
Identification of the Multiresistance Gene poxtA in Oxazolidinone-Susceptible Staphylococcus haemolyticus and Staphylococcus saprophyticus of Pig and Feed Origins

Abstract
:1. Introduction
2. Results
2.1. Identification of poxtA Gene in CoNS Isolates
2.2. Distribution of ARGs in poxtA-Positive CoNS Isolates and the Electrotransformants
2.3. Antimicrobial Susceptibility
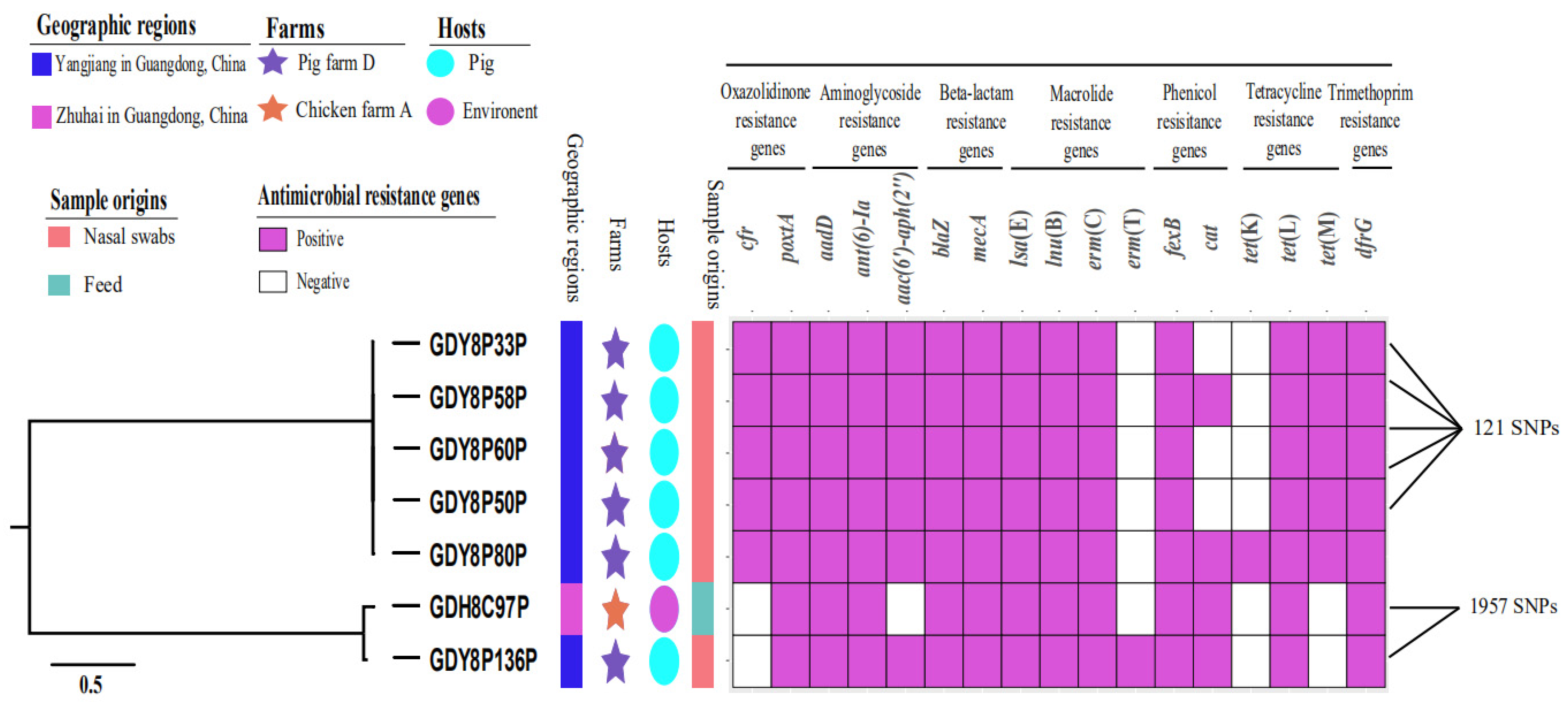
2.4. Phylogenetic Relatedness of poxtA-Positive CoNS Isolates
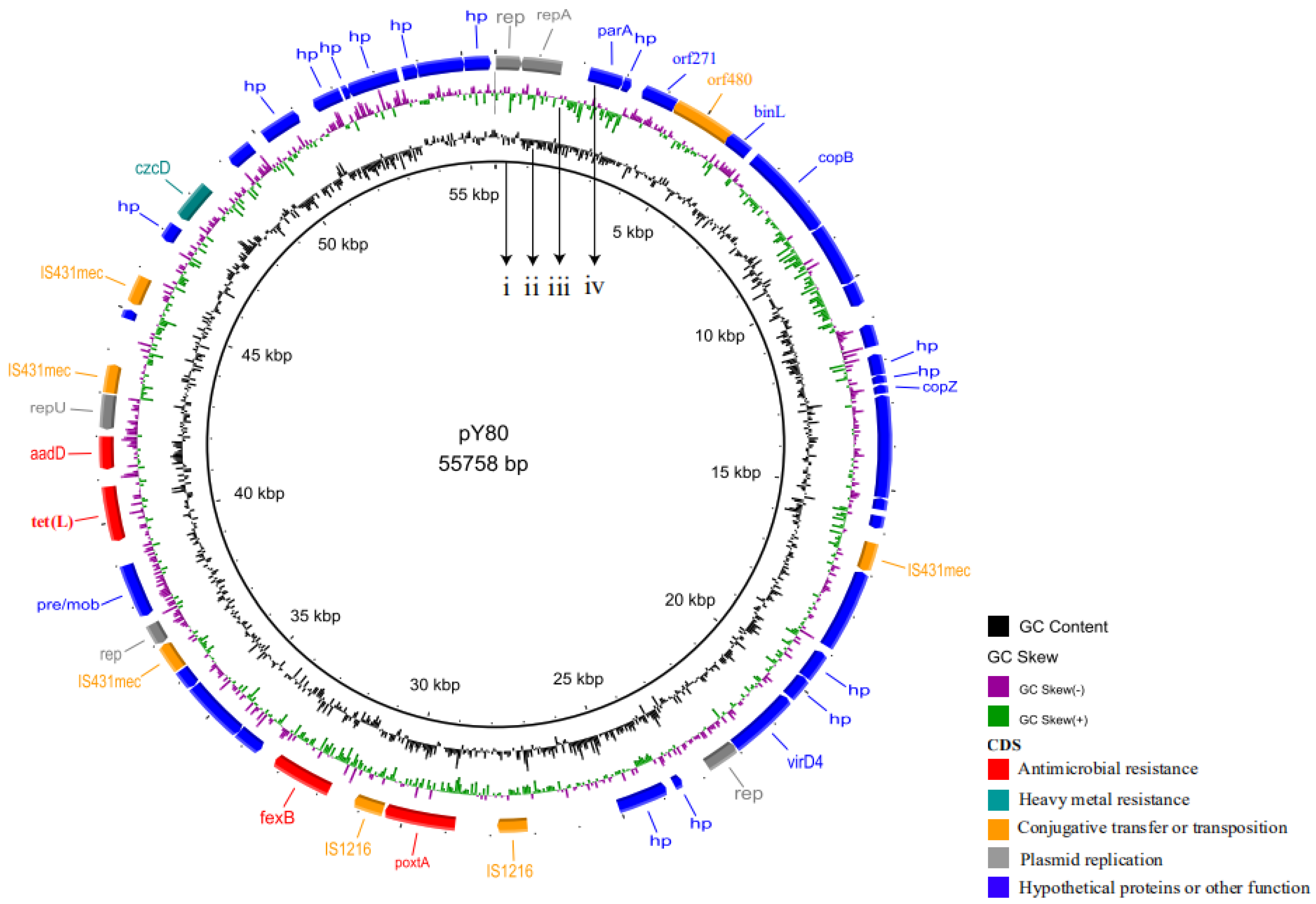
2.5. Plasmids Analysis
2.6. Genetic Environment of poxtA Gene
3. Discussion
4. Materials and Methods
4.1. Bacterial Isolations and Detection of poxtA Gene
4.2. Molecular Epidemiology Analysis and Transformation Assays
4.3. Antimicrobial Susceptibility Testing
4.4. S1-PFGE and WGS Analysis
4.5. Nucleotide Sequence Accession Numbers
4.6. Ethical Considerations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Al Tayyar, I.A.; Al-Zoubi, M.S.; Hussein, E.; Khudairat, S.; Sarosiekf, K. Prevalence and antimicrobial susceptibility pattern of coagulase-negative staphylococci (CoNS) isolated from clinical specimens in Northern of Jordan. Iran. J. Microbiol. 2015, 7, 294–301. [Google Scholar]
- Czekaj, T.; Ciszewski, M.; Szewczyk, E.M. Staphylococcus haemolyticus—An emerging threat in the twilight of the antibiotics age. Microbiology 2015, 161, 2061–2068. [Google Scholar] [CrossRef]
- Argemi, X.; Hansmann, Y.; Prola, K.; Prevost, G. Coagulase-Negative Staphylococci Pathogenomics. Int. J. Mol. Sci. 2019, 20, 1215. [Google Scholar] [CrossRef] [Green Version]
- Venugopal, N.; Mitra, S.; Tewari, R.; Ganaie, F.; Shome, R.; Rahman, H.; Shome, B.R. Molecular detection and typing of methicillin-resistant Staphylococcus aureus and methicillin-resistant coagulase-negative staphylococci isolated from cattle, animal handlers, and their environment from Karnataka, Southern Province of India. Vet. World. 2019, 12, 1760–1768. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teeraputon, S.; Santanirand, P.; Wongchai, T.; Songjang, W.; Lapsomthob, N.; Jaikrasun, D.; Toonkaew, S.; Tophon, P. Prevalence of methicillin resistance and macrolide-lincosamide-streptogramin B resistance in Staphylococcus haemolyticus among clinical strains at a tertiary-care hospital in Thailand. New Microbes New Infect. 2017, 19, 28–33. [Google Scholar] [CrossRef]
