
Characterization of Ocular Surface Microbial Profiles Revealed Discrepancies between Conjunctival and Corneal Microbiota


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
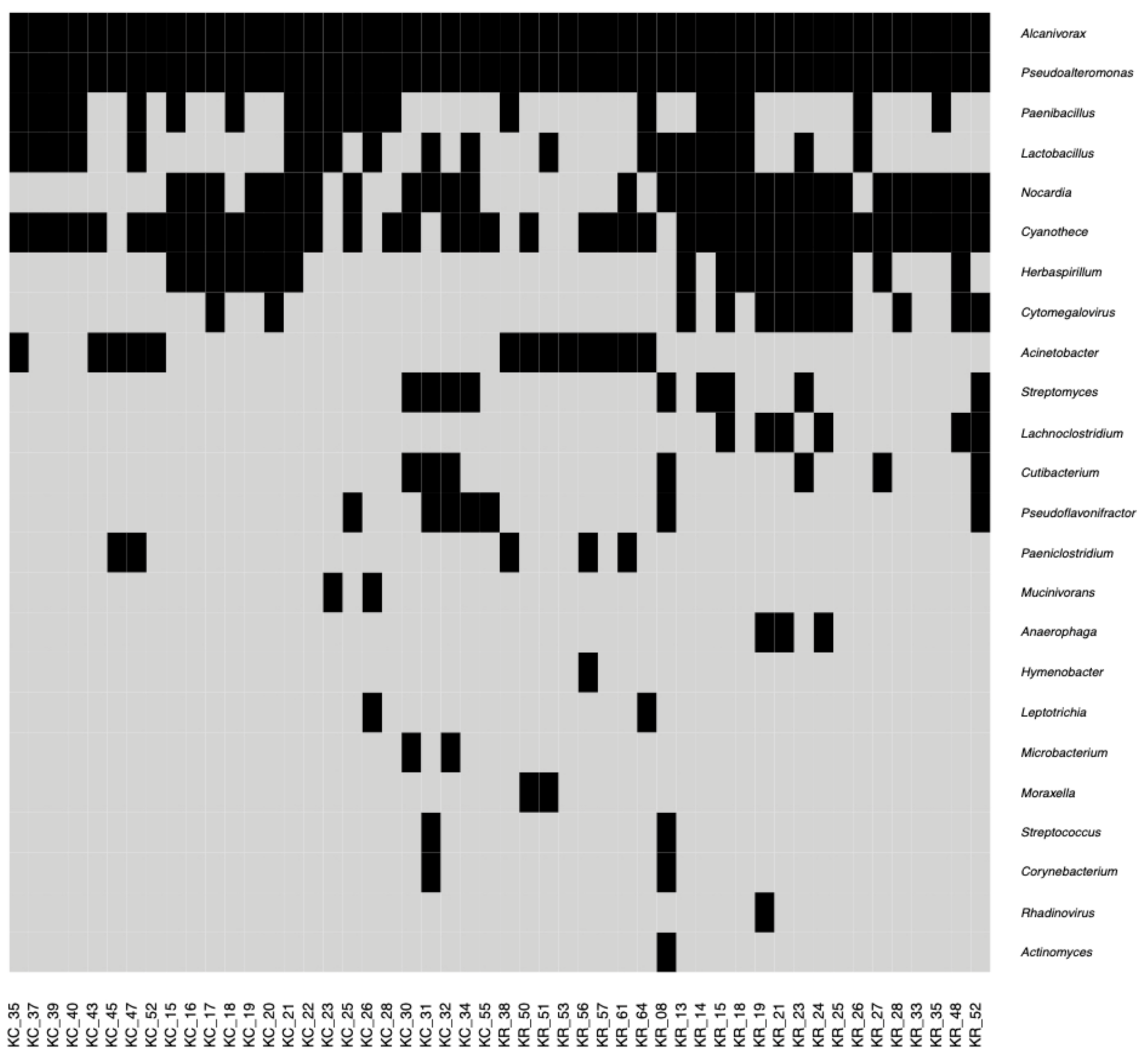
2.1. Characteristics of Bacterial, Archaeal and Viral Elements of the Human Corneal Microbiome Based on RNA-Sequencing Data
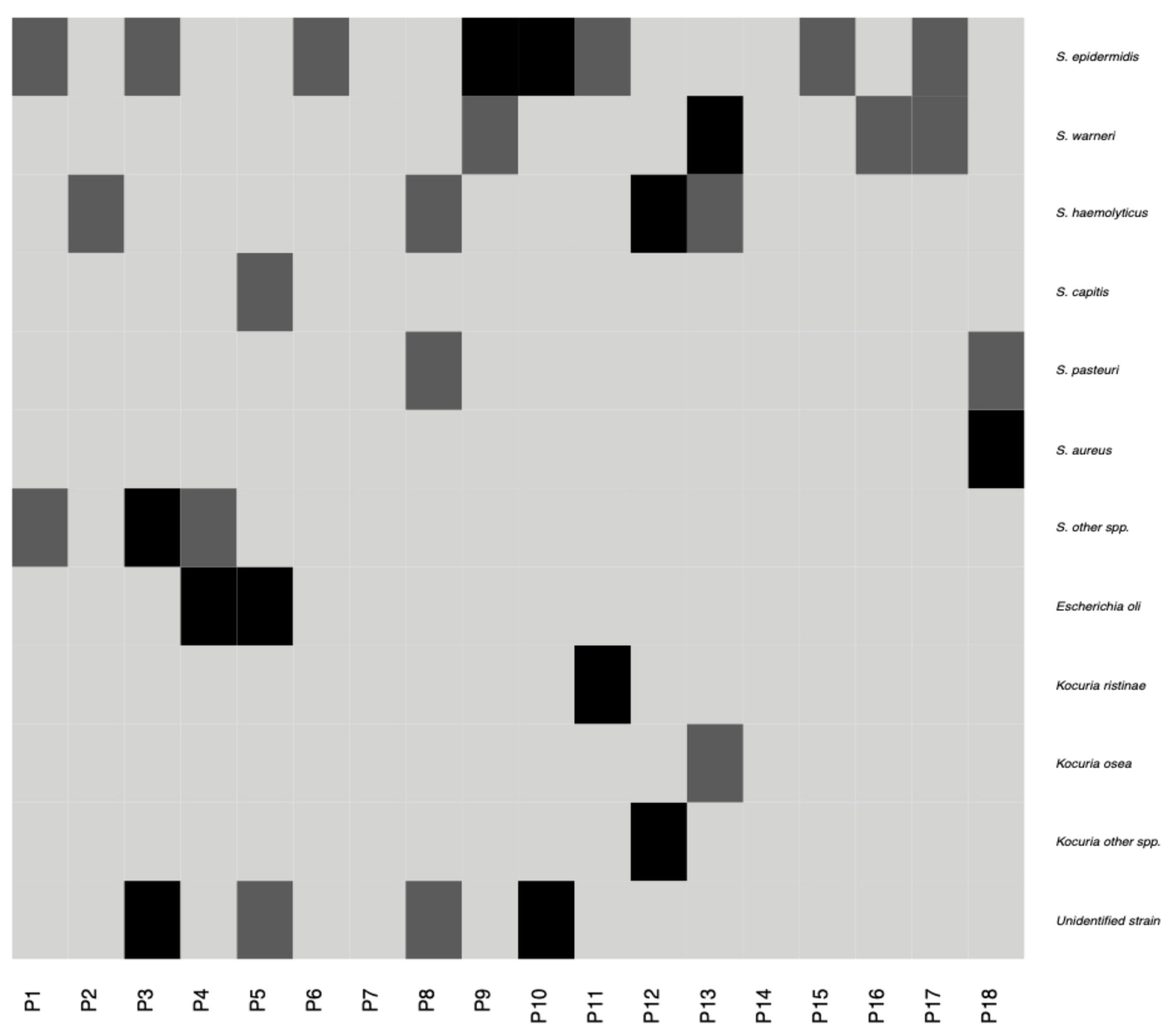
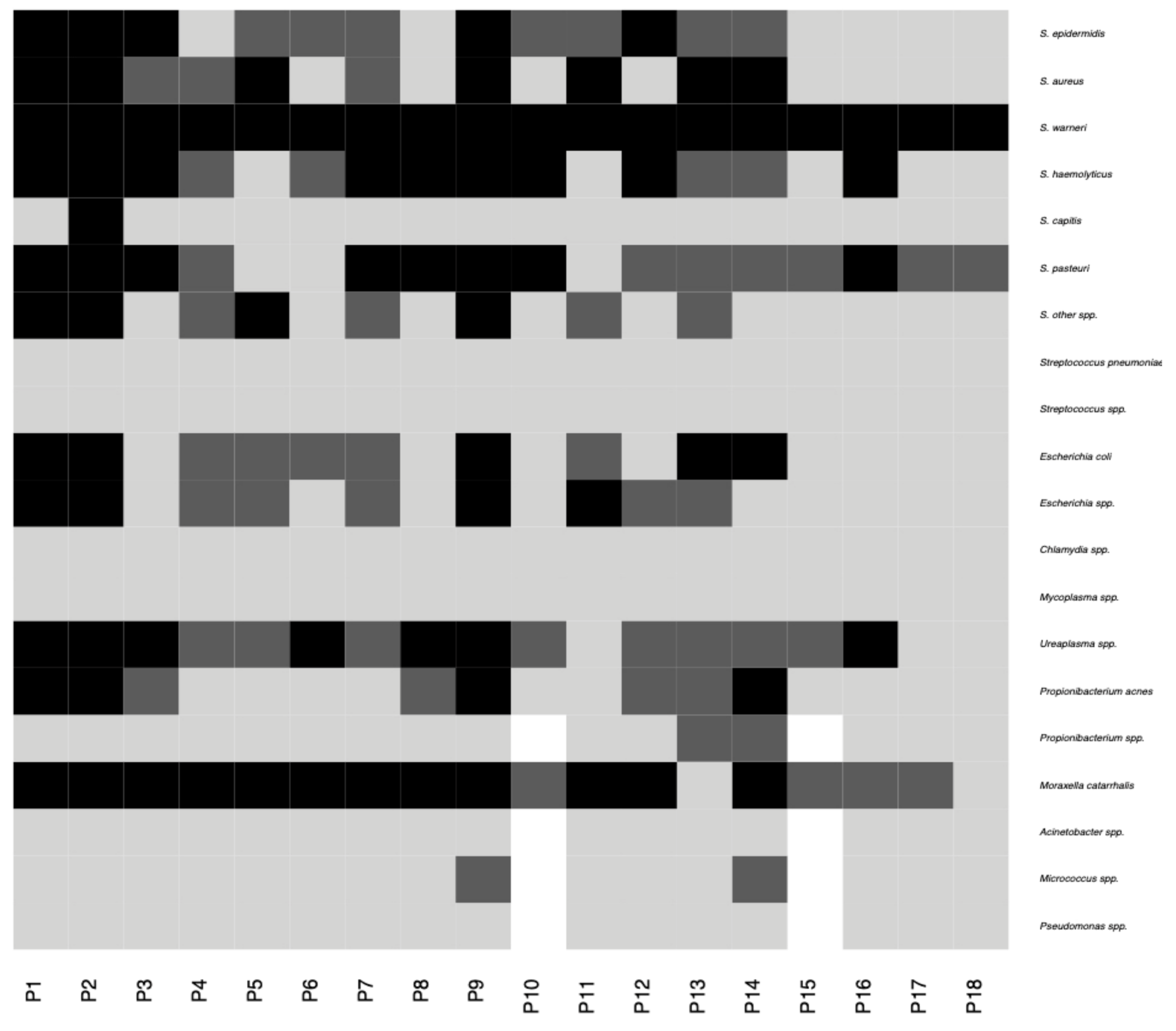
2.2. Characteristics of the Conjunctival Sample Microbiota
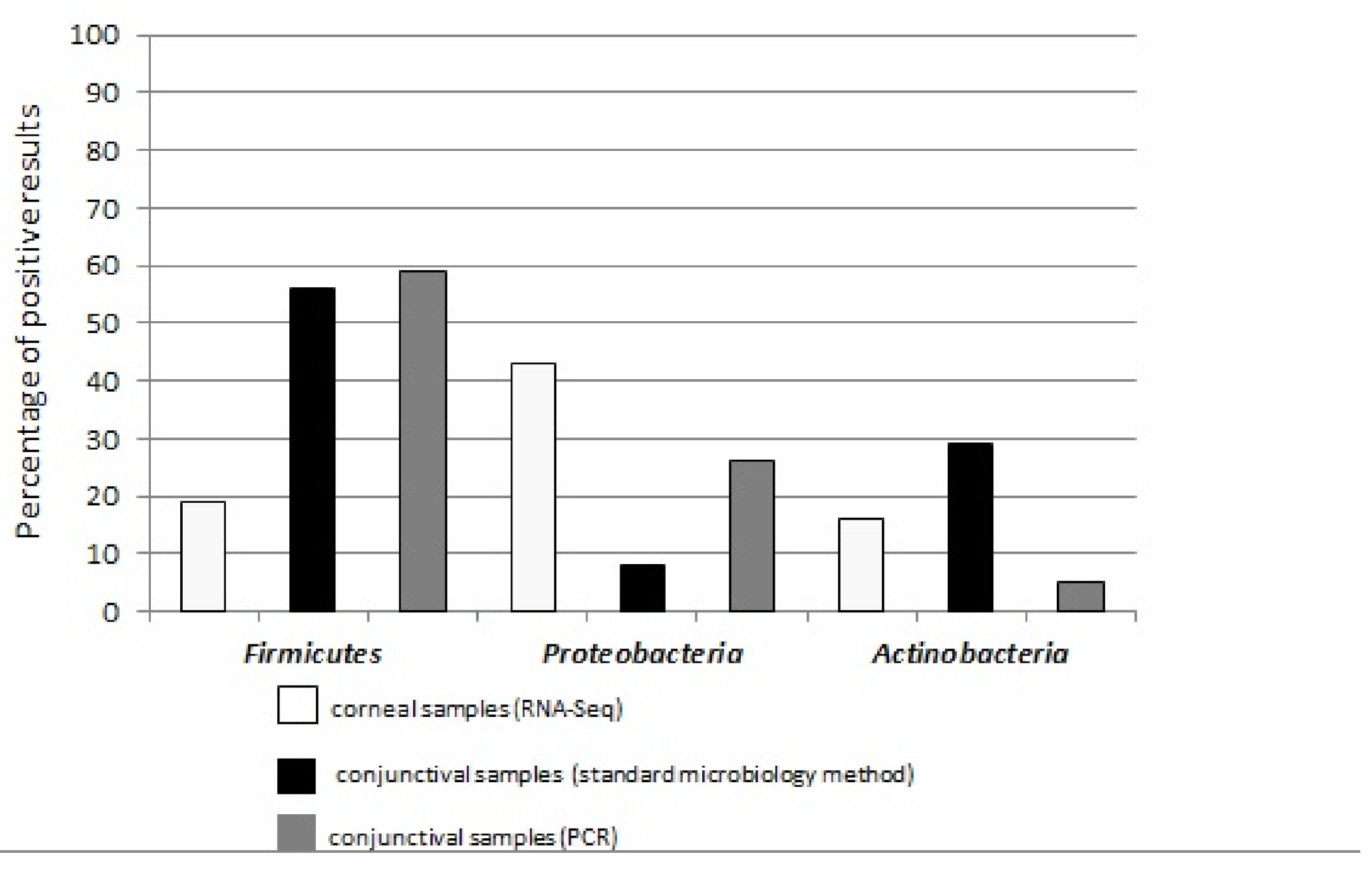
2.3. Similarities and Discrepancies in Conjunctival and Corneal Microbial Profiles
3. Discussion
4. Materials and Methods
4.1. Study Individuals and Materials
4.2. Identification of Bacterial, Archaeal and Viral Phyla in Corneal Samples
4.3. Assessment of Bacterial and Fungal Cultures Derived from Conjunctival Samples
4.4. Bacterial and Fungal Microbiome Elements—Molecular Detection in Conjunctival Samples
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Blum, H.E. The human microbiome. Adv. Med. Sci. 2017, 62, 414–420. [Google Scholar] [CrossRef]
- Zhou, Y.; Holland, M.J.; Makalo, P.; Joof, H.; Roberts, C.H.; Mabey, D.C.; Bailey, R.L.; Burton, M.J.; Weinstock, G.M.; E Burr, S. The conjunctival microbiome in health and trachomatous disease: A case control study. Genome Med. 2014, 6, 99. [Google Scholar] [CrossRef] [Green Version]
- De Almada, C.N.; de Almada, C.N.; Martinez, R.C.; Sant’Ana Ade, S. Characterization of the intestinal microbiota and its interaction with probiotics and health impacts. Appl. Microbiol. Biotechnol. 2015, 99, 4175–4199. [Google Scholar] [CrossRef]
