Interactions between Obesity-Related Copy Number Variants and Dietary Behaviors in Childhood Obesity

Abstract
:1. Introduction
2. Experimental Section
2.1. Subjects
2.2. Anthropometric Measurements and Epidemiologic Investigation
2.3. CNVs Selection

{kind=link}
{kind=link}
| CNV_ID | Locus | Position * | Size (bp) | Overlap Genes |
|---|---|---|---|---|
| CNV_1 | 11q11 | chr11: 55,130,596–55,210,165 | 79,569 | OR4P1P OR4V1P OR4P4 OR4S2 OR4C6 |
| CNV_2 | 5q13.2 | chr5: 68,903,038–70,343,313 | 1,440,275 | SMN1 SMN2 SERF1A SMA4 SMA5 GUSBP1 LOC643367 LOC643784 LOC653080 LOC653188 GTF2H2B LOC653391 LOC728488 SERF1B LOC728499 LOC728506 LOC728519 LOC728526 LOC728535 LOC728555 LOC100093625 LOC100132218 LOC100133280 |
| CNV_4 | 10q11.22(1) | chr10: 46,338,178–46,812,351 | 474,173 | GLUDP2 PPYR1 GPRIN2 SYT15 BMS1P2 LOC642826 LOC643650 ANXA8L1 CTGLF7 LOC728643 LOC728657 LOC100132646 FAM25B LOC100133189 |
| CNV_10 | 10q11.22(2) | chr10: 47,011,183–47,145,122 | 133,939 | LOC340844 LOC728684 |
| CNV_12 | 6q14.1 | chr6: 81,340,896–81,346,266 | 5370 | None |
| CNV_13 | 4q25 | chr4: 108,285,188–108,293,270 | 8082 | None |
2.4. CNVs Genotyping by AccuCopy™ Technique
2.5. Statistical Analysis
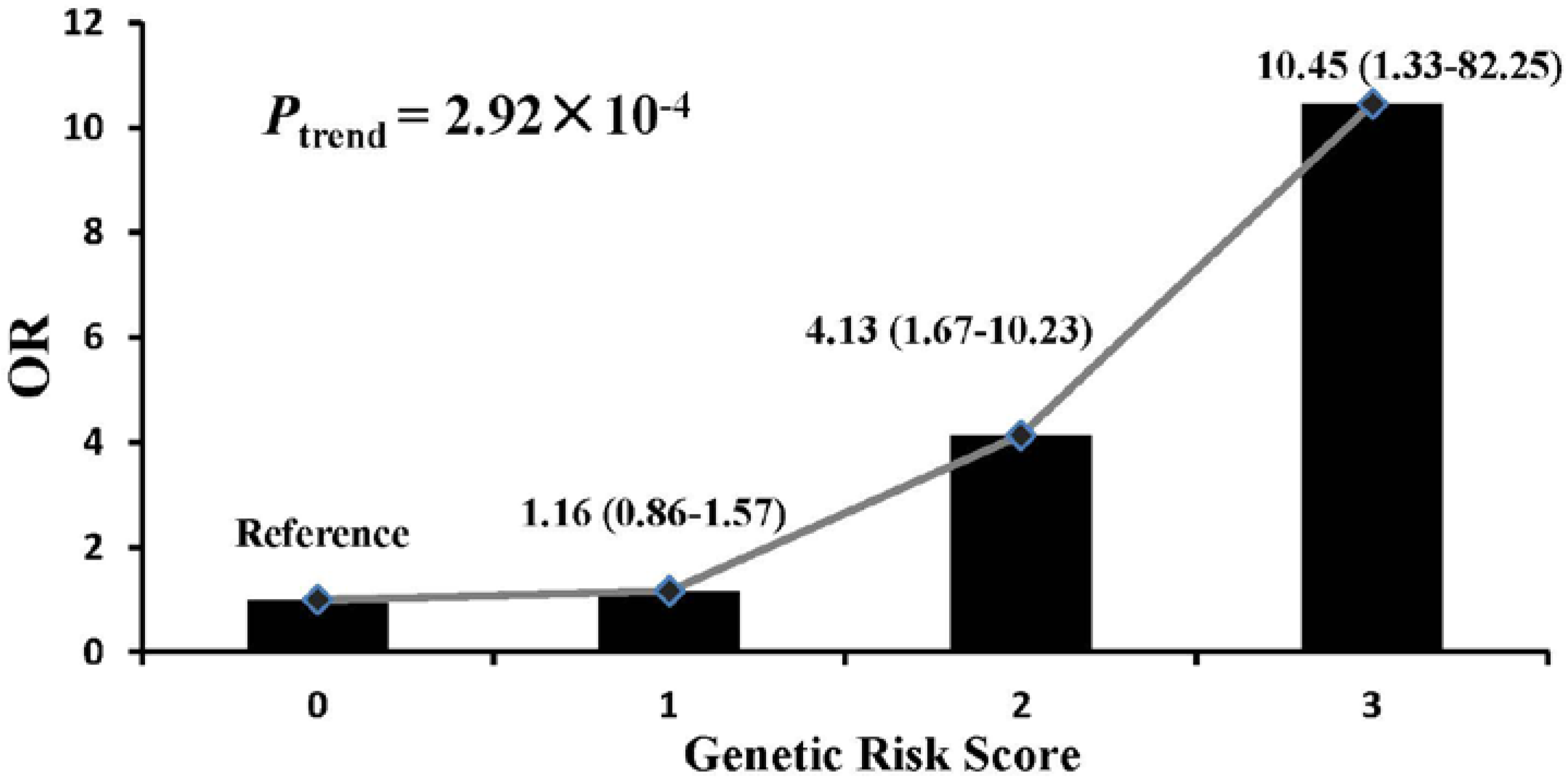
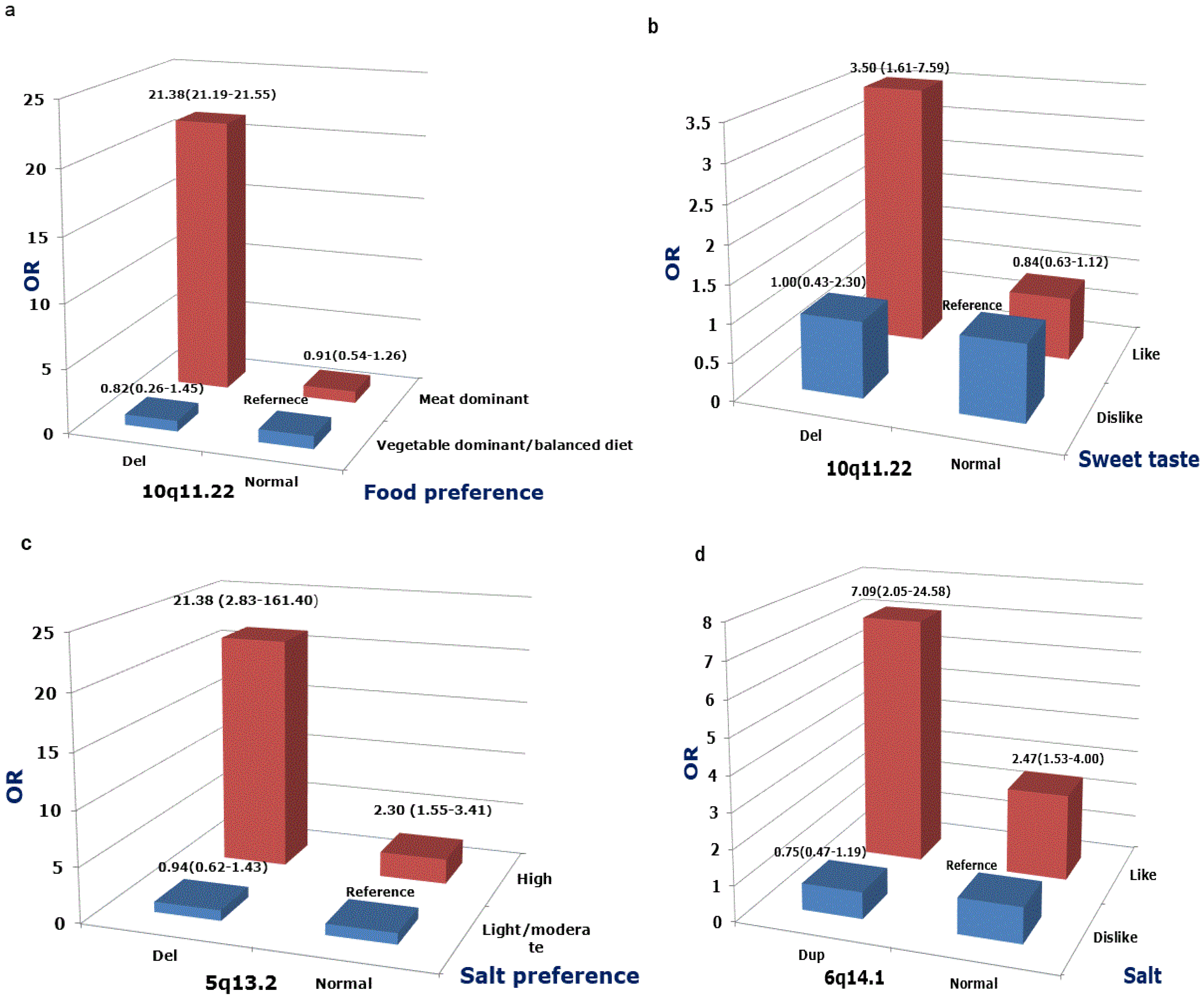
3. Results
| Characteristics | Obese Children (n = 529) | Non-Obese Children (n = 498) | p-Value |
|---|---|---|---|
| Age (years) | 11.7 ± 2.9 | 11.7 ± 2.9 | 0.85 |
| Gender (M/F) | 345/184 | 326/172 | 0.93 |
| BMI (Kg m−2) | 27.3 ± 4.2 | 17.6 ± 2.3 | <0.001 |
| BMI z-score | 2.71 ± 0.71 | −0.17 ± 0.88 | <0.001 |
| WHtR | 0.54 ± 0.06 | 0.41 ± 0.03 | <0.001 |
| WHR | 0.89 ± 0.08 | 0.81 ± 0.06 | <0.001 |
| WC (cm) | 82.8 ± 13.4 | 60.4 ± 8.1 | <0.001 |
| HC (cm) | 96.0 (82.0–102.0) | 75.0 (66.0–84.0) | <0.001 |
| Food preference (n, %) | <0.001 | ||
| Meat-based diet | 120 (23.5%) | 54 (11.0%) | |
| Balanced diet | 358 (70.1%) | 392 (79.7%) | |
| Vegetable-based diet | 33 (6.5%) | 46 (9.3%) | |
| Salt preference (n, %) | <0.001 | ||
| Like | 130 (25.8%) | 53 (10.8%) | |
| No strong preference/dislike | 374 (74.2%) | 439 (89.2%) | |
| Sweet taste (n, %) | 0.75 | ||
| Like | 319 (66.6%) | 327 (67.6%) | |