- Farrell, D.J.; Mendes, R.E.; Bensaci, M. In vitro activity of tedizolid against clinical isolates of Staphylococcus lugdunensis and Staphylococcus haemolyticus from Europe and the United States. Diagn. Microbiol. Infect. Dis. 2019, 93, 85–88. [Google Scholar] [CrossRef] [PubMed]
- Paiva-Santos, W.; Sousa, V.S.; Giambiagi-deMarval, M. Occurrence of virulence-associated genes among Staphylococcus saprophyticus isolated from different sources. Microb. Pathog. 2018, 119, 9–11. [Google Scholar] [CrossRef] [PubMed]
- Douros, A.; Grabowski, K.; Stahlmann, R. Drug-drug interactions and safety of linezolid, tedizolid, and other oxazolidinones. Expert Opin. Drug Metab. Toxicol. 2015, 11, 1849–1859. [Google Scholar] [CrossRef] [PubMed]
- Bender, J.K.; Cattoir, V.; Hegstad, K.; Sadowy, E.; Coque, T.M.; Westh, H.; Hammerum, A.M.; Schaffer, K.; Burns, K.; Murchan, S.; et al. Update on prevalence and mechanisms of resistance to linezolid, tigecycline and daptomycin in enterococci in Europe: Towards a common nomenclature. Drug Resist. Updates 2018, 40, 25–39. [Google Scholar] [CrossRef]
- Tewhey, R.; Gu, B.; Kelesidis, T.; Charlton, C.; Bobenchik, A.; Hindler, J.; Schork, N.J.; Humphries, R.M. Mechanisms of Linezolid Resistance among Coagulase-Negative Staphylococci Determined by Whole-Genome Sequencing. mBio 2014, 5, e00894-14. [Google Scholar] [CrossRef] [Green Version]
- Deshpande, L.M.; Castanheira, M.; Flamm, R.K.; Mendes, R.E. Evolving oxazolidinone resistance mechanisms in a worldwide collection of enterococcal clinical isolates: Results from the SENTRY Antimicrobial Surveillance Program. J. Antimicrob. Chemother. 2018, 73, 2314–2322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hao, H.H.; Sander, P.; Iqbal, Z.; Wang, Y.L.; Cheng, G.Y.; Yuan, Z.H. The Risk of Some Veterinary Antimicrobial Agents on Public Health Associated with Antimicrobial Resistance and their Molecular Basis. Front. Microbiol. 2016, 7, 1626. [Google Scholar] [CrossRef] [Green Version]