- Rajoka, M.S.R.; Zhao, H.; Li, N.; Lu, Y.; Lian, Z.; Shao, D.; Jin, M.; Li, Q.; Zhao, L.; Shi, J. Origination, change, and modulation of geriatric dis-ease-related gut microbiota during life. Appl. Microbiol. Biotechnol. 2018, 102, 8275–8289. [Google Scholar] [CrossRef]
- Ozkan, J.; Willcox, M.D. The Ocular Microbiome: Molecular Characterisation of a Unique and Low Microbial Environment. Curr. Eye Res. 2019, 44, 685–694. [Google Scholar] [CrossRef]
- Doan, T.; Akileswaran, L.; Andersen, D.; Johnson, B.; Ko, N.; Shrestha, A.; Shestopalov, V.; Lee, C.S.; Lee, A.Y.; Van Gelder, R.N. Paucibacterial Microbiome and Resident DNA Virome of the Healthy Conjunctiva. Investig. Opthalmol. Vis. Sci. 2016, 57, 5116–5126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Willcox, M.D. Characterization of the normal microbiota of the ocular surface. Exp. Eye Res. 2013, 117, 99–105. [Google Scholar] [CrossRef] [PubMed]
- Dong, Q.; Brulc, J.M.; Iovieno, A.; Bates, B.; Garoutte, A.; Miller, D.; Revanna, K.V.; Gao, X.; Antonopoulos, D.A.; Slepak, V.Z.; et al. Diversity of Bacteria at Healthy Human Conjunctiva. Investig. Opthalmol. Vis. Sci. 2011, 52, 5408–5413. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ozkan, J.; Nielsen, S.; Diez-Vives, C.; Coroneo, M.; Thomas, T.; Willcox, M. Temporal Stability and Composition of the Ocular Surface Microbiome. Sci. Rep. 2017, 7, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, Y.; Yang, B.; Li, W. Defining the normal core microbiome of conjunctival microbial communities. Clin. Microbiol. Infect. 2016, 22, 643.e7–643.e12. [Google Scholar] [CrossRef] [Green Version]