| No strong preference/dislike | 160 (33.4%) | 157 (32.4%) |
| Locus | Type | Obese Children (n, %) | Non-Obese Children (n, %) | OR (95%CI) 1 | p-Value 2 | p (FDR) 3 |
|---|---|---|---|---|---|---|
| 11q11 | normal | 218 (41.2) | 232 (46.6) | 1 | ||
| deletion | 123 (23.3) | 92 (18.5) | 1.43 (1.03–1.98) | 0.03 | 0.05 | |
| duplication | 188 (35.5) | 174 (34.9) | 1.15 (0.87–1.52) | 0.32 | 0.75 | |
| 5q13.2 | normal | 386 (73.0) | 369 (74.1) | 1 | ||
| deletion | 68 (12.9) | 59 (11.8) | 1.10 (0.76–1.61) | 0.61 | 0.61 | |
| duplication | 75 (14.2) | 70 (14.1) | 1.02 (0.72–1.46) | 0.90 | 0.90 | |
| 10q11.22 | normal | 459 (86.8) | 464 (93.2) | 1 | ||
| deletion | 52 (9.8) | 22 (4.4) | 2.40 (1.43–4.00) | 9.1 × 10−4 | 0.006 | |
| duplication | 18 (3.4) | 12 (2.4) | 1.51 (0.72–3.18) | 0.28 | 0.75 | |
| 10q11.22(2) | normal | 490 (92.6) | 472 (94.8) | 1 | ||
| deletion | 25 (4.7) | 15 (3.0) | 1.61 (0.84–3.10) | 0.15 | 0.21 | |
| duplication | 14 (2.6) | 11 (2.2) | 1.23 (0.55–2.73) | 0.62 | 0.87 | |
| 6q14.1 | normal | 270 (51.0) | 257 (51.6) | 1 | ||
| deletion | 202 (38.2) | 180 (36.1) | 1.07 (0.82–1.39) | 0.61 | 0.61 | |
| duplication | 56 (10.6) | 61 (12.2) | 0.87 (0.58–1.30) | 0.50 | 0.87 | |
| 4q25 | normal | 500 (94.5) | 487 (97.8) | 1 | ||
| deletion | 23 (4.3) | 6 (1.2) | 3.77 (1.52–9.36) | 0.004 | 0.01 | |
| duplication | 6 (1.1) | 5 (1.0) | 1.16 (0.35–3.83) | 0.81 | 0.90 |
| Locus | Type | BMI z-Score 1 | p-Value 3 | WHtR 2 | p-Value 4 |
|---|---|---|---|---|---|
| 11q11 | normal | 1.25 ± 1.55 | 0.47 ± 0.08 | ||
| deletion | 1.49 ± 1.69 | 0.063 | 0.49 ± 0.09 | 0.014 | |
| duplication | 1.29 ± 1.72 | 0.649 | 0.48 ± 0.08 | 0.258 | |
| 10q11.22 | normal | 1.26 ± 1.61 | 0.47 ± 0.08 | ||
| deletion | 1.87 ± 1.68 | 0.002 | 0.50 ± 0.08 | 0.006 | |
| duplication | 1.75 ± 2.12 | 0.069 | 0.51 ± 0.12 | 0.022 | |
| 4q25 | normal | 1.29 ± 1.64 | 0.48 ± 0.08 | ||
| deletion | 2.18 ± 1.45 | 0.007 | 0.52 ± 0.09 | 0.008 | |
| duplication | 1.31 ± 1.74 | 0.852 | 0.51 ± 0.08 | 0.187 |

4. Discussion

5. Conclusions
Supplementary Files
Supplementary File 1Acknowledgments
Author Contributions
Conflicts of Interest
References