- Shen, J.Z.; Wang, Y.; Schwarz, S. Presence and dissemination of the multiresistance gene cfr in Gram-positive and Gram-negative bacteria. J. Antimicrob. Chemother. 2013, 68, 1697–1706. [Google Scholar] [CrossRef]
- Huang, J.H.; Chen, L.; Wu, Z.W.; Wang, L.P. Retrospective analysis of genome sequences revealed the wide dissemination of optrA in Gram-positive bacteria. J. Antimicrob. Chemother. 2017, 72, 614–616. [Google Scholar] [CrossRef] [Green Version]
- Huang, J.H.; Wang, M.L.; Gao, Y.; Chen, L.; Wang, L.P. Emergence of plasmid-mediated oxazolidinone resistance gene poxtA from CC17 Enterococcus faecium of pig origin. J. Antimicrob. Chemother. 2020, 75, 1359–1361. [Google Scholar] [CrossRef] [PubMed]
- Antonelli, A.; D’Andrea, M.M.; Brenciani, A.; Galeotti, C.L.; Morroni, G.; Pollini, S.; Varaldo, P.E.; Rossolini, G.M. Characterization of poxtA, a novel phenicol-oxazolidinone-tetracycline resistance gene from an MRSA of clinical origin. J. Antimicrob. Chemother. 2018, 73, 1763–1769. [Google Scholar] [CrossRef] [Green Version]
- Egan, S.A.; Shore, A.C.; O’Connell, B.; Brennan, G.I.; Coleman, D.C. Linezolid resistance in Enterococcus faecium and Enterococcus faecalis from hospitalized patients in Ireland: High prevalence of the MDR genes optrA and poxtA in isolates with diverse genetic backgrounds. J. Antimicrob. Chemother. 2020, 75, 1704–1711. [Google Scholar] [CrossRef]
- Freitas, A.R.; Tedim, A.P.; Duarte, B.; Elghaieb, H.; Abbassi, M.S.; Hassen, A.; Read, A.; Alves, V.; Novais, C.; Peixe, L. Linezolid-resistant (Tn6246::fexB-poxtA) Enterococcus faecium strains colonizing humans and bovines on different continents: Similarity without epidemiological link. J. Antimicrob. Chemother. 2020, 75, 2416–2423. [Google Scholar] [CrossRef] [PubMed]
- Xiong, W.G.; Wang, Y.L.; Sun, Y.X.; Ma, L.P.; Zeng, Q.L.; Jiang, X.T.; Li, A.T.; Zeng, Z.L.; Zhang, T. Antibiotic-mediated changes in the fecal microbiome of broiler chickens define the incidence of antibiotic resistance genes. Microbiome 2018, 6, 34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fioriti, S.; Morroni, G.; Coccitto, S.N.; Brenciani, A.; Antonelli, A.; Di Pilato, V.; Baccani, I.; Pollini, S.; Cucco, L.; Morelli, A.; et al. Detection of oxazolidinone resistance genes and characterization of genetic environments in enterococci of swine origin, Italy. Microorganisms 2020, 8, 2021. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Lv, Y.; Cai, J.C.; Schwarz, S.; Cui, L.Q.; Hu, Z.D.; Zhang, R.; Li, J.; Zhao, Q.; He, T.; et al. A novel gene, optrA, that confers transferable resistance to oxazolidinones and phenicols and its presence in Enterococcus faecalis and Enterococcus faecium of human and animal origin. J. Antimicrob. Chemother. 2015, 70, 2182–2190. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.Y.; Li, X.W.; Fu, Y.L.; Chen, Y.Q.; Wang, Y.; Ye, D.Y.; Wang, C.F.; Hu, X.; Zhou, L.; Du, J.J.; et al. Association of florfenicol residues with the abundance of oxazolidinone resistance genes in livestock manures. J. Hazard. Mater. 2020, 399, 123059. [Google Scholar] [CrossRef]
- Brenciani, A.; Morroni, G.; Vincenzi, C.; Manso, E.; Mingoia, M.; Giovanetti, E.; Varaldo, P.E. Detection in Italy of two clinical Enterococcus faecium isolates carrying both the oxazolidinone and phenicol resistance gene optrA and a silent multiresistance gene cfr. J. Antimicrob. Chemother. 2016, 71, 1118–1119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Wang, Y.; Schwarz, S.; Wang, S.L.; Chen, L.R.; Wu, C.M.; Shen, J.Z. Investigation of a multiresistance gene cfr that fails to mediate resistance to phenicols and oxazolidinones in Enterococcus faecalis. J. Antimicrob. Chemother. 2014, 69, 892–898. [Google Scholar] [CrossRef] [Green Version]
- Subbiah, M.; Mitchell, S.M.; Ullman, J.L.; Call, D.R. β-Lactams and florfenicol antibiotics remain bioactive in soils while ciprofloxacin, neomycin, and tetracycline are neutralized. Appl. Environ. Microbiol. 2011, 77, 7255–7260. [Google Scholar] [CrossRef] [Green Version]
- Tenover, F.C.; Arbeit, R.D.; Goering, R.V.; Mickelsen, P.A.; Murray, B.E.; Persing, D.H.; Swaminathan, B. Interpreting chromosomal DNA restriction patterns produced by pulsed-field gel electrophoresis: Criteria for bacterial strain typing. J. Clin. Microbiol. 1995, 33, 2233–2239. [Google Scholar] [CrossRef] [Green Version]