- Capriotti, J.A.; Pelletier, J.S.; Shah, M.; Caivano, D.M.; Ritterband, D.C. Normal ocular flora in healthy eyes from a rural population in Sierra Leone. Int. Ophthalmol. 2008, 29, 81–84. [Google Scholar] [CrossRef]
- Eder, M.; Fariña, N.; Sanabria, R.R.; Ta, C.N.; Koss, M.; Samudio, M.; Cuevas, C.; Gines, A.; Simancas, M.; Klauss, V.; et al. Normal ocular flora in newborns delivered in two hospital centers in Argentina and Paraguay. Graefe’s Arch. Clin. Exp. Ophthalmol. 2005, 243, 1098–1107. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Rubio, M.E.; Rebolledo-Lara, L.; Martinez-García, M.; Alarcón-Tomás, M.; Cortés-Valdés, C.; E Fern, L.R.-L.M. The conjunctival bacterial pattern of diabetics undergoing cataract surgery. Eye 2009, 24, 825–834. [Google Scholar] [CrossRef] [Green Version]
- Graham, J.E.; Moore, J.E.; Jiru, X.; Moore, J.E.; Goodall, E.A.; Dooley, J.S.G.; Hayes, V.E.A.; Dartt, D.A.; Downes, C.S.; Moore, T.C.B. Ocular Pathogen or Commensal: A PCR-Based Study of Surface Bacterial Flora in Normal and Dry Eyes. Investig. Opthalmol. Vis. Sci. 2007, 48, 5616–5623. [Google Scholar] [CrossRef]
- Martins, E.N.; Alvarenga, L.S.; Höfling-Lima, A.L.; Freitas, D.; Zorat-Yu, M.C.; E Farah, M.; Mannis, M.J. Aerobic Bacterial Conjunctival Flora in Diabetic Patients. Cornea 2004, 23, 136–142. [Google Scholar] [CrossRef]
- Wu, T.G.; Mitchell, B.M.; Carothers, T.S.; Coats, D.K.; Brady-McCreery, K.M.; Paysse, E.A.; Wilhelmus, K.R. Molecular analysis of the pediatric ocular surface for fungi. Curr. Eye Res. 2003, 26, 33–36. [Google Scholar] [CrossRef]