- De Onis, M.; Blossner, M.; Borghi, E. Global prevalence and trends of overweight and obesity among preschool children. Am. J. Clin. Nutr. 2010, 92, 1257–1264. [Google Scholar] [CrossRef] [PubMed]
- Ji, C.Y.; Chen, T.J. Empirical changes in the prevalence of overweight and obesity among chinese students from 1985 to 2010 and corresponding preventive strategies. Biomed. Environ. Sci. 2013, 26, 1–12. [Google Scholar] [PubMed]
- Dong, B.; Wang, H.J.; Wang, Z.; Liu, J.S.; Ma, J. Trends in blood pressure and body mass index among chinese children and adolescents from 2005 to 2010. Am. J. Hypertens. 2013, 26, 997–1004. [Google Scholar] [CrossRef] [PubMed]
- Lu, X.; Shi, P.; Luo, C.Y.; Zhou, Y.F.; Yu, H.T.; Guo, C.Y.; Wu, F. Prevalence of hypertension in overweight and obese children from a large school-based population in shanghai, China. BMC Public Health 2013, 13, 24. [Google Scholar] [CrossRef] [PubMed]
- Rocchini, A.P. Childhood obesity and coronary heart disease. N. Engl. J. Med. 2011, 365, 1927–1929. [Google Scholar] [CrossRef] [PubMed]
- Chiarelli, F.; Marcovecchio, M.L. Insulin resistance and obesity in childhood. Eur. J. Endocrinol. 2008, 159, S67–S74. [Google Scholar] [CrossRef] [PubMed]
- Verbeeten, K.C.; Elks, C.E.; Daneman, D.; Ong, K.K. Association between childhood obesity and subsequent type 1 diabetes: A systematic review and meta-analysis. Diabet. Med. 2011, 28, 10–18. [Google Scholar] [CrossRef] [PubMed]
- Yang, R.; Mao, S.; Zhang, S.; Li, R.; Zhao, Z. Prevalence of obesity and overweight among chinese children with attention deficit hyperactivity disorder: A survey in zhejiang province, China. BMC Psychiatry 2013, 13, 133. [Google Scholar] [CrossRef] [PubMed]
- Cserjesi, R.; Molnar, D.; Luminet, O.; Lenard, L. Is there any relationship between obesity and mental flexibility in children? Appetite 2007, 49, 675–678. [Google Scholar] [CrossRef] [PubMed]
- Anderson, S.E.; Cohen, P.; Naumova, E.N.; Jacques, P.F.; Must, A. Adolescent obesity and risk for subsequent major depressive disorder and anxiety disorder: Prospective evidence. Psychosom. Med. 2007, 69, 740–747. [Google Scholar] [CrossRef] [PubMed]
