8, G11 | 4 | 1 | D2 | 4 | 4 | R7, R14, R17 | 4 | 4 | 16S-5, 16S-13, 16S-14 | 4 | 4 | 23S-7, 23S-12 |
| Hyalomma truncatum | 11 | 1 | G2 | 1 | - | - | 1 | 1 | R16 | 1 | - | - | 1 | 1 | 23S-16 |
| Ornithodoros faini | 10 | 2 | G21, G22 | 2 | - | - | 2 | - | - | 2 | - | - | 2 | - | - |
| Rhipicephalus appendiculatus | 13 | 4 | G14, G15 | 4 | 4 | D9, D18, D19 | 4 | 4 | R21, R24 | 4 | - | - | 4 | 4 | 23S-13, 23S-17, 23S-18 |
| Rhipicephalus camicasi | 1 | 1 | G20 | 1 | 1 | D16 | 1 | 1 | R22 | 1 | 1 | 16S-15 | 1 | 1 | 23S-4 |
| Rhipicephalus decoloratus | 10 | 4 | G4, G9 | 4 | 4 | D4, D10 | 4 | 4 | R6 | 4 | - | - | 4 | 4 | 23S-19 |
| Rhipicephalus evertsi evertsi | 1 | 1 | G18 | 1 | - | - | 1 | 1 | R21 | 1 | - | - | 1 | 1 | 23S-20 |
| Rhipicephalus geigyi | 5 | 1 | G17 | 1 | 1 | D11 | 1 | 1 | R6 | 1 | - | - | 1 | 1 | 23S-21 |
| Rhipicephalus guilhoni | 2 | 2 | G1, G8 | 2 | 1 | D7 | 2 | 2 | R15 | 2 | - | - | 2 | 2 | 23S-4 |
| Rhipicephalus lunulatus | 12 | 6 | G7 | 6 | 3 | D5, D6 | 6 | 6 | R4 | 6 | 2 | 16S-16, 16S-17 | 6 | 6 | 23S-9, 23S-22, 23S-23 |
| Rhipicephalus microplus | 10 | 4 | G9, G16 | 4 | 3 | D8 | 4 | 4 | R10 | 4 | 2 | 16S-3 | 4 | 4 | 23S-8 |
| Rhipicephalus muhsamae | 2 | 2 | G8 | 2 | 1 | D22 | 2 | 2 | R7 | 2 | - | - | 2 | 2 | 23S-12 |
| Rhipicephalus sanguineus | 13 | 7 | G1, G19 | 7 | 7 | D2, D13, D14, | 7 | 7 | R3, R20 | 7 | 3 | 16S-18, 16S-19, 16S-20 | 7 | 7 | 23S-1, 23S-24 |
| Rhipicephalus simus | 8 | 8 | G3 | 8 | 8 | D1 | 8 | 8 | R1 | 8 | 7 | 16S-1 | 8 | 8 | 23S-2 |
| Rhipicephalus sulcatus | 6 | 4 | G1 | 4 | 2 | D2 | 4 | 4 | R3 | 4 | 3 | 16S-21, 16S-22, 16S-23 | 4 | 4 | 23S-1, 23S-7 |
| Rhipicephalus turanicus | 10 | 4 | G4, G12 | 4 | 4 | D7, D12 | 4 | 4 | R9 | 4 | 3 | 16S-24, 16S-25 | 4 | 4 | 23S-4, 23S-25 |
| Rhipicephalus spp. | 10 | 7 | G10, G13 | 7 | 3 | D3, D15 D17, | 7 | 3 | R5 | 7 | 5 | 16S-4, 16S-26, 16S-27 | 7 | 7 | 23S-10, 23S-11, 23S-26 |
| Target Gene | Primer Name | Sequence 5’→ 3’ | PCR Type | Tm (°C) | Fragment Size (bp) | Reference |
|---|---|---|---|---|---|---|
| Ticks 16S rDNA | mt-rrs 1 | CTGCTCAATGATTTTTTAAATTGCTGTGG | Single | 55 | 401–416 | [61] |
| mt-rrs 2 | CCGGTCTGAACTCAGATCAAGTA | |||||
| Coxiella groEL | Cox-GrF1 | TTTGAAAAYATGGGCGCKCAAATGGT | 1st PCR | 56 | 655 | [19] |
| Cox-GrR2 | CGRTCRCCAAARCCAGGTGC | |||||
| Cox-GrF2 | GAAGTGGCTTCGCRTACWTCAGACG | 2nd PCR | 56 | 619 | [20] | |
| Cox-GrR1 | CCAAARCCAGGTGCTTTYAC | |||||
| Coxiella dnaK | Cox-dnaKF1 | CGTCARGCRACGAARGATGCA | 1st PCR | 54 | 777 | [20] |
| Cox-dnaKR | CGTCATGAYKCCGCCYAAGG | |||||
| Cox-dnaKF3 | GGTACKTTYGATATTTCCATC | Alternative 1st PCR | 54 | 636 | [20] | |
| Cox-dnaKR | CGTCATGAYKCCGCCYAAGG | |||||
| Cox-dnaKF2 | GAAGTGGATGGCGARCAYCAATT | 2nd PCR | 54 | 603 | [20] | |
| Cox-dnaKR | CGTCATGAYKCCGCCYAAGG | |||||