- Zegans, M.E.; Van Gelder, R.N. Considerations in Understanding the Ocular Surface Microbiome. Am. J. Ophthalmol. 2014, 158, 420–422. [Google Scholar] [CrossRef] [Green Version]
- Ge, C.; Wei, C.; Yang, B.X.; Cheng, J.; Huang, Y.S. Conjunctival microbiome changes associated with fungal keratitis: Meta-genomic analysis. Int. J. Ophthalmol. 2019, 12, 194–200. [Google Scholar]
- Ozkan, J.; Willcox, M.; Wemheuer, B.; Wilcsek, G.; Coroneo, M.; Thomas, T. Biogeography of the human ocular microbiota. Ocul. Surf. 2019, 17, 111–118. [Google Scholar] [CrossRef]
- Cavuoto, K.M.; Mendez, R.; Miller, D.; Galor, A.; Banerjee, S. Effect of clinical parameters on the ocular surface microbiome in children and adults. Clin. Ophthalmol. 2018, 12, 1189–1197. [Google Scholar] [CrossRef] [Green Version]
- Kugadas, A.; Gadjeva, M. Impact of Microbiome on Ocular Health. Ocul. Surf. 2016, 14, 342–349. [Google Scholar] [CrossRef] [Green Version]
- Owji, N.; Zareifard, A. Bacterial Flora of the Conjunctiva One Year after Dacryocystorhinostomy. Middle East Afr. J. Ophthalmol. 2018, 25, 35–39. [Google Scholar] [CrossRef]
- Scharschmidt, T.C. Establishing Tolerance to Commensal Skin Bacteria. Dermatol. Clin. 2017, 35, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cosseau, C.; Romano-Bertrand, S.; Duplan, H.; Lucas, O.; Ingrassia, I.; Pigasse, C.; Roques, C.; Jumas-Bilak, E. Proteobacteria from the human skin microbiota: Species-level diversity and hypotheses. One Health 2016, 2, 33–41. [Google Scholar] [CrossRef] [Green Version]