- Elks, C.E.; den Hoed, M.; Zhao, J.H.; Sharp, S.J.; Wareham, N.J.; Loos, R.J.; Ong, K.K. Variability in the heritability of body mass index: A systematic review and meta-regression. Front. Endocrinol. 2012, 3, 29. [Google Scholar] [CrossRef]
- Jarick, I.; Vogel, C.I.; Scherag, S.; Schafer, H.; Hebebrand, J.; Hinney, A.; Scherag, A. Novel common copy number variation for early onset extreme obesity on chromosome 11q11 identified by a genome-wide analysis. Hum. Mol. Genet. 2011, 20, 840–852. [Google Scholar] [CrossRef] [PubMed]
- Glessner, J.T.; Bradfield, J.P.; Wang, K.; Takahashi, N.; Zhang, H.; Sleiman, P.M.; Mentch, F.D.; Kim, C.E.; Hou, C.; Thomas, K.A.; et al. A genome-wide study reveals copy number variants exclusive to childhood obesity cases. Am. J. Hum. Genet. 2010, 87, 661–666. [Google Scholar] [CrossRef] [PubMed]
- Bochukova, E.G.; Huang, N.; Keogh, J.; Henning, E.; Purmann, C.; Blaszczyk, K.; Saeed, S.; Hamilton-Shield, J.; Clayton-Smith, J.; O’Rahilly, S.; et al. Large, rare chromosomal deletions associated with severe early-onset obesity. Nature 2010, 463, 666–670. [Google Scholar] [CrossRef] [PubMed]
- Wheeler, E.; Huang, N.; Bochukova, E.G.; Keogh, J.M.; Lindsay, S.; Garg, S.; Henning, E.; Blackburn, H.; Loos, R.J.; Wareham, N.J.; et al. Genome-wide snp and cnv analysis identifies common and low-frequency variants associated with severe early-onset obesity. Nat. Genet. 2013, 45, 513–517. [Google Scholar] [CrossRef] [PubMed]
- Bachmann-Gagescu, R.; Mefford, H.C.; Cowan, C.; Glew, G.M.; Hing, A.V.; Wallace, S.; Bader, P.I.; Hamati, A.; Reitnauer, P.J.; Smith, R.; et al. Recurrent 200-kb deletions of 16p11.2 that include the sh2b1 gene are associated with developmental delay and obesity. Genet. Med. 2010, 12, 641–647. [Google Scholar] [CrossRef] [PubMed]
- Willer, C.J.; Speliotes, E.K.; Loos, R.J.; Li, S.; Lindgren, C.M.; Heid, I.M.; Berndt, S.I.; Elliott, A.L.; Jackson, A.U.; Lamina, C.; et al. Six new loci associated with body mass index highlight a neuronal influence on body weight regulation. Nat. Genet. 2009, 41, 25–34. [Google Scholar] [CrossRef] [PubMed]