- Li, S.M.; Zhou, Y.F.; Li, L.; Fang, L.X.; Duan, J.H.; Liu, F.R.; Liang, H.Q.; Wu, Y.T.; Gu, W.Q.; Liao, X.P.; et al. Characterization of the Multi-Drug Resistance Gene cfr in Methicillin-Resistant Staphylococcus aureus (MRSA) Strains Isolated from Animals and Humans in China. Front. Microbiol. 2018, 9, 2925. [Google Scholar] [CrossRef]
- Clinical and Laboratory Standards Institute. Performance Standards for Antimicrobial Disk and Diffusion Susceptibility Tests for Bacteria Isolated from Animals: Second Informational Supplement VET01-S2; CLSI: Wayne, PA, USA, 2013. [Google Scholar]
- Clinical and Laboratory Standards Institute. Performance Standards for Antimicrobial Susceptibility Testing: Twenty-Sixth Informational Supplement M100-S30; CLSI: Wayne, PA, USA, 2020. [Google Scholar]
- Barton, B.M.; Harding, G.P.; Zuccarelli, A.J. A general method for detecting and sizing large plasmids. Anal. Biochem. 1995, 226, 235–240. [Google Scholar] [CrossRef] [PubMed]
- Chin, C.S.; Alexander, D.H.; Marks, P.; Klammer, A.A.; Drake, J.; Heiner, C.; Clum, A.; Copeland, A.; Huddleston, J.; Eichler, E.E.; et al. Nonhybrid, finished microbial genome assemblies from long-read SMRT seqfuencing data. Nat. Methods 2013, 10, 563–569. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Durbin, R. Fast and accurate long-read alignment with Burrows-Wheeler transform. Bioinformatics 2010, 26, 589–595. [Google Scholar] [CrossRef] [Green Version]
- Overbeek, R.; Olson, R.; Pusch, G.D.; Olsen, G.J.; Davis, J.J.; Disz, T.; Edwards, R.A.; Gerdes, S.; Parrello, B.; Shukla, M.; et al. The SEED and the Rapid Annotation of Microbial Genomes Using Subsystems Technology (RAST). Nucleic Acids Res. 2014, 42, D206–D214. [Google Scholar] [CrossRef] [PubMed]
- Zankari, E.; Hasman, H.; Cosentino, S.; Vestergaard, M.; Rasmussen, S.; Lund, O.; Aarestrup, F.M.; Larsen, M.V. Identification of acquired antimicrobial resistance genes. J. Antimicrob. Chemother. 2012, 67, 2640–2644. [Google Scholar] [CrossRef]
