Nutrition, One-Carbon Metabolism and Neural Tube Defects: A Review


1
Department of Food Science and Nutrition, Zhejiang University, Hangzhou 310058, China
2
Fuli Institute, Zhejiang University, Hangzhou 310058, China
3
Monash Asia Institute and Departments of Medicine and of Nutrition and Dietetics, Monash University, Melbourne 3006, Australia
*
Author to whom correspondence should be addressed.
Nutrients 2016, 8(11), 741; https://doi.org/10.3390/nu8110741
Submission received: 18 August 2016
/
Revised: 6 November 2016
/
Accepted: 16 November 2016
/
Published: 23 November 2016
(This article belongs to the Special Issue B-Vitamins and One-Carbon Metabolism)
{kind=link}
Abstract
:Neural tube defects (NTDs) are a group of severe congenital malformations, induced by the combined effects of genes and the environment. The most valuable finding so far has been the protective effect of folic acid supplementation against NTDs. However, many women do not take folic acid supplements until they are pregnant, which is too late to prevent NTDs effectively. Long-term intake of folic acid–fortified food is a good choice to solve this problem, and mandatory folic acid fortification should be further promoted, especially in Europe, Asia and Africa. Vitamin B2, vitamin B-6, vitamin B-12, choline, betaine and n-3 polyunsaturated fatty acids (PUFAs) can also reduce the NTD risk by interacting with the one-carbon metabolism pathway. This suggest that multivitamin B combined with choline, betaine and n-3 PUFAs supplementation may have a better protective effect against NTDs than folic acid alone. Genetic polymorphisms involved in one-carbon metabolism are associated with NTD risk, and gene screening for women of childbearing age prior to pregnancy may help prevent NTDs induced by the risk allele. In addition, the consumption of alcohol, tea and coffee, and low intakes of fruit and vegetable are also associated with the increased risk of NTDs, and should be avoided by women of childbearing age.
Keywords:
folate; neural tube defects; vitamin B; choline; betaine; one-carbon metabolism; tea; alcohol; coffee1. Introduction
Neural tube defects (NTDs) are a group of severe congenital malformations. It is estimated that approximately one out of 1000 newborns present with this type of defect [1]. The development and closure of neural tube happen during normal embryogenesis between the 18th and 28th days after fertilization. Failure of the neural tube to close in embryonic development results in NTDs [2]. The etiology of NTDs is still unknown. One possible reason may be the disturbance of the one-carbon metabolism pathway [3,4]. However, there are also folate-resistant NTDs, indicating that folate deficiency is not the unique reason for NTDs, and other potential pathogeneses may also responsible for NTDs. The deficiency of other nutritional factors involved in one-carbon metabolism, such as vitamin B-2, B-6, B-12, choline, betaine, and n-3 polyunsaturated fatty acids, may be also associated with NTDs. Genetic factors are another important cause of NTDs. Many loci on genes have been identified as associated with the risk of NTDs, especially on genes involved in the one-carbon metabolism pathway, such as methylenetetrahydrofolate reductase (MTHFR) and 5-methyltetrahydrofolate-homocysteine methyltransferase (MTR) [5,6].
In this study, we review the research progress on the effect of nutritional factors and genes involved in the one-carbon metabolism pathway on NTDs. This should provide a basis for better nutritional approaches to NTD prevention.
2. B-Vitamins, One-Carbon Metabolism and NTDs
2.1. Folate
Folate, also known as vitamin B-9, plays an important role in the homocysteine (Hcy) metabolism, one-carbon pathway and DNA synthesis (Figure 1). In 1991, the Medical Research Council Vitamin Study Research Group conducted a large-scale randomized controlled trial (RCT) with 1817 women from 33 centers in seven countries to assess the effect of folic acid on the reoccurrence of neural tube defects and found that 4 mg folic acid supplementation per day lowered the risk of NTDs by 72% compared with the control group (without folic acid supplementation) [7]. Another important RCT was conducted by Czeizel et al. in Hungarian women without a history of NTD-affected pregnancy [3]. In that study, no NTD cases occurred in 2104 women who received multivitamins containing 0.8 mg folic acid per day, while six NTD cases occurred in 2052 women who received only trace elements and vitamin C. In 1999, Berry et al. conducted a cohort intervention study in an area of China with high rates of neural tube defects (the northern region) and one with low rates (the southern region), and found that preconceptional supplementation (starting supplementation before the last menstrual period before conception and stopping at the end of the first trimester) of 400 μg folic acid (a synthetic form of folate) per day reduced the NTD risk by 85% in the northern subgroup of 31,960 women and by 40% in the southern subgroup of 215,871 women [4]. These results of intervention studies have provided the most persuasive evidence for the protective effect of folic acid against NTDs.
In 1995, a case-control study by Daly first reported a dose-response relationship between the red blood cell folate concentration of mothers and the risk of NTDs, and that a red blood cell folate concentration ≥906 nmol/L (400 ng/mL) can provide optimal protection against NTDs [8]. In 2014, Crider et al. analyzed data from two intervention studies in Chinese women (400 μg folic acid per day from preconception through the end of the first trimester) by a Bayesian model, and found a dose-response relationship between maternal red blood cell folate and the risk of NTDs: a folate concentration >1000 nmol/L substantially reduced the risk of NTDs [5]. This was consistent with the result of Daly’s study. Based on these results, in 2015 the World Health Organization (WHO) recommended a threshold value of the maternal red blood cell folate concentration of 906 nmol/L (400 ng/mL) to prevent NTDs. Folic acid supplementation is a solution for the insufficient dietary intake of folate. A daily intake of 0.4 mg (at least one month before conception through the first three months of conception) folic acid is recommended by the Centers for Disease Control and Prevention for women who do not have a history of a previous NTD-affected pregnancy; this dose can prevent 50% of NTDs [9]. As for women who have had a previous NTD-affected pregnancy, the dose of folic acid is increased to 4 mg per day (at least one month before pregnancy through the first three months of pregnancy) [9]. Although recommendations for preconceptional folic acid supplementation have existed for decades, only a small number of women were actually supplemented with folic acid before conception [10]. Most women started folic acid supplementation when they knew they were pregnant, and this was often too late for the effective prevention of NTDs [11]. Long-term intake of folic acid–fortified foods should complement preconceptional folic acid supplementation. Arth et al. reported that mandatory folic acid fortification of wheat and maize flour prevented 13.2% folic acid–preventable NTDs (35,500 of approximately 268,700 global cases) in 58 countries [12]. Seventy-eight countries have fortified flour with folic acid mandatorily, while most countries in Asia, Europe and Africa do not mandate folic acid fortification [13]. Khoshnood et al. conducted an observational study to assess the long-term trend in NTD prevalence in 19 European countries and found that, without mandatory folic acid fortification, the long-existent recommendation for preconceptional folic acid supplementation and voluntary folic acid fortification did not significantly decrease the prevalence of NTDs [14]. In 2016, an intervention study in 16,648 women in Shanxi, China (one of the most NTD-affected regions, with a 13.8‰ to 19.9‰ prevalence), found that folic acid–fortified flour decreased the NTD burden by 58.5%, which has informed the future implementation of mandatory folic acid fortification in China [15]. Further promotion of mandatory folic acid fortification is needed to prevent NTDs.
Several genes involved in the folate-dependent one-carbon metabolism have been shown to be associated with the risk of NTDs. One of the most important is the gene encoding MTHFR, an enzyme that catalyzes the conversion of 5,10-methylenetetrahydrofolate (5,10-MTHF) to 5-methyltetrahydrofolate (5-MTHF) and provides the methyl group needed for remethylation of Hcy to form methionine (Met) (Figure 1) [16]. The T allele of MTHFR 677C>T is associated with an increased risk of NTDs in the western population [17,18,19]. Similar observations are made in Chinese mothers even when they supplement with folic acid [5,16,20]. Two considerations may help explain this association between MTHFR 677C>T and NTDs. On the one hand, the TT genotype of MTHFR 677C>T can attenuate the plasma and red blood cell (RBC) folate response to folic acid supplementation [21]. In addition, previous studies have reported that the T allele carriers of MTHFR 677C>T had lower folate concentrations than non-carriers [18,22]. On the other hand, the MTHFR 677C>T mutation is associated with reduced MTHFR activity [18,23]. MTHFR 1298A>C is yet another mutation associated with decreased MTHFR activity [23]. However, the link between MTHFR 1298A>C and NTDs remains controversial: one study in Italy reported that the C allele of MTHFR 1298A>C was associated with a higher risk of NTDs [24], several other studies found no significant association between MTHFR 1298A>C and NTDs [6,25,26,27], while one study in China even found that the C allele of MTHFR 1298A>C had a protective role against NTDs [28].