- Sha, B.Y.; Yang, T.L.; Zhao, L.J.; Chen, X.D.; Guo, Y.; Chen, Y.; Pan, F.; Zhang, Z.X.; Dong, S.S.; Xu, X.H.; et al. Genome-wide association study suggested copy number variation may be associated with body mass index in the chinese population. J. Hum. Genet. 2009, 54, 199–202. [Google Scholar] [CrossRef] [PubMed]
- Kimm, S.Y. The role of dietary fiber in the development and treatment of childhood obesity. Pediatrics 1995, 96, 1010–1014. [Google Scholar] [PubMed]
- Gallos, L.K.; Barttfeld, P.; Havlin, S.; Sigman, M.; Makse, H.A. Collective behavior in the spatial spreading of obesity. Sci. Rep. 2012, 2, 454. [Google Scholar] [CrossRef] [PubMed]
- Hiroi, M.; Nagahara, Y.; Miyauchi, R.; Misaki, Y.; Goda, T.; Kasezawa, N.; Sasaki, S.; Yamakawa-Kobayashi, K. The combination of genetic variations in the prdx3 gene and dietary fat intake contribute to obesity risk. Obesity 2011, 19, 882–887. [Google Scholar] [CrossRef] [PubMed]
- Marti, A.; Martinez-Gonzalez, M.A.; Martinez, J.A. Interaction between genes and lifestyle factors on obesity. Proc. Nutr. Soc. 2008, 67, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Basic, M.; Butorac, A.; Landeka Jurcevic, I.; Bacun-Druzina, V. Obesity: Genome and environment interactions. Arh. Hig. Rada Toksikol. 2012, 63, 395–405. [Google Scholar] [CrossRef] [PubMed]
- Reinehr, T.; Hebebrand, J.; Friedel, S.; Toschke, A.M.; Brumm, H.; Biebermann, H.; Hinney, A. Lifestyle intervention in obese children with variations in the melanocortin 4 receptor gene. Obesity 2009, 17, 382–389. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.; Xu, Y.; Liang, L.; Fu, J.; Xiong, F.; Liu, G.; Gong, C.; Luo, F.; Chen, S.; Xu, C.; et al. The effects of genetic variation in fto rs9939609 on obesity and dietary preferences in Chinese han children and adolescents. PLoS ONE 2014, 9, e104574. [Google Scholar] [CrossRef] [PubMed]
- Group of China Obesity Task Force. Body mass index reference norm for screening overweight and obesity in chinese children and adolescents. Zhonghua Liu Xing Bing Xue Za Zhi 2004, 25, 97–102. [Google Scholar]
- Yu, Y.; Zhu, H.; Miller, D.T.; Gusella, J.F.; Platt, O.S.; Wu, B.L.; Shen, Y. Age- and gender-dependent obesity in individuals with 16p11.2 deletion. J. Genet. Genomics 2011, 38, 403–409. [Google Scholar] [CrossRef] [PubMed]