- Wilantho, A.; Deekaew, P.; Srisuttiyakorn, C.; Tongsima, S.; Somboonna, N. Diversity of bacterial communities on the facial skin of different age-group Thai males. PeerJ 2017, 5, e4084. [Google Scholar] [CrossRef] [Green Version]
- Almeida, A.; Mitchell, A.L.; Boland, M.; Forster, S.C.; Gloor, G.B.; Tarkowska, A.; Lawley, T.D.; Finn, R.D. A new genomic blueprint of the human gut microbiota. Nat. Cell Biol. 2019, 568, 499–504. [Google Scholar] [CrossRef] [Green Version]
- Bouslimani, A.; Da Silva, R.; Kosciolek, T.; Janssen, S.; Callewaert, C.; Amir, A.; Dorrestein, K.; Melnik, A.V.; Zaramela, L.S.; Kim, J.-N.; et al. The impact of skin care products on skin chemistry and microbiome dynamics. BMC Biol. 2019, 17, 47. [Google Scholar] [CrossRef]
- Theis, K.R.; Romero, R.; Winters, A.D.; Greenberg, J.M.; Gomez-Lopez, N.; Alhousseini, A.; Bieda, J.; Maymon, E.; Pacora, P.; Fettweis, J.M.; et al. Does the human placenta delivered at term have a microbiota? Results of cultivation, quantitative real-time PCR, 16S rRNA gene sequencing, and metagenomics. Am. J. Obstet. Gynecol. 2019, 220, 267.e1–267.e39. [Google Scholar] [CrossRef] [Green Version]
- Kugadas, A.; Wright, Q.; Geddes-McAlister, J.; Gadjeva, M. Role of Microbiota in Strengthening Ocular Mucosal Barrier Function Through Secretory IgA. Investig. Opthalmol. Vis. Sci. 2017, 58, 4593–4600. [Google Scholar] [CrossRef]