In addition to MTHFR, other genes involved in the folate metabolism have been demonstrated to influence the development of NTDs in the Chinese. MTR is an enzyme that catalyzes the remethylation of Hcy to Met and is dependent on the provision of methyl groups from 5-MTHF (Figure 1). The mutation of MTR is associated with the increased risk of NTDs [6,19,29,30]. Solute carrier family 19 member 1 (SCF19M1) can transport folate into cells. The mutation of SCF19M1 is associated with increased NTD risk even when mothers are supplemented with folic acid [6]. In addition, mutations on BHMT [6,19,31], CBS [19,32], MTRR [19,33,34], MTHFD1 [19,35,36,37,38,39,40,41], MTHFD2 [19], SHMT1 [36], FOLH1 [42], RFC1 [43,44], SARDH [45], PEMT [40], GART [40] and TYMS [19,36] have also been reported to be associated with the risk of NTDs.
2.2. Vitamins B-2, B-6 and B-12
Vitamin B-12 is the cofactor of MTR (Figure 1). Additionally, B-12 deficiency is associated with elevated Hcy [46,47]. In case-control studies, vitamin B-12 status has been found to be protective against NTDs in the Chinese [47,48]. According to a study in 1170 women in northwest China, the prevalence of vitamin B-12 deficiency was 45% [49], indicating that vitamin B-12 supplementation maybe also be needed to prevent NTDs in China. The negative association between vitamin B-12 and the risk of NTDs is also observed in other populations [50,51,52], and remains significant even in folic acid–fortified populations [53,54]. Transcobalamin II (TCN2) is a carrier protein that can bind vitamin B12. The mutation of TCN2 is associated with an increased NTD risk even when mothers are supplemented with folate [6]. CUBN is a gene that encodes the intestinal receptor responsible for the uptake of the vitamin B12–intrinsic factor complex. The mutation of CUBN is also associated with the risk of NTDs [40,55]. This genetic evidence also demonstrates the role of vitamin B-12 in the development of NTDs.
Besides folate and vitamin B-12, vitamin B-2 and B-6 are also important enzyme cofactors involved in the one-carbon metabolism. The conversion of 5,10-MTHF to 5-MTHF is catalyzed by MTHFR and depends on FADH2, the hydroquinone form of flavin adenine dinucleotide (FAD) (a derivative of vitamin B-2). Hustad et al., in a cross-sectional study, found that plasma vitamin B-2 was negatively associated with Hcy [56]. An intervention study by McNulty et al. in healthy adults found that vitamin B-2 supplementation lowered the concentration of Hcy only in subjects with a TT genotype of MTHFR 677C>T, but there was no effect in CC or CT genotypes [57]. In addition, vitamin B-2 can interact with folate to modulate Hcy concentrations. One intervention study found that folic acid supplementation (400 μg/day) had a greater Hcy-lowering effect in subjects with a high plasma level of vitamin B-2, and this effect was unrelated to MTHFR 677C>T polymorphism [58]. Several intervention studies have found that preconceptional multivitamin supplementation (including vitamin B-2 and several other vitamins such as folic acid, vitamin B-6, vitamin B-12, vitamin E, thiamin, vitamin A, vitamin D, nicotinamide, and ascorbic acid) could reduce the risk of NTDs [3,59]. Vitamin B-6 is the cofactor for betaine-Hcy methyltransferase (BHMT) (an enzyme that catalyzes the remethylation of Hcy to Met with betaine providing the methyl needed), cystathionine-beta-synthase (CBS) (an enzyme that catalyzes the reaction from Hcy to cystathionine) and cystathionine-gamma-lyase (CSE) (an enzyme that catalyzes the reaction from cystathionine to cysteine) (Figure 1). Vitamin B-6 deficiency is also associated with increased Hcy [46]. A French case-control study found that maternal plasma vitamin B-6 was negatively associated with NTD risk [33]. Thus, supplementation of vitamin B-2, vitamin B-6 and vitamin B-12 together with folic acid may have a better protective effect against NTDs than folic acid supplementation alone. However, this still needs to be demonstrated by well-designed intervention studies.
2.3. Potential Adverse Effects of B Vitamins
Despite the beneficial effect for NTD prevention, observational studies showed that very high doses of folic acid supplementation during conception also have several adverse effects. However, the evidence is still inconsistent. Observational studies showed that folic acid supplementation during conception was associated with the increased risk of wheezing in children through 18 months of age (dose was not reported) [60] and the increased risk of infant asthma (>72,000 μg∙day) [61]. Evidence from observational studies showed that folic acid supplementation was associated with the increased risk of infant clefts (dose was not reported) [62] and spontaneous preterm delivery (mean dose (interquartile range): 313 (167–558) μg/day) [63]. Valera-Gran et al. found that high folic acid supplementation for mothers during conception (>5000 μg/day) had a detrimental effect on the psychomotor development of children [64]. However, Tolarova et al. found that preconceptional supplementation of 10 mg/day folic acid plus multivitamins significantly reduced the risk of infant clefts [65]. One population-based cohort study in China found that preconceptional folic acid supplementation (400 μg/day) significantly reduced the risk of spontaneous preterm delivery [66]. McGarel et al. reported that folic acid supplementation had a beneficial effect on brain development and cognitive performance [67]. The FDA’s safe upper limit of folic acid is 1000 μg [68]. Folic acid is the synthetic form of folate. Before entry into the circulation system, folic acid undergoes reduction (by DHFR) and methylation to 5-MTHF, the circulating form of folic acid. However, when folic acid supplementation exceeds a certain dose, other transport mechanisms, such as passive diffusion, will complement the normal absorption mechanism, and thus unaltered folic acid enters the circulation system [69]. That is unmetabolized folic acid. One acute study found that folic acid supplementation of more than 800 μg/day can cause unmetabolized folic acid accumulation in the serum, but when the dose was no more than 400 μg/day, the unmetabolized folic acid in the serum was undetectable [69]. A prospective study of pregnant Canadian women found that unmetabolized folic acid was detectable in more than 90% of maternal and cord plasma samples, which may be a result of excess folic acid supplementation [70]. One RCT found that folic acid supplementation with a dose of 400 μg/day during conception had no significant influence on the unmetabolized folic acid concentration in maternal plasma or newborn cord blood plasma [71]. Therefore, a dose of 400 μg/day is safer than 1000 μg/day for folic acid supplementation.
Bailey reported that a higher vitamin B-6 intake (>1.85 mg/day) of mothers during the last six months of pregnancy was associated with a higher risk of childhood lymphoblastic leukemia [72]. However, one case-control study found that the per 1 mg increase of preconceptional vitamin B-6 intake from food and supplements was associated with a 11% decreased risk of childhood acute lymphoblastic leukemia. As suggested by US authorities, the level of vitamin B-6 with no observed adverse effect is set at 200 mg per day while the safe upper limit is at 100 mg per day [73]. Supplementation of vitamins B-2 and B-12 has not been associated with any adverse effects. Additionally, there is still insufficient evidence to set safe upper intake levels for vitamin B-2 and vitamin B-12 [74].
3. Choline, Betaine, One-Carbon Metabolism and NTDs
Choline is a nutrient associated with NTDs. Food sources of choline are principally those rich in lecithin (phosphatidylcholine) such as eggs and soy beans [75]. Betaine, which may be derived from choline, is found mostly in green leafy vegetables and beets (root vegetables). Choline can be synthesized in vivo by the methylation of phosphatidylethanolamine (PE) to phosphatidylcholine (PC) [76]. However, its biosynthesis is limited and dietary intake of choline is necessary [76]. Choline can be metabolized to betaine, catalyzed by choline oxidase (Figure 1). Similar to 5-MTHF, betaine can also provide the methyl needed in the remethylation of Hcy to Met, a reaction catalyzed by BHMT (Figure 1). The mutation of BHMT has been demonstrated to be associated with the risk of NTDs [6,19,31]. The ability of betaine supplementation to lower Hcy has been reported in the Netherlands [77]. Abnormal choline metabolism can lead to NTDs in the mouse [78]. An observational study found that preconceptional dietary intake of choline and betaine was negatively associated with the risk of fetal NTDs independent of folate intake in Americans [79].