- Du, R.; Lu, C.; Jiang, Z.; Li, S.; Ma, R.; An, H.; Xu, M.; An, Y.; Xia, Y.; Jin, L.; et al. Efficient typing of copy number variations in a segmental duplication-mediated rearrangement hotspot using multiplex competitive amplification. J. Hum. Genet. 2012, 57, 545–551. [Google Scholar] [CrossRef] [PubMed]
- De Onis, M.; Onyango, A.W.; Borghi, E.; Siyam, A.; Nishida, C.; Siekmann, J. Development of a who growth reference for school-aged children and adolescents. Bull. World Health Organ. 2007, 85, 660–667. [Google Scholar] [CrossRef] [PubMed]
- Cole, T.J.; Bellizzi, M.C.; Flegal, K.M.; Dietz, W.H. Establishing a standard definition for child overweight and obesity worldwide: International survey. BMJ 2000, 320, 1240–1243. [Google Scholar] [CrossRef] [PubMed]
- Andersson, T.; Alfredsson, L.; Kallberg, H.; Zdravkovic, S.; Ahlbom, A. Calculating measures of biological interaction. Eur. J. Epidemiol. 2005, 20, 575–579. [Google Scholar] [CrossRef] [PubMed]
- Sun, C.; Cao, M.; Shi, J.; Li, L.; Miao, L.; Hong, J.; Cui, B.; Ning, G. Copy number variations of obesity relevant loci associated with body mass index in young chinese. Gene 2013, 516, 198–203. [Google Scholar] [CrossRef] [PubMed]
- Asakawa, A.; Inui, A.; Yuzuriha, H.; Ueno, N.; Katsuura, G.; Fujimiya, M.; Fujino, M.A.; Niijima, A.; Meguid, M.M.; Kasuga, M.; et al. Characterization of the effects of pancreatic polypeptide in the regulation of energy balance. Gastroenterology 2003, 124, 1325–1336. [Google Scholar] [CrossRef] [PubMed]
- Batterham, R.L.; le Roux, C.W.; Cohen, M.A.; Park, A.J.; Ellis, S.M.; Patterson, M.; Frost, G.S.; Ghatei, M.A.; Bloom, S.R. Pancreatic polypeptide reduces appetite and food intake in humans. J. Clin. Endocrinol. Metab. 2003, 88, 3989–3992. [Google Scholar] [CrossRef] [PubMed]
- Ueno, N.; Inui, A.; Iwamoto, M.; Kaga, T.; Asakawa, A.; Okita, M.; Fujimiya, M.; Nakajima, Y.; Ohmoto, Y.; Ohnaka, M.; et al. Decreased food intake and body weight in pancreatic polypeptide-overexpressing mice. Gastroenterology 1999, 117, 1427–1432. [Google Scholar] [CrossRef] [PubMed]