- De Kaspar, H.M.; Kreutzer, T.C.; Aguirre-Romo, I.; Ta, C.N.; Dudichum, J.; Bayrhof, M.; Klauss, V.; Kampik, A. A Prospective Randomized Study to Determine the Efficacy of Preoperative Topical Levofloxacin in Reducing Conjunctival Bacterial Flora. Am. J. Ophthalmol. 2008, 145, 136–142.e2. [Google Scholar] [CrossRef]
- Salas-Jara, M.J.; Ilabaca, A.; Vega, M.; García, A. Biofilm Forming Lactobacillus: New Challenges for the Development of Probiotics. Microorganisms 2016, 4, 35. [Google Scholar] [CrossRef]
- Maroniche, G.A.; García, J.E.; Salcedo, F.; Creus, C.M. Molecular identification of Azospirillum spp.: Limitations of 16S rRNA and qualities of rpo D as genetic markers. Microbiol. Res. 2017, 195, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Velásquez-Mejía, E.P.; de la Cuesta-Zuluaga, J.; Escobar, J.S. Impact of DNA extraction, sample dilution, and reagent contamination on 16S rRNA gene sequencing of human feces. Appl. Microbiol. Biotechnol. 2018, 102, 403–411. [Google Scholar] [CrossRef] [PubMed]
- Kirstahler, P.; Bjerrum, S.S.; Friis-Møller, A.; la Cour, M.; Aarestrup, F.M.; Westh, H.; Pamp, S.J. Genomics-based identification of microorganisms in human ocular body fluid. Sci. Rep. 2018, 8, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Turnbaugh, P.J.; Ley, R.E.; Hamady, M.; Fraser-Liggett, C.M.; Knight, R.; Gordon, J.I. The Human Microbiome Project. Nat. Cell Biol. 2007, 449, 804–810. [Google Scholar] [CrossRef]
- Wen, X.; Miao, L.; Deng, Y.; Bible, P.W.; Hu, X.; Zou, Y.; Liu, Y.; Guo, S.; Liang, J.; Chen, T.; et al. The Influence of Age and Sex on Ocular Surface Microbiota in Healthy Adults. Investig. Opthalmol. Vis. Sci. 2017, 58, 6030–6037. [Google Scholar] [CrossRef]