However, choline and betaine intake can adversely affect serum lipid concentrations, such as increase total serum cholesterol, low density lipoprotein (LDL), high density lipoprotein (HDL) and triacylglycerol. Through the gut microbiome and trimethylamine production, they can also increase the risk of atherosclerotic vascular disease [80]. The safe upper level of choline may be 3500 mg/day for people with an age ≥19 years [81]. The safe upper level of betaine is unknown.
4. Other Dietary Factors Interact with the One-Carbon Metabolism to Influence the Development of NTDs
A case-control study has found that preconceptional tea consumption increases the risk of NTDs in Shanxi, China [82]. Similar findings have been observed in a case-control study in Atlanta [83]. In another case-control study conducted in the US, tea consumption was not associated with the risk of NTDs; however, when subjects were divided into subgroups according to the dose of folic acid intake, tea consumption was associated with an increased risk of NTDs in subjects with a folic acid intake >400 μg, and the authors suggested that tea consumption might interact with the folate metabolism pathway to influence the occurrence of NTDs [84]. The effects of tea consumption on the blood folate level are controversial [85,86]. One study has reported a lowering effect of tea consumption on blood folate [86]. A related case-control study found an association between the polymorphisms of catechol-O-methyltransferase (COMT) (encoding the enzyme that catalyzes the methylation of catecholamines with S-adenosylmethionine (SAM), a methyl donor) and the risk of NTDs in the Chinese: the mutant homozygotes of rs73,785 or rs4633 had a higher risk of NTDs than the wild homozygotes did, while the heterozygotes of rs4680 had a lower risk of NTDs than the wild homozygotes did, and the rs4680 genotype interacted with tea drinking to alter the risk of NTDs [87]. The effect of tea drinking on the development of NTDs can be explained by an inhibitory effect of tea catechins on dihydrofolatereductase [88]. Dihydrofolatereductase (DHFR) is an enzyme that catalyzes the conversion of dihydrofolate (DHF) to tetrahydrofolate (THF), the active form of folate. THF plays an important role in homocysteine metabolism and thymidine monophosphate (dTMP) synthesis. In summary, tea catechins inhibit the activity of DHFR, thus blocking DNA synthesis and homocysteine clearance, and this may help explain the association between tea consumption and the risk of NTDs (Figure 1).
One case-control study in Italians found that alcohol, low fruit and vegetable intake, and coffee were associated with an increased risk of NTDs [89]. Alcohol consumption is associated with lower blood folate as well as pyridoxal 5′-phosphate (the active form of vitamin B-6) and higher Hcy [90]. Animal studies showed that alcohol consumption can reduce the level of SAM (the major methyl donor) [91,92] and the activity of MTR (the enzyme catalyzing the remethylation of Hcy to Met) (Figure 1) [93]. Fruit and vegetables are rich in folate, and thus a low intake of these foods may be associated with inadequate folate intake (Figure 1). In pregnant Japanese women, it has been found that caffeine intake is associated with elevated Hcy only in subjects with a high intake of vitamin B-6 [94]. The positive association of caffeine with the level of Hcy and the risk of NTDs can be attributed to it having a similar chemical structure to theophylline, which can decrease pyridoxal 5′-phosphate (the active form of vitamin B-6) by acting as an inhibitor of pyridoxal kinase (Figure 1) [95,96].
In addition, n-3 PUFAs can lower Hcy [97,98,99,100], and upregulate the expression of several enzymes involved in the one-carbon metabolism, such as MTHFR, CBS, and CSE (Figure 1) [101]. Our unpublished case-control study in the Chinese has found that placental C18:3n-3, C20:5n-3 and C22:5n-3 are negatively associated with fetal NTD occurrence. The meta-analysis indicates that n-3 PUFAs combined with vitamin B supplementation have a greater Hcy-lowering effect than n-3 PUFAs alone [100]. However, there has been a concern that high-dose n-3 PUFA supplementation might induce bleeding [102]. However, there is little evidence to support this [103,104,105]. Previous studies showed that marine-derived n-3 PUFAs supplementation can promote LDL oxidation [106], but this is still controversial [107].
5. Perspective and Prospects
Although preconceptional folic acid supplementation has been recommended for decades, its overall ability to reduce the prevalence of NTDs is limited. Many women do not take a folic acid supplement until they are pregnant, which is too late for the effective prevention of NTDs. Long-term intake of folic acid–fortified foods should complement preconceptional folic acid supplementation. However, mandatory folic acid fortification is still not universally in place in Europe, Asia or Africa. However, folate-resistant NTDs exist, so there are other reasons for NTDs other than folate deficiency. In recent years, studies have indicated that the low intake and status of vitamin B-2, B-6, B-12, choline, betaine or n-3 PUFAs, and the consumption of alcohol, tea, or coffee are also associated with an increased risk of NTDs. Further cohort and intervention studies are needed to demonstrate whether multivitamin B (folate, vitamin B-2, B-6, B-12) combined with choline, betaine and n-3 PUFAs, or simply a biodiverse diet, has a better protective effect against NTDs than folic acid alone. In addition, genetic factors play an important role in the development of NTDs. There are many genetic variants involved in the one-carbon metabolism demonstrated to be associated with the risk of NTDs. Some variants can increase the risk of NTDs regardless of folic acid supplementation. Therefore, gene screening of women of childbearing age prior to pregnancy could enhance efforts to prevent NTDs.
6. Conclusions
Further cohort and intervention studies are needed to demonstrate whether multivitamin B (folate, vitamin B-2, B-6, B-12) combined with choline, betaine and n-3 PUFAs supplementation, or a biodiverse diet, has a better protective effect against NTDs than folic acid alone. Mandatory folic acid fortification and nutrition education, targeted at women in the reproductive age group, should be promoted and gene screening for women of childbearing age prior to pregnancy should be made available to prevent NTDs. These strategies would help decrease the burden of this oppressive health problem, especially in high-risk populations.
Acknowledgments
This study was funded by National Natural Science Foundation of China (NSFC, No. J20121077); by the Ph.D. Programs of the Foundation of Ministry of Education of China (J20130084); and by the National Basic Research Program of China (973 Program: 2015CB553600).