| Cox-dnaKF3 | GGTACKTTYGATATTTCCATC | Alternative 2nd PCR | 54 | 512 | [20] | |
| Cox-dnaKR3 | CTTGAATAGCYGCACCAATAGC | |||||
| Coxiella rpoB | Cox-rpoBF2 | GGGCGNCAYGGWAAYAAAGGSGT | 1st PCR | 56 | 607–610 | [20] |
| Cox-rpoBR1 | CACCRAAHCGTTGACCRCCAAATTG | |||||
| Cox-rpoBF3 | TCGAAGAYATGCCYTATTTAGAAG | 2nd PCR | 56 | 539–542 | [20] | |
| Cox-rpoBR3 | AGCTTTMCCACCSARGGGTTGCTG | |||||
| Coxiella 16S rDNA | Cox-16SF1 | CGTAGGAATCTACCTTRTAGWGG | 1st PCR | 52–56 | 1321–1429 | [19,20] |
| Cox-16SR2 | GCCTACCCGCTTCTGGTACAATT | |||||
| 16S-07F | AGAGTTTGATYMTGGCTCAG | Alternative 1st PCR | 52–56 | 1434–1542 | [19,20] | |
| Cox-16SR2 | GCCTACCCGCTTCTGGTACAATT | |||||
| Cox-16SF1 | CGTAGGAATCTACCTTRTAGWGG | 2nd PCR (fragment 1) | 52–56 | 719–826 | [20] | |
| Cox-16SR1 | ACTYYCCAACAGCTAGTTCTCA | |||||
| 16S-07F | AGAGTTTGATYMTGGCTCAG | Alternative 2nd PCR (fragment 1) | 52–56 | 832–939 | [20] | |
| Cox-16SR1 | ACTYYCCAACAGCTAGTTCTCA | |||||
| Cox-16SF2 | TGAGAACTAGCTGTTGGRRAGT | 2nd PCR (fragment 2) | 52–56 | 624–627 | [20] | |
| Cox-16SR2 | GCCTACCCGCTTCTGGTACAATT | |||||
| Cox16S_seq1 | TCTACGCATTTCACCGCTAC | Sequencing | This study | |||
| Cox16S_seq2 | AGTCGGATGTGAAAGCCCTA | |||||
| Cox16S_seq3 | CCTGTCACTCGGTTCCCAAA | |||||
| Cox16S_seq4 | CTGACACTGAGGCCGCGAAAGC | |||||
| Coxiella 23S rDNA | Cox-23SF1 | GCCTGCGAWAAGCTTCGGGGAG | 1st PCR | 56 | 694–1188 | [20] |
| Cox-23SR2 | CTCCTAKCCACASCTCATCCCC | |||||
| Cox-23SF2 | GATCCGGAGATWTCYGAATGGGG | 2nd PCR run | 56 | 583–867 | [20] | |
| Cox-23SR1 | TCGYTCGGTTTCGGGTCKACTC | |||||
| Cox-23SF1 | GCCTGCGAWAAGCTTCGGGGAG | Alternative 2nd PCR | 56 | 601–884 | [20] | |
| Cox-23SR2 | CTCCTAKCCACASCTCATCCCC |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kobayashi, T.; Chatanga, E.; Qiu, Y.; Simuunza, M.; Kajihara, M.; Hang’ombe, B.M.; Eto, Y.; Saasa, N.; Mori-Kajihara, A.; Simulundu, E.; et al. Molecular Detection and Genotyping of Coxiella-Like Endosymbionts in Ticks Collected from Animals and Vegetation in Zambia. Pathogens 2021, 10, 779. https://doi.org/10.3390/pathogens10060779
Kobayashi T, Chatanga E, Qiu Y, Simuunza M, Kajihara M, Hang’ombe BM, Eto Y, Saasa N, Mori-Kajihara A, Simulundu E, et al. Molecular Detection and Genotyping of Coxiella-Like Endosymbionts in Ticks Collected from Animals and Vegetation in Zambia. Pathogens. 2021; 10(6):779. https://doi.org/10.3390/pathogens10060779
Chicago/Turabian StyleKobayashi, Toshiya, Elisha Chatanga, Yongjin Qiu, Martin Simuunza, Masahiro Kajihara, Bernard Mudenda Hang’ombe, Yoshiki Eto, Ngonda Saasa, Akina Mori-Kajihara, Edgar Simulundu, and et al. 2021. "Molecular Detection and Genotyping of Coxiella-Like Endosymbionts in Ticks Collected from Animals and Vegetation in Zambia" Pathogens 10, no. 6: 779. https://doi.org/10.3390/pathogens10060779