- Kabza, M.; A Karolak, J.; Rydzanicz, M.; Szcześniak, M.W.; Nowak, D.M.; Ginter-Matuszewska, B.; Polakowski, P.; Ploski, R.; Szaflik, J.P.; Gajecka, M. Collagen synthesis disruption and downregulation of core elements of TGF-β, Hippo, and Wnt pathways in keratoconus corneas. Eur. J. Hum. Genet. 2017, 25, 582–590. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mangul, S.; Loohuis, L.M.; Ori, A.P.; Jospin, G.; Koslicki, D.; Yang, H.T.; Wu, T.; Boks, M.P.; Lomen-Hoerth, C.; Wiedau-Pazos, M.; et al. Total RNA Sequencing reveals microbial communities in human blood and disease specific effects. bioRxiv 2016. [Google Scholar] [CrossRef] [Green Version]
- Dobin, A.; Davis, C.A.; Schlesinger, F.; Drenkow, J.; Zaleski, C.; Jha, S.; Batut, P.; Chaisson, M.; Gingeras, T.R. STAR: Ultrafast universal RNA-seq aligner. Bioinformatics 2013, 29, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Tarasov, A.; Vilella, A.J.; Cuppen, E.; Nijman, I.J.; Prins, P. Sambamba: Fast processing of NGS alignment formats. Bioinformatics 2015, 31, 2032–2034. [Google Scholar] [CrossRef]
- Quinlan, A.R.; Hall, I.M. BEDTools: A flexible suite of utilities for comparing genomic features. Bioinformatics 2010, 26, 841–842. [Google Scholar] [CrossRef] [Green Version]
- Menzel, P.; Ng, K.L.; Krogh, A. Fast and sensitive taxonomic classification for metagenomics with Kaiju. Nat. Commun. 2016, 7, 11257. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brister, J.R.; Ako-Adjei, D.; Bao, Y.; Blinkova, O. NCBI Viral Genomes Resource. Nucleic Acids Res. 2015, 43, D571–D577. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mende, D.R.; Letunic, I.; Huerta-Cepas, J.; Li, S.S.; Forslund, K.; Sunagawa, S.; Bork, P. proGenomes: A resource for consistent functional and taxonomic annotations of prokaryotic genomes. Nucleic Acids Res. 2017, 45, D529–D534. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ratajczak, M.; Kubicka, M.M.; Kamińska, D.; Sawicka, P.; Długaszewska, J. Microbiological quality of non-sterile pharmaceutical products. Saudi Pharm. J. 2015, 23, 303–307. [Google Scholar] [CrossRef] [Green Version]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Matysiak, A.; Kabza, M.; Karolak, J.A.; Jaworska, M.M.; Rydzanicz, M.; Ploski, R.; Szaflik, J.P.; Gajecka, M. Characterization of Ocular Surface Microbial Profiles Revealed Discrepancies between Conjunctival and Corneal Microbiota. Pathogens 2021, 10, 405. https://doi.org/10.3390/pathogens10040405
Matysiak A, Kabza M, Karolak JA, Jaworska MM, Rydzanicz M, Ploski R, Szaflik JP, Gajecka M. Characterization of Ocular Surface Microbial Profiles Revealed Discrepancies between Conjunctival and Corneal Microbiota. Pathogens. 2021; 10(4):405. https://doi.org/10.3390/pathogens10040405
Chicago/Turabian StyleMatysiak, Anna, Michal Kabza, Justyna A. Karolak, Marcelina M. Jaworska, Malgorzata Rydzanicz, Rafal Ploski, Jacek P. Szaflik, and Marzena Gajecka. 2021. "Characterization of Ocular Surface Microbial Profiles Revealed Discrepancies between Conjunctival and Corneal Microbiota" Pathogens 10, no. 4: 405. https://doi.org/10.3390/pathogens10040405