Author Contributions
D.L. and M.L.W. conceived this paper; K.L. wrote the paper; D.L. and M.L.W. revised the paper.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Mitchell, L.E. Epidemiology of Neural Tube Defects. In American Journal of Medical Genetics Part C: Seminars in Medical Genetics, 2005; Wiley Online Library: Hoboken, NJ, USA, 2005; pp. 88–94. [Google Scholar]
- Detrait, E.R.; George, T.M.; Etchevers, H.C.; Gilbert, J.R.; Vekemans, M.; Speer, M.C. Human neural tube defects: Developmental biology, epidemiology, and genetics. Neurotoxicol. Teratol. 2005, 27, 515–524. [Google Scholar] [CrossRef] [PubMed]
- Czeizel, A.E.; Dudas, I. Prevention of the first occurrence of neural-tube defects by periconceptional vitamin supplementation. N. Engl. J. Med. 1992, 327, 1832–1835. [Google Scholar] [CrossRef] [PubMed]
- Berry, R.J.; Li, Z.; Erickson, J.D.; Li, S.; Moore, C.A.; Wang, H.; Mulinare, J.; Zhao, P.; Wong, L.Y.; Gindler, J.; et al. Prevention of neural-tube defects with folic acid in China. China-U.S. Collaborative project for neural tube defect prevention. N. Engl. J. Med. 1999, 341, 1485–1490. [Google Scholar] [CrossRef] [PubMed]
- Crider, K.S.; Devine, O.; Hao, L.; Dowling, N.F.; Li, S.; Molloy, A.M.; Li, Z.; Zhu, J.; Berry, R.J. Population red blood cell folate concentrations for prevention of neural tube defects: Bayesian model. BMJ 2014, 29, g4554. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Qi, J.; Yu, X.; Zhu, J.; Zhang, L.; Ning, Q.; Luo, X. Investigations of single nucleotide polymorphisms in folate pathway genes in Chinese families with neural tube defects. J. Neurol. Sci. 2014, 337, 61–66. [Google Scholar] [CrossRef] [PubMed]
- MRC Vitamin Study Research Group. Prevention of neural tube defects: Results of the medical research council vitamin study. Mrc vitamin study research group. Lancet 1991, 338, 131–137. [Google Scholar]
- Daly, L.E.; Kirke, P.N.; Molloy, A.; Weir, D.G.; Scott, J.M. Folate levels and neural tube defects. Implications for prevention. JAMA 1995, 274, 1698–1702. [Google Scholar] [CrossRef] [PubMed]
- Centers for Disease Control and Prevention. Recommendations for the use of folic acid to reduce the number of cases of spina bifida and other neural tube defects. MMWR Recomm. Rep. 1992, 41, 1–7. [Google Scholar]
- Bestwick, J.P.; Huttly, W.J.; Morris, J.K.; Wald, N.J. Prevention of neural tube defects: A cross-sectional study of the uptake of folic acid supplementation in nearly half a million women. PLoS ONE 2014, 9. [Google Scholar] [CrossRef] [PubMed]
- Ren, A.G. Prevention of neural tube defects with folic acid: The Chinese experience. World J. Clin. Pediatr. 2015, 4, 41–44. [Google Scholar] [CrossRef] [PubMed]
- Arth, A.; Kancherla, V.; Pachon, H.; Zimmerman, S.; Johnson, Q.; Oakley, G.P., Jr. A 2015 global update on folic acid-preventable spina bifida and anencephaly. Birth Defects Res. A Clin. Mol. Teratol. 2016, 106, 520–529. [Google Scholar] [CrossRef] [PubMed]
- Santos, L.M.; Lecca, R.C.; Cortez-Escalante, J.J.; Sanchez, M.N.; Rodrigues, H.G. Prevention of neural tube defects by the fortification of flour with folic acid: A population-based retrospective study in brazil. Bull. World Health Organ. 2016, 94, 22–29. [Google Scholar] [CrossRef] [PubMed]
- Khoshnood, B.; Loane, M.; de Walle, H.; Arriola, L.; Addor, M.C.; Barisic, I.; Beres, J.; Bianchi, F.; Dias, C.; Draper, E.; et al. Long term trends in prevalence of neural tube defects in europe: Population based study. BMJ 2015, 24, h5949. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.; De Steur, H.; Chen, G.; Zhang, X.; Pei, L.; Gellynck, X.; Zheng, X. Effectiveness of folic acid fortified flour for prevention of neural tube defects in a high risk region. Nutrients 2016, 8, 152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Z.Z.; Zhang, J.T.; Liu, D.; Hao, Y.H.; Chang, B.M.; Xie, J.; Li, P.Z. Interaction between maternal 5,10-methylenetetrahydrofolate reductase c677t and methionine synthase a2756g gene variants to increase the risk of fetal neural tube defects in a Shanxi han population. Chin. Med. J. 2013, 126, 865–869. [Google Scholar] [PubMed]
- Morales de Machin, A.; Mendez, K.; Solis, E.; Borjas de Borjas, L.; Bracho, A.; Hernandez, M.L.; Negron, A.; Delgado, W.; Sanchez, Y. c677t polymorphism of the methylentetrahydrofolate reductase gene in mothers of children affected with neural tube defects. Investig. Clin. 2015, 56, 284–295. [Google Scholar]
- Van der Put, N.M.; Steegers-Theunissen, R.P.; Frosst, P.; Trijbels, F.J.; Eskes, T.K.; van den Heuvel, L.P.; Mariman, E.C.; den Heyer, M.; Rozen, R.; Blom, H.J. Mutated methylenetetrahydrofolate reductase as a risk factor for spina bifida. Lancet 1995, 346, 1070–1071. [Google Scholar] [CrossRef]
- Shaw, G.M.; Lu, W.; Zhu, H.; Yang, W.; Briggs, F.B.; Carmichael, S.L.; Barcellos, L.F.; Lammer, E.J.; Finnell, R.H. 118 snps of folate-related genes and risks of spina bifida and conotruncal heart defects. BMC Med. Genet. 2009, 10, 1471–2350. [Google Scholar] [CrossRef] [PubMed]
- Yan, L.; Zhao, L.; Long, Y.; Zou, P.; Ji, G.; Gu, A.; Zhao, P. Association of the maternal mthfr c677t polymorphism with susceptibility to neural tube defects in offsprings: Evidence from 25 case-control studies. PLoS ONE 2012, 7, 3. [Google Scholar] [CrossRef] [PubMed]
- Crider, K.S.; Zhu, J.H.; Hao, L.; Yang, Q.H.; Yang, T.P.; Gindler, J.; Maneval, D.R.; Quinlivan, E.P.; Li, Z.; Bailey, L.B.; et al. Mthfr 677c→t genotype is associated with folate and homocysteine concentrations in a large, population-based, double-blind trial of folic acid supplementation. Am. J. Clin. Nutr. 2011, 93, 1365–1372. [Google Scholar] [CrossRef] [PubMed]
- Tsang, B.L.; Devine, O.J.; Cordero, A.M.; Marchetta, C.M.; Mulinare, J.; Mersereau, P.; Guo, J.; Qi, Y.P.; Berry, R.J.; Rosenthal, J.; et al. Assessing the association between the methylenetetrahydrofolate reductase (mthfr) 677c>t polymorphism and blood folate concentrations: A systematic review and meta-analysis of trials and observational studies. Am. J. Clin. Nutr. 2015, 101, 1286–1294. [Google Scholar] [CrossRef] [PubMed]
- Van der Put, N.M.; Gabreels, F.; Stevens, E.M.; Smeitink, J.A.; Trijbels, F.J.; Eskes, T.K.; van den Heuvel, L.P.; Blom, H.J. A second common mutation in the methylenetetrahydrofolate reductase gene: An additional risk factor for neural-tube defects? Am. J. Hum. Genet. 1998, 62, 1044–1051. [Google Scholar] [CrossRef] [PubMed]