- Sullivan, M.J.; Petty, N.K.; Beatson, S.A. Easyfig: A genome comparison visualizer. Bioinformatics 2011, 27, 1009–1010. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Isolate | Sampling Time | Origin (Farm Type) | Species | MLST |
|---|---|---|---|---|
| GDH8C97P | June 2018 | Feed sample (chicken farm A) | S. saprophyticus | – |
| GDY8P33P | December 2018 | swine nasal swab (pig farm D) | S. haemolyticus | ST64 |
| GDY8P50P | December 2018 | swine nasal swab (pig farm D) | S. haemolyticus | ST64 |
| GDY8058P | December 2018 | swine nasal swab (pig farm D) | S. haemolyticus | ST64 |
| GDY8P60P | December 2018 | swine nasal swab (pig farm D) | S. haemolyticus | ST64 |
| GDY8P80P | December 2018 | swine nasal swab (pig farm D) | S. haemolyticus | ST64 |
| GDY8P136P | December 2018 | swine nasal swab (pig farm D) | S. saprophyticus | – |
| Bacterial Isolate | MICs (mg/L) | Resistance Genes | ||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| AMO | PEN | FOX | GEN | AMI | NEO | KAN | DOX | TET | TIG | FFC | ERY | RIF | VAN | CIP | ENR | LZD | TZD | SXT | ||
| S. aureusRN4220 | 0.125 | 0.125 | 2 | 0.25 | 1 | 0.25 | 0.25 | 0.125 | 0.5 | 0.06 | 2 | 0.25 | 0.008 | 1 | 0.5 | 0.25 | 0.5 | 0.06 | 0.25 | - |
| GDH8C97P | 8 | 16 | 8 | 0.125 | 0.25 | 1 | 0.25 | >64 | >64 | 0.25 | >64 | >256 | 0.015 | 1 | 4 | 16 | 0.5 | 0.06 | 0.5 | aadD, fexB, poxtA, tet(L) |
| RN4220/pH97 | 0.06 | 0.125 | 2 | 0.125 | 0.25 | 0.25 | 0.25 | 1 | 2 | 0.06 | 32 | >256 | 0.008 | 1 | 0.25 | 0.125 | 0.5 | 0.06 | 0.25 | fexB, poxtA, tet(L) |
| GDY8P33P | 2 | 4 | 16 | 8 | 2 | 4 | 8 | >64 | >64 | 0.25 | >64 | >256 | 0.004 | 1 | >64 | 32 | 2 | 0.06 | 2 | aadD, fexB, poxtA, tet(L), tet(M) |
| RN4220/pY33 | 0.06 | 0.125 | 2 | 0.125 | 2 | 1 | 0.25 | 1 | 2 | 0.125 | 32 | 0.125 | 0.008 | 1 | 0.5 | 0.125 | 0.5 | 0.06 | 0.25 | aadD, fexB, poxtA, tet(L) |
| GDY8P50P | 4 | 4 | 32 | 8 | 2 | 4 | 8 | 64 | >64 | 0.25 | >64 | >256 | 0.004 | 1 | >64 | >64 | 2 | 0.25 | 8 | aadD, fexB, poxtA, tet(L), tet(M) |
| RN4220/pY50 | 0.06 | 0.125 | 2 | 0.125 | 2 | 1 | 0.25 | 2 | 2 | 0.125 | 32 | 0.125 | 0.008 | 1 | 0.5 | 0.125 | 0.5 | 0.06 | 0.25 | aadD, fexB, poxtA, tet(L) |
| GDY8058P | 4 | 4 | 32 | 8 | 2 | 4 | 8 | 64 | >64 | 0.25 | >64 | >256 | 0.004 | 1 | >64 | >64 | 4 | 0.5 | 16 | aadD, fexB, poxtA, tet(L), tet(M) |