- Balasubramaniam, A.; Mullins, D.E.; Lin, S.; Zhai, W.; Tao, Z.; Dhawan, V.C.; Guzzi, M.; Knittel, J.J.; Slack, K.; Herzog, H.; et al. Neuropeptide y (npy) y4 receptor selective agonists based on npy(32–36): Development of an anorectic y4 receptor selective agonist with picomolar affinity. J. Med. Chem. 2006, 49, 2661–2665. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.; Shi, Y.C.; Yulyaningsih, E.; Aljanova, A.; Zhang, L.; Macia, L.; Nguyen, A.D.; Lin, E.J.; During, M.J.; Herzog, H.; et al. Critical role of arcuate y4 receptors and the melanocortin system in pancreatic polypeptide-induced reduction in food intake in mice. PLoS ONE 2009, 4, e8488. [Google Scholar] [CrossRef] [PubMed]
- Sainsbury, A.; Shi, Y.C.; Zhang, L.; Aljanova, A.; Lin, Z.; Nguyen, A.D.; Herzog, H.; Lin, S. Y4 receptors and pancreatic polypeptide regulate food intake via hypothalamic orexin and brain-derived neurotropic factor dependent pathways. Neuropeptides 2010, 44, 261–268. [Google Scholar] [CrossRef] [PubMed]
- Karhunen, L.J.; Juvonen, K.R.; Huotari, A.; Purhonen, A.K.; Herzig, K.H. Effect of protein, fat, carbohydrate and fibre on gastrointestinal peptide release in humans. Regul. Pept. 2008, 149, 70–78. [Google Scholar] [CrossRef] [PubMed]
- Klopocki, E.; Mundlos, S. Copy-number variations, noncoding sequences, and human phenotypes. Annu. Rev. Genomics Hum. Genet. 2011, 12, 53–72. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, D.; Li, Z.; Wang, H.; Yang, M.; Liang, L.; Fu, J.; Wang, C.; Ling, J.; Zhang, Y.; Zhang, S.; et al. Interactions between Obesity-Related Copy Number Variants and Dietary Behaviors in Childhood Obesity. Nutrients 2015, 7, 3054-3066. https://doi.org/10.3390/nu7043054
Zhang D, Li Z, Wang H, Yang M, Liang L, Fu J, Wang C, Ling J, Zhang Y, Zhang S, et al. Interactions between Obesity-Related Copy Number Variants and Dietary Behaviors in Childhood Obesity. Nutrients. 2015; 7(4):3054-3066. https://doi.org/10.3390/nu7043054
Chicago/Turabian StyleZhang, Dandan, Zhenli Li, Hao Wang, Min Yang, Li Liang, Junfen Fu, Chunling Wang, Jie Ling, Yan Zhang, Shuai Zhang, and et al. 2015. "Interactions between Obesity-Related Copy Number Variants and Dietary Behaviors in Childhood Obesity" Nutrients 7, no. 4: 3054-3066. https://doi.org/10.3390/nu7043054