- De Marco, P.; Calevo, M.G.; Moroni, A.; Arata, L.; Merello, E.; Finnell, R.H.; Zhu, H.; Andreussi, L.; Cama, A.; Capra, V. Study of mthfr and ms polymorphisms as risk factors for ntd in the italian population. J. Hum. Genet. 2002, 47, 319–324. [Google Scholar] [CrossRef] [PubMed]
- Stegmann, K.; Ziegler, A.; Ngo, E.T.; Kohlschmidt, N.; Schroter, B.; Ermert, A.; Koch, M.C. Linkage disequilibrium of mthfr genotypes 677c/t-1298a/c in the german population and association studies in probands with neural tube defects(ntd). Am. J. Med. Genet. 1999, 87, 23–29. [Google Scholar] [CrossRef]
- Barber, R.; Shalat, S.; Hendricks, K.; Joggerst, B.; Larsen, R.; Suarez, L.; Finnell, R. Investigation of folate pathway gene polymorphisms and the incidence of neural tube defects in a texas hispanic population. Mol. Genet. Metab. 2000, 70, 45–52. [Google Scholar] [CrossRef] [PubMed]
- Volcik, K.A.; Blanton, S.H.; Tyerman, G.H.; Jong, S.T.; Rott, E.J.; Page, T.Z.; Romaine, N.K.; Northrup, H. Methylenetetrahydrofolate reductase and spina bifida: Evaluation of level of defect and maternal genotypic risk in hispanics. Am. J. Med. Genet. 2000, 95, 21–27. [Google Scholar] [CrossRef]
- Wang, Y.; Liu, Y.; Ji, W.; Qin, H.; Wu, H.; Xu, D.; Turtuohut, T.; Wang, Z. Variants in mthfr gene and neural tube defects susceptibility in China. Metab. Brain Dis. 2015, 30, 1017–1026. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Liu, Y.; Ji, W.; Qin, H.; Wu, H.; Xu, D.; Tukebai, T.; Wang, Z. Analysis of mtr and mtrr polymorphisms for neural tube defects risk association. Medicine 2015, 94, 1367. [Google Scholar] [CrossRef] [PubMed]
- Sliwerska, E.; Szpecht-Potocka, A. Mutations of MTHFR, MTR, MTRR genes as high risk factors for neural tube defects. Med. Wieku Rozwoj 2002, 6, 371–382. [Google Scholar] [PubMed]
- Boyles, A.L.; Billups, A.V.; Deak, K.L.; Siegel, D.G.; Mehltretter, L.; Slifer, S.H.; Bassuk, A.G.; Kessler, J.A.; Reed, M.C.; Nijhout, H.F.; et al. Neural tube defects and folate pathway genes: Family-based association tests of gene-gene and gene-environment interactions. Environ. Health Perspect. 2006, 114, 1547–1552. [Google Scholar] [CrossRef] [PubMed]
- Richter, B.; Stegmann, K.; Roper, B.; Boddeker, I.; Ngo, E.T.; Koch, M.C. Interaction of folate and homocysteine pathway genotypes evaluated in susceptibility to neural tube defects (ntd) in a german population. J. Hum. Genet. 2001, 46, 105–109. [Google Scholar] [CrossRef] [PubMed]
- Candito, M.; Rivet, R.; Herbeth, B.; Boisson, C.; Rudigoz, R.C.; Luton, D.; Journel, H.; Oury, J.F.; Roux, F.; Saura, R.; et al. Nutritional and genetic determinants of vitamin B and homocysteine metabolisms in neural tube defects: A multicenter case-control study. Am. J. Med. Genet. A 2008, 1, 1128–1133. [Google Scholar] [CrossRef] [PubMed]
- Yadav, U.; Kumar, P.; Yadav, S.K.; Mishra, O.P.; Rai, V. Polymorphisms in folate metabolism genes as maternal risk factor for neural tube defects: An updated meta-analysis. Metab. Brain Dis. 2015, 30, 7–24. [Google Scholar] [CrossRef] [PubMed]
- De Marco, P.; Merello, E.; Calevo, M.G.; Mascelli, S.; Raso, A.; Cama, A.; Capra, V. Evaluation of a methylenetetrahydrofolate-dehydrogenase 1958g>a polymorphism for neural tube defect risk. J. Hum. Genet. 2006, 51, 98–103. [Google Scholar] [CrossRef] [PubMed]
- Etheredge, A.J.; Finnell, R.H.; Carmichael, S.L.; Lammer, E.J.; Zhu, H.; Mitchell, L.E.; Shaw, G.M. Maternal and infant gene-folate interactions and the risk of neural tube defects. Am. J. Med. Genet. A 2012, 10, 17. [Google Scholar] [CrossRef] [PubMed]
- Jiang, J.; Zhang, Y.; Wei, L.; Sun, Z.; Liu, Z. Association between mthfd1 g1958a polymorphism and neural tube defects susceptibility: A meta-analysis. PLoS ONE 2014, 9. [Google Scholar] [CrossRef] [PubMed]
- Meng, J.; Han, L.; Zhuang, B. Association between mthfd1 polymorphisms and neural tube defect susceptibility. J. Neurol. Sci. 2015, 348, 188–194. [Google Scholar] [CrossRef] [PubMed]
- Parle-McDermott, A.; Pangilinan, F.; O’Brien, K.K.; Mills, J.L.; Magee, A.M.; Troendle, J.; Sutton, M.; Scott, J.M.; Kirke, P.N.; Molloy, A.M.; et al. A common variant in mthfd1l is associated with neural tube defects and mrna splicing efficiency. Hum. Mutat. 2009, 30, 1650–1656. [Google Scholar] [CrossRef] [PubMed]
- Pangilinan, F.; Molloy, A.M.; Mills, J.L.; Troendle, J.F.; Parle-McDermott, A.; Signore, C.; O’Leary, V.B.; Chines, P.; Seay, J.M.; Geiler-Samerotte, K.; et al. Evaluation of common genetic variants in 82 candidate genes as risk factors for neural tube defects. BMC Med. Genet. 2012, 13, 62. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Bao, Y.; Lu, X.; Wu, L.; Zhang, T.; Guo, J.; Yang, J. Polymorphisms in mthfd1 gene and susceptibility to neural tube defects: A case-control study in a Chinese han population with relatively low folate levels. Med. Sci. Monit. 2015, 21, 2630–2637. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.; Xie, H.; Wang, J.; Zhao, H.; Wang, F.; Liu, C.; Wang, L.; Lu, X.; Bao, Y.; Zou, J.; et al. The maternal folate hydrolase gene polymorphism is associated with neural tube defects in a high-risk Chinese population. Genes Nutr. 2013, 8, 191–197. [Google Scholar] [CrossRef] [PubMed]
- Pei, L.J.; Zhu, H.P.; Li, Z.W.; Zhang, W.; Ren, A.G.; Zhu, J.H.; Li, Z. Interaction between maternal periconceptional supplementation of folic acid and reduced folate carrier gene polymorphism of neural tube defects. Zhonghua Yi Xue Yi Chuan Xue Za Zhi 2005, 22, 284–287. [Google Scholar] [PubMed]
- Zhang, T.; Lou, J.; Zhong, R.; Wu, J.; Zou, L.; Sun, Y.; Lu, X.; Liu, L.; Miao, X.; Xiong, G. Genetic variants in the folate pathway and the risk of neural tube defects: A meta-analysis of the published literature. PLoS ONE 2013, 8, e59570. [Google Scholar] [CrossRef] [PubMed]
- Piao, W.; Guo, J.; Bao, Y.; Wang, F.; Zhang, T.; Huo, J.; Zhang, K. Analysis of polymorphisms of genes associated with folate-mediated one-carbon metabolism and neural tube defects in Chinese han population. Birth Defects Res. A Clin. Mol. Teratol. 2016, 106, 232–239. [Google Scholar] [CrossRef] [PubMed]
- Hao, L.; Ma, J.; Zhu, J.; Stampfer, M.J.; Tian, Y.; Willett, W.C.; Li, Z. High prevalence of hyperhomocysteinemia in Chinese adults is associated with low folate, vitamin B-12, and vitamin B-6 status. J. Nutr. 2007, 137, 407–413. [Google Scholar] [PubMed]