| RN4220/pY58 | 0.06 | 0.125 | 2 | 0.125 | 2 | 1 | 0.25 | 2 | 1 | 0.125 | 32 | 0.125 | 0.008 | 1 | 0.5 | 0.125 | 0.5 | 0.06 | 0.25 | aadD, fexB, poxtA, tet(L) |
| GDY8P60P | 2 | 1 | 32 | 8 | 2 | 4 | 8 | 64 | 64 | 0.25 | >64 | >256 | 0.008 | 1 | >64 | >64 | 2 | 0.5 | 8 | aadD, fexB, poxtA, tet(L), tet(M) |
| RN4220/pY60 | 0.06 | 0.125 | 2 | 0.125 | 2 | 1 | 0.25 | 1 | 1 | 0.25 | 32 | >256 | 0.002 | 1 | 0.5 | 0.125 | 0.5 | 0.06 | 0.25 | aadD, fexB, poxtA, tet(L) |
| GDY8P80P | 2 | 4 | 16 | 8 | 2 | 4 | 8 | 64 | >64 | 0.25 | >64 | >256 | 0.004 | 1 | >64 | >64 | 4 | 0.25 | 16 | aadD, fexB, poxtA, tet(L), tet(M) |
| RN4220/pY80 | 0.06 | 0.125 | 2 | 0.125 | 2 | 1 | 0.25 | 1 | 2 | 0.25 | 32 | 0.125 | 0.008 | 1 | 0.5 | 0.125 | 0.5 | 0.06 | 0.25 | aadD, fexB, poxtA, tet(L) |
| GDY8P136P | 16 | 16 | 16 | 8 | 0.5 | 4 | 8 | 32 | 64 | 0.25 | 64 | >256 | 0.03 | 2 | 4 | 2 | 2 | 0.25 | 1 | aadD, fexB, poxtA, tet(L) |
| RN4220/pY136 | 0.06 | 0.125 | 1 | 0.125 | 0.5 | 0.25 | 0.25 | 0.06 | 2 | 0.03 | 16 | 0.125 | 0.002 | 1 | 0.25 | 0.125 | 0.5 | 0.06 | 0.25 | fexB, poxtA, tet(L) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, L.; Hu, J.-X.; Liu, C.; Liu, J.; Ma, Z.-B.; Tang, Z.-Y.; Li, Y.-F.; Zeng, Z.-L. Identification of the Multiresistance Gene poxtA in Oxazolidinone-Susceptible Staphylococcus haemolyticus and Staphylococcus saprophyticus of Pig and Feed Origins. Pathogens 2021, 10, 601. https://doi.org/10.3390/pathogens10050601
Chen L, Hu J-X, Liu C, Liu J, Ma Z-B, Tang Z-Y, Li Y-F, Zeng Z-L. Identification of the Multiresistance Gene poxtA in Oxazolidinone-Susceptible Staphylococcus haemolyticus and Staphylococcus saprophyticus of Pig and Feed Origins. Pathogens. 2021; 10(5):601. https://doi.org/10.3390/pathogens10050601
Chicago/Turabian StyleChen, Lin, Jian-Xin Hu, Chang Liu, Jiao Liu, Zhen-Bao Ma, Zi-Yun Tang, Ya-Fei Li, and Zhen-Ling Zeng. 2021. "Identification of the Multiresistance Gene poxtA in Oxazolidinone-Susceptible Staphylococcus haemolyticus and Staphylococcus saprophyticus of Pig and Feed Origins" Pathogens 10, no. 5: 601. https://doi.org/10.3390/pathogens10050601