- Gu, Q.; Li, Y.; Cui, Z.L.; Luo, X.P. Homocysteine, folate, vitamin B12 and B6 in mothers of children with neural tube defects in Xinjiang, China. Acta Paediatr. 2012, 101, 1651–2227. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Xin, R.; Gu, X.; Wang, F.; Pei, L.; Lin, L.; Chen, G.; Wu, J.; Zheng, X. Maternal serum vitamin B12, folate and homocysteine and the risk of neural tube defects in the offspring in a high-risk area of China. Public Health Nutr. 2009, 12, 680–686. [Google Scholar] [CrossRef] [PubMed]
- Dang, S.; Yan, H.; Zeng, L.; Wang, Q.; Li, Q.; Xiao, S.; Fan, X. The status of vitamin B12 and folate among Chinese women: A population-based cross-sectional study in northwest China. PLoS ONE 2014, 9. [Google Scholar] [CrossRef] [PubMed]
- Gaber, K.R.; Farag, M.K.; Soliman, S.E.; El-Bassyouni, H.T.; El-Kamah, G. Maternal vitamin B12 and the risk of fetal neural tube defects in egyptian patients. Clin. Lab. 2007, 53, 69–75. [Google Scholar] [PubMed]
- Molloy, A.M.; Kirke, P.N.; Troendle, J.F.; Burke, H.; Sutton, M.; Brody, L.C.; Scott, J.M.; Mills, J.L. Maternal vitamin B12 status and risk of neural tube defects in a population with high neural tube defect prevalence and no folic acid fortification. Pediatrics 2009, 123, 917–923. [Google Scholar] [CrossRef] [PubMed]
- Nasri, K.; Ben Fradj, M.K.; Touati, A.; Aloui, M.; Ben Jemaa, N.; Masmoudi, A.; Elmay, M.V.; Omar, S.; Feki, M.; Kaabechi, N.; et al. Association of maternal homocysteine and vitamins status with the risk of neural tube defects in tunisia: A case-control study. Birth Defects Res. A Clin. Mol. Teratol. 2015, 103, 1011–1020. [Google Scholar] [CrossRef] [PubMed]
- Ray, J.G.; Wyatt, P.R.; Thompson, M.D.; Vermeulen, M.J.; Meier, C.; Wong, P.Y.; Farrell, S.A.; Cole, D.E. Vitamin B12 and the risk of neural tube defects in a folic-acid-fortified population. Epidemiology 2007, 18, 362–366. [Google Scholar] [CrossRef] [PubMed]
- Thompson, M.D.; Cole, D.E.; Ray, J.G. Vitamin B-12 and neural tube defects: The canadian experience. Am. J. Clin. Nutr. 2009, 89, 30. [Google Scholar] [CrossRef] [PubMed]
- Franke, B.; Vermeulen, S.H.; Steegers-Theunissen, R.P.; Coenen, M.J.; Schijvenaars, M.M.; Scheffer, H.; den Heijer, M.; Blom, H.J. An association study of 45 folate-related genes in spina bifida: Involvement of cubilin (cubn) and trna aspartic acid methyltransferase 1 (trdmt1). Birth Defects Res. A Clin. Mol. Teratol. 2009, 85, 216–226. [Google Scholar] [CrossRef] [PubMed]
- Hustad, S.; Ueland, P.M.; Vollset, S.E.; Zhang, Y.; Bjorke-Monsen, A.L.; Schneede, J. Riboflavin as a determinant of plasma total homocysteine: Effect modification by the methylenetetrahydrofolate reductase c677t polymorphism. Clin. Chem. 2000, 46, 1065–1071. [Google Scholar] [PubMed]
- McNulty, H.; Dowey le, R.C.; Strain, J.J.; Dunne, A.; Ward, M.; Molloy, A.M.; McAnena, L.B.; Hughes, J.P.; Hannon-Fletcher, M.; Scott, J.M. Riboflavin lowers homocysteine in individuals homozygous for the mthfr 677c→t polymorphism. Circulation 2006, 113, 74–80. [Google Scholar] [CrossRef] [PubMed]
- Moat, S.J.; Ashfield-Watt, P.A.; Powers, H.J.; Newcombe, R.G.; McDowell, I.F. Effect of riboflavin status on the homocysteine-lowering effect of folate in relation to the mthfr (c677t) genotype. Clin. Chem. 2003, 49, 295–302. [Google Scholar] [CrossRef] [PubMed]
- Smithells, R.W.; Sheppard, S.; Schorah, C.J.; Seller, M.J.; Nevin, N.C.; Harris, R.; Read, A.P.; Fielding, D.W. Apparent prevention of neural tube defects by periconceptional vitamin supplementation. Arch. Dis. Child. 1981, 56, 911–918. [Google Scholar] [CrossRef] [PubMed]
- Haberg, S.E.; London, S.J.; Stigum, H.; Nafstad, P.; Nystad, W. Folic acid supplements in pregnancy and early childhood respiratory health. Arch. Dis. Child. 2009, 94, 180–184. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Jiang, L.; Bi, M.; Jia, X.; Wang, Y.; He, C.; Yao, Y.; Wang, J.; Wang, Z. High dose of maternal folic acid supplementation is associated to infant asthma. Food Chem. Toxicol. 2015, 75, 88–93. [Google Scholar] [CrossRef] [PubMed]
- Rozendaal, A.M.; van Essen, A.J.; te Meerman, G.J.; Bakker, M.K.; van der Biezen, J.J.; Goorhuis-Brouwer, S.M.; Vermeij-Keers, C.; de Walle, H.E. Periconceptional folic acid associated with an increased risk of oral clefts relative to non-folate related malformations in the northern netherlands: A population based case-control study. Eur. J. Epidemiol. 2013, 28, 875–887. [Google Scholar] [CrossRef] [PubMed]
- Sengpiel, V.; Bacelis, J.; Myhre, R.; Myking, S.; Devold Pay, A.S.; Haugen, M.; Brantsaeter, A.L.; Meltzer, H.M.; Nilsen, R.M.; Magnus, P.; et al. Folic acid supplementation, dietary folate intake during pregnancy and risk for spontaneous preterm delivery: A prospective observational cohort study. BMC Pregnancy Childbirth 2014, 14, 014–0375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valera-Gran, D.; Garcia de la Hera, M.; Navarrete-Munoz, E.M.; Fernandez-Somoano, A.; Tardon, A.; Julvez, J.; Forns, J.; Lertxundi, N.; Ibarluzea, J.M.; Murcia, M.; et al. Folic acid supplements during pregnancy and child psychomotor development after the first year of life. JAMA Pediatr. 2014, 168, 3. [Google Scholar] [CrossRef] [PubMed]
- Tolarova, M.; Harris, J. Reduced recurrence of orofacial clefts after periconceptional supplementation with high-dose folic acid and multivitamins. Teratology 1995, 51, 71–78. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Ye, R.; Zhang, L.; Li, H.; Liu, J.; Ren, A. Periconceptional folic acid supplementation and the risk of preterm births in China: A large prospective cohort study. Int. J. Epidemiol. 2014, 43, 1132–1139. [Google Scholar] [CrossRef] [PubMed]
- McGarel, C.; Pentieva, K.; Strain, J.J.; McNulty, H. Emerging roles for folate and related B-vitamins in brain health across the lifecycle. Proc. Nutr. Soc. 2015, 74, 46–55. [Google Scholar] [CrossRef] [PubMed]
- Quinlivan, E.P.; Gregory, J.F., 3rd. Effect of food fortification on folic acid intake in the united states. Am. J. Clin. Nutr. 2003, 77, 221–225. [Google Scholar] [PubMed]
- Kelly, P.; McPartlin, J.; Goggins, M.; Weir, D.G.; Scott, J.M. Unmetabolized folic acid in serum: Acute studies in subjects consuming fortified food and supplements. Am. J. Clin. Nutr. 1997, 65, 1790–1795. [Google Scholar] [PubMed]
- Plumptre, L.; Masih, S.P.; Ly, A.; Aufreiter, S.; Sohn, K.J.; Croxford, R.; Lausman, A.Y.; Berger, H.; O’Connor, D.L.; Kim, Y.I. High concentrations of folate and unmetabolized folic acid in a cohort of pregnant canadian women and umbilical cord blood. Am. J. Clin. Nutr. 2015, 102, 848–857. [Google Scholar] [CrossRef] [PubMed]
- Pentieva, K.; Selhub, J.; Paul, L.; Molloy, A.M.; McNulty, B.; Ward, M.; Marshall, B.; Dornan, J.; Reilly, R.; Parle-McDermott, A.; et al. Evidence from a randomized trial that exposure to supplemental folic acid at recommended levels during pregnancy does not lead to increased unmetabolized folic acid concentrations in maternal or cord blood. J. Nutr. 2016, 146, 494–500. [Google Scholar] [CrossRef] [PubMed]
- Bailey, H.D.; Miller, M.; Langridge, A.; de Klerk, N.H.; van Bockxmeer, F.M.; Attia, J.; Scott, R.J.; Armstrong, B.K.; Milne, E. Maternal dietary intake of folate and vitamins B6 and B12 during pregnancy and the risk of childhood acute lymphoblastic leukemia. Nutr. Cancer 2012, 64, 1122–1130. [Google Scholar] [CrossRef] [PubMed]
- Katan, M.B. How much vitamin B6 is toxic? Ned. Tijdschr. Geneeskd. 2005, 149, 2545–2546. [Google Scholar] [PubMed]
- Rogovik, A.L.; Vohra, S.; Goldman, R.D. Safety considerations and potential interactions of vitamins: Should vitamins be considered drugs? Ann. Pharmacother. 2010, 44, 311–324. [Google Scholar] [CrossRef] [PubMed]
- Chu, D.-M.; Wahlqvist, M.L.; Chang, H.-Y.; Yeh, N.-H. Choline and betaine food sources and intakes in taiwanese. Asia Pac. J. Clin. Nutr. 2012, 21, 547–557. [Google Scholar] [PubMed]
- Zeisel, S.H.; da Costa, K.A. Choline: An essential nutrient for public health. Nutr. Rev. 2009, 67, 615–623. [Google Scholar] [CrossRef] [PubMed]
- Steenge, G.R.; Verhoef, P.; Katan, M.B. Betaine supplementation lowers plasma homocysteine in healthy men and women. J. Nutr. 2003, 133, 1291–1295. [Google Scholar] [PubMed]
- Fisher, M.C.; Zeisel, S.H.; Mar, M.H.; Sadler, T.W. Perturbations in choline metabolism cause neural tube defects in mouse embryos in vitro. FASEB J. 2002, 16, 619–621. [Google Scholar] [CrossRef] [PubMed]
- Shaw, G.M.; Carmichael, S.L.; Yang, W.; Selvin, S.; Schaffer, D.M. Periconceptional dietary intake of choline and betaine and neural tube defects in offspring. Am. J. Epidemiol. 2004, 160, 102–109. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Klipfell, E.; Bennett, B.J.; Koeth, R.; Levison, B.S.; Dugar, B.; Feldstein, A.E.; Britt, E.B.; Fu, X.; Chung, Y.M.; et al. Gut flora metabolism of phosphatidylcholine promotes cardiovascular disease. Nature 2011, 472, 57–63. [Google Scholar] [CrossRef] [PubMed]
- Sanders, L.M.; Zeisel, S.H. Choline: Dietary requirements and role in brain development. Nutr. Today 2007, 42, 181–186. [Google Scholar] [CrossRef] [PubMed]
- Ye, R.; Ren, A.; Zhang, L.; Li, Z.; Liu, J.; Pei, L.; Zheng, X. Tea drinking as a risk factor for neural tube defects in northern China. Epidemiology 2011, 22, 491–496. [Google Scholar] [CrossRef] [PubMed]
- Correa, A.; Stolley, A.; Liu, Y. Prenatal tea consumption and risks of anencephaly and spina bifida. Ann. Epidemiol. 2000, 10, 476–477. [Google Scholar] [CrossRef]
- Yazdy, M.M.; Tinker, S.C.; Mitchell, A.A.; Demmer, L.A.; Werler, M.M. Maternal tea consumption during early pregnancy and the risk of spina bifida. Birth Defects Res. Part A Clin. Mol. Teratol. 2012, 94, 756–761. [Google Scholar] [CrossRef] [PubMed]
- Augustin, K.; Frank, J.; Augustin, S.; Langguth, P.; Ohrvik, V.; Witthoft, C.M.; Rimbach, G.; Wolffram, S. Greeen tea extracts lower serum folates in rats at very high dietary concentrations only and do not affect plasma folates in a human pilot study. J. Physiol. Pharmacol. 2009, 60, 103–108. [Google Scholar] [PubMed]
- Shiraishi, M.; Haruna, M.; Matsuzaki, M.; Ota, E.; Murayama, R.; Murashima, S. Association between the serum folate levels and tea consumption during pregnancy. Biosci. Trends 2010, 4, 225–230. [Google Scholar] [PubMed]
- Liu, J.; Wang, L.; Fu, Y.; Li, Z.; Zhang, Y.; Zhang, L.; Jin, L.; Ye, R.; Ren, A. Association between maternal comt gene polymorphisms and fetal neural tube defects risk in a Chinese population. Birth Defects Res. A Clin. Mol. Teratol. 2014, 100, 22–29. [Google Scholar] [CrossRef] [PubMed]
- Navarro-Peran, E.; Cabezas-Herrera, J.; Campo, L.S.; Rodriguez-Lopez, J.N. Effects of folate cycle disruption by the green tea polyphenol epigallocatechin-3-gallate. Int. J. Biochem. Cell Biol. 2007, 39, 2215–2225. [Google Scholar] [CrossRef] [PubMed]
- De Marco, P.; Merello, E.; Calevo, M.G.; Mascelli, S.; Pastorino, D.; Crocetti, L.; De Biasio, P.; Piatelli, G.; Cama, A.; Capra, V. Maternal periconceptional factors affect the risk of spina bifida-affected pregnancies: An italian case-control study. Childs Nerv. Syst. 2011, 27, 1073–1081. [Google Scholar] [CrossRef] [PubMed]
- Cravo, M.L.; Gloria, L.M.; Selhub, J.; Nadeau, M.R.; Camilo, M.E.; Resende, M.P.; Cardoso, J.N.; Leitao, C.N.; Mira, F.C. Hyperhomocysteinemia in chronic alcoholism: Correlation with folate, vitamin B-12, and vitamin B-6 status. Am. J. Clin. Nutr. 1996, 63, 220–224. [Google Scholar] [PubMed]
- Stickel, F.; Choi, S.W.; Kim, Y.I.; Bagley, P.J.; Seitz, H.K.; Russell, R.M.; Selhub, J.; Mason, J.B. Effect of chronic alcohol consumption on total plasma homocysteine level in rats. Alcohol. Clin. Exp. Res. 2000, 24, 259–264. [Google Scholar] [CrossRef] [PubMed]
- Murillo-Fuentes, M.L.; Artillo, R.; Ubeda, N.; Varela-Moreiras, G.; Murillo, M.L.; Carreras, O. Hepatic s-adenosylmethionine after maternal alcohol exposure on offspring rats. Addict. Biol. 2005, 10, 139–144. [Google Scholar] [CrossRef] [PubMed]
- Barak, A.J.; Beckenhauer, H.C.; Junnila, M.; Tuma, D.J. Dietary betaine promotes generation of hepatic s-adenosylmethionine and protects the liver from ethanol-induced fatty infiltration. Alcohol. Clin. Exp. Res. 1993, 17, 552–555. [Google Scholar] [CrossRef] [PubMed]
- Shiraishi, M.; Haruna, M.; Matsuzaki, M.; Ota, E.; Murayama, R.; Sasaki, S.; Yeo, S.; Murashima, S. Relationship between plasma total homocysteine level and dietary caffeine and vitamin B6 intakes in pregnant women. Nurs. Health Sci. 2014, 16, 164–170. [Google Scholar] [CrossRef] [PubMed]
- Ubbink, J.B.; Delport, R.; Becker, P.J.; Bissbort, S. Evidence of a theophylline-induced vitamin B6 deficiency caused by noncompetitive inhibition of pyridoxal kinase. J. Lab. Clin. Med. 1989, 113, 15–22. [Google Scholar] [PubMed]
- Ubbink, J.B.; Delport, R.; Bissbort, S.; Vermaak, W.J.; Becker, P.J. Relationship between vitamin B-6 status and elevated pyridoxal kinase levels induced by theophylline therapy in humans. J. Nutr. 1990, 120, 1352–1359. [Google Scholar] [PubMed]
- Huang, T.; Li, K.; Asimi, S.; Chen, Q.; Li, D. Effect of vitamin B-12 and n-3 polyunsaturated fatty acids on plasma homocysteine, ferritin, c-reaction protein, and other cardiovascular risk factors: A randomized controlled trial. Asia Pac. J. Clin. Nutr. 2015, 24, 403–411. [Google Scholar] [PubMed]
- Huang, T.; Yu, X.; Shou, T.; Wahlqvist, M.L.; Li, D. Associations of plasma phospholipid fatty acids with plasma homocysteine in Chinese vegetarians. Br. J. Nutr. 2013, 109, 1688–1694. [Google Scholar] [CrossRef] [PubMed]
- Huang, T.; Asimi, S.; Lou, D.; Li, D. Plasma phospholipid polyunsaturated fatty acids and homocysteine in Chinese type 2 diabetes patients. Asia Pac. J. Clin. Nutr. 2012, 21, 394–399. [Google Scholar] [PubMed]
- Dawson, S.L.; Bowe, S.J.; Crowe, T.C. A combination of omega-3 fatty acids, folic acid and b-group vitamins is superior at lowering homocysteine than omega-3 alone: A meta-analysis. Nutr. Res. 2016, 36, 499–508. [Google Scholar] [CrossRef] [PubMed]
- Huang, T.; Hu, X.; Khan, N.; Yang, J.; Li, D. Effect of polyunsaturated fatty acids on homocysteine metabolism through regulating the gene expressions involved in methionine metabolism. Sci. World J. 2013, 23, 931626. [Google Scholar] [CrossRef] [PubMed]
- Detopoulou, P.; Papamikos, V. Gastrointestinal bleeding after high intake of omega-3 fatty acids, cortisone and antibiotic therapy: A case study. Int. J. Sport Nutr. Exerc. Metab. 2014, 24, 253–257. [Google Scholar] [CrossRef] [PubMed]
- Wachira, J.K.; Larson, M.K.; Harris, W.S. N-3 fatty acids affect haemostasis but do not increase the risk of bleeding: Clinical observations and mechanistic insights. Br. J. Nutr. 2014, 111, 1652–1662. [Google Scholar] [CrossRef] [PubMed]
- Meredith, D.S.; Kepler, C.K.; Huang, R.C.; Hirsch, B.; Nguyen, J.; Farmer, J.C.; Girardi, F.P.; O’Leary, P.F.; Cammisa, F.P. The effect of omega-3 fatty-acid supplements on perioperative bleeding following posterior spinal arthrodesis. Eur. Spine J. 2012, 21, 2659–2663. [Google Scholar] [CrossRef] [PubMed]
- Salisbury, A.C.; Harris, W.S.; Amin, A.P.; Reid, K.J.; O’Keefe, J.H., Jr.; Spertus, J.A. Relation between red blood cell omega-3 fatty acid index and bleeding during acute myocardial infarction. Am. J. Cardiol. 2012, 109, 13–18. [Google Scholar] [CrossRef] [PubMed]
- Finnegan, Y.E.; Minihane, A.M.; Leigh-Firbank, E.C.; Kew, S.; Meijer, G.W.; Muggli, R.; Calder, P.C.; Williams, C.M. Plant- and marine-derived n-3 polyunsaturated fatty acids have differential effects on fasting and postprandial blood lipid concentrations and on the susceptibility of ldl to oxidative modification in moderately hyperlipidemic subjects. Am. J. Clin. Nutr. 2003, 77, 783–795. [Google Scholar] [PubMed]
- Mori, T.A.; Woodman, R.J.; Burke, V.; Puddey, I.B.; Croft, K.D.; Beilin, L.J. Effect of eicosapentaenoic acid and docosahexaenoic acid on oxidative stress and inflammatory markers in treated-hypertensive type 2 diabetic subjects. Free Radic. Biol. Med. 2003, 35, 772–781. [Google Scholar] [CrossRef]
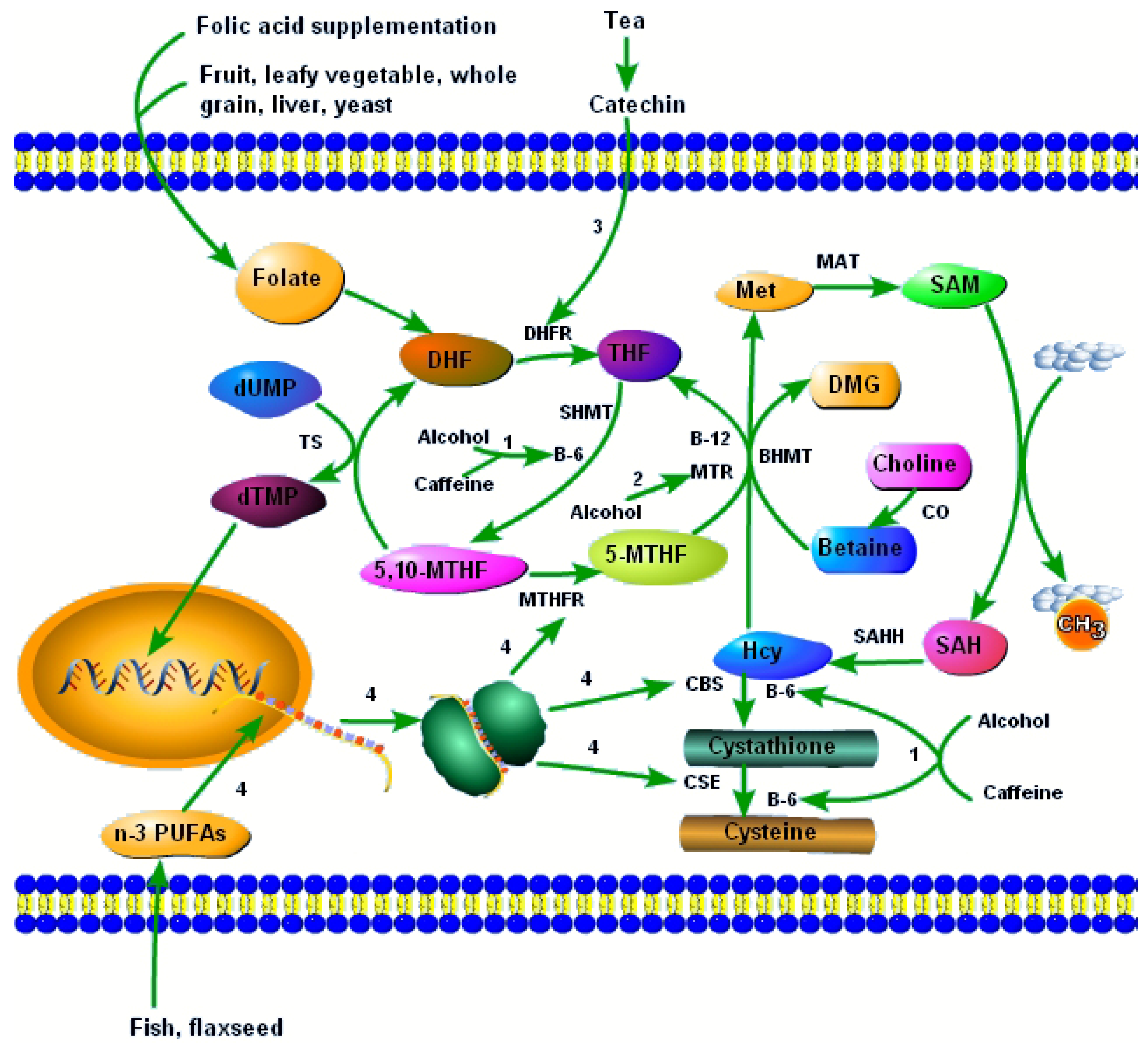
Figure 1.
B vitamins and other dietary factors interact with the one-carbon metabolism to influence the development of NTDs. 1, alcohol and caffeine lower vitamin B-6, and thus disturb vitamin B-6–dependent one-carbon metabolism pathways; 2, alcohol reduces the activity of MTR, leading to increased Hcy and reduced SAM; 3, catechins in tea reduce the activity of DHFR, and hinder the synthesis of THF; 4, n-3 PUFAs increase the mRNA expression of enzymes involved in one-carbon metabolism, such as MTHFR, CBS, and CSE. Abbreviations: DHF, dihydrofolate; THF, tetrahydrofolate; 5,10-MTHF, 5,10-methylenetetrahydrofolate; 5-MTHF, 5-methyltetrahydrofolate; dUMP, deoxyuridine monophosphate; dTMP, thymidinemonophosphate; Hcy, homocysteine; Met, methionine; SAM, S-adenosylmethionine; SAH, S-adenosylhomocysteine; DMG, dimethylglycine; DHFR, dihydrofolate reductase; SHMT, serine hydroxymethyltransferase; MTHFR, methylenetetrahydrofolate reductase; MTR, 5-methyltetrahydrofolate-homocysteine methyltransferase; TS, thymidylate synthase; BHMT, betaine-homocysteine methyltransferase; MAT, methionine adenosyltransferase; SAHH, S-adenosylhomocysteine hydrolase; CBS, cystathionine-beta-synthase; CSE, cystathionine-gamma-lyase.
Figure 1.
B vitamins and other dietary factors interact with the one-carbon metabolism to influence the development of NTDs. 1, alcohol and caffeine lower vitamin B-6, and thus disturb vitamin B-6–dependent one-carbon metabolism pathways; 2, alcohol reduces the activity of MTR, leading to increased Hcy and reduced SAM; 3, catechins in tea reduce the activity of DHFR, and hinder the synthesis of THF; 4, n-3 PUFAs increase the mRNA expression of enzymes involved in one-carbon metabolism, such as MTHFR, CBS, and CSE. Abbreviations: DHF, dihydrofolate; THF, tetrahydrofolate; 5,10-MTHF, 5,10-methylenetetrahydrofolate; 5-MTHF, 5-methyltetrahydrofolate; dUMP, deoxyuridine monophosphate; dTMP, thymidinemonophosphate; Hcy, homocysteine; Met, methionine; SAM, S-adenosylmethionine; SAH, S-adenosylhomocysteine; DMG, dimethylglycine; DHFR, dihydrofolate reductase; SHMT, serine hydroxymethyltransferase; MTHFR, methylenetetrahydrofolate reductase; MTR, 5-methyltetrahydrofolate-homocysteine methyltransferase; TS, thymidylate synthase; BHMT, betaine-homocysteine methyltransferase; MAT, methionine adenosyltransferase; SAHH, S-adenosylhomocysteine hydrolase; CBS, cystathionine-beta-synthase; CSE, cystathionine-gamma-lyase.

© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Li, K.; Wahlqvist, M.L.; Li, D. Nutrition, One-Carbon Metabolism and Neural Tube Defects: A Review. Nutrients 2016, 8, 741. https://doi.org/10.3390/nu8110741
AMA Style
Li K, Wahlqvist ML, Li D. Nutrition, One-Carbon Metabolism and Neural Tube Defects: A Review. Nutrients. 2016; 8(11):741. https://doi.org/10.3390/nu8110741
Chicago/Turabian StyleLi, Kelei, Mark L. Wahlqvist, and Duo Li. 2016. "Nutrition, One-Carbon Metabolism and Neural Tube Defects: A Review" Nutrients 8, no. 11: 741. https://doi.org/10.3390/nu8110741
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.